Узел Ранвье
| Узел Ранвье | |
|---|---|
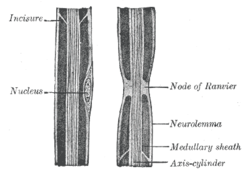 Рисунок аксона периферического нерва (с надписью «осевой цилиндр»), показывающий узел Ранвье вместе с другими особенностями. | |
 Узлы Ранвье | |
| Подробности | |
| Система | Нервная система |
| Расположение | Миелинизированный аксон нерва |
| Идентификаторы | |
| латинский | вырезка миелина |
| МеШ | D011901 |
| ТД | Х2.00.06.2.03015 |
| Анатомические термины микроанатомии | |
В нейробиологии и анатомии , узлы Ранвье ( / ˈ r ɑː n v i eɪ / RAHN -vee-ay ) [1] [2] Также известные как разрывы миелиновой оболочки , возникают вдоль миелинизированного аксона , где аксолемма выходит во внеклеточное пространство . Узлы Ранвье неизолированы и сильно обогащены ионными каналами , что позволяет им участвовать в обмене ионов, необходимых для регенерации потенциала действия . Нервная проводимость в миелинизированных аксонах называется скачкообразной проводимостью (от латинского saltus «прыжок», прыжок) из-за того, как потенциал действия «перескакивает» от одного узла к другому вдоль аксона. Это приводит к более быстрому проведению потенциала действия.
Обзор
[ редактировать ]
Аксоны многих позвоночных окружены миелиновой оболочкой, что обеспечивает быстрое и эффективное скачкообразное («прыгающее») распространение потенциалов действия. Контакты между нейронами и глиальными клетками характеризуются очень высоким уровнем пространственной и временной организации в миелинизированных волокнах. Миелинизирующие глиальные клетки — олигодендроциты в центральной нервной системе (ЦНС) и шванновские клетки в периферической нервной системе (ПНС) — обертываются вокруг аксона, оставляя аксолемму относительно непокрытой в регулярно расположенных узлах Ранвье.
Межузловые глиальные мембраны сливаются, образуя компактный миелин , тогда как заполненные цитоплазмой паранодальные петли миелинизирующих клеток спирально обертываются вокруг аксона по обе стороны от узлов. Такая организация требует жесткого контроля развития и формирования множества специализированных зон контакта между различными областями миелинизирующей клеточной мембраны. Каждый узел Ранвье окружен паранодальными областями, где спирально завернутые глиальные петли прикреплены к аксональной мембране с помощью перегородчатого соединения.
Сегмент между узлами Ранвье называется междоузлием , а его крайняя часть, соприкасающаяся с паранодами, называется юкстапаранодальной областью. Узлы инкапсулированы микроворсинками, исходящими из внешней части мембраны шванновских клеток в ПНС, или перинодальными отростками астроцитов в ЦНС.
Структура
[ редактировать ]Междоузлия представляют собой сегменты миелина , а промежутки между ними называются узлами. Размер и расстояние между междоузлиями изменяются в зависимости от диаметра волокна по криволинейной зависимости, оптимизированной для максимальной скорости проводимости. [3] Размер узлов варьируется от 1–2 мкм, тогда как длина междоузлий может достигать (а иногда и более) 1,5 миллиметров, в зависимости от диаметра аксона и типа волокна.
Структура узла и фланкирующих паранодальных областей отличается от междоузлий под компактной миелиновой оболочкой, но очень сходна в ЦНС и ПНС. Аксон подвергается воздействию внеклеточной среды в узле и сужается в диаметре. Уменьшенный размер аксона отражает более высокую плотность упаковки нейрофиламентов в этой области, которые менее фосфорилированы и транспортируются медленнее. [4] В узлах также увеличивается количество везикул и других органелл, что позволяет предположить, что существует узкое место в аксональном транспорте в обоих направлениях, а также в локальной аксонально-глиальной передаче сигналов.
