Аминоацилаза
| аминоацилаза | |||
|---|---|---|---|
| Идентификаторы | |||
| Номер ЕС. | 3.5.1.14 | ||
| Номер CAS. | 9012-37-7 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
В энзимологии аминоацилаза , ( КФ 3.5.1.14 ) — фермент катализирующий реакцию химическую .
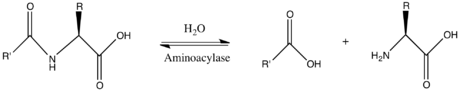
Таким образом, двумя субстратами этого фермента являются N-ацил-L-аминокислота и H 2 O , тогда как двумя его продуктами являются карбоксилат и L-аминокислота .
Этот фермент принадлежит к семейству гидролаз , действующих на связи углерод-азот , отличные от пептидных связей , особенно в линейных амидах . Систематическое название этого ферментов класса N-ацил-L-аминокислот — амидогидролаза . Другие широко используемые названия включают дегидропептидазу II , гистозим , гиппуриказу , бензамидазу , ацилазу I , гиппуразу , деацилазу аминокислот , L-аминоацилазу , ацилазу , аминоацилазу I , ацилазу L-аминокислот , альфа-N-ациламинокислоты гидролазу , длинную ациламидоацилазу. и короткая ациламидоацилаза . Этот фермент участвует в цикле мочевины и метаболизме аминогрупп .
Структура фермента
[ редактировать ]
По состоянию на конец 2007 года две структуры были решены для этого класса ферментов с PDB кодами доступа 1Q7L и 1YSJ . Эти структуры также соответствуют двум известным аминокислотным последовательностям аминоацилаз первичным . В связанных статьях идентифицируются два типа доменов , включающих аминоацилазы: цинксвязывающие домены , которые связывают Zn. 2+ ионы- и домены , которые облегчают димеризацию цинк - связывающих доменов . [ 1 ] [ 2 ] Именно эта димеризация позволяет происходить катализу аминоацилазы , поскольку активный центр находится между двумя цинксвязывающими ее доменами . [ 1 ]
Связанный цинк облегчает связывание субстрата L-аминокислоты N-ацил - , вызывая конформационный сдвиг , который объединяет белка субъединицы вокруг субстрата и позволяет катализ . осуществить [ 3 ] Аминоацилаза 1 существует в гетеротетрамерной структуре, то есть 2 цинка домена связывания и 2 домена димеризации объединяются, образуя аминоацилазы 1 четвертичную структуру .
Ферментативный механизм
[ редактировать ]
Аминоацилаза — это металлофермент , которому необходим цинк ( Zn 2+ ) как кофактор для функционирования. [ 3 ] [ 4 ] Ионы цинка , внутри аминоацилазы координированы гистидином , глутаматом с аспартатом и водой . [ 1 ] [ 3 ] [ 5 ] Ион цинка остатком поляризует воду . способствуя депротонированию близлежащим основным , ее [ 3 ] [ 5 ] Отрицательно заряженный ион является нуклеофильным и атакует электрофильный карбонильный углерод субстрата гидроксид - ацильной группы . [ 5 ] Точный механизм после этого момента неизвестен, возможно, карбонил затем преобразуется, разрывает амидную связь и образует два продукта . В какой-то момент механизма входит другая молекула воды и координирует свои действия с цинком , возвращая фермент в исходное состояние. [ 5 ]

Нуклеофильная атака водой является лимитирующей стадией каталитического механизма аминоацилазы . [ 6 ] Эта нуклеофильная атака обратима, тогда как последующие этапы являются быстрыми и необратимыми. [ 6 ] последовательность реакций является примером кинетики Михаэлиса-Ментен , позволяющей определить KM Ментен , Kcat с , Vmax Эта , число оборотов и специфичность субстрата помощью классических Михаэлиса- экспериментов с ферментами . [ 6 ] реакции Второй и третий этапы вперед вызывают образование и выделение продуктов . [ 6 ]
Биологическая функция
[ редактировать ]
Аминоацилазы экспрессируются в почках , где они перерабатывают N-ацил-L-аминокислоты в L-аминокислоты и способствуют регуляции цикла мочевины.
