Деметилаза
Деметилазы — это ферменты , удаляющие метильные (СН 3 ) группы из нуклеиновых кислот , белков (особенно гистонов ) и других молекул. Деметилазы являются важными эпигенетическими белками, поскольку они ответственны за транскрипционную регуляцию генома , контролируя метилирование ДНК и гистонов и, как следствие, состояние хроматина в определенных генов локусах .
Деметилирование лизина гистонов
[ редактировать ]
Метилирование гистонов первоначально считалось фактически необратимым процессом, поскольку период полураспада метилирования гистонов был примерно равен периоду полураспада гистонов. [1] Гистон лизиндеметилаза LSD1 (позже классифицированная как KDM1A) была впервые идентифицирована в 2004 году как гомолог ядерной аминоксидазы. [2] Существуют два основных класса гистон-лизиндеметилаз, определяемые их механизмами: флавинадениндинуклеотид (FAD) -зависимые аминоксидазы и α-кетоглутарат-зависимые гидроксилазы .
Гистон-лизиндеметилазы обладают множеством доменов , которые отвечают за распознавание гистонов, связывание ДНК, связывание субстрата метилированных аминокислот и каталитическую активность. К ним относятся:
- FAD-зависимые домены аминоксидазы, содержащие активный каталитический сайт KDM1
- Домены Jumonji-C, содержащие активный каталитический сайт от KDM2 до KDM8. [3] [4]
- Домены Jumonji-N, ответственные за стабильность конформации домена Jumonji-C.
- Домены SWIRM (SWI3P, RSC8P и Moira) предложены в качестве якорного сайта для субстратов гистонов и ответственны за хроматина . стабильность
- PHD, CXXC и C5HC2, Домены цинковых пальцев ответственные за распознавание и связывание гистонов.
Гистон-лизиндеметилазы классифицируются в соответствии с их доменами и уникальными субстратными специфичностями. Субстраты лизина идентифицируются по их положению в соответствующей аминокислотной последовательности гистонов и состоянию метилирования (например, H3K9me3 относится к триметилированному гистону 3 лизину 9).


- КДМ1
- Гомологи KDM1 включают KDM1A и KDM1B . KDM1A деметилирует H3K4me1/2 и H3K9me1/2, а KDM1B эметилирует H3K4me1/2. Активность KDM1 имеет решающее значение для эмбриогенеза и тканеспецифической дифференцировки , а также роста ооцитов. [1] Делеция гена KDM1A может влиять на рост и дифференцировку эмбриональных стволовых клеток и всегда приводит к летальному исходу у нокаутных мышей . [5] [6] Экспрессия гена KDM1A повышается при некоторых видах рака. [7] [8] и поэтому ингибирование KDM1A считается возможным эпигенетическим лечением рака. [9] [10] [11] :KDM1B, однако, в основном участвует в развитии ооцитов . Делеция этого гена приводит к материнской смертности у мышей. [12] Ортологи KDM1 у D. melanogaster и C. elegans , по-видимому, функционируют аналогично KDM1B, а не KDM1A. [13] [14]
- КДМ2
- Гомологи KDM2 включают KDM2A и KDM2B . KDM2A и KDM2B деметилируют H3K4me3 и H3K36me2/3. KDM2A играет роль либо в стимулировании, либо в ингибировании функции опухоли, а KDM2B играет роль в онкогенезе . [1] KDM2A и KDM2B обладают доменами цинковых пальцев CXXC, ответственными за связывание с неметилированными CpG-островками, и считается, что они могут связываться со многими регуляторными элементами гена в отсутствие специфичных для последовательности факторов транскрипции. [15] :Сверхэкспрессия KDM2B наблюдалась при лимфоме и аденокарциноме человека , а пониженная экспрессия KDM2B наблюдалась при раке предстательной железы и глиобластоме человека. Кроме того, было показано, что KDM2B предотвращает старение в некоторых клетках посредством эктопической экспрессии . [16]
- КДМ3