При продольном срезе миелинизированной шванновской клетки в узле представлены три отличительных сегмента: стереотипное междоузлие , паранодальная область и сам узел. В межузловой области шванновская клетка имеет наружный воротник цитоплазмы, компактную миелиновую оболочку, внутренний воротник цитоплазмы и аксолемму. В паранодальных областях паранодальные петли цитоплазмы контактируют с утолщениями аксолеммы, образуя перегородчатоподобные соединения. Только в узле аксолемма контактирует с несколькими шванновскими микроворсинками и содержит плотный цитоскелетный подпокрытие.
Различия в центральной и периферической нервной системе
[ редактировать ]Хотя исследования замороженных переломов показали, что узловая аксолемма как в ЦНС, так и в ПНС обогащена внутримембранозными частицами (IMP) по сравнению с междоузлиями, существуют некоторые структурные различия, отражающие их клеточные составляющие. [4] В ПНС специализированные микроворсинки выступают из внешнего воротника шванновских клеток и очень близко подходят к узловой аксолемме крупных волокон. Проекции шванновских клеток перпендикулярны узлу и расходятся от центральных аксонов. Однако в ЦНС один или несколько астроцитарных отростков располагаются в непосредственной близости от узлов. Исследователи заявляют, что эти процессы происходят из-за многофункциональных астроцитов, а не из популяции астроцитов, предназначенных для контакта с узлом. С другой стороны, в ПНС базальная пластинка, окружающая шванновские клетки, непрерывна поперек узла.
Состав
[ редактировать ]Узлы обменников Ранвье Na+/Ca2+ и высокая плотность потенциалзависимых Na+-каналов, генерирующих потенциалы действия. Натриевый канал состоит из порообразующей α-субъединицы и двух вспомогательных β-субъединиц, которые привязывают канал к внеклеточным и внутриклеточным компонентам. Узлы Ранвье в центральной и периферической нервной системе состоят преимущественно из субъединиц αNaV1.6 и β1. [5] Внеклеточная область β-субъединиц может ассоциироваться сама с собой и с другими белками, такими как тенасцин R и молекулы клеточной адгезии нейрофасцин и контактин. Контактин также присутствует в узлах ЦНС, и взаимодействие с этой молекулой усиливает поверхностную экспрессию каналов Na+.
анкирин Было обнаружено, что связан со спектрином βIV, изоформой спектрина, обогащенной перехватами Ранвье и начальными сегментами аксона. Узлы ПНС окружены микроворсинками шванновских клеток, которые содержат ERM и EBP50, которые могут обеспечивать связь с актиновыми микрофиламентами. Несколько белков внеклеточного матрикса обогащены перехватами Ранвье, включая тенасцин-R , Bral-1 и протеогликан NG2, а также фосфакан и версикан V2. В узлах ЦНС аксональные белки также включают контактин; однако микроворсинки шванновских клеток заменены перинодальными отростками астроцитов .
Молекулярная организация
[ редактировать ]Молекулярная организация узлов соответствует их специализированной функции по распространению импульсов. Уровень натриевых каналов в узле по сравнению с междоузлием позволяет предположить, что количество ИМП соответствует натриевым каналам. Калиевые каналы практически отсутствуют в узловой аксолемме, тогда как они высоко сконцентрированы в паранодальной аксолемме и мембранах шванновских клеток в узелке. [4] Точная функция калиевых каналов не совсем выяснена, но известно, что они могут способствовать быстрой реполяризации потенциалов действия или играть жизненно важную роль в буферизации ионов калия в узлах. Это крайне асимметричное распределение потенциал-управляемых натриевых и калиевых каналов разительно контрастирует с их диффузным распределением в немиелинизированных волокнах. [4] [6]
Нитчатая сеть, расположенная под узловой мембраной, содержит белки цитоскелета, называемые спектрином и анкирином . Высокая плотность анкирина в узлах может быть функционально значимой, поскольку несколько белков, населяющих узлы, обладают способностью связываться с анкирином с чрезвычайно высоким сродством. Все эти белки, включая анкирин, сконцентрированы в начальном сегменте аксонов, что предполагает функциональную связь. Сейчас связь этих молекулярных компонентов с кластеризацией натриевых каналов в узлах до сих пор не известна. Хотя сообщалось, что некоторые молекулы клеточной адгезии непостоянно присутствуют в узлах; однако известно, что множество других молекул густо заселены глиальными мембранами паранодальных областей, где они способствуют ее организации и структурной целостности.