N-ацил-L-аминокислоты образуются, когда L-аминокислот ковалентно N-конец связан с ацильной группой . Ацильная группа обеспечивает стабильность аминокислоты , делая ее более устойчивой к деградации. Кроме того, N-ацил-L-аминокислоты не могут использоваться непосредственно в качестве строительных блоков для белков и сначала должны быть преобразованы в L-аминокислоты с помощью аминоацилазы. Опять же, продукты L-аминокислот можно использовать для биосинтеза или катаболизма энергии .
Аминоацилаза участвует в регуляции цикла мочевины . N-ацетил-L-глутамат является аллостерическим активатором карбамоилфосфатсинтетазы . , важнейшего фермента , который коммитирует NH4 + молекул в цикл мочевины . [ 7 ] Цикл мочевины избавляется от избытка аммиака ( NH 4 + ) в организме, этот процесс необходимо активировать во время повышенного катаболизма белков , поскольку при распаде аминокислот образуется большое количество NH 4 . + . [ 7 ] Когда катаболизм аминокислот усиливается, N-ацетилглутаматсинтаза активируется, производя больше N-ацетил-L-глутамата, который активирует карбамоилфосфатсинтетазу и позволяет ей избавиться от избытка NH 4 . + от катаболизма . [ 7 ]
Аминоацилаза активируется во время дефицита питательных веществ или голодания , вызывая распад N-ацетил-L-глутамата , что снижает регуляцию карбамоилфосфатсинтетазы и остальной части цикла мочевины . не так много . Этот ответ эволюционно выгоден, поскольку дефицит питательных веществ означает, что NH 4 + от этого необходимо избавиться, и поскольку организм хочет сохранить как можно больше аминокислот. [ 7 ]
Актуальность заболевания
[ редактировать ]Дефицит аминоацилазы 1 ( A1D ) — редкое заболевание вызванное аутосомно-рецессивной мутацией гена аминоацилазы 1 , ( ACY1 ) на хромосоме 3p21 . [ 8 ] [ 9 ] [ 10 ] [ 11 ] [ 12 ] Отсутствие функциональной аминоацилазы 1, вызванное A1D , приводит к нарушению цикла мочевины , вызывая ряд неврологических расстройств, включая судороги , мышечную гипотонию , умственную отсталость и нарушение психомоторного развития . [ 8 ] [ 13 ] [ 14 ] [ 15 ] A1D также связан с аутизмом . [ 16 ] У пациентов с A1D симптомы часто начинают проявляться вскоре после рождения , но, похоже, полностью выздоравливают в течение следующих нескольких лет. [ 13 ] [ 14 ] [ 15 ]
Дефицит аминоацилазы 2 , также известный как болезнь Канавана , является еще одним редким заболеванием вызванным мутацией гена ASPA , (на хромосоме 17 ), которая приводит к дефициту фермента аминоацилазы 2 . Аминоацилаза 2 известна тем, что она может гидролизовать N-ацетиласпартат, а аминоацилаза 1 - нет. [ 17 ]
Промышленная значимость
[ редактировать ]Аминоацилазы используются для производства L- аминокислот в промышленных условиях с конца 1950-х годов. [ 18 ] Поскольку аминоацилазы являются субстрат -специфичными для N-ацил-L-аминокислот , а не для N-ацил-D-аминокислот , аминоацилазы можно использовать для надежного получения смеси этих двух реагентов и превращения только L -энантиомеров в продукты , которые затем можно использовать. быть выделены путем растворимости из непрореагировавших N-ацил-D-аминокислот . [ 18 ] [ 19 ] Хотя этот процесс в течение многих лет осуществлялся в реакторе периодического действия более быстрый и менее затратный процесс , в конце 1970-х годов был разработан , в котором аминоацилазы помещались в колонку N -ацил - аминокислоты . , через которую затем непрерывно промывали [ 18 ] [ 20 ] Этот процесс до сих пор используется в промышленности для преобразования N-ациламинокислот в аминокислоты специфичным способом энантиомерно- .
