Мышечное сокращение

Сокращение мышц -это активация сайтов, генерирующих напряжение в мышечных клетках . [ 1 ] [ 2 ] В физиологии сокращение мышц не обязательно означает укорочение мышц, потому что мышечное напряжение может быть вызвано без изменений в длине мышц, например, при хранении чего -то тяжелого в одном и том же положении. [ 1 ] За завершением сокращения мышц сопровождается релаксацией мышц , которое представляет собой возвращение мышечных волокон в их низкое состояние, порождающее напряжение. [ 1 ]
Для того, чтобы схватки произошли, мышечные клетки должны полагаться на изменение действия двух типов филаментов: тонких и толстых филаментов.
Основной составляющей тонких филаментов является цепь, образованная спиральной панелью двух цепей актина , и толстые филаменты, преимущественно, состоят из цепей моторного белка миозина . Вместе эти два нити образуют миофибриллы - основные функциональные органеллы в скелетной мышечной системе.
У позвоночных , сокращения скелетных мышц являются нейрогенными поскольку они требуют синаптического ввода от моторных нейронов . Один моторный нейрон способен иннервировать множество мышечных волокон, что приводит к тому, что волокна сокращаются одновременно. Оказавшись иннервируемым, белковые филаменты в каждой скелетной мышечной волокне проскользнули мимо друг друга, чтобы вызвать сокращение, которое объясняется теорией скользящей филаменты . Полученное сокращение может быть описано как подергивание , суммирование или столб, в зависимости от частоты потенциалов действия . В скелетных мышцах мышечное напряжение в самом наибольшем количестве, когда мышца растягивается до промежуточной длины, как описано в зависимости от отношения длины.
В отличие от скелетных мышц, сокращения гладких и сердечных мышц являются миогенными (что означает, что они инициируются самими клетками гладких или сердечных мышц, а не стимулируются внешним событием, таким как нервное стимуляция), хотя они могут модулироваться стимулами из вегетативная нервная система . Механизмы сокращения в этих мышечных тканях аналогичны механизмам в тканях скелетных мышц.
Сокращение мышц также может быть описано с точки зрения двух переменных: длина и напряжение. [ 1 ] В естественных движениях, лежащих в основе локомоторной активности , мышечные сокращения многогранны, поскольку они способны производить изменения в длине и напряжении изменяющимся временем. [ 3 ] Следовательно, ни длина, ни напряжение, вероятно, не останутся одинаковыми в скелетных мышцах , которые сокращаются во время передвижения. Сокращения могут быть описаны как изометрические, если мышечное напряжение изменяется, но длина мышц остается прежней. [ 1 ] [ 4 ] [ 5 ] [ 6 ] Напротив, мышечное сокращение описывается как изотоническое , если мышечное напряжение остается неизменным на протяжении всего сокращения. [ 1 ] [ 4 ] [ 5 ] [ 6 ] Если длина мышц сокращается, сокращение является концентрическим; [ 1 ] [ 7 ] Если длина мышц удлиняется, сокращение является эксцентричным.
Типы
[ редактировать ]
Мышечные сокращения могут быть описаны на основе двух переменных: сила и длина. Сама сила может быть дифференцирована как натяжение или нагрузку. Мышечное напряжение - это сила, оказываемая мышцей на объекте, тогда как нагрузка - это сила, оказываемое объектом на мышцах. [ 1 ] Когда мышечное напряжение изменяется без каких -либо соответствующих изменений в длине мышц, сокращение мышц описывается как изометрическое. [ 1 ] [ 4 ] [ 5 ] [ 6 ] Если длина мышц меняется, когда мышечное напряжение остается неизменным, то сокращение мышц является изотоническим. [ 1 ] [ 4 ] [ 5 ] [ 6 ] В изотоническом сокращении длина мышц может либо сократить, чтобы произвести концентрическое сокращение, либо удлинять, чтобы вызвать эксцентрическое сокращение. [ 1 ] [ 7 ] В естественных движениях, лежащих в основе локомоторной активности, мышечные сокращения многогранны, поскольку они способны производить изменения в длине и напряжении изменяющимся временем. [ 3 ] Следовательно, ни длина, ни напряжение, вероятно, не останутся постоянными, когда мышца активна во время локомоторной активности.
Изометрическое сокращение
[ редактировать ]Изометрическое сокращение мышцы генерирует напряжение без изменения длины. [ 1 ] [ 4 ] [ 5 ] [ 6 ] Пример можно найти, когда мышцы руки и предплечья захватывают объект; Суставы руки не движутся, но мышцы генерируют достаточную силу , чтобы предотвратить сброс объекта.
Изотоническое сокращение
[ редактировать ]В изотоническом сокращении напряжение в мышцах остается постоянным, несмотря на изменение длины мышц. [ 1 ] [ 4 ] [ 5 ] [ 6 ] Это происходит, когда сила сжатия мышцы соответствует общей нагрузке на мышцу.
Концентрическое сокращение
[ редактировать ]В концентрическом сокращении мышечное напряжение достаточно для преодоления нагрузки, и мышцы сокращаются при сжимании. [ 8 ] Это происходит, когда сила, генерируемая мышцей, превышает нагрузку, противостоящую ее сокращению.
Во время концентрического сокращения мышца стимулируется для сжимания в соответствии с теорией скольжения . Это происходит по всей длине мышцы, генерируя силу при происхождении и вставке, в результате чего мышца сокращается и изменяет угол сустава. По отношению к локте , концентрическое сокращение бицепса заставило бы руку сгибаться в локте, когда рука перемещалась с ноги к плечу ( сгибание бицепса ). Концентрическое сокращение трицепса изменяет угол сустава в противоположном направлении, выпрямив руку и перемещая руку к ноге.
Эксцентричное сокращение
[ редактировать ]В эксцентрическом сокращении напряжение, генерируемое, в то время как изометрическое недостаточно для преодоления внешней нагрузки на мышцы, а мышечные волокна удлиняются при сжимании. [ 9 ] Вместо того, чтобы работать над тем, чтобы потянуть сустав в направлении сжатия мышц, мышца действует, чтобы замедлить сустав в конце движения или иным образом контролировать перемещение нагрузки. Это может произойти невольно (например, при попытке перемещать вес, слишком тяжелый, чтобы мышцы могли поднять) или добровольно (например, когда мышца «сглаживает» движение или сопротивляется гравитации, например, во время ходьбы вниз). В краткосрочной перспективе силовые тренировки с участием как эксцентрических, так и концентрических сокращений, по-видимому, увеличивают мышечную силу больше, чем тренировки с концентрическими сокращениями. [ 10 ] Однако вызванное физическим упражнением повреждение мышц также больше во время удлинительных сокращений. [ 11 ]
Во время эксцентричного сокращения мышцы бицепса локоть начинает движение, а затем выпрямляется, когда рука уходит от плеча . Во время эксцентричного сокращения мышцы трицепса локоть начинает движение прямо, а затем изгибается, когда рука движется к плечу. Desmin , Titin и другие z-line белки участвуют в эксцентрических сокращениях, но их механизм плохо изучен по сравнению с цикличностью поперечного моста в концентрических сокращениях. [ 9 ]
Хотя мышца выполняет негативное количество механической работы , (работа выполняется на мышцах), тем не менее потребляется химическая энергия (жира или глюкозы или временно хранится в АТФ ), хотя и меньше, чем поглощена во время концентрического сокращения. той же силы. Например, каждый тратит больше энергии, поднимающееся по лестнице, чем на один и тот же рейс.
