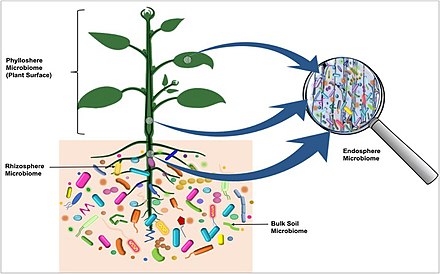
Растительный микробиом
| Часть серии о |
| Микробиомы |
|---|
 |
, Растительный микробиом также известный как фитомикробиом , играет роль в здоровье и продуктивности растений, и в последние годы ему уделяется значительное внимание. [ 1 ] [ 2 ] Микробиом . был определен как «характерное микробное сообщество, занимающее достаточно четко определенную среду обитания и обладающее отчетливыми физико-химическими свойствами. Таким образом, этот термин не только относится к вовлеченным микроорганизмам, но также охватывает и область их активности» [ 3 ] [ 4 ]
Растения живут в ассоциации с разнообразными микробными консорциумами . Эти микробы, называемые микробиотой растения , живут как внутри (эндосфера), так и снаружи (эписфера) тканей растения и играют важную роль в экологии и физиологии растений. [ 5 ] «Считается, что основной микробиом растения состоит из ключевых микробных таксонов, которые важны для приспособленности растений и созданы посредством эволюционных механизмов отбора и обогащения микробных таксонов, содержащих гены, важные функции для приспособленности растительного холобионта». [ 6 ]
Микробиомы растений формируются как факторами, связанными с самим растением, такими как генотип, орган, вид и состояние здоровья, так и факторами, связанными с окружающей средой растения, такими как управление, землепользование и климат. [ 7 ] В некоторых исследованиях сообщалось, что состояние здоровья растения отражается его микробиомом или связано с ним. [ 8 ] [ 1 ] [ 9 ] [ 2 ]
Обзор
[ редактировать ]
Изучение связи растений с микроорганизмами предшествует изучению микробиомов животных и человека, особенно роли микробов в поглощении азота и фосфора. Наиболее яркими примерами являются растений корненно - арбускулярная микоризная (АМ) и бобово-ризобиальный симбиозы , оба из которых сильно влияют на способность корней поглощать различные питательные вещества из почвы. Некоторые из этих микробов не могут выжить в отсутствие растения-хозяина ( к облигатным симбионтам относятся вирусы, а также некоторые бактерии и грибы), который обеспечивает микроорганизмы пространством, кислородом, белками и углеводами. Связь АМ-грибов с растениями известна с 1842 г., с ними связано более 80% наземных растений. [ 11 ] Считается, что AM-грибы помогли одомашнить растения. [ 5 ]

Традиционно исследования взаимодействия растений и микробов ограничивались культивируемыми микробами . Многочисленные микробы, которые невозможно было культивировать, остались неисследованными, поэтому знания об их роли в значительной степени неизвестны. [ 5 ] Возможности раскрытия типов и результатов этих взаимодействий растений и микробов вызвали значительный интерес среди экологов, биологов-эволюционистов, биологов-растений и агрономов. [ 8 ] [ 12 ] [ 1 ] Недавние разработки в области мультиомики и создание больших коллекций микроорганизмов резко расширили знания о составе и разнообразии микробиома растений. Секвенирование , маркерных генов целых микробных сообществ, называемое метагеномикой проливает свет на филогенетическое разнообразие микробиомов растений. Это также дополняет знания об основных биотических и абиотических факторах, микробиомов растений ответственных за формирование сообществ . [ 12 ] [ 5 ]
Состав микробных сообществ, связанных с разными видами растений, коррелирует с филогенетической дистанцией между видами растений, то есть близкородственные виды растений имеют тенденцию иметь более сходные микробные сообщества, чем отдаленные виды. [ 13 ] Основное внимание в исследованиях микробиома растений было уделено модельным растениям, таким как Arabidopsis thaliana , а также важным экономическим видам культур, включая ячмень (Hordeum vulgare), кукурузу (Zea mays), рис (Oryza sativa), сою (Glycine max), пшеница (Triticum aestivum), тогда как фруктовым культурам и древесным породам уделяется меньше внимания. [ 14 ] [ 2 ]
Микробиота растений
[ редактировать ]Цианобактерии являются примером микроорганизмов, которые широко симбиотически взаимодействуют с наземными растениями . [ 15 ] [ 16 ] [ 17 ] [ 18 ] Цианобактерии могут проникать в растение через устьица и колонизировать межклеточное пространство, образуя петли и внутриклеточные клубки. [ 19 ] Анабаена виды. колонизировать корни растений пшеницы и хлопчатника. [ 20 ] [ 21 ] [ 22 ] Калотрикс сп. также был обнаружен в корневой системе пшеницы. [ 21 ] [ 22 ] Однодольные растения , такие как пшеница и рис, были заселены видами Nostoc . [ 23 ] [ 24 ] [ 25 ] [ 26 ] В 1991 году Гантер и другие выделили из корней растений и почвы разнообразные гетероцистные азотфиксирующие цианобактерии, в том числе Nostoc , Anabaena и Cylindrospermum . Оценка корней проростков пшеницы выявила два типа паттернов ассоциации: свободную колонизацию корневых волосков Anabaena и плотную колонизацию поверхности корня в ограниченной зоне Nostoc . [ 23 ] [ 27 ]


(2) На поверхности корня цианобактерии демонстрируют два типа колонизации; в корневых волосках нити видов Anabaena и Nostoc образуют рыхлые колонии, а в ограниченной зоне на поверхности корня специфические виды Nostoc образуют колонии цианобактерий.
(3) Совместная инокуляция 2,4-D и Nostoc spp. увеличивает образование параклубеньков и фиксацию азота. Большое количество Nostoc spp. изоляты колонизируют корневую эндосферу и образуют параклубеньки. [ 27 ]
Ризосферный микробиом
[ редактировать ]
микробы и корневые выделения [ 29 ]
Ризосфера корни включает зону почвы толщиной 1–10 мм, непосредственно окружающую , которая находится под влиянием растения за счет отложения корневых экссудатов , слизи и мертвых растительных клеток. [ 30 ] В ризосфере живут разнообразные организмы, включая бактерии , грибы , оомицеты , нематоды , водоросли , простейшие , вирусы и археи . [ 31 ]
«Экспериментальные данные подчеркивают важность корневого микробиома для здоровья растений, и становится все более очевидным, что растение способно контролировать состав своего микробиома. их репродуктивному успеху будет способствовать эволюционный отбор. Похоже, что такое давление отбора привело к множеству специфических взаимодействий между растениями и микробами, и накапливаются доказательства того, что растения требуют микробной помощи в случае необходимости».
