Морфология насекомых

Легенда о частях тела
Тагматы: А – Голова, Б – Грудная клетка, С – Брюшко.
- антенна
- глазки (нижние)
- глазки (верхние)
- сложный глаз
- мозг (мозговые ганглии )
- переднегрудь
- спинной кровеносный сосуд
- трахеальные трубки (ствол со дыхальцем )
- среднегруди
- заднегруди
- переднее крыло
- заднее крыло
- средняя кишка (желудок)
- спинная трубка (сердце)
- яичник
- задняя кишка (кишечник, прямая кишка и анус)
- анус
- яйцевод
- нервный канатик (брюшные ганглии)
- Мальпигиевы трубки
- предплюсневые подушечки
- когти
- предплюсна
- большеберцовая кость
- бедренная кость
- вертел
- передняя кишка (зоб, желудок)
- грудной ганглий
- бедро
- слюнная железа
- подпищеводный ганглий
- ротовой аппарат
Морфология насекомых это изучение и описание физической формы насекомых — . Терминология, используемая для описания насекомых, аналогична терминологии, используемой для других членистоногих, из-за их общей истории эволюции. Три физических особенности отличают насекомых от других членистоногих: у них есть тело, разделенное на три области (так называемые тагматы) (голова, грудь и брюшко), три пары ног и ротовой аппарат, расположенный за пределами головной капсулы . Такое положение ротового аппарата отделяет их от ближайших родственников, ненасекомых, шестиногих , к которым относятся Protura , Diplura и Collembola .
У разных видов насекомых существуют огромные различия в строении тела. Особи могут иметь размер от 0,3 мм ( мушки ) до 30 см в поперечнике ( большая совушка ); [ 1 ] : 7 глаз нет или много; хорошо развитые крылья или отсутствуют; а ноги модифицированы для бега, прыжков, плавания или даже рытья. Эти модификации позволяют насекомым занять почти все экологические ниши , кроме глубин океана. В этой статье описывается основное тело насекомого и некоторые вариации различных частей тела; при этом он определяет многие технические термины, используемые для описания тел насекомых.
Краткое изложение анатомии
[ редактировать ]Насекомые, как и все членистоногие, не имеют внутреннего скелета; вместо этого у них есть экзоскелет — твердый внешний слой, состоящий в основном из хитина , который защищает и поддерживает тело. Тело насекомого разделено на три части : голову, грудь и брюшко . [ 2 ] Голова специализируется на сенсорной обработке и приеме пищи; грудная клетка, которая является точкой опоры для ног и крыльев (если они есть), предназначена для передвижения; а живот предназначен для пищеварения , дыхания , выделения и размножения. [ 1 ] : 22–48 Хотя общая функция трех областей тела одинакова для всех видов насекомых, существуют серьезные различия в базовой структуре: крылья, ноги, усики и ротовой аппарат варьируются от группы к группе. [ 3 ]
Внешний
[ редактировать ]Экзоскелет
[ редактировать ]Внешний скелет насекомого, кутикула , состоит из двух слоев; эпикутикула называется , представляющая собой тонкий восковой водостойкий внешний слой, лишенный хитина, а слой под ним прокутикулой . Он хитиновый, намного толще эпикутикулы и имеет два слоя: внешний — экзокутикула, а внутренний — эндокутикула. Жесткая и гибкая эндокутикула построена из многочисленных слоев волокнистого хитина и белков, перекрещивающихся друг с другом в виде сэндвича, тогда как экзокутикула жесткая и склеротизована . [ 1 ] : 22–24 Экзокутикула сильно редуцирована у многих мягкотелых насекомых, особенно на личиночных стадиях (например, у гусениц ). Химически хитин представляет собой длинноцепочечный полимер , N-ацетилглюкозамина производного глюкозы. В неизмененной форме хитин полупрозрачен, податлив и эластичен. Однако у членистоногих он часто видоизменяется, внедряясь в затвердевший белковый матрикс, который образует большую часть экзоскелета . В чистом виде он кожистый, но при инкрустации карбонатом кальция становится намного твёрже. [ 4 ] Разница между неизмененной и видоизмененной формами видна при сравнении стенки тела гусеницы ( немодифицированной) и жука (измененной).