Мышцы, подвергающиеся тяжелой эксцентрической нагрузке, испытывают больший ущерб при перегрузке (например, во время наращивания мышечных или силовых тренировок ) по сравнению с концентрической нагрузкой. Когда эксцентрические сокращения используются в силовых тренировках, они обычно называют негативами . Во время концентрического сокращения сократительные мышечные проскользнули мимо друга друг миофиламенты миозина и актина , сталкивая Z-линии вместе. Во время эксцентрического сокращения миофиламенты проскользнули мимо друг друга противоположным образом, хотя фактическое движение голов миозина во время эксцентрического сокращения неизвестно. Упражнения с тяжелой эксцентрической нагрузкой могут фактически поддерживать больший вес (мышцы примерно на 40% сильнее во время эксцентрических сокращений, чем во время концентрических сокращений), а также приводит к большему мышечному повреждению и задержке мышечной болезненности мышечной массы через один -два дня после тренировки. Упражнения, которые включают как эксцентрические, так и концентрические мышечные сокращения (то есть, включающие сильное сокращение и контролируемое снижение веса), могут привести к большему увеличению силы, чем только концентрические сокращения. [ 10 ] [ 12 ] В то время как неубедительные эксцентричные сокращения могут легко привести к перетренированию , умеренная тренировка может обеспечить защиту от травм. [ 10 ]
Эксцентричные сокращения в движении
[ редактировать ]Эксцентрические сокращения обычно возникают в виде тормозной силы в противодействии концентрическому сокращению для защиты суставов от повреждения. Во время практически любого обычного движения эксцентричные сокращения помогают поддерживать плавные движения, но также могут замедлить быстрые движения, такие как удар или бросок. Часть обучения для быстрых движений, таких как подача во время бейсбола, включает в себя снижение эксцентричного торможения, позволяя развивать большую силу на протяжении всего движения.
Эксцентрические сокращения исследуются за их способность ускорить реабилитацию слабых или раненых сухожилий. Ахиллес тендинит [ 13 ] [ 14 ] и коленник [ 15 ] (Также было показано, что также известно как коленное судно-коленное джемпер), как было показано, выигрывают от эксцентрических сокращений с высокой нагрузкой.
Позвоночные
[ редактировать ]
У животных позвоночных есть три типа мышечных тканей : скелетные, гладкие и сердечные. Скелетная мышца представляет собой большую часть мышечной массы в организме и отвечает за локомоторную активность. Гладкие мышцы образуют кровеносные сосуды , желудочно -кишечный тракт и другие области тела, которые вызывают устойчивые сокращения. Сердечная мышца составляет сердце, которое перекачивает кровь. Скелетные и сердечные мышцы называются полосатыми мышцами из -за их полосатого появления под микроскопом, который связан с высокоорганизованным чередующимся рисунком полос и I полос.
Скелетная мышца
[ редактировать ]
За исключением рефлексов, все сокращения скелетных мышц происходят в результате сигналов, возникающих в мозге. Мозг посылает электрохимические сигналы через нервную систему в моторный нейрон , который иннервирует несколько мышечных волокон. [ 16 ] В случае некоторых рефлексов сигнал к сокращению может возникнуть в спинном мозге через петлю обратной связи с серого вещества. Другие действия, такие как локомоция, дыхание и жевание, имеют для них рефлекторный аспект: сокращения могут быть инициированы либо сознательно, либо неосознанно.
Нервно -мышечное соединение
[ редактировать ]
Нервно -мышечное соединение - это химический синапс, образованный контактом между моторным нейроном и мышечным волокном . [ 17 ] Это место, в котором моторный нейрон передает сигнал в мышечное волокно, чтобы инициировать сокращение мышц. Последовательность событий, которые приводят к деполяризации мышечного волокна в нервно -мышечном соединении, начинается, когда потенциал действия инициируется в теле клеток моторного нейрона, который затем распространяется в результате соленой проводимости вдоль его аксона в направлении нервно -мышечного соединения. Как только он достигнет терминального бутона , потенциал действия вызывает CA 2+
Ионовый приток в терминал посредством управляемых напряжением кальциевых каналов . Ca 2+
Приток вызывает синаптические везикулы, содержащие нейротрансмиттер ацетилхолин , с помощью плазматической мембраны, высвобождая ацетилхолин в синаптическую расщелину между терминалом моторного нейрона и нервно -мышечным соединением волокна скелетных мышц. Ацетилхолиновые диффузы через синапс и связывают и активирует никотиновые ацетилхолиновые рецепторы на нервно -мышечном соединении. Активация никотинового рецептора открывает свой внутренний канал натрия / калия , вызывая натрия в спешке и калий, чтобы вытечь. В результате сарколемма реверсирует полярность, и ее напряжение быстро прыгает с мембранного потенциала покоя от -90 мВ до +75 мВ, как поступает натрий. Затем мембранный потенциал становится гиперполяризованным при выходе из калия, а затем скорректируется обратно к потенциалу покоящейся мембраны. Это быстрое колебание называется потенциалом конечной пластины. [ 18 ] Ионные каналы, управляемые напряжением сарколеммы рядом с конечной пластиной, открываются в ответ на потенциал конечной пластины. Они специфичны для натрия и калия и позволяют только один. Эта волна ионных движений создает потенциал действия, который распространяется от конечной пластины двигателя во всех направлениях. [ 18 ] Если потенциалы действия перестают прибывать, то ацетилхолин перестает освобождать от терминального бутона. Оставшийся ацетилхолин в синаптической расщелине либо деградируется активной ацетилхолинэстеразой , либо реабсорбируется синаптической ручкой, и ни один из них не остается для замены разлагаемого ацетилхолина.
Объединение возбуждения - конструкции
[ редактировать ]Связание с возбуждением -контролем (ECC) - это процесс, посредством которого потенциал мышечного действия в мышечном волокне вызывает миофибриллов сокращение . В скелетных мышцах соединение возбуждения-контроля зависит от прямой связи между двумя ключевыми белками, каналом высвобождения кальция саркоплазматического ретикулума (SR), идентифицированный как кальциевый Ryanodine , и кальциевый канал L-типа, управляемый напряжением рецептор . , (DHPR). DHPR расположены на сарколемме (которая включает в себя поверхностную сарколемму и поперечные канальцы ), в то время как Ryrs проживают через мембрану SR. Тесное приложение поперечного канальца и двух областей SR, содержащих Ryrs, описывается как триада и преимущественно, когда происходит связь возбуждения -контроля.