- Гомологи KDM3 включают KDM3A , KDM3B и KDM3C . KDM3A, KDM3B и KDM3C деметилируют H3K9me1/2. KDM3A играет роль в сперматогенезе и метаболических функциях, однако активность KDM3B и KDM3C конкретно не известна. [1] : Исследования нокдауна KDM3A на мышах привели к мужскому бесплодию и ожирению у взрослых. Дополнительные исследования показали, что KDM3A может играть роль в регуляции генов, зависимых от андрогенных рецепторов, а также генов, участвующих в плюрипотентности , что указывает на потенциальную роль KDM3A в онкогенезе. [17]
- КДМ4
- Гомологи KDM4 включают KDM4A , KDM4B , KDM4C , KDM4D , KDM4E и KDM4F. KDM4A, KDM4B и KDM4C деметилируют H3K9me2/3, H3K9me3 и H3K36me2/3, а KDM4D, KDM4E и KDM4F деметилируют H3K9me2/3. KDM4A, KDM4B, KDM4C и KDM4D играют роль в онкогенезе , однако активность KDM4E и KDM4F конкретно не известна. [1] Повышение регуляции KDM4B наблюдалось при медуллобластоме, а амплификация KDM4C была зарегистрирована при плоскоклеточном раке пищевода, медуллобластоме и раке молочной железы. [18] [19] [20] [21] Другие данные по экспрессии генов также свидетельствуют о том, что KDM4A, KDM4B и KDM4C сверхэкспрессируются при раке простаты. [22]
- КДМ5
- Гомологи KDM5 включают KDM5A , KDM5B , KDM5C и KDM5D . KDM5A, KDM5B, KDM5C и KDM5D деметилируют H3K4me2/3. [1] Семейство KDM5, по-видимому, регулирует ключевые функции развития, включая клеточную дифференцировку, функцию митохондрий и развитие клеточного цикла . [23] [24] [25] [26] [27] [28] KDM5B и KDM5C также продемонстрировали взаимодействие с белками PcG, которые участвуют в репрессии транскрипции. Мутации KDM5C в Х-хромосоме также наблюдались у пациентов с Х-сцепленной умственной отсталостью. [29] Истощение гомологов KDM5C у D. rerio выявило дефекты структуры мозга и гибель нейрональных клеток. [30]
- КДМ6
- Семейство KDM6 включает KDM6A , KDM6B и KDM6C . KDM6A и KDM6B деметилируют H3K27me2/3, а KDM4C деметилирует H3K27me3. KDM6A и KDM6B обладают опухолесупрессирующими свойствами. Нокдаун KDM6A в фибробластах приводит к немедленному увеличению популяции фибробластов. KDM6B, экспрессируемый в фибробластах, индуцирует онкогены пути Ras/Raf/MEK/ERK. [31] Точечные мутации KDM6A были идентифицированы как одна из причин синдрома Кабуки — врожденного заболевания, приводящего к умственной отсталости. [32] [33] Делеция KDM6A у D. rerio приводит к снижению экспрессии генов HOX, которые играют роль в регуляции формирования паттерна тела во время развития. [34] Исследования на млекопитающих показали, что KDM6A также регулирует гены HOX. [35] [36] Мутация KDM5B нарушает развитие гонад у C.elegans . [35] Другие исследования показали, что экспрессия KDM6B усиливается в активированных макрофагах и динамически экспрессируется во время дифференцировки стволовых клеток . [37] [38]
Деметилирование эфира
[ редактировать ]
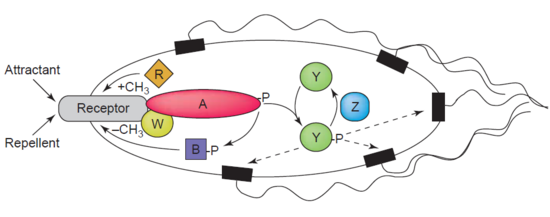
Другим примером деметилазы является протеин-глутамат-метилэстераза также известная как белок CheB (EC 3.1.1.61), которая деметилирует MCP ( м -этил-принимающие белки хемотаксиса , ) посредством гидролиза сложноэфирных связей карбоновых кислот. Ассоциация рецептора хемотаксиса с агонистом приводит к фосфорилированию CheB. Фосфорилирование белка CheB усиливает его каталитическую деметилирующую активность MCP, что приводит к адаптации клетки к стимулам окружающей среды. [39] MCP реагируют на внеклеточные аттрактанты и репелленты у бактерий, таких как E. coli, в регуляции хемотаксиса . CheB более конкретно называют метилэстеразой , поскольку она удаляет метильные группы из остатков метилглутамата , расположенных на MCP, посредством гидролиза, образуя глутамат, сопровождающийся высвобождением метанола . [40]
CheB представляет особый интерес для исследователей, поскольку может быть терапевтической мишенью для смягчения распространения бактериальных инфекций. [41]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж Педерсен М.Т., Хелин К. (ноябрь 2010 г.). «Гистоновые деметилазы в развитии и заболеваниях». Тенденции в клеточной биологии . 20 (11): 662–71. дои : 10.1016/j.tcb.2010.08.011 . ПМИД 20863703 .