На эмбриональных стадиях слой столбчатых или кубовидных эпителиальных клеток дает начало внешней кутикуле и внутренней базальной мембране. Большая часть материала насекомых находится внутри эндокутикулы. Кутикула обеспечивает мышечную поддержку и действует как защитный щит по мере развития насекомого. Однако, поскольку она не может расти, внешняя склеротизированная часть кутикулы периодически отбрасывается в процессе, называемом «линькой». По мере приближения времени линьки большая часть материала экзокутикулы реабсорбируется. При линьке старая кутикула отделяется от эпидермиса ( аполиз ). Затем между старой кутикулой и эпидермисом выделяется ферментативная жидкость линьки, которая отделяет экзокутикулу, переваривая эндокутикулу и изолируя ее материал для новой кутикулы. Когда новая кутикула сформировалась в достаточной степени, эпикутикула и редуцированная экзокутикула отслаиваются в результате шелушения . [ 5 ] : 16–20
Четырьмя основными областями сегмента тела насекомого являются тергум или спинная часть, грудина или вентральная часть и две плевры или боковые части. Затвердевшие пластинки экзоскелета называются склеритами и представляют собой подразделения основных областей - тергитов, стернитов и плевритов для соответствующих областей тергума, грудины и плевры. [ 6 ]
Голова
[ редактировать ]


Голова головную у большинства насекомых заключена в твердую, сильно склеротизированную экзоскелетную капсулу . Основное исключение составляют те виды, личинки которых не полностью склеротизированы, главным образом некоторые голометаболы; но даже большинство несклеротизированных или слабо склеротизированных личинок имеют тенденцию иметь хорошо склеротизованные головные капсулы, например личинки жесткокрылых и перепончатокрылых. Однако у личинок Cyclorrhapha , как правило, практически нет головной капсулы.
Головная капсула содержит большую часть органов чувств, включая усики, глазки и сложные глаза, а также ротовой аппарат. У взрослого насекомого головная капсула кажется несегментированной, хотя эмбриологические исследования показывают, что она состоит из шести сегментов, несущих парные головные придатки, включая ротовой аппарат, каждая пара на определенном сегменте. [ 7 ] Каждая такая пара занимает один сегмент, хотя не все сегменты современных насекомых несут видимые придатки.
Из всех отрядов насекомых прямокрылые демонстрируют наибольшее разнообразие особенностей головы насекомых, включая швы и склериты . [ 6 ] Здесь вершина , или вершина (дорсальная область), расположена между сложными глазами насекомых с гипогнатной и опистогнатной головой. У прогнатных насекомых вершина находится не между сложными глазами, а там, где глазки обычно находятся . Это связано с тем, что основная ось головы повернута на 90° и становится параллельной основной оси тела. У некоторых видов этот регион видоизменяется и получает другое название. [ 8 ] : 13
Экдизиальный шов состоит из венечных, лобных и эпикраниальных швов, а также экдизиальных линий и линий расщепления, которые различаются у разных видов насекомых. Экдизиальный шов продольно накладывают на макушку, разделяя эпикраниальные половины головы на левую и правую стороны. В зависимости от насекомого шов может иметь разную форму: Y, U или V. Расходящиеся линии, составляющие экдизиальный шов, называются лобными или лобногенными швами. швы раскрываются Не у всех видов насекомых есть лобные швы, но у тех, у кого они есть, во время шелушения , что обеспечивает выход нового возраста из покровов.