Соединение возбуждения -контроля (ECC) происходит, когда деполяризация скелетных мышц (обычно посредством нейронной иннервации) приводит к потенциалу действий мышц. Этот потенциал действия распространяется по поверхности мышцы и в сеть Т-тубов мышечных волокон , деполяризуя внутреннюю часть мышечного волокна. Это активирует дигидропиридиновые рецепторы в терминальных цистерне , которые находятся в непосредственной близости от рецепторов райанодина в соседней саркоплазматической ретикулуме . Активированные дигидропиридиновые рецепторы физически взаимодействуют с рецепторами раинодина, чтобы активировать их посредством процессов стопы (включающих конформационные изменения, которые аллостерически активируют рецепторы раинодина). Как открываются рецепторы райанодина, CA 2+ высвобождается из саркоплазматической ретикулумы в локальное соединительное пространство и диффундирует в объемную цитоплазму, чтобы вызвать кальциевую искру . [ 19 ] Потенциал действия создает почти синхронную активацию тысяч искры кальция и вызывает увеличение кальция по всему клеткам, что приводит к повышению уровня переходного процесса кальция . Ca 2+ Выпущенные в цитозоль связываются с тропонином С актиновыми филаментами . Эта связь позволяет актиновым филаментам выполнять циклу поперечного моста , производя силу и, в некоторых ситуациях, движусью.
Когда желаемое движение будет достигнуто, расслабление может быть быстро достигнуто по многочисленным путям. Расслабление быстро достигается через CA 2+ буфер с различными цитоплазматическими белками, связывающими с CA 2+ с очень высоким сродством. [ 20 ] Эти цитоплазматические белки обеспечивают быструю релаксацию в быстрых мышцах. Несмотря на медленнее, SARCO/Эндоплазматический ретикулум кальций-АТФаза (SERCA) активно накачивает CA 2+ Вернуться в саркоплазматический ретикулум, что приводит к постоянному расслаблению до появления следующего потенциала действия. [ 19 ]
Митохондрия также участвует в СА 2+ обратный захват, в конечном итоге доставив их собрание CA 2+ к SERCA для хранения в саркоплазматической ретикулуме. Несколько механизмов релаксации (NCX, насосы CA2+ и каналы утечки CA2+) также полностью выводят CA2+ из клеток. [ 21 ] Как 2+ Концентрация снижается до уровня покоя, CA2+ выпускает от тропонина С, не давая перекрестного мостового цикла, что приводит к снижению силы и возникновению релаксации. После того, как релаксация полностью произошла, мышца сможет снова сжиматься, таким образом, полностью сбросить цикл.
Теория скользящей нити
[ редактировать ]
Теория скользящей нити описывает процесс, используемый мышцами для сокращения. Это цикл повторяющихся событий, который заставляет тонкую нить скользить по толстой нити и генерировать натяжение в мышцах. [ 22 ] Это было независимо разработано Эндрю Хаксли и Рольфом Нидергером, а также Хью Хаксли и Джин Хансон в 1954 году. [ 23 ] [ 24 ] Физиологически, это сокращение не является равномерным по всему саркоме; Центральное положение толстых филаментов становится нестабильным и может сдвигаться во время сокращения, но это противоречит действию упругого миофиламента титина . Этот тонкий миофиламент поддерживает равномерное натяжение через саркомер, вытягивая толстую нить в центральное положение. [ 25 ]
Цикл перекрестного моста
[ редактировать ]
Цикл перекрестного моста-это последовательность молекулярных событий, которые лежат в основе теории скользящей филаменты. Поперечный мост -это проекция миозина, состоящая из двух голов миозина, которая простирается от толстых филаментов. [ 1 ] Каждая головка миозина имеет два сайта связывания: один для аденозин трифосфат (АТФ) и другой для актина. Связывание АТФ с головой миозина отделяет миозин от актина , что позволяет миозину связываться с другой молекулой актина. После прикрепления АТФ гидролизуется миозином, который использует высвобожденную энергию, чтобы перейти в «петушеное положение», в результате чего он слабо связывается с частью сайта связывания актина. Остальная часть сайта связывания актина блокируется тропомиозином . [ 26 ] С гидролизованным АТФ головка миозина с миозином содержит аденозинфосфат (ADP) + P i . Два ок 2+
Ионы связываются с тропонином C на актиновых филаментах. Troponin- ca 2+
Комплекс заставляет тропомиозин скользить и разблокировать оставшуюся часть сайта связывания актина. Распыление остальных сайтов связывания актина позволяет двум головкам миозина закрываться, а миозин тесно связывается с актином. [ 26 ] Затем головка миозина высвобождает неорганический фосфат и инициирует силовой удар, который генерирует силу 2 пн. Силовой удар перемещает актиновую нить внутрь, тем самым сокращая саркомер . Затем миозин выпускает ADP, но все еще остается тесно связанным с актином. В конце мощного удара ADP освобождается от головы миозина, оставляя миозин прикреплен к актину в строгого состоянии, пока другой АТФ не свяжется с миозином. Отсутствие АТФ приведет к характеристике строгого состояния . Когда-то другой АТФ связывается с миозином, голова миозина снова отречься от актина, и происходит еще один цикл поперечного моста.
Cross Bridge Cycling может продолжаться до тех пор, пока существует достаточное количество ATP и CA 2+
в цитоплазме. [ 26 ] Окончание велосипедного велосипеда может произойти, когда CA 2+
обратно активно перекачивается в саркоплазматический ретикулум. Когда ок 2+
Больше не присутствует на тонкой нити, тропомиозин изменяет конформацию обратно в свое предыдущее состояние, чтобы снова заблокировать сайты связывания. Миозин перестает связывать с тонкой нити, и мышца расслабляется. Ca 2+
Ионы покидают молекулу тропонина, чтобы поддерживать CA 2+
Концентрация ионов в саркоплазме. Активная накачка CA 2+
Ионы в саркоплазматическую ретикулуму создают дефицит жидкости вокруг миофибриллов. Это вызывает удаление CA 2+
Ионы из тропонина. Таким образом, комплекс тропомиозин-тропонина снова охватывает сайты связывания на актиновых филаментах, а сокращение прекращается.
Градация скелетных сокращений мышц
[ редактировать ]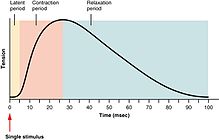

Сила сокращений скелетных мышц может быть широко разделена на подергивание , суммирование и столб . Подергивание - это единственный сжатие и цикл релаксации, порождаемый потенциалом действия в самом мышечном волокне. [ 27 ] Время между стимулом к двигательному нерву и последующим сокращением иннервированной мышцы называется скрытым периодом , который обычно занимает около 10 мс и вызвано временем для потенциала нервного действия для распространения, время для химической передачи на Нервно-мышечное соединение, затем последующие шаги в связи с возбуждением. [ 28 ]
Если еще один потенциал мышечного действия должен был быть получен до полного расслабления мышечного подергивания, то следующий подергивание просто подведет под предыдущий подергивание, что приведет к суммированию . [ 29 ] Суммирование может быть достигнуто двумя способами: [ 30 ] суммирование частоты и суммирование множественного волокна . При суммировании частоты сила, оказываемая скелетной мышцей, контролируется изменением частоты, с которой потенциалы действия отправляются в мышечные волокна. Потенциалы действия не приходят к мышцам синхронно, и во время сокращения некоторая часть волокон в мышцах будет стрелять в любой момент времени. В типичных обстоятельствах, когда люди оказывают свои мышцы так же сильно, как они сознательно способны, примерно одна треть волокна в каждой из этих мышц будет сразу же стрелять [ Цитация необходима ] , хотя это соотношение может зависеть от различных физиологических и психологических факторов (включая органы сухожилия Гольджи и клетки Renshaw ). Этот «низкий» уровень сокращения является защитным механизмом для предотвращения авульсии сухожилия - сила, полученная 95% сокращением всех волокон, достаточна для повреждения тела. В суммировании множественного волокна , если центральная нервная система посылает слабый сигнал для сжимания мышцы, более мелкие моторные единицы , более возбудимые, чем более крупные, сначала стимулируются. По мере увеличения силы сигнала , в дополнение к более крупным моторным единицам, при этом самые большие моторные единицы в 50 раз превышают сократительную прочность, чем меньшие. По мере того, как все больше и больше моторных единиц активируются, сила сокращения мышц становится все более сильной. Концепция, известная как принцип размера, позволяет градации мышечной силы во время слабых сокращений, возникающих небольшими шагами, что затем становятся все больше, когда требуется большее количество силы.