- ^ Ши Ю, Лан Ф, Мэтсон С, Маллиган П, Уэтстайн-младший, Коул П.А., Касеро Р.А., Ши Ю (декабрь 2004 г.). «Деметилирование гистонов, опосредованное гомологом ядерной аминоксидазы LSD1» . Клетка . 119 (7): 941–53. дои : 10.1016/j.cell.2004.12.012 . ПМИД 15620353 .
- ^ Мосаммапараст Н., Ши Ю (2010). «Обращение метилирования гистонов: биохимические и молекулярные механизмы деметилаз гистонов». Ежегодный обзор биохимии . 79 : 155–79. doi : 10.1146/annurev.biochem.78.070907.103946 . ПМИД 20373914 .
- ^ Апреликова О, Чен К., Эль Туни Л.Х., Бриньяц-Гиттард С., Хань Дж., Цю Т., Ян Х.Х., Ли М.П., Чжу М., Грин Дж.Э. (апрель 2016 г.). «Эпигенетический модификатор JMJD6 амплифицируется в опухолях молочной железы и взаимодействует с c-Myc, усиливая клеточную трансформацию, прогрессирование опухоли и метастазирование» . Клин Эпигенетика . 8 (38): 38. дои : 10.1186/s13148-016-0205-6 . ПМЦ 4831179 . ПМИД 27081402 .
- ^ Ван Дж., Хеви С., Кураш Дж.К., Лей Х., Гей Ф., Байко Дж., Су Х., Сунь В., Чанг Х., Сюй Г., Годе Ф., Ли Э., Чен Т. (январь 2009 г.). «Лизиндеметилаза LSD1 (KDM1) необходима для поддержания глобального метилирования ДНК». Природная генетика . 41 (1): 125–9. дои : 10.1038/ng.268 . ПМИД 19098913 . S2CID 2010695 .
- ^ Ван Дж., Скалли К., Чжу Икс, Кай Л., Чжан Дж., Префонтейн Г.Г., Кронес А., Оги К.А., Чжу П., Гарсиа-Бассетс И., Лю Ф., Тейлор Х., Лозак Дж., Джейс Ф.Л., Корах К.С., Гласс С.К., Фу XD, Розенфельд М.Г. (апрель 2007 г.). «Противоположные комплексы LSD1 участвуют в программах активации и репрессии генов развития». Природа . 446 (7138): 882–7. Бибкод : 2007Natur.446..882W . дои : 10.1038/nature05671 . ПМИД 17392792 . S2CID 4387240 .
- ^ Каль П., Гуллотти Л., Хойкамп Л.С., Вольф С., Фридрихс Н., Форройтер Р., Солледер Г., Бастиан П.Дж., Эллингер Дж., Мецгер Э., Шюле Р., Бюттнер Р. (декабрь 2006 г.). «Коактиваторы андрогенных рецепторов лизин-специфическая гистондеметилаза 1 и белок 2 домена четырех с половиной LIM предсказывают риск рецидива рака простаты». Исследования рака . 66 (23): 11341–7. дои : 10.1158/0008-5472.CAN-06-1570 . PMID 17145880 .