– Берендсен и др., 2012 г. [ 8 ]

на корнях Arabidopsis thaliana
а) Общий вид корня A. thaliana (первичный корень) с многочисленными корневыми волосками. б) Бактерии , образующие биопленки . в) грибов или оомицетов, Гифы окружающие поверхность корня. г) Первичный корень густо покрыт спорами и протистами . д, е) Протисты , скорее всего, относящиеся к классу Bacillariophyceae . г) Бактерии и бактериальные нити . з, и) Различные бактериальные особи, демонстрирующие большое разнообразие форм и морфологических особенностей. [ 32 ]
Микоризные грибы являются многочисленными представителями ризосферного сообщества, они были обнаружены у более чем 200 000 видов растений и, по оценкам, ассоциированы с более чем 80% всех растений. [ 33 ] Микориза-корневые ассоциации играют важную роль в наземных экосистемах, регулируя циклы питательных веществ и углерода . Микоризы являются неотъемлемой частью здоровья растений, поскольку они обеспечивают до 80% потребностей растений в азоте и фосфоре. Взамен грибы получают углеводы и липиды от растений-хозяев. [ 34 ] Недавние исследования арбускулярных микоризных грибов с использованием технологий секвенирования показывают большее межвидовое и внутривидовое разнообразие, чем было известно ранее. [ 35 ] [ 5 ]
Наиболее часто изучаемыми полезными ризосферными организмами являются микоризы , бактерии-ризобии , ризобактерии, способствующие росту растений (PGPR), и микробы биоконтроля . Предполагалось, что один грамм почвы может содержать более миллиона различных бактериальных геномов. [ 36 ] и более 50 000 OTU ( оперативных таксономических единиц ) были обнаружены в ризосфере картофеля. [ 37 ] Среди прокариотов в ризосфере наиболее часто встречаются бактерии Acidobacteriota , Pseudomonadota , Planctomycetota , Actinomycetota , Bacteroidota и Bacillota . [ 38 ] [ 39 ] В некоторых исследованиях не было обнаружено существенных различий в составе микробного сообщества между основной почвой (почвой, не прикрепленной к корню растения) и ризосферной почвой. [ 40 ] [ 41 ] Определенные группы бактерий (например, Actinomycetota, Xanthomonadaceae ) менее распространены в ризосфере, чем в близлежащей основной массе почвы. [ 38 ] [ 5 ]
Эндосферный микробиом
[ редактировать ]Некоторые микроорганизмы, например эндофиты , проникают и занимают внутренние ткани растений, образуя эндосферный микробиом . Арбускулярные микоризные и другие эндофитные грибы являются доминирующими колонизаторами эндосферы. [ 42 ] Бактерии и в некоторой степени археи являются важными членами сообществ эндосферы. Некоторые из этих эндофитных микробов взаимодействуют со своим хозяином и приносят очевидную пользу растениям. [ 38 ] [ 43 ] [ 44 ] В отличие от ризосферы и ризопланы, эндосферы содержат весьма специфические микробные сообщества. Корневое эндофитное сообщество может сильно отличаться от сообщества прилегающей почвы. В целом разнообразие эндофитного сообщества ниже разнообразия микробного сообщества вне растения. [ 41 ] Идентичность и разнообразие эндофитного микробиома надземных и подземных тканей также могут различаться внутри растения. [ 45 ] [ 42 ] [ 5 ]
Филлосферный микробиом
[ редактировать ]
Воздушная поверхность растения (стебель, лист, цветок, плод) называется филлосферой и считается сравнительно бедной питательными веществами по сравнению с ризосферой и эндосферой. Среда филлосферы более динамична, чем среда ризосферы и эндосферы. Микробные колонизаторы подвержены суточным и сезонным колебаниям тепла, влаги и радиации. Кроме того, эти элементы окружающей среды влияют на физиологию растений (например, фотосинтез, дыхание, поглощение воды и т. д.) и косвенно влияют на состав микробиома. [ 5 ] Дождь и ветер также вызывают временные изменения микробиома филлосферы. [ 47 ]
Взаимодействия между растениями и связанными с ними микроорганизмами во многих из этих микробиомов могут играть ключевую роль в растений-хозяев . здоровье, функционировании и эволюции [ 48 ] Поверхность листа, или филлосфера , содержит микробиом, состоящий из разнообразных сообществ бактерий, грибов, водорослей, архей и вирусов. [ 49 ] [ 50 ] Взаимодействия между растением-хозяином и филлосферными бактериями могут влиять на различные аспекты физиологии растения-хозяина. [ 51 ] [ 52 ] [ 53 ] Однако по состоянию на 2020 год знания об этих бактериальных ассоциациях в филлосфере остаются относительно скромными, и существует необходимость в расширении фундаментальных знаний о динамике микробиома филлосферы. [ 54 ] [ 55 ]
В целом сохраняется высокое видовое богатство филлосферных сообществ. Грибные сообщества весьма изменчивы в филлосфере умеренных регионов и более разнообразны, чем в тропических регионах. [ 56 ] На поверхности листьев растений может присутствовать до 107 микробов на квадратный сантиметр, а бактериальная популяция филлосферы в глобальном масштабе оценивается в 10 26 клетки. [ 57 ] Размер популяции грибной филлосферы, вероятно, будет меньше. [ 58 ]
Микробы филлосферы разных растений кажутся в некоторой степени сходными у таксонов высоких уровней, но у таксонов более низких уровней сохраняются существенные различия. Это указывает на то, что микроорганизмам может потребоваться тонко настроенная метаболическая корректировка, чтобы выжить в среде филлосферы. [ 56 ] Pseudomonadota , по-видимому, являются доминирующими колонизаторами, при этом Bacteroidota и Actinomycetota также преобладают в филлосферах. [ 59 ] обнаружено очень мало сходства . Хотя между ризосферными и почвенными микробными сообществами существуют сходства, между филлосферными сообществами и микроорганизмами, плавающими на открытом воздухе ( аэропланктоном ), [ 42 ] [ 5 ]
Сборка микробиома филлосферы, которую можно строго определить как эпифитные бактериальные сообщества на поверхности листьев, может формироваться микробными сообществами, присутствующими в окружающей среде (т. е. стохастическая колонизация ) и растении-хозяине (т. е. биотический отбор). [ 49 ] [ 57 ] [ 55 ] Однако, хотя поверхность листьев обычно считается дискретной средой обитания микробов, [ 60 ] [ 61 ] нет единого мнения относительно доминирующего фактора формирования сообществ в микробиомах филлосферы. Например, в филлосфере одновременно встречающихся видов растений были зарегистрированы бактериальные сообщества, специфичные для хозяина, что указывает на доминирующую роль отбора хозяина. [ 61 ] [ 42 ] [ 62 ] [ 55 ]
И наоборот, сообщается, что микробиомы окружающей среды являются основным определяющим фактором состава сообщества филлосферы. [ 60 ] [ 63 ] [ 56 ] [ 64 ] В результате процессы, которые управляют сборкой филлосферных сообществ, недостаточно изучены, но вряд ли они будут универсальными для всех видов растений. Однако существующие данные указывают на то, что филлосферные микробиомы, демонстрирующие ассоциации с хозяином, с большей вероятностью будут взаимодействовать с хозяином, чем те, которые в первую очередь рекрутируются из окружающей среды. [ 51 ] [ 65 ] [ 66 ] [ 67 ] [ 55 ]
Поиск основного микробиома в микробных сообществах, связанных с хозяином, является полезным первым шагом в попытке понять взаимодействия, которые могут происходить между хозяином и его микробиомом. [ 68 ] [ 69 ] Преобладающая концепция основного микробиома основана на представлении о том, что выживание таксона за пространственно-временными границами экологической ниши напрямую отражает его функциональную значимость в той нише, которую он занимает; следовательно, он обеспечивает основу для идентификации функционально важных микроорганизмов, которые постоянно ассоциируются с видом-хозяином. [ 68 ] [ 70 ] [ 40 ] [ 55 ]
В научной литературе возникли различные определения «основного микробиома», при этом исследователи по-разному определяют «основные таксоны» как те, которые сохраняются в различных микросредах обитания хозяина. [ 71 ] [ 72 ] и даже разные виды. [ 62 ] [ 65 ] Учитывая функциональные различия микроорганизмов у разных видов хозяев. [ 62 ] и микросреды обитания, [ 73 ] определение основных таксонов в строгом смысле слова как таксонов, устойчивых на больших географических расстояниях в ткане- и видоспецифичных микробиомах хозяина, представляет собой наиболее биологически и экологически приемлемое применение этой концептуальной основы. [ 74 ] [ 55 ] О ткане- и видоспецифичных основных микробиомах в популяциях хозяев, разделенных большими географическими расстояниями, в филлосфере не сообщалось широко, если использовать строгое определение, установленное Руиненом. [ 52 ] [ 55 ]
Пример: филлосфера манука.