Наконец, если частота потенциалов действий мышц увеличивается так, что сокращение мышц достигает своей пиковой силы и плато на этом уровне, то сокращение является столбцом .
Отношения натяжения длины
[ редактировать ]
Отношение натяжения длины связывает силу изометрического сокращения с длиной мышц, при которой происходит сокращение. Мышцы работают с наибольшим активным напряжением вблизи идеальной длины (часто их длины покоя). При растяжении или сокращении этого (будь то из -за действия самой мышцы или внешней силой), максимальное активное напряжение уменьшается. [ 31 ] Это уменьшение является минимальным для небольших отклонений, но напряжение быстро падает, поскольку длина отклоняется от идеала. Из -за присутствия упругих белков в мышечной клетке (таких как титин ) и внеклеточного матрикса, поскольку мышца растягивается за пределами данной длины, существует совершенно пассивное напряжение, которое противостоит удлинению. В совокупности существует сильная устойчивость к удлинению активной мышцы далеко за пределами пика активного напряжения.
Силовая скорость отношения
[ редактировать ]
Отношения силы и скорости связывают скорость, с которой мышца изменяет свою длину (обычно регулируемой внешними силами, такими как нагрузка или другие мышцы) с количеством силы, которое она генерирует. Сила снижается гиперболической модой по сравнению с изометрической силой, поскольку скорость сокращения увеличивается, в конечном итоге достигая нуля с некоторой максимальной скоростью. Обратная сторона верно для того, когда мышца растягивается - сила увеличивается выше изометрического максимума, пока, наконец, достигнет абсолютного максимума. Это внутреннее свойство активной мышечной ткани играет роль в активном демпфировании суставов, которые приводятся в действие одновременно активными противоположными мышцами. В таких случаях профиль скорости силы усиливает силу, вызванную удлиняющей мышцей за счет сокращения мышц. Это предпочтение любой мышцы возвращает сустав в равновесие эффективно увеличивает демпфирование сустава. Более того, сила демпфирования увеличивается с мышечной силой. Таким образом, моторная система может активно контролировать демпфирование суставов посредством одновременного сокращения (совместно-конструкции) противоположных групп мышц. [ 32 ]
Гладкая мышца
[ редактировать ]
Гладкие мышцы можно разделить на две подгруппы: одноразовые и мультиногические . Клетки гладких мышц одноразового мышца можно найти в кишечнике и кровеносных сосудах. Поскольку эти клетки связаны вместе с помощью щелевых соединений, они способны сокращаться как функциональный синцитик . Клетки гладких мышц одноразового мышца сокращаются миогенно, что может модулироваться вегетативной нервной системой.
В отличие от одноразовых клеток гладких мышц, в мышце глаз и в основании волосяных фолликулов обнаружены мунитные клетки гладких мышц. Клетки гладких мышц с мультиюнитами сокращаются, когда их отдельно стимулируют нервы вегетативной нервной системы. Таким образом, они обеспечивают тонкий контроль и постепенные ответы, очень похожие на рекрутирование двигателя в скелетных мышцах.
Механизмы сокращения гладких мышц
[ редактировать ]
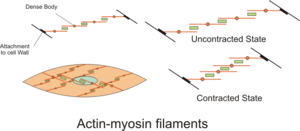
Сократительная активность клеток гладких мышц может быть тонизированным (устойчивым) или фазическим (переходным) [ 33 ] и зависит от нескольких входов, таких как спонтанная электрическая активность, нервные и гормональные входы, локальные изменения в химическом составе и растяжение. [ 1 ] Это в отличие от сократительной активности клеток скелетных мышц, что зависит от одного нейронного ввода. Некоторые типы клеток гладких мышц способны спонтанно генерировать свои собственные потенциалы действия, которые обычно происходят после потенциала кардиостимулятора или потенциала медленного волны . Эти потенциалы действия генерируются притоком внеклеточного CA 2+
, а не на +
Полем Как скелетные мышцы, цитозольное CA 2+
Ионы также необходимы для велосипедного перекрестного моста в клетках гладких мышц.
Два источника цитозольной CA 2+
В клетках гладких мышц находятся внеклеточный CA 2+
вход через кальциевые каналы и CA 2+
ионы, которые высвобождаются из саркоплазматической ретикулумы. Повышение цитозольного CA 2+
приводит к большему количеству CA 2+
Связывание с кальмодулином , который затем связывает и активирует миозиновую легкую цепь киназу . Кальций-кальмодулин-миозиновый комплекс киназы киназы фосфорилирует миозин на миозиновых цепях миозина 20 килодальтона (KDA) на аминокислотных остатков-сирине 19, обеспечивая молекулярное взаимодействие миозина и актина и инициируя контракту и активируя миозин-атпазу . В отличие от клеток скелетных мышц, клетки гладких мышц не имеют тропонина, даже если они содержат тропомиозин белка тонкого нити и другие известные белки - Caldesmon и кальпонин. Таким образом, сокращения гладких мышц инициируются CA 2+
-активированное фосфорилирование миозина, а не CA 2+
Связывание с комплексом тропонина, который регулирует сайты связывания миозина на актина, как в скелетных и сердечных мышцах.
Прекращение велосипедного моста (и оставление мышц в защелке) происходит, когда миозиновая легкая цепь фосфатаза удаляет фосфатные группы из голов миозина. Фосфорилирование световых цепей миозина 20 кДа хорошо коррелирует со скоростью укорочения гладких мышц. В течение этого периода наблюдается быстрый всплеск использования энергии, измеренный с помощью потребления кислорода. В течение нескольких минут после начала уровня кальция заметно уменьшается, фосфорилирование световых цепей миозина на 20 кДа уменьшается, а потребление энергии уменьшается; Однако сила в тонизированных гладких мышцах сохраняется. Во время сокращения мышц быстро циклически образуется перекрестные мощные мосты между активированным актином и фосфорилированным миозином, генерируя силу. Предполагается, что поддержание силы является результатом дефосфорилированных «защелкивающихся мостиков», которые медленно ездят на велосипеде и поддерживают силу. Считается, что ряд киназ, таких как Rho -киназа , DAPK3 и протеинкиназа C, участвуют в устойчивой фазе сокращения и CA 2+
Поток может быть значительным.