- ^ Лим С., Янцер А., Беккер А., Циммер А., Шюле Р., Бюттнер Р., Кирфель Дж. (март 2010 г.). «Лизин-специфическая деметилаза 1 (LSD1) высоко экспрессируется при ER-негативном раке молочной железы и является биомаркером, предсказывающим агрессивную биологию». Канцерогенез . 31 (3): 512–20. дои : 10.1093/carcin/bgp324 . ПМИД 20042638 .
- ^ Мецгер Э., Виссманн М., Инь Н., Мюллер Дж.М., Шнайдер Р., Петерс А.Х., Гюнтер Т., Бюттнер Р., Шюле Р. (сентябрь 2005 г.). «LSD1 деметилирует репрессивные метки гистонов, чтобы способствовать транскрипции, зависимой от андрогенных рецепторов». Природа . 437 (7057): 436–9. Бибкод : 2005Nature.437..436M . дои : 10.1038/nature04020 . ПМИД 16079795 . S2CID 4308627 .
- ^ Шульте Й.Х., Лим С., Шрамм А., Фридрихс Н., Костер Дж., Верстеег Р., Ора И., Пайтлер К., Кляйн-Хитпасс Л., Кухфиттиг-Кулле С., Мецгер Е., Шуле Р., Эггерт А., Бюттнер Р., Кирфель Дж. (Мар. 2009). «Лизин-специфическая деметилаза 1 сильно экспрессируется при низкодифференцированной нейробластоме: значение для терапии» . Исследования рака . 69 (5): 2065–71. дои : 10.1158/0008-5472.CAN-08-1735 . ПМИД 19223552 .
- ^ Ван Ю, Чжан Х, Чен Ю, Сунь Ю, Ян Ф, Юй В, Лян Дж, Сунь Л, Ян Х, Ши Л, Ли Р, Ли Ю, Чжан Ю, Ли Ц, И Х, Шан Ю (август 2009 г.) ). «LSD1 является субъединицей комплекса NuRD и нацелен на программы метастазирования рака молочной железы» . Клетка . 138 (4): 660–72. дои : 10.1016/j.cell.2009.05.050 . ПМИД 19703393 .
- ^ Чикконе Д.Н., Су Х., Хеви С., Гей Ф., Лей Х., Байко Дж., Сюй Г., Ли Э., Чен Т. (сентябрь 2009 г.). «KDM1B представляет собой деметилазу гистона H3K4, необходимую для установления материнских геномных отпечатков». Природа . 461 (7262): 415–8. Бибкод : 2009Natur.461..415C . дои : 10.1038/nature08315 . ПМИД 19727073 . S2CID 1143128 .
- ^ Рудольф Т., Йонезава М., Лейн С., Хайдрих К., Кубичек С., Шефер С., Фальке С., Вальтер М., Шмидт А., Дженувейн Т., Рейтер Г. (апрель 2007 г.). «Формирование гетерохроматина у дрозофилы инициируется путем активного удаления метилирования H3K4 гомологом LSD1 SU(VAR)3-3» . Молекулярная клетка . 26 (1): 103–15. doi : 10.1016/j.molcel.2007.02.025 . ПМИД 17434130 .
- ^ Ди Стефано Л., Джи Дж.И., Мун Н.С., Герр А., Дайсон Н. (май 2007 г.). «Мутация Lsd1 дрозофилы нарушает метилирование H3-K4, что приводит к тканеспецифичным дефектам во время развития» . Современная биология . 17 (9): 808–12. дои : 10.1016/j.cub.2007.03.068 . ПМК 1909692 . ПМИД 17462898 .
- ^ Блэкледж Н.П., Чжоу Дж.К., Толсторуков М.Ю., Фаркас А.М., Парк П.Дж., Клозе Р.Дж. (апрель 2010 г.). «Островки CpG рекрутируют деметилазу лизина 36 гистона H3» . Молекулярная клетка . 38 (2): 179–90. doi : 10.1016/j.molcel.2010.04.009 . ПМК 3098377 . ПМИД 20417597 .
- ^ Хэ Дж., Каллин Э.М., Цукада Ю., Чжан Ю. (ноябрь 2008 г.). «Деметилаза H3K36 Jhdm1b/Kdm2b регулирует пролиферацию и старение клеток посредством p15 (Ink4b)» . Структурная и молекулярная биология природы . 15 (11): 1169–75. дои : 10.1038/nsmb.1499 . ПМК 2612995 . ПМИД 18836456 .