Лоб — это часть головной капсулы, расположенная вентрально или кпереди от темени. Размер лба зависит от насекомого, и у многих видов определение его границ произвольно, даже у некоторых таксонов насекомых, которые имеют четко выраженные головные капсулы. Однако у большинства видов лоб спереди окаймлен лобно-клипеальной или эпистомальной бороздой над наличником. С латеральной стороны она ограничена лобно-половинной бороздой, если она имеется, а граница с макушкой - линией экдизиального расщепления, если она видна. Если есть срединный глазок, то обычно он находится на лбу, хотя у некоторых насекомых, например у многих перепончатокрылых, все три глазка появляются на макушке. Более формальное определение состоит в том, что это склерит, из которого возникают мышцы-расширители глотки, но во многих случаях это тоже бесполезно. [ 7 ] В анатомии некоторых таксонов, например многих Cicadomorpha , передняя часть головы довольно четко выражена и имеет тенденцию быть широкой и субвертикальной; эту срединную область обычно принимают за лоб. [ 9 ]
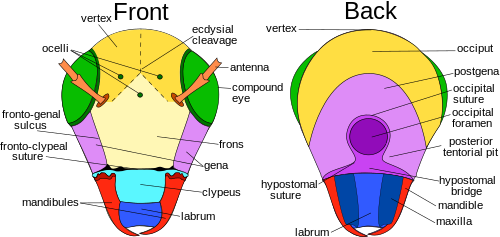
Наличник представляет собой склерит между лицом и верхней губой, который у примитивных насекомых отделен от лба лобно-клипеальным швом. Клипеогенный шов латерально отграничивает клипеус, причем наличник вентрально отделен от верхней губы клипеолабральным швом. Наличник различается по форме и размеру, как, например, у видов чешуекрылых с крупным наличником с удлиненным ротовым аппаратом. Щека или щека образуют склеротизированную область на каждой стороне головы ниже сложных глаз, доходящую до горлового шва. Как и многие части головы насекомого, щека варьируется у разных видов, и ее границы трудно установить. У стрекоз и стрекоз он находится между сложными глазами, наличником и ротовым аппаратом. Постгена — это область непосредственно сзади, сзади или ниже щеки крыловидных насекомых , образующая латеральную и вентральную части затылочной дуги. Затылочная дуга представляет собой узкую полосу, образующую задний край головной капсулы, выгибающуюся дорсально над отверстием. Субгенальная область обычно узкая, расположена над ротовым аппаратом; эта область также включает в себя гипостома и плевростома . [ 8 ] : 13–14 Темя выступает вперед над основаниями усиков в виде выступающего, заостренного, вогнутого рострума. Задняя стенка головной капсулы пронизана крупным отверстием — отверстием. Через него проходят системы органов, такие как нервный канатик , пищевод , слюнные протоки и мускулатура , соединяющая голову с грудной клеткой . [ 10 ]
На задней стороне головы расположены затылок , постгенальная кость , затылочное отверстие , задняя тенториальная ямка , губа, постгенальный мостик , гипостомальный шов и мостик , а также нижние челюсти , губа и верхняя челюсть . Затылочный шов хорошо развит у видов прямокрылых, но не так сильно у других отрядов. Затылочный шов там, где он обнаружен, представляет собой дугообразную подковообразную бороздку на затылке, заканчивающуюся на задней части каждой нижней челюсти. Постокципитальный шов является ориентиром на задней поверхности головы и обычно располагается вблизи затылочных костей. У крыловидных заднезатылочный отдел образует крайнюю заднюю часть, часто U-образную, образующую край головы, доходящий до постзатылочного шва. У крыловидных, например прямокрылых, затылочное отверстие и рот не разделены. Три типа затылочных замыканий или точек под затылочным отверстием, которые разделяют две нижние половины постгена, - это гипостомальный мост, постгенальный мост и гула. Гипостомальный мост обычно встречается у насекомых гипогнатной ориентации. Постгенальный мост встречается у взрослых видов высших Diptera и шиповидные перепончатокрылые , а гула встречается у некоторых Coleoptera , Neuroptera и Isoptera , у которых обычно ротовой аппарат ориентирован прогнатно. [ 8 ] : 15
Сложные глаза и глазки
[ редактировать ]У большинства насекомых есть одна пара больших, выступающих сложных глаз, состоящих из единиц, называемых омматидиями ( ommatidium , единственное число), до 30 000 в одном сложном глазу, например, у крупных стрекоз. Глаза этого типа дают меньшую разрешающую способность, чем глаза позвоночных, но дают острое восприятие движения и обычно обладают чувствительностью к ультрафиолету и зеленому свету, а также могут иметь дополнительные пики чувствительности в других областях зрительного спектра. Часто способность обнаруживать E-вектор поляризованного света существует при поляризации света. [ 11 ] Также могут быть дополнительные два или три глазка, которые помогают обнаружить слабую освещенность или небольшие изменения интенсивности света. Воспринимаемое изображение представляет собой комбинацию входных сигналов от многочисленных омматидиев, расположенных на выпуклой поверхности и, таким образом, направленных в несколько разных направлениях. По сравнению с простыми глазами сложные глаза обладают очень большими углами зрения и лучшей остротой, чем дорсальные глазки насекомого, но некоторые стеблевые (= личиночные глаза), например, у личинок пилильщиков ( Tenthredinidae ) с остротой зрения 4 градуса и очень высокой поляризацией. чувствительность, соответствует производительности сложных глаз. [ 12 ] [ 13 ]