Нейромодуляция
[ редактировать ]Хотя сокращения гладких мышц являются миогенными, скорость и сила их сокращений могут модулироваться вегетативной нервной системой . Постганглионовые нервные волокна парасимпатической нервной системы высвобождают ацетилхолин нейротрансмиттер, который связывается с мускариновыми ацетилхолиновыми рецепторами (MACHRS) на клетках гладких мышц. Эти рецепторы являются метаботропными или G-белковыми рецепторами, которые инициируют второй каскад мессенджера. И наоборот, постганглионовые нервные волокна симпатической нервной системы высвобождают нейротрансмиттеры адреналин и норэпинефрин, которые связываются с адренергическими рецепторами, которые также являются метаботропными. Точное влияние на гладкую мышцу зависит от конкретных характеристик активированного рецептора - как парасимпатического ввода, так и симпатического ввода могут быть либо возбуждающими (сократительными), либо ингибирующими (расслабляющими).
Сердечная мышца
[ редактировать ]
Существует два типа клеток сердца мышц : авторитмический и сократительный. Авторитмические клетки не сжимаются, а вместо этого устанавливают темпы сокращения для других клеток сердца мышц, которые могут модулироваться вегетативной нервной системой. Напротив, сократительные мышечные клетки (кардиомиоциты) составляют большую часть сердечной мышцы и способны сокращаться.
Связание с борьбой с возбуждением
[ редактировать ]Как в скелетной, так и в сердечной мышечной связи с взаимодействием (EC), деполяризационная проводимость и CA 2+ Процессы выпуска происходят. Однако, хотя вовлеченные белки схожи, они отличаются по структуре и регуляции. Дигидропиридиновые рецепторы (DHPR) кодируются различными генами, а рецепторы райанодина (RYRS) являются отдельными изоформами. Кроме того, контакты с DHPR с RYR1 (основная изоформа RYR в скелетных мышцах) для регулирования CA 2+ Выброс в скелетных мышцах, в то время как кальциевый канал L-типа (DHPR на миоцитах сердца) и Ryr2 (основная изоформа RYR в сердечной мышце) не физически связаны в сердечной мышце, но сталкиваются друг с другом с помощью соединения. [ 34 ]
В отличие от скелетных мышц, Считается, что сочетание ЕС в сердечной мышце зависит главным образом от механизма, называемого кальцием, индуцированным высвобождением кальция , [ 35 ] который основан на соединительной структуре между Т -булкой и саркоплазматической ретикулумом. Juntophilin-2 (JPH2) необходим для поддержания этой структуры, а также целостности Т-тубулы . [ 36 ] [ 37 ] [ 38 ] Другой белок, рецепторный аксессуарный белок 5 (REEP5), функционирует, чтобы сохранить нормальную морфологию соединения SR. [ 39 ] Дефекты соединительной связи могут быть результатом недостатков любого из двух белков. Во время процесса высвобождения кальция, вызванного кальцием, RYR2 активируются с помощью триггера кальция, который вызван потоком CA 2+ через кальциевые каналы L-типа. После этого сердечная мышца имеет тенденцию демонстрировать диада структуры, а не триады .
Соединение возбуждения в клетках сердца мышц возникает, когда потенциал действия инициируется клетками кардиостимулятора в синоатриальном узле или атриовентрикулярном узле и проводится ко всем клеткам в сердце через разрывные соединения . Потенциал действия перемещается вдоль поверхностной мембраны в Т-тубов (последние не наблюдаются во всех типах сердечных клеток), а деполяризация вызывает внеклеточную CA 2+
Войти в клетку через кальциевые каналы L-типа и, возможно, натриевой кальциум-обменник (NCX) во время ранней части фазы плато . Хотя это Ca 2+ приток соответствует только около 10% CA 2+ Необходимый для активации, он относительно больше, чем у скелетных мышц. Это ок 2+
приток вызывает небольшое локальное увеличение внутриклеточного CA 2+
Полем Увеличение внутриклеточного CA 2+
обнаружен Ryr2 в мембране саркоплазматической ретикулумы, которая выпускает CA 2+
В положительном физиологическом ответе. Эта положительная обратная связь известна как выброс кальция, вызванный кальцием [ 35 ] и приводит к появлению искры кальция ( CA 2+
искры [ 40 ] ) Пространственное и временное суммирование ~ 30 000 CA 2+
Sparks дает общетоковое увеличение цитоплазматической концентрации кальция. [ 41 ] Увеличение цитозольного кальция после потока кальция через клеточную мембрану и саркоплазматический ретикулум смягчается кальциевыми буферами , которые связывают большую долю внутриклеточного кальция. В результате значительное увеличение общего кальция приводит к относительно небольшому росту бесплатного CA 2+
. [ 42 ]
Цитоплазматический кальций связывается с тропонином С, перемещая комплекс тропомиозина из сайта связывания актина, позволяя головке миозина связываться с актиновым филаментом. С этого момента сократительный механизм по существу такой же, как и для скелетных мышц (выше). Вкратце, используя гидролиз АТФ, головка миозина тянет актиновую нить в сторону центра саркории.

После систолы внутриклеточный кальций поднимается накачкой SARCO/Эндоплазматической ретикулумной атпазы (SERCA) обратно в саркоплазматический ретикулум, готовую для начала следующего цикла. Кальций также вытесняется из ячейки в основном кальциевым обменом натрия (NCX) и, в меньшей степени, плазматической мембраной кальциевой атпазой . Некоторый кальций также занимается митохондриями. [ 43 ] Фермент, фосфоламбан , служит тормозом для SERCA. При низкой частоте сердечных сокращений фосфоламбан активен и замедляет активность АТФазы, так что CA 2+
Не должен оставлять ячейку полностью. При высокой частоте сердечных сокращений фосфоламбан фосфорилируется и деактивируется, таким образом, принимая большую часть CA 2+
от цитоплазмы обратно в саркоплазматический ретикулум. Еще раз, буферы кальция умеренно этой осенью в CA 2+
концентрация, разрешающая относительно небольшое снижение бесплатного CA 2+
Концентрация в ответ на большое изменение общего кальция. Падение ок 2+
Концентрация позволяет комплексу тропонина диссоциации от актиновой нити, тем самым заканчивая сокращение. Сердце расслабляется, позволяя желудочкам наполняться кровью и снова начать сердечный цикл.
Беспозвоночные
[ редактировать ]Круглые и продольные мышцы
[ редактировать ]
В таких аннолидах , как дождевые черви и пиявки , клетки круглых и продольных мышц образуют стенку тела этих животных и отвечают за их движение. [ 44 ] Например, в дождевой червях, которая движется через почву, сокращения круговых и продольных мышц возникают взаимно, в то время как целомическая жидкость служит гидроскелетом , поддерживая дивант с дождевой черви. [ 45 ] Когда круглые мышцы в передних сегментах сокращаются, передняя часть тела животного начинает сужать радиально, что толкает несжимаемую целомическую жидкость вперед и увеличивает длину животного. В результате передняя часть животного движется вперед. Когда передний конец дождевого червя становится закрепленным, а круговые мышцы в передних сегментах становятся расслабленными, волна продольных сокращений мышц проходит назад, что тянет остальное затягивающее тело животного вперед. [ 44 ] [ 45 ] Эти чередующиеся волны круговых и продольных сокращений называются перистальтикой , которая лежит в основе ползучего движения дождевых червей.