- ^ Ло Ю.Х., Чжан В., Чен Икс, Джордж Дж., Нг Х.Х. (октябрь 2007 г.). «Деметилазы гистона H3 Jmjd1a и Jmjd2c Lys 9 регулируют самообновление эмбриональных стволовых клеток» . Гены и развитие . 21 (20): 2545–57. дои : 10.1101/gad.1588207 . ПМК 2000320 . ПМИД 17938240 .
- ^ Эрбрехт А, Мюллер У, Вольтер М, Хойшен А, Кох А, Радлвиммер Б, Актер Б, Минчева А, Питш Т, Лихтер П, Райфенбергер Г, Вебер Р.Г. (март 2006 г.). «Комплексный геномный анализ десмопластических медуллобластом: идентификация новых амплифицированных генов и отдельная оценка различных гистологических компонентов». Журнал патологии . 208 (4): 554–63. дои : 10.1002/путь.1925 . ПМИД 16400626 . S2CID 1463027 .
- ^ Лю Дж., Боллиг-Фишер А., Крайке Б., ван де Вийвер М.Дж., Абрамс Дж., Этье С.П., Ян ZQ (декабрь 2009 г.). «Геномная амплификация и онкогенные свойства гена деметилазы гистонов GASC1 при раке молочной железы» . Онкоген . 28 (50): 4491–500. дои : 10.1038/onc.2009.297 . ПМК 2795798 . ПМИД 19784073 .
- ^ Норткотт П.А., Накахара И., Ву Х, Фейк Л., Эллисон Д.В., Кроул С., Мак С., Конгкхам П.Н., Пикок Дж., Дубук А., Ра Ю.С., Зильберберг К., МакЛеод Дж., Шерер С.В., Сунил Рао Дж., Эберхарт К.Г., Грайковска В., Гиллеспи Ю., Лах Б., Гранди Р., Поллак И.Ф., Гамильтон Р.Л., Ван Метер Т., Карлотти К.Г., Буп Ф., Бигнер Д., Гилбертсон Р.Дж., Рутка Дж.Т., Тейлор МД (апрель 2009 г.). «Множественные повторяющиеся генетические события сходятся в контроле метилирования лизина гистонов при медуллобластоме» . Природная генетика . 41 (4): 465–72. дои : 10.1038/ng.336 . ПМЦ 4454371 . ПМИД 19270706 .
- ^ Ян ZQ, Имото И, Фукуда Й, Пимкхаокхам А, Симада Й, Имамура М, Сугано С, Накамура Й, Инадзава Дж (сентябрь 2000 г.). «Идентификация нового гена, GASC1, в ампликоне 9p23-24, часто обнаруживаемого в клеточных линиях рака пищевода». Исследования рака . 60 (17): 4735–9. ПМИД 10987278 .
- ^ Клоос П.А., Кристенсен Дж., Аггер К., Майолика А., Раппсилбер Дж. , Антал Т., Хансен К.Х., Хелин К. (июль 2006 г.). «Предполагаемый онкоген GASC1 деметилирует три- и диметилированный лизин 9 на гистоне H3». Природа . 442 (7100): 307–11. Бибкод : 2006Natur.442..307C . дои : 10.1038/nature04837 . ПМИД 16732293 . S2CID 2874903 .
- ^ Ли Н., Эрджумент-Бромаж Х., Темпст П., Джонс Р.С., Чжан Ю. (март 2009 г.). «Покрышка деметилазы H3K4 связывается с деацетилазой гистонов Rpd3 и ингибирует ее» . Молекулярная и клеточная биология . 29 (6): 1401–10. дои : 10.1128/MCB.01643-08 . ПМЦ 2648242 . ПМИД 19114561 .
- ^ Беневоленская Е.В., Мюррей Х.Л., Брэнтон П., Янг Р.А., Кэлин В.Г. (июнь 2005 г.). «Связывание pRB с белком PHD RBP2 способствует клеточной дифференцировке» . Молекулярная клетка . 18 (6): 623–35. doi : 10.1016/j.molcel.2005.05.012 . ПМИД 15949438 .