Поскольку отдельные линзы настолько малы, эффекты дифракции налагают ограничение на возможное разрешение, которое можно получить (при условии, что они не функционируют как фазированные решетки ). Противостоять этому можно только увеличением размера и количества линз. Чтобы видеть с разрешением, сравнимым с разрешением наших простых глаз, людям потребовались бы сложные глаза, каждый из которых достиг бы размера их головы. Сложные глаза делятся на две группы: аппозиционные глаза, формирующие несколько перевернутых изображений, и суперпозиционные глаза, формирующие одно вертикальное изображение. [ 14 ] [ 15 ] Сложные глаза растут по краям с добавлением новых омматидий. [ 16 ]
Антенны
[ редактировать ]
Усики , иногда называемые «щупальцами», представляют собой гибкие придатки, расположенные на голове насекомого и используемые для зондирования окружающей среды. Насекомые могут чувствовать свои усики благодаря тонким волоскам ( щетинкам ), которые их покрывают. [ 17 ] : 8–11 Однако прикосновение — не единственное, что могут обнаружить усики; многочисленные крошечные сенсорные структуры на усиках позволяют насекомым чувствовать запахи, температуру, влажность, давление и даже потенциально ощущать себя в космосе . [ 17 ] : 8–11 [ 18 ] [ 19 ] Некоторые насекомые, в том числе пчелы и некоторые группы мух, также могут улавливать звук своими усиками. [ 20 ]
Число сегментов антенны у разных насекомых варьируется: у высших мух 3-6 сегментов. [ 21 ] в то время как у взрослых тараканов их может быть более 140. [ 22 ] Общая форма усиков также весьма разнообразна, но первый членик (тот, что прикреплен к голове) всегда называется скапусом, а второй сегмент — цветоножкой. Остальные сегменты усиков или жгутики называются жгутиками. [ 17 ] : 8–11
Общие типы антенн насекомых показаны ниже:
Аристат |
Ты случаешься |
Булавовидный |
нитевидный |
Фантастически |
коленчатый |
щетинистый |
Ламеллат |
Монилиформный |
пектинат |
Перистый |
Зубчатый |
Стилет |
Ротовые органы
[ редактировать ]Ротовой аппарат насекомых состоит из верхней челюсти, губ, а у некоторых видов — жвал. [ 8 ] : 16 [ 23 ] Верхняя губа представляет собой простой сросшийся склерит, часто называемый верхней губой, подвижный в продольном направлении. Он шарнирно прикреплен к наличнику. Нижние челюсти (челюсти) представляют собой пару сильно склеротизированных структур, которые движутся под прямым углом к телу и используются для откусывания, жевания и разделения пищи. Максиллы представляют собой парные структуры, которые также могут двигаться под прямым углом к телу и имеют сегментированные щупики. Губа (нижняя губа) представляет собой сросшуюся структуру, движущуюся в продольном направлении и имеющую пару сегментированных щупиков. [ 24 ]

Легенда: а – усики
в – сложный глаз
фунт – губа
лр – верхняя губа
мд – нижние челюсти
мх – челюсти
Ротовой аппарат и остальная часть головы могут сочленяться как минимум в трех различных положениях: прогнатном, опистогнатном и гипогнатном. У видов с прогнатным сочленением голова расположена вертикально на одной линии с телом, например у видов Formicidae ; при гипогнатном типе голова расположена горизонтально и прилегает к телу. Опистогнатная голова расположена диагонально, как, например, у видов Blattodea и некоторых Coleoptera . [ 25 ] Ротовой аппарат сильно различается у насекомых разных отрядов, но есть две основные функциональные группы: мандибула и хвостовик. Ротовой аппарат Haustellate используется для всасывания жидкостей и может быть дополнительно классифицирован по наличию стилетов , которые включают колюще-сосущее, обтирание и сифонирование. Стилеты представляют собой игольчатые выступы, используемые для проникновения в ткани растений и животных. Стилеты и питательная трубка образуют видоизмененные нижние челюсти, верхнюю челюсть и гортаноглотку. [ 24 ]
- Нижнечелюстной ротовой аппарат, один из наиболее распространенных у насекомых, используется для откусывания и измельчения твердой пищи.
- Колюще-сосущий ротовой аппарат имеет стилеты и служит для проникновения в твердые ткани и последующего всасывания жидкой пищи.
- Губочный ротовой аппарат используется для высасывания и всасывания жидкостей и не имеет стилетов (например, у большинства двукрылых).
- В ротовом аппарате для сифонирования отсутствуют стилеты, они используются для всасывания жидкостей и обычно встречаются среди видов чешуекрылых .