Эволюция
[ редактировать ]Во многих научных исследованиях за последние полвека аминоацилаза свиньи использовалась в качестве модельного фермента аминоацилазы . [ 21 ] аминокислотная аминоацилазы свиньи. последовательность и первичная структура Определены [ 4 ] Свиная аминоацилаза 1 состоит из двух идентичных гетеродимерных субъединиц, каждая из которых состоит из 406 аминокислот, с ацетилаланином на N-конце каждой. [ 4 ] Аминоацилаза свиньи отличается от человека аминоацилазы по структуре , но повторяет ее функцию. [ 1 ] [ 4 ] [ 22 ] На основе этих данных можно сделать вывод, что эти два фермента произошли от общего предкового белка , сохранив функцию, но разошлись по структуре . со временем [ 1 ] [ 4 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и Линднер Х.А., Лунин В.В., Алари А., Хекер Р., Сиглер М., Менар Р. (ноябрь 2003 г.). «Основная роль лигирования цинка и димеризации ферментов в катализе в семействе аминоацилазы-1/M20» . Журнал биологической химии . 278 (45): 44496–504. дои : 10.1074/jbc.M304233200 . ПМИД 12933810 .
- ^ Фонс В.С., Ли М. (апрель 1953 г.). «Гидролиз N-ацилпроизводных аланина и фенилаланина ацилазой I и карбоксипептидазой» . Журнал биологической химии . 201 (2): 847–56. ПМИД 13061423 .
- ^ Jump up to: а б с д Линднер Х.А., Алари А., Уилке М., Сулеа Т. (апрель 2008 г.). «Исследование ацилсвязывающего кармана аминоацилазы-1». Биохимия . 47 (14): 4266–75. дои : 10.1021/bi702156h . ПМИД 18341290 .
- ^ Jump up to: а б с д и Митта М., Оноги Х., Ямамото А., Като И., Сакияма Ф., Цунасава С. (декабрь 1992 г.). «Первичная структура свиной аминоацилазы 1, выведенная из последовательности кДНК». Журнал биохимии . 112 (6): 737–42. doi : 10.1093/oxfordjournals.jbchem.a123968 . ПМИД 1284246 .
- ^ Jump up to: а б с д Херник М., Фиерке, Калифорния (январь 2005 г.). «Цинковые гидролазы: механизмы цинкзависимых деацетилаз». Архив биохимии и биофизики . 433 (1): 71–84. дои : 10.1016/j.abb.2004.08.006 . ПМИД 15581567 .
- ^ Jump up to: а б с д Отвос Л., Моравчик Э., Мади Г. (сентябрь 1971 г.). «Исследование механизма катализируемого ацилазой-I гидролиза ациламинокислот». Связь с биохимическими и биофизическими исследованиями . 44 (5): 1056–64. дои : 10.1016/S0006-291X(71)80192-4 . ПМИД 5160398 .
- ^ Jump up to: а б с д Берг, Джереми М.; Тимочко, Джон Л.; Страйер, Люберт (2012). Биохимия . Нью-Йорк: WH Freeman and Company. п. 688. ИСБН 978-1-4292-2936-4 .
- ^ Jump up to: а б Саммер А., Кристенсен Э., Швенгер С. и др. (июнь 2011 г.). «Молекулярная основа дефицита аминоацилазы 1» (PDF) . Биохимия и биофизика Acta (BBA) - Молекулярные основы болезней . 1812 (6): 685–90. дои : 10.1016/j.badis.2011.03.005 . ПМИД 21414403 .
- ^ Ферри Л., Фунгини С., Фиораванти А. и др. (октябрь 2013 г.). «Дефицит аминоацилазы I из-за пропуска экзона мРНК ACY1». Клиническая генетика . 86 (4): 367–372. дои : 10.1111/cge.12297 . ПМИД 24117009 . S2CID 24017306 .
- ^ Миллер Ю.Е., Минна Дж.Д., Газдар А.Ф. (июнь 1989 г.). «Отсутствие экспрессии аминоацилазы-1 при мелкоклеточном раке легких. Доказательства инактивации генов, кодируемых хромосомой 3p» . Журнал клинических исследований . 83 (6): 2120–4. дои : 10.1172/JCI114125 . ПМК 303939 . ПМИД 2542383 .