Наклонно полосатые мышцы
[ редактировать ]Беспозвоночные, такие как Annelids, Mollusks и нематоды , обладают наклонно полосатыми мышцами, которые содержат полосы толстых и тонких филаментов, которые расположены на спиральности, а не поперечно, как в скелетных или сердечных мышцах позвоночных. [ 46 ] В двустворчатых моллюсках мышцы склонной стыки могут поддерживать напряжение в течение длительных периодов, не используя слишком много энергии. Двухцеполисты используют эти мышцы, чтобы держать их раковины закрытыми.
Асинхронные мышцы
[ редактировать ]
У продвинутых насекомых, таких как осы , мухи , пчелы и жуки , обладают асинхронными мышцами , которые составляют мышцы полетов у этих животных. [ 46 ] Эти летные мышцы часто называют фибриллярными мышцами, потому что они содержат миофибрилл, которые являются густыми и заметными. [ 47 ] Замечательной особенностью этих мышц является то, что они не требуют стимуляции для каждого сокращения мышц. Следовательно, они называются асинхронными мышцами , потому что количество сокращений в этих мышцах не соответствует (или синхронизируется) с количеством потенциалов действия. Например, мышца крыла привязанной мухи может получать потенциалы действия на частоте 3 Гц, но она может бить на частоте 120 Гц. [ 46 ] Высокочастотное избиение стало возможным, потому что мышцы связаны с резонансной системой, которая приводится к естественной частоте вибрации.
История
[ редактировать ]
В 1780 году Луиджи Гальвани обнаружил, что мышцы мертвых лягушек держались, когда ударила электрическую искру. [ 49 ] Это был один из первых набег на изучение биоэлектричества , поле, которое по -прежнему изучает электрические паттерны и сигналы в тканях, таких как нервы и мышцы.
В 1952 году была придумана термин -связь возбуждения -контроля для описания физиологического процесса преобразования электрического стимула в механический отклик. [ 50 ] Этот процесс имеет основополагающее значение для физиологии мышц, в результате чего электрический стимул обычно является потенциалом действия, а механический ответ - это сокращение. Связание возбуждения -контроля может быть нарушена при многих заболеваниях. Несмотря на то, что связь возбуждения и контроля была известна более полувека, она все еще является активной областью биомедицинских исследований. Общая схема заключается в том, что потенциал действия приходит для деполяризации клеточной мембраны. С помощью механизмов, специфичных для мышц, эта деполяризация приводит к увеличению цитозольного кальция , который называется переходным процессом кальция. Это увеличение кальция активирует чувствительные к кальция сократительные белки, которые затем используют АТФ, чтобы вызвать укорочение клеток.
Механизм мышечного сокращения в течение многих лет уклонялся от ученых и требует дальнейших исследований и обновления. [ 51 ] Теория скользящих нити была независимо разработана Эндрю Ф. Хаксли и Рольфом Нидергерке, а также Хью Хаксли и Джин Хансон . Их выводы были опубликованы как две последовательные статьи, опубликованные в выпуске природы 22 мая 1954 года под общей темой «Структурные изменения в мышцах во время сокращения». [ 23 ] [ 24 ]
Смотрите также
[ редактировать ]- Анатомические термины движения
- Индуцированное кальцием высвобождение кальция
- Потенциал сердечного действия
- Судороги
- Дистония
- Физиология упражнений
- Фасцикуляция
- Модель мышц Хилла
- Гипновый придурок
- Тестирование мышц in vitro
- Парадокс Ломбарда
- Миоклонус
- СРЕДНАЯ СМЕРТИ
- Спазм
- Сокращение матки
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а Widmaier, Eric P.; Рафф, Херсел; Странг, Кевин Т. (2010). "Мышца". Человеческая физиология Вандера: механизмы функции организма (12 -е изд.). Нью-Йорк, Нью-Йорк: МакГроу-Хилл. С. 250–291. ISBN 978-0-321-98122-6 .
- ^ Silverthorn, Dee Close (2016). "Мышцы" Интегрированный подход (7 -е изд.). Святой Франциск, Калифорния: Пирсон. стр. 377–4 ISBN 978-0-321-98122-6 .
- ^ Jump up to: а беременный БИВЕНЕР, Эндрю А. (2003). «Мышцы и скелеты: строительные блоки движения животных». АВТОМОБИЛЬНОЕ ДОЛЖНОСТЬ . Оксфордская серия биологии животных. Нью -Йорк, Нью -Йорк: издательство Оксфордского университета. С. 15–45. ISBN 978-0-198-50022-3 .
- ^ Jump up to: а беременный в дюймовый и фон Эйдли, Дэвид Дж. (1998). «Механика и энергия мышечного сокращения». Физиология возбудимых клеток (4 -е изд.). Нью -Йорк, Нью -Йорк: издательство Кембриджского университета. С. 323–335 . ISBN 978-0-521-57421-1 .
- ^ Jump up to: а беременный в дюймовый и фон Sircar, Sabyasachi (2008). «Мышечная эластичность». Принципы медицинской физиологии (1 -е изд.). Нью -Йорк, Нью -Йорк: Тим. п. 113. ISBN 978-1-588-90572-7 .
- ^ Jump up to: а беременный в дюймовый и фон Баллок, Джон; Бойл, Джозеф; Ван, Майкл Б. (2001). «Мышечное сокращение». NMS Физиология . Тол. 578 (4 -е изд.). Балтимор, Мэриленд: Липпинкотт Уильямс и Уилкинс. С. 37–56.
- ^ Jump up to: а беременный Кумар, Шраван (2008). «Введение и терминология». В Кумаре, Шраван (ред.). Мышечная сила (1 -е изд.). Boca Raton, FL: CRC Press. п. 113. ISBN 978-0-415-36953-4 .
- ^ Фолкнер, JA (2003). «Терминология для сокращений мышц во время укорочения, в то время как изометрическое и во время удлинения». Журнал прикладной физиологии . 95 (2): 455–459. doi : 10.1152/japplphysiol.00280.2003 . PMID 12851415 . S2CID 28649208 .
- ^ Jump up to: а беременный «Типы сокращений» . 31 мая 2006 г. Получено 2 октября 2007 года .
- ^ Jump up to: а беременный в Коллиандер Э.Б., Тесч П.А. (1990). «Влияние эксцентрических и концентрических мышечных действий в тренировках с сопротивлением». Acta Physiol. Скандал 140 (1): 31–9. doi : 10.1111/j.1748-1716.1990.tb08973.x . PMID 2275403 .
- ^ Nikolaidis MG, Kyparos A, Spanou C, Paschalis V, Theodorou AA, Vrabas IS (2012). «Окислительно -восстановительная биология упражнений: интегративное и сравнительное рассмотрение некоторых упущенных проблем» . J. Exp. Биол . 215 (Pt 10): 1615–25. doi : 10.1242/jeb.067470 . PMID 22539728 .