Разработка
[ редактировать ]Миелинизация нервных волокон
[ редактировать ]Сложные изменения, которые претерпевает шванновская клетка в процессе миелинизации периферических нервных волокон, наблюдались и изучались многими. Первоначальное охватывание аксона происходит непрерывно на всем протяжении шванновской клетки. Этот процесс осуществляется за счет складывания поверхности шванновских клеток так, что образуется двойная мембрана противоположных сторон свернутой поверхности шванновских клеток. Эта мембрана растягивается и спирально заворачивается снова и снова по мере того, как продолжается сворачивание поверхности шванновских клеток. В результате легко констатируется увеличение толщины расширения миелиновой оболочки в диаметре ее поперечного сечения. Также очевидно, что каждый из последовательных витков спирали увеличивается в размерах по длине аксона по мере увеличения числа витков. Однако неясно, можно ли объяснить увеличение длины миелиновой оболочки исключительно увеличением длины аксона, охватываемого каждым последующим витком спирали, как объяснялось ранее. В месте соединения двух шванновских клеток по аксону направления пластинчатого нависания миелиновых окончаний имеют противоположный смысл. [7] Это соединение, примыкающее к шванновским клеткам, образует область, называемую узлом Ранвье.
Ранние стадии
[ редактировать ]Исследователи доказывают, что в развивающейся ЦНС Nav1.2 изначально экспрессируется во всех формирующих узлах Ранвье. [8] По мере созревания узловой Nav1.2 подавляется и заменяется Nav1.6. Nav1.2 также экспрессируется во время формирования узла ПНС, что позволяет предположить, что переключение подтипов Nav-каналов является общим явлением в ЦНС и ПНС. В этом же исследовании было показано, что Nav1.6 и Nav1.2 колокализуются во многих узлах Ранвье во время ранней миелинизации. Это также привело к предположению, что ранним кластерам каналов Nav1.2 и Nav1.6 суждено позже стать узлами Ранвье. нейрофасцин Сообщается также, что является одним из первых белков, которые накапливаются во вновь образующихся узлах Ранвье. Также обнаружено, что они обеспечивают место зародышеобразования для прикрепления анкирина G, каналов Nav и других белков. [9] Недавняя идентификация белка микроворсинок шванновских клеток глиомедина в качестве вероятного партнера по связыванию аксонального нейрофасцина дает существенные доказательства важности этого белка в рекрутировании Nav-каналов в перехваты Ранвье. Более того, Ламберт и др. и Эшед и др. также указывает на то, что нейрофасцин накапливается перед каналами Nav и, вероятно, играет решающую роль в самых ранних событиях, связанных с формированием узла Ранвье. Таким образом, может существовать множество механизмов, которые работают синергетически для облегчения кластеризации навигационных каналов в узлах Ранвье.
Узловое образование
[ редактировать ]Первым событием, по-видимому, является накопление молекул клеточной адгезии, таких как NF186 или NrCAM. Внутриклеточные области этих молекул клеточной адгезии взаимодействуют с анкирином G, который служит якорем для натриевых каналов. В то же время периаксональное расширение глиальной клетки оборачивается вокруг аксона, образуя паранодальные области. Это движение вдоль аксона вносит значительный вклад в общее формирование узлов Ранвье, позволяя геминодам, образовавшимся на краях соседних глиальных клеток, сливаться в полные узлы.Септатоподобные соединения формируются в паранодах при обогащении NF155 в глиальных паранодальных петлях. Сразу после ранней дифференцировки нодальной и паранодальной областей калиевые каналы, Caspr2 и TAG1 накапливаются в околопаранодальных областях. Это накопление непосредственно совпадает с образованием компактного миелина. В зрелых узловых регионах взаимодействия с внутриклеточными белками кажутся жизненно важными для стабильности всех узловых регионов. В ЦНС, олигодендроциты не обладают микроворсинками, но, по-видимому, способны инициировать кластеризацию некоторых аксональных белков посредством секретируемых факторов. Комбинированные эффекты таких факторов с последующими движениями, вызванными обертыванием периаксонального расширения олигодендроцитов, могли бы объяснить организацию узлов Ранвье в ЦНС.