[ редактировать ]Цветущее чайное дерево, широко известное как манука , произрастает в Новой Зеландии. [ 75 ] Мед манука , получаемый из нектара цветов манука, известен своими неперекисными антибактериальными свойствами. [ 76 ] [ 77 ] Микроорганизмы изучались в ризосфере и эндосфере манука. [ 78 ] [ 79 ] [ 80 ] Более ранние исследования в основном были сосредоточены на грибах, а исследование 2016 года представило первое исследование эндофитных бактериальных сообществ из трех географически и экологически различных популяций манука с использованием методов дактилоскопии и выявило тканеспецифичные основные эндомикробиомы. [ 81 ] [ 55 ]

(B) На диаграмме справа показано, как OTU в филлосфере и связанных с ней почвенных сообществах различались по относительной численности. [ 55 ]
Исследование 2020 года выявило специфичный для среды обитания и относительно многочисленный основной микробиом в филлосфере манука, который сохранялся во всех образцах. Напротив, неосновные микроорганизмы филлосферы демонстрировали значительные различия между отдельными деревьями-хозяевами и популяциями, что в значительной степени было обусловлено экологическими и пространственными факторами. Результаты продемонстрировали существование доминирующего и повсеместного основного микробиома в филлосфере мануки. [ 55 ]


Семенной микробиом
[ редактировать ]Семена растений могут служить естественными переносчиками вертикальной передачи полезных эндофитов, например тех, которые придают устойчивость к болезням. В исследовательской статье 2021 года объясняется: «Имеет смысл, что их наиболее важные симбионты будут передаваться вертикально через семена, а не делать ставку на то, что все правильные микробы, обитающие в почве, могут быть доступны в месте прорастания». [ 82 ]
Новая парадигма относительно мутуалистических грибов и передачи бактерий через семена растений-хозяев возникла в основном благодаря исследованиям, касающимся растений, имеющих сельскохозяйственное значение. [ 82 ] [ 83 ] Было обнаружено, что семена риса характеризуются высоким микробным разнообразием, причем наибольшее разнообразие обитает в зародыше, а не в околоплоднике . [ 84 ] Было обнаружено, что грибы рода Fusarium, передающиеся через семена, являются доминирующими членами микробиома стеблей кукурузы . [ 82 ] Этот аспект микробиома растения стал известен как микробиом семян. [ 85 ]
Исследователи лесного хозяйства также начали выявлять членов микробиома семян, принадлежащих к ценным породам деревьев. Вертикальная передача грибковых и бактериальных мутуалистов подтверждена в 2021 году желудях дубов . на [ 45 ] [ 86 ] Если окажется, что исследования дубов применимы и к другим породам деревьев, то станет ясно, что надпочвенные части растения ( филлосфера ) получают почти все свои полезные грибы из тех, которые содержатся в семенах. [ 45 ] Напротив, корни ( ризосфера ) приобретают из семени лишь небольшую часть своих мутуалистов. Большинство из них попадает через окружающую почву, включая их жизненно важные связи с арбускулярными микоризными грибами . [ 83 ]
Виды микробов, постоянно обнаруживаемые в семенах растений, известны как «основной микробиом». [ 82 ] [ 87 ] Преимущества для растения-хозяина включают их способность способствовать выработке противомикробных соединений, детоксикации, поглощению питательных веществ и стимулированию роста. [ 45 ] Выявление функций симбиотических микробов в семенах отодвигает сельскохозяйственную парадигму от селекции и подготовки семян, которые традиционно стремились свести к минимуму присутствие грибковых и бактериальных размножений. Вероятность того, что микроб, обнаруженный в семени, является мутуалистическим, теперь является обычным предположением. Такие партнеры могут способствовать «покою и прорастанию семян, адаптации к окружающей среде, устойчивости и толерантности к болезням, а также стимулированию роста». [ 87 ]
Было предложено применить новое понимание полезных микробов, населяющих семена, не только в сельском хозяйстве, но и для сохранения биоразнообразия. [ 83 ] Группа граждан, выступающая за содействие миграции на север исчезающего дерева в США, указала на изменение парадигмы семенного микробиома как на причину, по которой официальные учреждения снимают запрет на передачу семян за пределы ex situ насаждений, предназначенных для сохранения в северной Джорджии. [ 88 ]
Растение холобионт
[ редактировать ]С момента колонизации земли предковыми линиями растений 450 миллионов лет назад растения и связанные с ними микробы взаимодействовали друг с другом, образуя совокупность видов, которую часто называют холобионтом . Селективное давление, действующее на компоненты холобионта, вероятно, сформировало микробные сообщества, связанные с растениями, и отобрало адаптированные к хозяину микроорганизмы, влияющие на приспособленность растений. Однако высокая плотность микробов, обнаруженная в тканях растений, вместе с быстрым временем образования микробов и их более древним происхождением по сравнению с их хозяином, позволяют предположить, что межмикробные взаимодействия также являются важными селективными силами, формирующими сложные микробные комплексы в филлосфере , ризосфере , и растительные отсеки эндосферы . [ 32 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с Тернер, Томас Р.; Джеймс, Юан К.; Пул, Филип С. (2013). «Микробиом растений» . Геномная биология . 14 (6): 209. doi : 10.1186/gb-2013-14-6-209 . ПМК 3706808 . ПМИД 23805896 .
- ^ Jump up to: а б с Пурахонг, Витун; Орру, Луиджи; Донати, Ирен; Перпетуини, Джорджия; Челлини, Антонио; Ламонтанара, Антонелла; Микелотти, Ваня; Таккони, Джанни; Спинелли, Франческо (2018). «Микробиом растений и его связь со здоровьем растений: виды-хозяева, органы и Pseudomonas syringae pv. Инфекция Actinidiae, формирующая бактериальные филлосферные сообщества растений киви» . Границы в науке о растениях . 9 : 1563. doi : 10.3389/fpls.2018.01563 . ПМК 6234494 . ПМИД 30464766 . .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Уиппс, Дж. М.; Льюис, Карен; Кук, Р.К. (1988). «Микопаразитизм и борьба с болезнями растений». В Бердже, Майкл Н. (ред.). Грибы в системах биологического контроля . Манчестер, Великобритания, Нью-Йорк: Издательство Манчестерского университета. стр. 161–187. ISBN 978-0-7190-1979-1 .
- ^ Берг, Габриэле ; Рыбакова Дарья; Фишер, Дорин; Чернава, Томислав; Верже, Мари-Кристин Шампомье; Чарльз, Тревор; Чен, Сяоюлун; Коколин, Лука; Эверсол, Келли; Коррал, Хема Эрреро; Казу, Мария; Кинкель, Линда; Ланге, Лене; Лима, Нельсон; Лой, Александр; и др. (декабрь 2020 г.). «Повторное рассмотрение определения микробиома: старые концепции и новые проблемы» . Микробиом . 8 (1). дои : 10.1186/s40168-020-00875-0 . hdl : 1822/65842 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Jump up to: а б с д и ж г час я дж Дастогер, К.М., Тумпа, Ф.Х., Султана, А., Актер, М.А. и Чакраборти, А. (2020) «Микробиом растений – отчет о факторах, которые формируют состав и разнообразие сообщества». Текущая биология растений : 100161. дои : 10.1016/j.cpb.2020.100161 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Компант, Стефан; Самад, Абдул; Фаист, Ханна; Сессич, Анжела (2019). «Обзор микробиома растений: экология, функции и новые тенденции в применении микробов» . Журнал перспективных исследований . 19 :29–37. дои : 10.1016/j.jare.2019.03.004 . ПМК 6630030 .