- ^ Лопес-Бигас Н., Кисиэль Т.А., Деваал Д.С., Холмс К.Б., Волкерт Т.Л., Гупта С., Лав Дж., Мюррей Х.Л., Янг Р.А., Беневоленская Е.В. (август 2008 г.). «Полногеномный анализ деметилазы гистонов H3K4 RBP2 выявил программу транскрипции, контролирующую дифференцировку» . Молекулярная клетка . 31 (4): 520–30. doi : 10.1016/j.molcel.2008.08.004 . ПМЦ 3003864 . ПМИД 18722178 .
- ^ Пасини Д., Хансен К.Х., Кристенсен Дж., Аггер К., Клоос П.А., Хелин К. (май 2008 г.). «Координированная регуляция репрессии транскрипции с помощью деметилазы RBP2 H3K4 и поликомб-репрессивного комплекса 2» . Гены и развитие . 22 (10): 1345–55. дои : 10.1101/gad.470008 . ПМК 2377189 . ПМИД 18483221 .
- ^ ван Овелен С., Ван Дж., Асп П., Ян К., Кэлин В.Г., Клюгер Ю., Динлахт Б.Д. (ноябрь 2008 г.). «Роль Sin3 млекопитающих в постоянном молчании генов» . Молекулярная клетка . 32 (3): 359–70. doi : 10.1016/j.molcel.2008.10.015 . ПМК 3100182 . ПМИД 18995834 .
- ^ Цзэн Дж, Ге З, Ван Л, Ли Q, Ван Н, Бьёркхольм М, Цзя Дж, Сюй Д (март 2010 г.). «Гистондеметилаза RBP2 сверхэкспрессируется при раке желудка, и ее ингибирование вызывает старение раковых клеток». Гастроэнтерология . 138 (3): 981–92. дои : 10.1053/j.gastro.2009.10.004 . ПМИД 19850045 .
- ^ Йенсен Л.Р., Аменде М., Гурок У., Мозер Б., Гиммел В., Чах А., Джанеке А.Р., Таривердиан Г., Челли Дж., Фринс Дж.П., Ван Эш Х., Клифстра Т., Хамель Б., Морейн С., Геч Дж., Тернер Г., Рейнхардт Р., Кальшойер В.М., Роперс Х.Х., Ленцнер С. (февраль 2005 г.). «Мутации в гене JARID1C, который участвует в регуляции транскрипции и ремоделировании хроматина, вызывают Х-сцепленную умственную отсталость» . Американский журнал генетики человека . 76 (2): 227–36. дои : 10.1086/427563 . ПМК 1196368 . ПМИД 15586325 .
- ^ Ивасе С., Лан Ф., Бэйлисс П., де ла Торре-Убьета Л., Уарте М., Ци Х.Х., Уэтстайн Дж.Р., Бонни А., Робертс Т.М., Ши Ю. (март 2007 г.). «Х-сцепленный ген умственной отсталости SMCX/JARID1C определяет семейство лизин-4-деметилаз гистона H3» . Клетка . 128 (6): 1077–88. дои : 10.1016/j.cell.2007.02.017 . ПМИД 17320160 .
- ^ Аггер К., Клоос П.А., Радкьяер Л., Уильямс К., Андерсен Г., Кристенсен Дж., Хелин К. (май 2009 г.). «Деметилаза H3K27me3 JMJD3 способствует активации локуса INK4A-ARF в ответ на старение, вызванное онкогеном и стрессом» . Гены и развитие . 23 (10): 1171–6. дои : 10.1101/gad.510809 . ПМЦ 2685535 . ПМИД 19451217 .
- ^ Ледерер Д., Грисар Б., Диджилио М.К., Бенуа В., Креспен М., Гариани СК, Майстадт И., Даллапиккола Б., Вереллен-Дюмулен К. (январь 2012 г.). «Удаление KDM6A, деметилазы гистонов, взаимодействующей с MLL2, у трех пациентов с синдромом Кабуки» . Американский журнал генетики человека . 90 (1): 119–24. дои : 10.1016/j.ajhg.2011.11.021 . ПМЦ 3257878 . ПМИД 22197486 .