- ^ Брукс, Джорджия; Фахей, ТД; White, TP (1996). Физиология упражнений: биоэнергетика человека и ее приложения . Mayfield Publishing Co.
- ^ Альфредсон, ч; Pietilä, T; Jonsson, P; Lorentzon, R (1998). «Тренировка эксцентричной мышцы с тяжелой нагрузкой для лечения хронического ахиллового тендиноза» (PDF) . Американский журнал спортивной медицины . 26 (3): 360–6. doi : 10.1177/03635465980260030301 . PMID 9617396 . S2CID 30259362 .
- ^ Satyendra L, BYL N (2006). «Эффективность физической терапии для ахилловой тендинопатии: фактическое обзор эксцентричных упражнений» . Изокинетика и наука о физических упражнениях . 14 (1): 71–80. doi : 10.3233/IES-2006-0223 . Архивировано из оригинала 9 июля 2012 года.
- ^ Cannell LJ, Taunton JE, Clement DB, Smith C, Khan KM (2001). «Рандомизированное клиническое исследование эффективности приседаний капель или упражнений по скручиванию ног/сгибания ног для лечения клинически диагностированного колена перемычки у спортсменов: пилотное исследование» . Br J Sports Med . 35 (1): 60–4. doi : 10.1136/bjsm.35.1.60 . PMC 1724276 . PMID 11157465 .
- ^ Тассинар; Cacioppo (2000). «Скелемоторная система: поверхностная электромиография». В Cacioppo, Джон Т.; Тасинар, Луа Г.; Бернтсон, Гэри Г. (ред.). Справочник по психофизиологии (второе изд.). Кембридж: издательство Кембриджского университета. ISBN 978-0-521-62634-7 .
- ^ Левитан, Ирвин; Качмарек, Леонард (19 августа 2015 г.). «Межклеточная связь». Нейрон: клеточная и молекулярная биология (4 -е изд.). Нью -Йорк, Нью -Йорк: издательство Оксфордского университета. С. 153–328. ISBN 978-0199773893 .
- ^ Jump up to: а беременный Саладин, Кеннет С., Стивен Дж. Салливан и Кристина А. Ган. Анатомия и физиология: единство формы и функции. 7 -е изд. Нью-Йорк: McGraw-Hill Education, 2015. Печать.
- ^ Jump up to: а беременный Lanner, Johanna T.; Georgiou, Dimitra K.; Джоши, Адитья Д.; Гамильтон, Сьюзен Л. (ноябрь 2010). «Райанодиновые рецепторы: структура, экспрессия, молекулярные детали и функция в высвобождении кальция» . Перспективы Cold Spring Harbor в биологии . 2 (11): A003996. doi : 10.1101/cshperspect.a003996 . ISSN 1943-0264 . PMC 2964179 . PMID 20961976 .
- ^ Calderón, Juan C.; Боланьос, Пура; Капуто, Карло (март 2014 г.). «Механизм связывания возбуждения в скелетных мышцах» . Биофизические обзоры . 6 (1): 133–160. doi : 10.1007/s12551-013-0135-x . ISSN 1867-2450 . PMC 5425715 . PMID 28509964 .
- ^ Уэбб, Р. Клинтон (декабрь 2003 г.). «Кравление гладких мышц и расслабление» . Достижения в области физиологического образования . 27 (4): 201–206. doi : 10.1152/advan.00025.2003 . ISSN 1043-4046 .
- ^ Саладин, Кеннет (2012). Анатомия и физиология: единство формы и функции . Нью -Йорк: МакГроу Хилл. ISBN 978-0-07-337825-1 .
- ^ Jump up to: а беременный Huxley AF, Niedergerke R (1954). «Структурные изменения в мышцах во время сокращения: интерференционная микроскопия живых мышечных волокон». Природа . 173 (4412): 971–973. Bibcode : 1954natur.173..971H . doi : 10.1038/173971a0 . PMID 13165697 . S2CID 4275495 .
- ^ Jump up to: а беременный Хаксли Х., Хансон Дж. (1954). «Изменения в перекрестной полосе мышц во время сокращения и растяжения и их структурной интерпретации». Природа . 173 (4412): 973–976. Bibcode : 1954natur.173..973H . doi : 10.1038/173973a0 . PMID 13165698 . S2CID 4180166 .
- ^ Horowits R, Podolsky RJ (ноябрь 1987 г.). «Позиционная стабильность толстых филаментов в активированных скелетных мышцах зависит от длины саркомера: доказательства роли титина» . J. Cell Biol . 105 (5): 2217–23. doi : 10.1083/jcb.105.5.2217 . PMC 2114850 . PMID 3680378 .
- ^ Jump up to: а беременный в Enoka, Roger M.; Пирсон, Кейр Г. (2013). «Моторная единица и мышечное действие». В Канделе, Эрик Р.; Шварц, Джеймс Х.; Джесселл, Томас М.; Зигельбаум, Стивен А.; Hudspeth, AJ (Eds.). Принципы нейронной науки (5 -е изд.). Нью-Йорк, Нью-Йорк: McGraw-Hill Medical. С. 768–789. ISBN 978-0-071-39011-8 .
- ^ Фехер, Джозеф (2012). «Глава 3.4: Механика скелетных мышц». Количественная физиология человека: введение . Академическая пресс -серия в биомедицинской инженерии (1 -е изд.). Нью -Йорк, Нью -Йорк: Академическая пресса. С. 239–248. ISBN 978-0-123-82163-8 .
- ^ Хурана, Инду (2006). «Характеристики мышечной возбудимости и сократимости». Учебник медицинской физиологии (1 -е изд.). Elsevier. С. 101–2.
- ^ Смит, Ян С.; Адам, Хелен; Герцог, Уолтер (апрель 2020 г.). «Краткое сокращение оказывает сложное влияние на суммирование паров Twitch в adductor volicis» . Экспериментальная физиология . 105 (4): 676–689. doi : 10.1113/ep088401 . PMID 32052487 . S2CID 211100581 . Получено 5 апреля 2022 года .
- ^ Shwedyk, E.; Balasubramanian, R.; Скотт, Р.Н. (1977). «Нестационарная модель для электромиограммы». IEEE транзакции по биомедицинской инженерии . 24 (5): 417–424. doi : 10.1109/tbme.1977.326175 . PMID 892834 . S2CID 1770255 .
- ^ Гордон А.М., Хаксли А.Ф., Джулиан Ф.Дж. (1966). «Изменение изометрического напряжения с длиной саркомера в мышечных волокнах позвоночных» . J. Physiol . 184 (1): 170–92. doi : 10.1113/jphysiol.1966.sp007909 . PMC 1357553 . PMID 5921536 .
- ^ Хейтманн, Стюарт; Папоротники, норм; Breakpsear, Michael (2011). «Мышечная совместная связь модулирует демпфирование и стабильность сустава в биомеханической конечности из трех связей» . Границы в нейророботике . 5 : 5. doi : 10.3389/fnbot.2011.00005 . ISSN 1662-5218 . PMC 3257849 . PMID 22275897 .