- ^ Брингель, Франсуаза; Куэ, Иван (2015). «Основная роль микроорганизмов филлосферы на стыке между функционированием растений и динамикой газовых примесей в атмосфере» . Границы микробиологии . 06 : 486. дои : 10.3389/fmicb.2015.00486 . ПМК 4440916 . ПМИД 26052316 .
- ^ Jump up to: а б с Берендсен, Роланд Л.; Питерс, Корне MJ; Баккер, Питер АХМ (2012). «Ризосферный микробиом и здоровье растений». Тенденции в науке о растениях . 17 (8): 478–486. doi : 10.1016/j.tplants.2012.04.001 . hdl : 1874/255269 . ПМИД 22564542 . S2CID 32900768 .
- ^ Берг, Габриэле; Грубе, М.; Шлотер, М.; Смолла, К. (2014). «Микробиом растений и его значение для здоровья растений и человека» . Границы микробиологии . 5 : 491. дои : 10.3389/fmicb.2014.00491 . ПМК 4166366 . ПМИД 25278934 .
- ^ Шелаке, Р.М., Праманик, Д. и Ким, Дж.Я. (2019) «Исследование взаимодействия растений и микробов для устойчивого сельского хозяйства в эпоху CRISPR». Микроорганизмы , 7 (8): 269. doi : 10.3390/microorganisms7080269 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Койде, Роджер Т.; Мосс, Барбара (2004). «История исследований арбускулярной микоризы». Микориза . 14 (3): 145–163. Бибкод : 2004Mycor..14..145K . дои : 10.1007/s00572-004-0307-4 . ПМИД 15088135 . S2CID 1809402 .
- ^ Jump up to: а б Булгарелли, Давиде; Шлеппи, Клаус; Спепен, Стейн; Ван Темаат, Эмиэль Вер Лорен; Шульце-Леферт, Пол (2013). «Структура и функции бактериальной микробиоты растений». Ежегодный обзор биологии растений . 64 : 807–838. doi : 10.1146/annurev-arplant-050312-120106 . ПМИД 23373698 .
- ^ Абдельфаттах, Ахмед; Тэк, Эйко Дж. М.; Вассерман, Биргит; Лю, Цзя; Берг, Габриэле; Норелли, Джон; Дроби, Самир; Вишневский, Майкл (2021). «Доказательства коэволюции хозяина и микробиома яблока» . Новый фитолог . 234 (6): 2088–2100. дои : 10.1111/nph.17820 . ISSN 1469-8137 . ПМЦ 9299473 . ПМИД 34823272 . S2CID 244661193 .
- ^ Басби, Пози Э.; Соман, Чинмей; Вагнер, Мэгги Р.; Фризен, Марен Л.; Кремер, Джеймс; Беннетт, Элисон; Морси, Мустафа; Эйзен, Джонатан А.; Лич, Ян Э.; Дангл, Джеффри Л. (2017). «Приоритеты исследований по использованию растительных микробиомов в устойчивом сельском хозяйстве» . ПЛОС Биология . 15 (3): e2001793. дои : 10.1371/journal.pbio.2001793 . ПМК 5370116 . ПМИД 28350798 . S2CID 6434145 .
- ^ Гантар, М.; Эльхай, Дж. (1999). «Колонизация параклубеньков пшеницы N2-фиксирующей цианобактерией Nostocsp . Штамм 2S9B» . Новый фитолог . 141 (3): 373–379. дои : 10.1046/j.1469-8137.1999.00352.x .
- ^ Гантар, М. (2000). «Механическое повреждение корней обеспечивает усиленную колонизацию эндоризосферы пшеницы диазотфиксирующей цианобактерией Nostoc sp. Штамм 2S9B». Биология и плодородие почв . 32 (3): 250–255. Бибкод : 2000BioFS..32..250G . дои : 10.1007/s003740000243 . S2CID 7590731 .
- ^ Тревес, Хаим; Раанан, Аггей; Кедем, Исаак; Мюрик, Омер; Керен, Нир; Зер, Хаггит; Беркович, Саймон М.; Джордано, Марио; Норичи, Алессандра; Шотланд, Джорам; Охад, Ицхак; Каплан, Аарон (2016). «Механизмы, благодаря которым зеленая водоросль Chlorella ohadii, выделенная из корки пустынной почвы, демонстрирует беспрецедентную устойчивость к фотоповреждениям» . Новый фитолог . 210 (4): 1229–1243. дои : 10.1111/nph.13870 . ПМИД 26853530 .
- ^ Чжу, Хуан; Ли, Шуин; Ху, Чжэнъюй; Лю, Госян (2018). «Молекулярная характеристика эукариотических сообществ водорослей тропической филлосферы на основе секвенирования гена 18S рДНК в реальном времени» . Биология растений BMC . 18 (1): 365. дои : 10.1186/s12870-018-1588-7 . ПМК 6299628 . ПМИД 30563464 .
- ^ Крингс, Майкл; Хасс, Хаген; Керп, Ганс; Тейлор, Томас Н.; Агерер, Рейнхард; Доцлер, Нора (2009). «Эндофитные цианобактерии в наземном растении возрастом 400 миллионов лет: сценарий возникновения симбиоза?». Обзор палеоботаники и палинологии . 153 (1–2): 62–69. Бибкод : 2009RPaPa.153...62K . дои : 10.1016/j.revpalbo.2008.06.006 .
- ^ Картикеян Н.; Прасанна, Р.; Суд, А.; Джайсвал, П.; Наяк, С.; Кошик, Б.Д. (2009). «Физиологическая характеристика и электронно-микроскопическое исследование цианобактерий, связанных с ризосферой пшеницы». Фолиа микробиологическая . 54 (1): 43–51. дои : 10.1007/s12223-009-0007-8 . ПМИД 19330544 . S2CID 23420342 .
- ^ Jump up to: а б Бабу, Сантош; Прасанна, Радха; Бидьярани, Нгангом; Сингх, Раджендра (2015). «Анализ колонизации инокулированных цианобактерий в растениях пшеницы с использованием биохимических и молекулярных инструментов». Журнал прикладной психологии . 27 (1): 327–338. Бибкод : 2015JAPco..27..327B . дои : 10.1007/s10811-014-0322-6 . S2CID 17353123 .
- ^ Jump up to: а б Бидьярани, Нгангом; Прасанна, Радха; Чавла, Гаутама; Бабу, Сантош; Сингх, Раджендра (2015). «Расшифровка факторов, связанных с заселением растений риса цианобактериями». Журнал фундаментальной микробиологии . 55 (4): 407–419. дои : 10.1002/jobm.201400591 . ПМИД 25515189 . S2CID 5401526 .
- ^ Jump up to: а б Гантар, М.; Керби, Северо-Запад; Роуэлл, П. (1991). «Колонизация пшеницы (Triticum vulgare L.) N2-фиксирующими цианобактериями: II. Ультраструктурное исследование». Новый фитолог . 118 (3): 485–492. дои : 10.1111/j.1469-8137.1991.tb00031.x .
- ^ Ахмед, Мехбуб; Сталь, Лукас Дж.; Хаснайн, Шахида (2010). «Ассоциация негетероцистных цианобактерий с сельскохозяйственными растениями» . Растение и почва . 336 (1–2): 363–375. Бибкод : 2010ПлСой.336..363А . дои : 10.1007/s11104-010-0488-x . S2CID 21309970 .
- ^ Хусейн, Анвар; Хамаюн, Мухаммед; Шах, Сайед Тарик (2013). «Корневая колонизация и фитостимуляция фитогормонами, продуцирующими энтофитный Nostoc sp. AH-12». Современная микробиология . 67 (5): 624–630. дои : 10.1007/s00284-013-0408-4 . ПМИД 23794014 . S2CID 14704537 .