Функция
[ редактировать ]Потенциал действия
[ редактировать ]Потенциал действия — это всплеск как положительных, так и отрицательных ионных разрядов, который проходит вдоль мембраны клетки. [10] Создание и проведение потенциалов действия представляет собой фундаментальное средство коммуникации в нервной системе. Потенциалы действия представляют собой быстрое изменение напряжения на плазматической мембране аксонов. Эти быстрые изменения опосредуются потенциалзависимыми ионными каналами, обнаруженными в плазматической мембране . Потенциал действия перемещается из одного места клетки в другое, но поток ионов через мембрану происходит только в перехватах Ранвье. В результате сигнал потенциала действия скачет по аксону, от узла к узлу, а не распространяется плавно, как это происходит в аксонах, лишенных миелиновой оболочки. Кластеризация потенциал-управляемых ионных каналов натрия и калия в узлах допускает такое поведение.
Сальтаторная проводимость
[ редактировать ]Поскольку аксон может быть немиелинизированным или миелинизированным, у потенциала действия есть два способа перемещения по аксону. Эти методы называются непрерывной проводимостью для немиелинизированных аксонов и скачкообразной проводимостью для миелинизированных аксонов. Сальтаторная проводимость определяется как потенциал действия, движущийся дискретными скачками вниз по миелинизированному аксону.
Этот процесс описывается как пассивное распространение заряда на следующий узел Ранвье, чтобы деполяризовать его до порога, который затем запускает потенциал действия в этой области, который затем пассивно распространяется на следующий узел и так далее.
Сальтаторная проводимость дает одно преимущество перед проводимостью, которая происходит по аксону без миелиновой оболочки. Дело в том, что повышенная скорость, обеспечиваемая этим способом проводимости, обеспечивает более быстрое взаимодействие между нейронами. С другой стороны, в зависимости от средней скорости возбуждения нейрона расчеты показывают, что энергетические затраты на поддержание потенциала покоя олигодендроцитов могут перевесить экономию энергии потенциалов действия. [11] Таким образом, миелинизация аксонов не обязательно экономит энергию.
Регламент формирования
[ редактировать ]Регуляция паранодов посредством накопления митохондрий
[ редактировать ]Митохондрии и другие мембранные органеллы обычно богаты PNP-областью периферических миелинизированных аксонов, особенно аксонов большого калибра. [12] Фактическая физиологическая роль этого накопления и факторы, регулирующие его, не понятны; однако известно, что митохондрии обычно присутствуют в тех областях клетки, которые испытывают высокую потребность в энергии. Предполагается, что в этих же областях они содержат конусы роста, синаптические терминали и места инициации и регенерации потенциала действия, такие как узлы Ранвье. В синаптических окончаниях митохондрии производят АТФ, необходимую для мобилизации везикул для нейротрансмиссии. В узлах Ранвье митохондрии играют важную роль в проведении импульса, производя АТФ, необходимую для поддержания активности энергоемких ионных насосов. В подтверждение этого факта в аксоплазме ПНП крупных периферических аксонов присутствует примерно в пять раз больше митохондрий, чем в соответствующих межузловых участках этих волокон. [12]
Узловое регулирование
[ редактировать ]Через αII-спектрин
[ редактировать ]Сальтаторная проводимость в миелинизированных аксонах требует организации узлов Ранвье, тогда как потенциалзависимые натриевые каналы густонаселены. Исследования показывают, что αII-спектрин, компонент цитоскелета, на ранних стадиях обогащается в узлах и паранодах, а по мере созревания узлов экспрессия этой молекулы исчезает. [13] Также доказано, что αII-спектрин в аксональном цитоскелете абсолютно необходим для стабилизации кластеров натриевых каналов и организации зрелого узла Ранвье.
Возможная регуляция через молекулу узнавания OMgp.