- ^ , Цурусаки Ю., Накашима М., Сайтсу Х., Ниикава Н., Мацумото Н. (январь 2013 г.) KDM6A вызывают синдром Кабуки» . «Точковые мутации Мияке Н., Мизуно С., Окамото Н., Охаси Х., Шиина М. , Огата К. : 108–10. doi : 10.1002/humu.22229 . PMID 23076834. (1 ) S2CID 1745473 .
- ^ Лан Ф., Бейлисс П.Е., Ринн Дж.Л., Уэтстин Дж.Р., Ван Дж.К., Чен С., Ивасе С., Алпатов Р., Исаева И., Канаани Э., Робертс Т.М., Чанг ХИ, Ши Ю (октябрь 2007 г.). «Деметилаза лизина 27 гистона H3 регулирует заднее развитие животных». Природа . 449 (7163): 689–94. Бибкод : 2007Natur.449..689L . дои : 10.1038/nature06192 . ПМИД 17851529 . S2CID 612144 .
- ^ Jump up to: а б Аггер К., Клоос П.А., Кристенсен Дж., Пасини Д., Роуз С., Раппсилбер Дж., Исаева И., Канаани Е., Сальчини А.Е., Хелин К. (октябрь 2007 г.). «UTX и JMJD3 представляют собой деметилазы гистонов H3K27, участвующие в регуляции и развитии гена HOX». Природа 449 (7163): 731–4. Бибкод : 2007Nature.449..731A . дои : 10.1038/nature06145 . ПМИД 17713478 . S2CID 4413812 .
- ^ Ван Дж.К., Цай MC, Пулин Г., Адлер А.С., Чен С., Лю Х., Ши Ю, Чанг ХИ (февраль 2010 г.). «Гистондеметилаза UTX обеспечивает RB-зависимый контроль судьбы клеток» . Гены и развитие . 24 (4): 327–32. дои : 10.1101/gad.1882610 . ПМЦ 2816731 . ПМИД 20123895 .
- ^ Де Санта Ф, Тотаро МГ, Просперини Э, Нотарбартоло С, Теста Дж, Натоли Дж (сентябрь 2007 г.). «Гистон H3 лизин-27 деметилаза Jmjd3 связывает воспаление с ингибированием молчания генов, опосредованного поликомбами» . Клетка . 130 (6): 1083–94. дои : 10.1016/j.cell.2007.08.019 . ПМИД 17825402 .
- ^ Бургольд Т., Спреафико Ф., Де Санта Ф., Тотаро М.Г., Просперини Э., Натоли Г., Теста Г. (2008). «Деметилаза Jmjd3, специфичная для гистона H3, лизин-27, необходима для нейронной активности» . ПЛОС ОДИН . 3 (8): е3034. Бибкод : 2008PLoSO...3.3034B . дои : 10.1371/journal.pone.0003034 . ПМК 2515638 . ПМИД 18716661 .
- ^ Jump up to: а б Владимиров Н., Лёвдок Л., Лебедз Д., Суржик В. (декабрь 2008 г.). «Зависимость бактериального хемотаксиса от формы градиента и скорости адаптации» . PLOS Вычислительная биология . 4 (12): e1000242. Бибкод : 2008PLSCB...4E0242V . дои : 10.1371/journal.pcbi.1000242 . ПМЦ 2588534 . ПМИД 19096502 .
- ^ Парк С.Ю., Борбат П.П., Гонсалес-Бонет Дж., Бхатнагар Дж., Поллард А.М., Фрид Дж.Х., Билвес А.М., Крейн Б.Р. (май 2006 г.). «Реконструкция сборки рецептор-киназа хемотаксиса». Структурная и молекулярная биология природы . 13 (5): 400–7. дои : 10.1038/nsmb1085 . ПМИД 16622408 . S2CID 859928 .
- ^ Вест АХ, Мартинес-Хакерт Э., Сток AM (июль 1995 г.). «Кристаллическая структура каталитического домена метилэстеразы рецептора хемотаксиса, CheB». Журнал молекулярной биологии . 250 (2): 276–90. дои : 10.1006/jmbi.1995.0376 . ПМИД 7608974 .