- ^ Хусейн, Анвар; Шах, Сайед Т.; Рахман, Хазир; Иршад, Мухаммед; Икбал, Амджад (2015). «Влияние IAA на рост in vitro и колонизацию Nostoc в корнях растений» . Границы в науке о растениях . 6 : 46. дои : 10.3389/fpls.2015.00046 . ПМЦ 4318279 . ПМИД 25699072 .
- ^ Jump up to: а б с Ли, Сан-Му; Рю, Чунг Мин (4 февраля 2021 г.). «Водоросли как новенькие в полезном микробиоме растений» . Границы в науке о растениях . 12 . Frontiers Media SA: 599742. doi : 10.3389/fpls.2021.599742 . ISSN 1664-462X . ПМЦ 7889962 . ПМИД 33613596 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Jump up to: а б Чаудри, Васви; Рунге, Пол; Сенгупта, Приямедха; Дёлеманн, Гюнтер; Паркер, Джейн Э; Кемен, Эрик (10 сентября 2020 г.). Коприва, Станислав (ред.). «Формирование микробиоты листа: взаимодействие растение-микроб-микроб» . Журнал экспериментальной ботаники . 72 (1). Издательство Оксфордского университета (OUP): 36–56. дои : 10.1093/jxb/eraa417 . ISSN 0022-0957 . ПМК 8210630 . ПМИД 32910810 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Вишвакарма, Канчан; Кумар, Нитин; Шандилья, Читракши; Мохапатра, Свати; Бхайана, Сахил; Варма, Аджит (2020). «Возврат к взаимодействиям растений и микробов и использованию микробных консорциумов для улучшения устойчивого сельского хозяйства: обзор» . Границы микробиологии . 11 : 560406. doi : 10.3389/fmicb.2020.560406 . ПМЦ 7779480 . ПМИД 33408698 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Хинсингер, Филипп; Бенго, А. Глин; Веттерлейн, Дорис; Янг, Иэн М. (2009). «Ризосфера: биофизика, биогеохимия и экологическая значимость». Растение и почва . 321 (1–2): 117–152. Бибкод : 2009PlSoi.321..117H . дои : 10.1007/s11104-008-9885-9 . S2CID 8997382 .
- ^ Бонковски, Майкл; Вильенав, Сесиль; Гриффитс, Брайан (2009). «Ризосферная фауна: функциональное и структурное разнообразие тесных взаимодействий почвенной фауны с корнями растений». Растение и почва . 321 (1–2): 213–233. Бибкод : 2009ПлСой.321..213Б . дои : 10.1007/s11104-009-0013-2 . S2CID 35701713 .
- ^ Jump up to: а б Хасани, М. Амин; Дуран, Палома; Хаккар, Стефан (март 2018 г.). «Микробные взаимодействия внутри холобионта растений» . Микробиом . 6 (1): 58. дои : 10.1186/s40168-018-0445-0 . ПМЦ 5870681 . ПМИД 29587885 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Ван дер Хейден, Марсель Г.А.; Мартин, Фрэнсис М.; Селосс, Марк-Андре; Сандерс, Ян Р. (2015). «Микоризная экология и эволюция: прошлое, настоящее и будущее». Новый фитолог . 205 (4): 1406–1423. дои : 10.1111/nph.13288 . ПМИД 25639293 .
- ^ Рич, Мелани К.; Нури, Ева; Курти, Пьер-Эммануэль; Рейнхардт, Дидье (2017). «Диета арбускулярных микоризных грибов: хлеб с маслом?» (PDF) . Тенденции в науке о растениях . 22 (8): 652–660. doi : 10.1016/j.tplants.2017.05.008 . ПМИД 28622919 .
- ^ Ли, Ын-Хва; Эо, Джу-Кён; Ка, Кан Хён; Эом, Ан-Хым (2013). «Разнообразие арбускулярных микоризных грибов и их роль в экосистемах» . Микобиология . 41 (3): 121–125. дои : 10.5941/MYCO.2013.41.3.121 . ПМЦ 3817225 . ПМИД 24198665 .
- ^ Ганс, Дж.; Волински, М.; Данбар, Дж. (2005). «Усовершенствования вычислений показывают большое бактериальное разнообразие и высокую токсичность металлов в почве». Наука . 309 (5739): 1387–1390. Бибкод : 2005Sci...309.1387G . дои : 10.1126/science.1112665 . ПМИД 16123304 . S2CID 130269020 .
- ^ i̇Nceoğlu, Озгюль; Аль-Суд, Валид Абу; Саллес, Жоана Фалькао; Семенов Александр В.; Ван Эльзас, Ян Дирк (2011). «Сравнительный анализ бактериальных сообществ на картофельном поле, определенный методом пиросеквенирования» . ПЛОС ОДИН . 6 (8): e23321. Бибкод : 2011PLoSO...623321I . дои : 10.1371/journal.pone.0023321 . ПМК 3158761 . ПМИД 21886785 .
- ^ Jump up to: а б с Булгарелли, Давиде; Ротт, Матиас; Шлеппи, Клаус; Вер Лорен Ван Темаат, Эмиэль; Ахмадинежад, Нахаль; Ассенца, Федерика; Рауф, Филип; Хюттель, Бруно; Рейнхардт, Ричард; Шмельцер, Элмон; Пеплиес, Йорг; Глекнер, Фрэнк Оливер; Аманн, Рудольф; Эйкхорст, Тило; Шульце-Леферт, Пол (2012). «Выявление структуры и признаков сборки бактериальной микробиоты, обитающей в корнях арабидопсиса». Природа . 488 (7409): 91-95. Стартовый код : 2012Природа.488...91Б . дои : 10.1038/nature11336 . ПМИД 22859207 . S2CID 4393146 .
- ^ Уроз, Стефан; Буэ, Марк; Мюрат, Клод; Фрей-Клетт, Паскаль; Мартин, Фрэнсис (2010). «Пиросеквенирование выявляет контрастное бактериальное разнообразие между ризосферой дуба и окружающей почвой». Отчеты по экологической микробиологии . 2 (2): 281–288. Бибкод : 2010EnvMR...2..281U . дои : 10.1111/j.1758-2229.2009.00117.x . ПМИД 23766079 .
- ^ Jump up to: а б Лундберг, Дерек С.; Лебейс, Сара Л.; Стены, Южный Кузнец; Вашстон, Скотт; Геринг, Джейс; Малфатти, Стефани; Трамбле, Жюльен; Энгельбректсон, Анна; Сегодня, Виктор; Рио, Тихана Главина; Эдгар, Роберт С.; Эйкхорст, Вещи; Ло, Рут Э.; Гугенгольц, Филип; Тринге, Сюзанна Грин; Дангл, Джеффри Л. (2012). «Определение основного микробиома корня Arabidopsis thaliana» . Природа 488 (7409): 86–90. Бибкод : 2012Природа.488...86Л . дои : 10.1038/nature11237 . ПМК 4074413 . ПМИД 22859206 .
- ^ Jump up to: а б Шлеппи, К.; Домбровский, Н.; Отер, Р.Г.; Вер Лорен Ван Темаат, Э.; Шульце-Леферт, П. (2014). «Количественное расхождение бактериальной корневой микробиоты у родственников Arabidopsis thaliana» . Труды Национальной академии наук . 111 (2): 585–592. Бибкод : 2014PNAS..111..585S . дои : 10.1073/pnas.1321597111 . ПМК 3896156 . ПМИД 24379374 . S2CID 13806811 .
- ^ Jump up to: а б с д Воку, Деспойна; Варели, Катерина; Зарали, Екатерини; Караманоли, Катерина; Константиниду, Хелен-Исида А.; Монокрусос, Николаос; Хэлли, Джон М.; Сайнис, Иоаннис (2012). «Изучение биоразнообразия бактериального сообщества средиземноморской филлосферы и его связи с бактериями, передающимися по воздуху». Микробная экология . 64 (3): 714–724. Бибкод : 2012MicEc..64..714V . дои : 10.1007/s00248-012-0053-7 . ПМИД 22544345 . S2CID 17291303 .