[ редактировать ]Ранее было показано, что OMgp (олигодендроцитарный миелиновый гликопротеин) кластеризуется в узлах Ранвье и может регулировать паранодальную архитектуру, длину узла и отрастание аксонов в узлах. [14] Однако последующее исследование показало, что антитело, использованное ранее для идентификации OMgp в узлах, перекрестно реагирует с другим компонентом, обогащенным узлами, версиканом V2, и что OMgp не требуется для целостности узлов и паранодов, что противоречит ранее сообщенной локализации и предложенным функциям. OMgp на узлах. [15]
Клиническое значение
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( март 2018 г. ) |
Белки в этих возбудимых доменах нейронов при повреждении могут привести к когнитивным расстройствам и различным нейропатическим заболеваниям.
История
[ редактировать ]
Миелиновую оболочку длинных нервов открыл и назвал немецкий патологоанатом . Рудольф Вирхов [16] в 1854 году. [17] Французский патологоанатом и анатом Луи-Антуан Ранвье позже обнаружил узлы или разрывы в миелиновой оболочке, которые теперь носят его имя. Ранвье родился в Лионе и был одним из самых выдающихся гистологов конца 19 века. Ранвье отказался от патологических исследований в 1867 году и стал ассистентом физиолога Клода Бернара . Он был председателем общей анатомии в Коллеж де Франс в 1875 году.
Его усовершенствованные гистологические методы и работы как с поврежденными, так и с нормальными нервными волокнами стали всемирно известными. Его наблюдения за узлами волокон, а также за дегенерацией и регенерацией перерезанных волокон оказали большое влияние на парижскую неврологию в Сальпетриере . Вскоре после этого он обнаружил разрывы в оболочках нервных волокон, которые позже были названы узлами Ранвье. Это открытие позже привело Ранвье к тщательному гистологическому исследованию миелиновых оболочек и шванновских клеток. [18]
Дополнительные изображения
[ редактировать ] Полная схема нейронных клеток
Полная схема нейронных клеток Медуллированные нервные волокна, окрашенные нитратом серебра.
Медуллированные нервные волокна, окрашенные нитратом серебра.


См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «узел Ранвье» . Lexico Британский словарь английского языка . Издательство Оксфордского университета . Архивировано из оригинала 16 октября 2021 года.
- ^ «узел Ранвье» . Словарь Merriam-Webster.com .
- ^ gxnSalzer JL (1997). «Кластеризация натриевых каналов в узле Ранвье: близкие контакты типа аксон-глия» . Нейрон . 18 (6): 843–846. дои : 10.1016/S0896-6273(00)80323-2 . ПМИД 9208851 . S2CID 6743084 .
- ^ Jump up to: а б с д Зальцер Дж.Л. (1997). «Кластеризация натриевых каналов в узле Ранвье: близкие контакты типа аксон-глия» . Нейрон . 18 (6): 843–846. дои : 10.1016/S0896-6273(00)80323-2 . ПМИД 9208851 . S2CID 6743084 .
- ^ Каплан М.Р.; Чо М.Х.; Уллиан ЭМ; Исом ЛЛ; Левинсон С.Р.; Баррес Б.А. (2001). «Дифференциальный контроль кластеризации натриевых каналов Na(v)1,2 и Na(v)1,6 при развивающихся узлах Ранвье ЦНС» . Нейрон . 30 (1): 105–119. дои : 10.1016/S0896-6273(01)00266-5 . ПМИД 11343648 . S2CID 10252129 .
- ^ Блэк, Дж. А., Сонтхаймер, Х., О, Ю. и Ваксман, С. Г. (1995). В «Аксоне » С. Ваксман, Дж. Кочис и П. Стис, ред. Издательство Оксфордского университета , Нью-Йорк , стр. 116–143.
- ^ Узмман Б.Г.; Ногейра-Граф Г. (1957). «Электронно-микроскопическое исследование формирования узлов Ранвье седалищных нервов мыши» . Журнал биофизической и биохимической цитологии . 3 (4): 589–597. дои : 10.1083/jcb.3.4.589 . ПМК 2224104 . ПМИД 13449102 .