- ^ EnterGene 95
- ^ Миллер Ю.Е., Драбкин Х., Джонс С., Фишер Дж.Х. (сентябрь 1990 г.). «Человеческая аминоацилаза-1: клонирование, региональное отнесение к дистальной хромосоме 3p21.1 и идентификация перекрестно-гибридизирующей последовательности на хромосоме 18». Геномика . 8 (1): 149–54. дои : 10.1016/0888-7543(90)90237-О . ПМИД 1707030 .
- ^ Jump up to: а б Сасс Дж.О., Мор В., Ольбрих Х. и др. (март 2006 г.). «Мутации в ACY1, гене, кодирующем аминоацилазу 1, вызывают новое врожденное нарушение метаболизма» . Американский журнал генетики человека . 78 (3): 401–9. дои : 10.1086/500563 . ПМК 1380284 . ПМИД 16465618 .
- ^ Jump up to: а б Сасс Д.О., Ольбрих Х., Мор В. и др. (июнь 2007 г.). «Неврологические данные при дефиците аминоацилазы 1». Неврология . 68 (24): 2151–3. дои : 10.1212/01.wnl.0000264933.56204.e8 . ПМИД 17562838 . S2CID 43376960 .
- ^ Jump up to: а б Ван Костер Р.Н., Герло Э.А., Джардина Т.Г. и др. (декабрь 2005 г.). «Дефицит аминоацилазы I: новая врожденная ошибка метаболизма». Связь с биохимическими и биофизическими исследованиями . 338 (3): 1322–6. дои : 10.1016/j.bbrc.2005.10.126 . ПМИД 16274666 .
- ^ Тылки-Шиманска А., Градовска В., Зоммер А. и др. (декабрь 2010 г.). «Дефицит аминоацилазы 1, связанный с аутистическим поведением». Журнал наследственных метаболических заболеваний . 33 Приложение 3: S211–4. дои : 10.1007/s10545-010-9089-3 . ПМИД 20480396 . S2CID 13374954 .
- ^ Се Ц, Го Т, Ван Т, Лу Дж, Чжоу Х. М. (ноябрь 2003 г.). «Аспартат-индуцированное аминоацилазное сворачивание и образование расплавленной глобулы». Международный журнал биохимии и клеточной биологии . 35 (11): 1558–72. дои : 10.1016/S1357-2725(03)00131-6 . ПМИД 12824065 .
- ^ Jump up to: а б с Сато, Тадаши; Тоса, Тецуя (2010). «Производство L-аминокислот с помощью аминоацилазы». Энциклопедия промышленной биотехнологии . стр. 1–20. дои : 10.1002/9780470054581.eib497 . ISBN 978-0-470-05458-1 .
- ^ Бирнбаум С.М., Левинтов Л., Кингсли Р.Б., Гринштейн Дж.П. (январь 1952 г.). «Специфичность ацилаз аминокислот» . Журнал биологической химии . 194 (1): 455–70. ПМИД 14927637 .
- ^ Хуан М.К., Чжоу Х.М. (1994). «Щелочное разворачивание и индуцированное солью сворачивание аминоацилазы при высоком pH». Ферменты и белки . 48 (4): 229–37. дои : 10.1159/000474993 . ПМИД 8821711 .
- ^ Корейши М., Асаяма Ф., Иманака Х. и др. (октябрь 2005 г.). «Очистка и характеристика новой аминоацилазы из Streptomyces mobaraensis» . Бионауки, биотехнологии и биохимия . 69 (10): 1914–22. дои : 10.1271/bbb.69.1914 . ПМИД 16244442 .
- ^ Митта М., Като И., Цунасава С. (август 1993 г.). «Нуклеотидная последовательность аминоацилазы-1 человека». Biochimica et Biophysical Acta (BBA) – Структура и экспрессия генов . 1174 (2): 201–3. дои : 10.1016/0167-4781(93)90116-У . ПМИД 8357837 .