- ^ Дастогер, Хондокер М.Г.; Ли, Хуа; Сиваситхампарам, Кришнапиллай; Джонс, Майкл Г.К.; Ду, Синь; Рен, Юнлинь; Уайли, Стивен Дж. (2017). «Метаболические реакции эндофитных растений Nicotiana benthamiana, испытывающих водный стресс» . Экологическая и экспериментальная ботаника . 143 : 59–71. Бибкод : 2017EnvEB.143...59D . дои : 10.1016/j.envexpbot.2017.08.008 .
- ^ Родригес, Р.Дж.; Уайт-младший, Дж. Ф.; Арнольд, А.Е.; Редман, RS (2009). «Грибные эндофиты: разнообразие и функциональные роли» . Новый фитолог . 182 (2): 314–330. дои : 10.1111/j.1469-8137.2009.02773.x . ПМИД 19236579 .
- ^ Jump up to: а б с д Абдельфаттах, Ахмед; Вишневский, Майкл; Шена, Леонардо; Тэк, Айко Дж. М. (2021). «Экспериментальные доказательства наследования микробов у растений и путей передачи от семян к филлосфере и корням» . Экологическая микробиология . 23 (4): 2199–2214. Бибкод : 2021EnvMi..23.2199A . дои : 10.1111/1462-2920.15392 . ISSN 1462-2920 . ПМИД 33427409 .
- ^ Хэ, Шэн-Ян (28 августа 2020 г.). «Когда растения и их микробы не синхронизированы, результаты могут быть катастрофическими» . Разговор .
- ^ Линдоу, Стивен Э. (1996). «Роль иммиграции и других процессов в определении популяций эпифитных бактерий». Воздушная поверхностная микробиология растений . стр. 155–168. дои : 10.1007/978-0-585-34164-4_10 . ISBN 978-0-306-45382-3 .
- ^ Фризен, Марен Л.; Портер, Стефани С.; Старк, Скотт С.; фон Веттберг, Эрик Дж.; Сакс, Джоэл Л.; Мартинес-Ромеро, Эсперанса (2011). «Микробно-опосредованные функциональные особенности растений». Ежегодный обзор экологии, эволюции и систематики . 42 : 23–46. doi : 10.1146/annurev-ecolsys-102710-145039 .
- ^ Jump up to: а б Лево, Йохан Х.Дж. (2019). «Краткое описание с листа: Последние исследования, способствующие нашему пониманию микробиома филлосферы». Современное мнение в микробиологии . 49 : 41–49. дои : 10.1016/j.mib.2019.10.002 . ПМИД 31707206 . S2CID 207946690 .
- ^ Руинен, Дж. (февраль 1956 г.). «Распространение видов Beijerinckia в« филлосфере » ». Природа . 177 (4501): 220–221. дои : 10.1038/177220a0 . ISSN 1476-4687 .
- ^ Jump up to: а б Фогель, Кристина ; Боденхаузен, Наташа; Грюиссем, Вильгельм; Ворхольт, Джулия А. (2016). «Транскриптом листьев арабидопсиса демонстрирует различные, но также перекрывающиеся реакции на колонизацию филлосферными комменсалами и патогенную инфекцию, оказывающую влияние на здоровье растений» (PDF) . Новый фитолог . 212 (1): 192–207. дои : 10.1111/nph.14036 . hdl : 20.500.11850/117578 . ПМИД 27306148 .
- ^ Jump up to: а б Сид, Фернанда П.; Маруяма, Фумито; Мурасе, Кадзунори; Гретер, Стивен П.; Ларама, Джованни; Браво, Леон А.; Хоркера, Милко А. (2018). «Проект последовательности генома бактерий, выделенных из филлосферы Deschampsia antarctica». Экстремофилы . 22 (3): 537–552. дои : 10.1007/s00792-018-1015-x . ПМИД 29492666 . S2CID 4320165 .
- ^ Кумаравел, Совья; Танкаппан, Сугита; Рагхупати, Шридар; Утанди, Сивакумар (2018). «Проект геномной последовательности стимулирующей рост растений и устойчивой к засухе Bacillus altitudinis FD48, выделенной из филлопланы риса» . Геномные объявления . 6 (9). doi : 10.1128/genomeA.00019-18 . ПМЦ 5834328 . ПМИД 29496824 .
- ^ Лафорест-Лапуант, Изабель; Уитакер, Бриана К. (2019). «Расшифровка микробиоты филлосферы: прогресс и проблемы» . Американский журнал ботаники . 106 (2): 171–173. дои : 10.1002/ajb2.1229 . ПМИД 30726571 .
- ^ Jump up to: а б с д и ж г час я дж к Благородная, Аня С.; Нет, Стиви; Клируотер, Майкл Дж.; Ли, Чарльз К. (2020). «Основной микробиом филлосферы существует в отдаленных популяциях видов деревьев, произрастающих в Новой Зеландии» . ПЛОС ОДИН . 15 (8): e0237079. Бибкод : 2020PLoSO..1537079N . дои : 10.1371/journal.pone.0237079 . ПМЦ 7425925 . ПМИД 32790769 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Jump up to: а б с Финкель, Омри М.; Берч, Эдриан Ю.; Линдоу, Стивен Э.; Пост, Антон Ф.; Белкин, Шимшон (2011). «Географическое положение определяет популяционную структуру филлосферных микробных сообществ соляного пустынного дерева» . Прикладная и экологическая микробиология . 77 (21): 7647–7655. Бибкод : 2011ApEnM..77.7647F . дои : 10.1128/АЕМ.05565-11 . ПМК 3209174 . ПМИД 21926212 .
- ^ Jump up to: а б Ворхольт, Джулия А. (2012). «Микробная жизнь в филлосфере». Обзоры природы Микробиология . 10 (12): 828–840. дои : 10.1038/nrmicro2910 . hdl : 20.500.11850/59727 . ПМИД 23154261 . S2CID 10447146 .
- ^ Линдоу, Стивен Э.; Брандл, Мария Т. (2003). «Микробиология филлосферы» . Прикладная и экологическая микробиология . 69 (4): 1875–1883. Бибкод : 2003ApEnM..69.1875L . дои : 10.1128/АЕМ.69.4.1875-1883.2003 . ПМК 154815 . ПМИД 12676659 . S2CID 2304379 .
- ^ Боденхаузен, Наташа; Хортон, Мэтью В.; Бергельсон, Джой (2013). «Бактериальные сообщества, связанные с листьями и корнями Arabidopsis thaliana» . ПЛОС ОДИН . 8 (2): e56329. Бибкод : 2013PLoSO...856329B . дои : 10.1371/journal.pone.0056329 . ПМЦ 3574144 . ПМИД 23457551 .
- ^ Jump up to: а б Стоун, Брэм В.Г.; Джексон, Колин Р. (2016). «Биогеографические закономерности между бактериальными филлосферными сообществами южной магнолии (Magnolia grandiflora) в небольшом лесу». Микробная экология . 71 (4): 954–961. Бибкод : 2016MicEc..71..954S . дои : 10.1007/s00248-016-0738-4 . ПМИД 26883131 . S2CID 17292307 .
- ^ Jump up to: а б Редфорд, Аманда Дж.; Бауэрс, Роберт М.; Найт, Роб; Линхарт, Ян; Фирер, Ной (2010). «Экология филлосферы: Географическая и филогенетическая изменчивость распределения бактерий на листьях деревьев» . Экологическая микробиология . 12 (11): 2885–2893. Бибкод : 2010EnvMi..12.2885R . дои : 10.1111/j.1462-2920.2010.02258.x . ПМК 3156554 . ПМИД 20545741 .