- ^ Бойко Т., Расбанд М.Н., Левинсон С.Р., Колдуэлл Дж.Х., Мандель Г., Триммер Дж.С. и др. (2001). «Компактный миелин диктует различное нацеливание двух изоформ натриевых каналов в один и тот же аксон» . Нейрон . 30 (1): 91–104. дои : 10.1016/S0896-6273(01)00265-3 . ПМИД 11343647 . S2CID 7168889 .
- ^ Ламберт С., Дэвис Дж.К., Беннетт В. (1997). «Морфогенез узла Ранвье: совместные кластеры анкирина и анкирин-связывающих интегральных белков определяют ранние промежуточные продукты развития» . Журнал неврологии . 17 (18): 7025–7036. doi : 10.1523/JNEUROSCI.17-18-07025.1997 . ПМК 6573274 . ПМИД 9278538 .
- ^ Фрай, К. (2007). «Клеточная физиология I». Хирургия (Оксфорд) . 25 (10): 425–429. дои : 10.1016/j.mpsur.2007.07.007 . S2CID 57536809 .
- ^ Харрис; Этвуд (2012). «Энергетика белого вещества ЦНС» . Журнал неврологии . 32 (1): 356–371. doi : 10.1523/JNEUROSCI.3430-11.2012 . ПМЦ 3272449 . ПМИД 22219296 .
- ^ Jump up to: а б Эйнхебер С., Бхат М.А., Зальцер Дж.Л. (август 2006 г.). «Нарушение аксо-глиальных соединений приводит к накоплению аномальных митохондрий в узлах Ранвье» . Биология нейронов глии . 2 (3): 165–174. дои : 10.1017/S1740925X06000275 . ПМЦ 1855224 . ПМИД 17460780 .
- ^ Воас М.Г., Лайонс Д.А., Нейлор С.Г., Арана Н., Расбанд М.Н., Талбот В.С. (март 2007 г.). «АльфаII-спектрин необходим для сборки узлов Ранвье в миелинизированных аксонах» . Современная биология . 17 (6): 562–8. Бибкод : 2007CBio...17..562V . дои : 10.1016/j.cub.2007.01.071 . ПМИД 17331725 . S2CID 14537696 .
- ^ Хуанг, Дж. К.; Филлипс, Греция; Рот, AD; Педраса, Л; Шан, В; Белкаид, В; Ми, С; Фекс-Свеннингсен, А; Флоренс, Л; Йейтс III, младший; Колман, Д.Р. (2005). «Глиальные мембраны узла Ранвье предотвращают рост нейритов» . Наука . 310 (5755): 1813–17. Бибкод : 2005Sci...310.1813H . дои : 10.1126/science.1118313 . ПМИД 16293723 . S2CID 17410200 .
- ^ Чанг, К.Дж.; Сусуки, К; Дурс-Циммерманн, Монтана; Циммерманн, доктор медицинских наук; Расбанд, Миннесота (2010). «Олигодендроцитарный миелиновый гликопротеин не влияет на структуру или сборку узла Ранвье» . Дж. Нейроски . 30 (43): 14476–81. doi : 10.1523/JNEUROSCI.1698-10.2010 . ПМЦ 2976578 . ПМИД 20980605 .
- ^ Synd/3816 в Who Named It?
- ^ Вирхов Р. (1854). «О широком распространении вещества, аналогичного нервному мозгу, в тканях животных» . Архивы по патологической анатомии и физиологии и клинической медицине . 6 (4): 562–572. дои : 10.1007/BF02116709 . S2CID 20120269 .
- ^ Барбара Дж.Г. (2005). «Кольцевые дроссели Луи Ранвье (1871 г.)» (PDF) . Письмо по неврологии . 28 :3–5.
Внешние ссылки
[ редактировать ]- База данных, ориентированная на ячейки – узел Ранвье
- Анатомическое фото: нервный/pns/nerve2/nerve5 - Сравнительная органология Калифорнийского университета в Дэвисе - «ПНС, нерв (LM, средний)»
| Базы данных органов управления : Национальные |
|---|