- ^ Чжан, y; Германсон, я; Eddinger, TJ (2013). «Тонизированное и фазовое сокращение гладких мышц не регулируется путем PKCα - CPI -17 в антруме и глазного дна свиней и глазного дна» . Plos один . 8 (9): E74608. BIBCODE : 2013PLOSO ... 874608Z . doi : 10.1371/journal.pone.0074608 . PMC 3776813 . PMID 24058600 .
- ^ Martonosi, Энтони Н.; Пикула, Славомир (2003). «Сеть регуляции кальция в мышцах» . Acta Biochimica Polonica . 50 (1): 1–30. doi : 10.18388/abp.2003_3711 . ISSN 0001-527X . PMID 12673344 .
- ^ Jump up to: а беременный Фабиато А. (1983). «Индуцированное кальцием высвобождение кальция из сердечной саркоплазматической ретикулумы». Американский журнал физиологии . 245 (1): C1–14. doi : 10.1152/ajpcell.1983.245.1.c1 . PMID 6346892 .
- ^ Го, Анг; Чжан, Xiaoying; Айер, Венкат Рамеш; Чен, Бийя; Чжан, Каймеи; Кутшке, Уильям Дж.; Вайс, Роберт М.; Franzini-Armstrong, Clara; Песня, Лонг-Шен (19 августа 2014 г.). «Сверхэкспрессия Juntophilin-2 не усиливает базовую функцию, но ослабляет развитие сердечной недостаточности после сердечного стресса» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (33): 12240–12245. BIBCODE : 2014PNAS..11112240G . doi : 10.1073/pnas.1412729111 . ISSN 1091-6490 . PMC 4143026 . PMID 25092313 .
- ^ Вэй, Шэн; Го, Анг; Чен, Бийя; Кутшке, Уильям; Xie, Yu-Ping; Циммерман, Кэти; Вайс, Роберт М.; Андерсон, Марк Э.; Ченг, Хепинг; Песня, Лонг-Шен (20 августа 2010 г.). «Ремоделирование T-Tubule во время перехода от гипертрофии к сердечной недостаточности» . Исследование циркуляции . 107 (4): 520–531. doi : 10.1161/circresaha.109.212324 . ISSN 1524-4571 . PMC 2927862 . PMID 20576937 .
- ^ Takehima, H.; Komazaki, S.; Nishi, M.; Iino, M.; Кангава, К. (июль 2000 г.). «Джанкнофилины: новое семейство белков соединительного мембранного комплекса» . Молекулярная клетка . 6 (1): 11–22. doi : 10.1016/s1097-2765 (00) 00003-4 . ISSN 1097-2765 . PMID 10949023 .
- ^ Яо, Лей; Xie, Duanyang; Генг, Ли; Ши, Дэн; Хуан, Цзянь; Ву, Юфэй; Lv, fei; Лян, Дандан; Ли, Ли; Лю, Йи; Ли, июнь (3 февраля 2018 г.). «REEP5 (рецепторный аксессуарный белок 5) действует как саркоплазматический скульптор ретикулума для модуляции сердечной функции» . Журнал Американской кардиологической ассоциации . 7 (3). doi : 10.1161/jaha.117.007205 . ISSN 2047-9980 . PMC 5850239 . PMID 29431104 .
- ^ Cheng H, Lederer WJ, Cannell MB (октябрь 1993 г.). «Sparks кальция: элементарные события, лежащие в основе соединения возбуждения в сердечных мышцах». Наука . 262 (5134): 740–4. Bibcode : 1993sci ... 262..740c . doi : 10.1126/science.8235594 . PMID 8235594 .
- ^ Cannell MB, Cheng H, Lederer WJ (ноябрь 1994 г.). "Пространственные неравномерность в CA 2+
I во время связания с контролем возбуждения в миоцитах сердца » . Биофиз. J. 67 ( 5): 1942–56. : 1994bpj .... 67.1942c . DOI : 10.1016/S0006-3495 (94) 80677-0 . Bibcode . PMC 1255569/S0006-3495 (94) 80677-0 . - ^ Bers, MD (2001). Связание с контролем возбуждения и сократительная сила сердца (2-е изд.). Дордрехт: академические издатели Kluwer. ISBN 9780792371571 Полем OCLC 47659382 .
- ^ Креспо Л.М., Грантхэм К.Дж., Каннелл М.Б. (июнь 1990 г.). «Кинетика, стехиометрия и роль обменного механизма Na-CA в изолированных миоцитах сердца». Природа . 345 (6276): 618–21. Bibcode : 1990natur.345..618c . doi : 10.1038/345618A0 . PMID 2348872 . S2CID 4348240 .
- ^ Jump up to: а беременный Хиллис, Дэвид М.; Садава, Дэвид Э.; Прайс, Мэри В. (2014). «Мышцы и движение». Принципы жизни (2 -е изд.). Сандерленд, Массачусетс: Sinauer Associates. С. 681–698. ISBN 978-1-464-10947-8 .
- ^ Jump up to: а беременный Гарднер, Кр (1976). «Нейрональный контроль передвижения в дождевой червях». Биологические обзоры Кембриджского философского общества . 51 (1): 25–52. doi : 10.1111/j.1469-185x.1976.tb01119.x . PMID 766843 . S2CID 9983649 .
- ^ Jump up to: а беременный в Александр, Р. Макнил (2003). «Мышца, мотор». Принципы передвижения животных (2 -е изд.). Принстон, Нью -Джерси: издательство Принстонского университета. С. 15–37. ISBN 978-0-691-12634-0 .
- ^ Джозефсон, RK; Маламуд, JG; Стоукс, DR (15 сентября 2000 г.). «Асинхронная мышца: праймер» . Журнал экспериментальной биологии . 203 (18): 2713–2722. doi : 10.1242/jeb.203.18.2713 . ISSN 0022-0949 . PMID 10952872 .
- ^ Уэллс, Дэвид Эймс (1859). «Как было обнаружено гальваническое электричество» . Наука общих вещей: знакомое объяснение первых принципов физической науки . Нью -Йорк: Ивисон и Финни. п. 290.
- ^ Whittaker, et (1951), история теорий эфира и электричества. Том 1 , Нельсон, Лондон
- ^ Сандоу, А (1952). «Соединение по борьбе с возбуждением в мышечном ответе» . Yale J Biol Med . 25 (3): 176–201. PMC 2599245 . PMID 13015950 .
- ^ Хаксли, он (апрель 2000 г.). «Прошлые, настоящие и будущие эксперименты на мышцах» . Философские транзакции: биологические науки . 355 (1396): 539–543. doi : 10.1098/rstb.2000.0595 . JSTOR 3066716 . PMC 1692762 . PMID 10836507 .
Дальнейшее чтение
[ редактировать ]- Krans, JL (2010) Теория скользящей нити о сжатии мышц. Nature Education 3 (9): 66
- Саладин, Кеннет С., Стивен Дж. Салливан и Кристина А. Ган. (2015). Анатомия и физиология: единство формы и функции. 7 -е изд. Нью-Йорк: McGraw-Hill Education.