- ^ Jump up to: а б с Лафорест-Лапуант, Изабель; Мессье, Кристиан; Кембель, Стивен В. (2016). «Идентичность вида-хозяина, место и время определяют структуру бактериального сообщества филлосферы деревьев умеренного климата» . Микробиом . 4 (1): 27. дои : 10.1186/s40168-016-0174-1 . ПМЦ 4912770 . ПМИД 27316353 .
- ^ Сарраонаиндия, Ираче; Оуэнс, Сара М.; Вайзенхорн, Памела; Уэст, Кристин; Хэмптон-Марселл, Джаррад; Лакс, Саймон; Бокулич, Николай Александрович; Миллс, Дэвид А.; Мартин, Жиль; Тагави, Сафий; Ван дер Лели, Дэниел; Гилберт, Джек А. (2015). «Микробиом почвы влияет на микробиоту, связанную с виноградной лозой» . мБио . 6 (2). дои : 10.1128/mBio.02527-14 . ПМЦ 4453523 . ПМИД 25805735 .
- ^ Финкель, Омри М.; Берч, Эдриан Ю.; Элад, Таль; Хьюз, Сьюзен М.; Линдоу, Стивен Э.; Пост, Антон Ф.; Белкин, Шимшон (2012). «Зависимость расстояния от распада частично определяет структуру разнообразия филлосферных бактерий на деревьях тамрикс в пустыне Сонора» . Прикладная и экологическая микробиология . 78 (17): 6187–6193. Бибкод : 2012ApEnM..78.6187F . дои : 10.1128/AEM.00888-12 . ПМЦ 3416633 . ПМИД 22752165 .
- ^ Jump up to: а б Кембель, Юго-Запад; О'Коннор, ТК; Арнольд, Гонконг; Хаббелл, СП; Райт, С.Дж.; Грин, Дж.Л. (2014). «Связь между бактериальными сообществами филлосферы и функциональными особенностями растений в неотропическом лесу» . Труды Национальной академии наук . 111 (38): 13715–13720. Бибкод : 2014PNAS..11113715K . дои : 10.1073/pnas.1216057111 . ПМК 4183302 . ПМИД 25225376 . S2CID 852584 .
- ^ Иннеребнер, Герд; Книф, Клаудия; Ворхольт, Джулия А. (2011). «Защита Arabidopsis thaliana от патогенных для листьев Pseudomonas syringae штаммами Sphingomonas в контролируемой модельной системе» . Прикладная и экологическая микробиология . 77 (10): 3202–3210. Бибкод : 2011ApEnM..77.3202I . дои : 10.1128/AEM.00133-11 . ПМК 3126462 . ПМИД 21421777 .
- ^ Лажуа, Женевьева; Мальоне, Реми; Кембель, Стивен В. (2020). «Адаптивное соответствие между филлосферными бактериями и их древесными хозяевами в неотропическом лесу» . Микробиом . 8 (1): 70. дои : 10.1186/s40168-020-00844-7 . ПМЦ 7243311 . ПМИД 32438916 .
- ^ Jump up to: а б Шейд, Эшли; Хандельсман, Джо (2012). «За пределами диаграммы Венна: охота за основным микробиомом» . Экологическая микробиология . 14 (1): 4–12. Бибкод : 2012EnvMi..14....4S . дои : 10.1111/j.1462-2920.2011.02585.x . ПМИД 22004523 .
- ^ Берг, Габриэле; Рыбакова Дарья; Фишер, Дорин; Чернава, Томислав; Верже, Мари-Кристин Шампомье; Чарльз, Тревор; Чен, Сяоюлун; Коколин, Лука; Эверсол, Келли; Коррал, Хема Эрреро; Казу, Мария; Кинкель, Линда; Ланге, Лене; Лима, Нельсон; Лой, Александр; и др. (2020). «Повторное рассмотрение определения микробиома: старые концепции и новые проблемы» . Микробиом . 8 (1): 103. дои : 10.1186/s40168-020-00875-0 . ПМЦ 7329523 . ПМИД 32605663 .
- ^ Тернбо, Питер Дж.; Хамади, Мика; Яцуненко Таня; Кантарел, Брэнди Л.; Дункан, Алексис; Лей, Рут Э.; Согин, Митчелл Л.; Джонс, Уильям Дж.; Роу, Брюс А.; Аффуртит, Джейсон П.; Эгхольм, Майкл; Анрисса, Бернар; Хит, Эндрю С.; Найт, Роб; Гордон, Джеффри И. (2009). «Основной микробиом кишечника у тучных и худых близнецов» . Природа . 457 (7228): 480–484. Бибкод : 2009Natur.457..480T . дои : 10.1038/nature07540 . ПМЦ 2677729 . ПМИД 19043404 .
- ^ Хамонтс, Келли; Триведи, Панкадж; Гарг, Аншу; Яниц, Кэролайн; Гринье, Жасмин; Холфорд, Пол; Бота, Фредерик К.; Андерсон, Ян К.; Сингх, Браджеш К. (2018). «Полевые исследования показывают основную микробиоту растений и относительную важность ее движущих сил». Экологическая микробиология . 20 (1): 124–140. Бибкод : 2018EnvMi..20..124H . дои : 10.1111/1462-2920.14031 . ПМИД 29266641 . S2CID 10650949 .
- ^ Чернава, Томислав; Эрлахер, Армин; Сох, Юнг; Сенсен, Кристоф В.; Грубе, Мартин; Берг, Габриэле (2019). «Энтеробактерии доминируют в основном микробиоме и вносят вклад в резистом рукколы (Eruca sativa Mill.)» . Микробиом . 7 (1): 13. дои : 10.1186/s40168-019-0624-7 . ПМК 6352427 . ПМИД 30696492 .
- ^ Лефф, Джонатан В.; Дель Тредичи, Питер; Фридман, Уильям Э.; Фирер, Ной (2015). «Пространственное структурирование бактериальных сообществ внутри отдельных двудольных деревьев гинкго ». Экологическая микробиология . 17 (7): 2352–2361. Бибкод : 2015EnvMi..17.2352L . дои : 10.1111/1462-2920.12695 . ПМИД 25367625 .
- ^ Эрнандес-Агреда, Алехандра; Гейтс, Рут Д.; Эйнсворт, Трейси Д. (2017). «Определение основного микробиома в микробном супе кораллов». Тенденции в микробиологии . 25 (2): 125–140. дои : 10.1016/j.tim.2016.11.003 . ПМИД 27919551 .
- ^ Стивенс, JMC; Молан, ПК; Кларксон, Б.Д. (2005). «Обзор Leptospermum scoparium (Myrtaceae) в Новой Зеландии» . Новозеландский журнал ботаники . 43 (2): 431–449. Бибкод : 2005NZJB...43..431S . дои : 10.1080/0028825X.2005.9512966 . S2CID 53515334 .
- ^ Купер, РА; Молан, ПК; Хардинг, КГ (2002). «Чувствительность к меду грамположительных кокков клинического значения, выделенных из ран». Журнал прикладной микробиологии . 93 (5): 857–863. дои : 10.1046/j.1365-2672.2002.01761.x . ПМИД 12392533 . S2CID 24517001 .
- ^ Раби, Эрика; Серем, Джун Чепту; Оберхольцер, Хестер Магдалена; Гаспар, Анабелла Регина Маркес; Бестер, Меган Джин (2016). «Как метилглиоксаль убивает бактерии: ультраструктурное исследование». Ультраструктурная патология . 40 (2): 107–111. дои : 10.3109/01913123.2016.1154914 . hdl : 2263/52156 . ПМИД 26986806 . S2CID 13372064 .
- ^ Джонстон, Питер Р. (1998). «Эндофиты листьев мануки (Leptospermum scoparium)». Микологические исследования . 102 (8): 1009–1016. дои : 10.1017/S0953756297005765 .
- ^ Маккензи, EHC; Джонстон, PR; Бьюкенен, ПК (2006). «Контрольный список грибов чайного дерева (виды Kunzea и Leptospermum ) в Новой Зеландии» . Новозеландский журнал ботаники . 44 (3): 293–335. Бибкод : 2006NZJB...44..293M . дои : 10.1080/0028825X.2006.9513025 . S2CID 84538904 .
- ^ Викаксоно, Вишну Ади; Сансом, Кэтрин Э.; Эйриан Джонс, Э.; Перри, Найджел Б.; Монк, Яна; Риджуэй, Хейли Дж. (2018). «Арбускулярные микоризные грибы, связанные с Leptospermum scoparium (Mānuka): влияние на рост растений и содержание эфирного масла». Симбиоз . 75 (1): 39–50. Бибкод : 2018Symbi..75...39W . дои : 10.1007/s13199-017-0506-3 . S2CID 4819178 .
- ^ Викаксоно, Вишну Ади; Джонс, Э. Эйриан ; Монк, Яна; Риджуэй, Хейли Дж. (2016). «Бактериальная подпись Leptospermum scoparium (Mānuka) выявляет основные и вспомогательные сообщества с биоактивными свойствами» . ПЛОС ОДИН . 11 (9): e0163717. Бибкод : 2016PLoSO..1163717W . дои : 10.1371/journal.pone.0163717 . ПМК 5038978 . ПМИД 27676607 .
- ^ Jump up to: а б с д Джонстон-Монже, Дэвид; Гутьеррес, Джаннет П; Лопес-Лаваль, Луис АБ (2021). «Бактерии и грибы, передающиеся с семенами, доминируют в микробиомах юных растений» . Границы микробиологии . 12 . дои : 10.3389/fmicb.2021.737616 . ПМЦ 8569520 . ПМИД 34745040 .
- ^ Jump up to: а б с Война, Аадиль Фарук; Башир, Икра; Реши, Зафар А.; Кардол, Пол; Рашид, Ирфан (апрель 2023 г.). «Понимание микробиома семян и его экологического значения в жизни растений» . Микробиологические исследования . 269 . дои : 10.1016/j.micres.2023.127318 . ПМИД 36753851 . S2CID 256563377 .
- ^ Мацумото, Харуна; Ван, Кусстатчер, Цзе, Саньлин; Ван, Илин; Чжу, Хашидоко, Габриэле; Чернава, Томислав; Ван, Менгсен (4 января 2021 г.). «Бактериальный эндофит семян формирует устойчивость природных растений к болезням». .7 33398157. : 60–72. doi : 10.1038/s41477-020-00826-5 . PMID ) ( 1 S2CID 230508404 .
- ^ Нельсон, Эрик Б. (2018). «ОБЗОР: Микробиом семян: происхождение, взаимодействие и влияние». Растение и почва . 422 : 7–34. дои : 10.1007/s11104-017-3289-7 .
- ^ «Семена передают свои микробы следующему поколению» . ЭврекАлерт! АААС . Проверено 21 января 2021 г.
- ^ Jump up to: а б Абдельфаттах, Ахмед; Тэк, Эйко Дж. М.; Лобато, Каролина; Вассерманн, Биргит; Берг, Габриэле (апрель 2023 г.). «От семени к семени: роль микробной наследственности в формировании микробиома растения» . Тенденции в микробиологии . 31 (4): 346–355. дои : 10.1016/j.tim.2022.10.009 . ПМИД 36481186 .
- ^ Барлоу, Конни. «Опубликованные документы о причинах угрозы исчезновения Torreya Taxifolia во Флориде» . Стражи Торрея . Проверено 22 февраля 2024 г.
Справочники
[ редактировать ]- Салим М. (2015) Экология микробиомного сообщества: основы и приложения Springer. ISBN 9783319116655 .
- Кумар В., Прасад Р., Кумар М. и Чоудхари Д.К. (2019) Микробиом в здоровье и болезнях растений: проблемы и возможности Springer. ISBN 9789811384950 .
- Кумар, Вивек; Прасад, Рам; Кумар, Манодж; Чоудхари, Девендра К. (10 августа 2019 г.). Микробиом в здоровье и болезнях растений: проблемы и возможности . Спрингер. ISBN 9789811384950 .
- Грубе, Мартин; Шлотер, Майкл; Смолла, Корнелия; Берг, Габриэле (22 января 2015 г.). Микробиом растений и его значение для здоровья растений и человека . Том. 5. с. 491. дои : 10.3389/fmicb.2014.00491 . ISBN 9782889193783 . ПМК 4166366 . ПМИД 25278934 .
- Эгамбердиева, Дилфуза; Ахмад, Парваиз (6 февраля 2018 г.). Микробиом растений: реакция на стресс . Спрингер. ISBN 9789811055140 .
- Кастильоне, Стефано; Чикателли, Анджела; Феррол, Нурия; Розпадек, Петр (22 августа 2019 г.). Влияние взаимодействия растений и микробиома на способность к фито- и биовосстановлению . Фронтирс Медиа С.А. ISBN 9782889459322 .
- Кастильоне, Стефано; Чикателли, Анджела; Феррол, Нурия; Розпадек, Петр (22 августа 2019 г.). Влияние взаимодействия растений и микробиома на способность к фито- и биовосстановлению . Фронтирс Медиа С.А. ISBN 9782889459322 .
- КАРВАЛЬЯИС, Лилия К.; Деннис, Пол Г. (30 декабря 2020 г.). Растительный микробиом: методы и протоколы . Спрингер. ISBN 9781071610398 .
- Кумар, Вивек; Прасад, Рам; Кумар, Манодж; Чоудхари, Девендра К. (10 августа 2019 г.). Микробиом в здоровье и болезнях растений: проблемы и возможности . Спрингер. ISBN 9789811384950 .
- Соланки, Манодж Кумар; Кашьяп, Прем Лал; Ансари, Ризван Али; Кумари, Бэби (28 августа 2020 г.). Микробиомы и здоровье растений: Паноплия и их применение . Академическая пресса. ISBN 9780128226018 .
- Ядав, Аджар (2020). Достижения в области микробиома растений и устойчивого сельского хозяйства, функциональная аннотация и будущие задачи . Сингапур: Спрингер. ISBN 978-981-15-3204-7 . OCLC 1193118045 .
- Варма, Аджит (28 августа 2020 г.). Парадигма микробиома растений . Спрингер. ISBN 9783030503956 .
- Доти, Шэрон Лафферти (21 сентября 2017 г.). Функциональное значение микробиома растений: значение для сельского хозяйства, лесного хозяйства и биоэнергетики . Спрингер. ISBN 9783319658971 .
- Антвис, Рэйчел Э.; Харрисон, Ксавье А.; Кокс, Майкл Дж. (12 марта 2020 г.). Микробиомы почв, растений и животных: комплексный подход . Издательство Кембриджского университета. ISBN 9781108473712 .
- Варма, Аджит; Трипати, Свати; Прасад, Рам (20 октября 2020 г.). Парадигма микробиома растений . Спрингер. ISBN 9783030503949 .
- Розенберг, Юджин; Зильбер-Розенберг, Илана (31 января 2014 г.). Концепция гологенома: микробиота человека, животных и растений . Спрингер. ISBN 9783319042411 .
- Ядав, Аджар Натх; Сингх, Джогиндер; Растегари, Али Асгар; Ядав, Нилам (6 марта 2020 г.). Растительные микробиомы для устойчивого сельского хозяйства . Спрингер. ISBN 9783030384531 .