Нижнечелюстной ротовой аппарат встречается у видов Odonata , взрослых Neuroptera , Coleoptera , Hymenoptera , Blattodea , Orthoptera и Lepidoptera . Однако у большинства взрослых чешуекрылых есть сифонирующий ротовой аппарат, а у их личинок (обычно называемых гусеницами ) есть челюсти .
мандибулировать
[ редактировать ]Верхняя губа представляет собой широкую лопасть, образующую крышу предротовой полости, подвешенную к наличнику перед ртом и образующую верхнюю губу. [ 1 ] : 22–24 На внутренней стороне он перепончатый и может образовывать срединную долю, эпиглотку , несущую некоторые сенсиллы . Верхняя губа поднимается от нижней челюсти двумя мышцами, выходящими на головку, и вставляется медиально в передний край верхней губы. Он частично закрыт от нижних челюстей двумя мышцами, возникающими в голове и прикрепляющимися к задне-боковым краям на двух небольших склеритах, тормах, и, по крайней мере у некоторых насекомых, резилиновой пружиной в кутикуле в месте соединения нижней челюсти. губа с наличником. [ 26 ] До недавнего времени считалось, что верхняя губа связана с первым сегментом головы. Однако недавние исследования эмбриологии, экспрессии генов и иннервации верхней губы показывают, что она иннервируется тритоцеребрумом головного мозга, который представляет собой сросшиеся ганглии третьего сегмента головы. Он образуется в результате слияния частей пары предковых придатков, обнаруженных на третьем сегменте головы, что свидетельствует об их взаимосвязи. [ 1 ] : 22–24 Его вентральная, или внутренняя, поверхность обычно мембранозная и образует лопастный эпифаринкс, несущий механосенсиллы и хемосенсиллы. [ 27 ] [ 28 ]
Жевательные насекомые имеют две челюсти, по одной с каждой стороны головы. Нижние челюсти расположены между верхней губой и верхней челюстью . Жвалы режут и измельчают пищу и могут использоваться для защиты; как правило, у них есть апикальный режущий край, а более базальная область коренных зубов измельчает пищу. Они могут быть чрезвычайно твердыми (около 3 по шкале Мооса или твердостью при вдавливании около 30 кг/мм). 2 ); таким образом, многие термиты и жуки без труда просверливают фольгу, сделанную из таких распространенных металлов, как медь, свинец, олово и цинк. [ 1 ] : 22–24 Режущие кромки обычно укрепляются добавлением цинка, марганца или, реже, железа в количестве примерно до 4% от сухого веса. [ 27 ] Обычно это самые крупные ротовые части жевательных насекомых, которые используются для пережевывания (разрезания, разрывания, раздавливания, пережевывания) пищевых продуктов. Они открываются наружу (в стороны головы) и сходятся медиально. У плотоядных жевательных насекомых челюсти могут быть изменены и стать более ножевидными, тогда как у травоядных жевательных насекомых они обычно широкие и плоские на противоположных сторонах (например, у гусениц ). У самцов жуков-оленей нижние челюсти модифицированы до такой степени, что не выполняют никакой функции питания, а вместо этого используются для защиты мест спаривания от других самцов. У муравьев жвалы также выполняют защитную функцию (особенно у каст солдат). У муравьев -быков челюсти удлиненные и зубчатые, используемые в качестве охотничьих (и защитных) придатков. [ нужна ссылка ]
Расположенные под нижними челюстями парные верхнечелюстные кости манипулируют пищей во время жевания . Максиллы могут иметь волосы и «зубы» по внутреннему краю. На внешнем крае галея представляет собой чашевидную или ковшовую структуру, которая располагается над внешним краем губы. У них также есть щупики , которые используются для определения характеристик потенциальной пищи. Максиллы занимают боковое положение, по одной с каждой стороны головы позади нижних челюстей. Проксимальная часть верхней челюсти состоит из базального кардо, имеющего одно сочленение с головкой, и плоской пластинки — ножки, шарнирно прикрепленной к кардо . И кардо, и ножки свободно соединены с головой перепонкой, поэтому способны двигаться. Дистально на ножках расположены две доли: внутренняя пластинка и наружная галея, одна или обе из которых могут отсутствовать. Сбоку на ножках находится членистый ножкообразный щупик, состоящий из множества сегментов; у прямокрылых их пять. Передние и задние мышцы-вращатели прикрепляются к кардо, а вентральные приводящие мышцы, возникающие на намете, прикрепляются как к кардо, так и к ножке. От ножки отходят мышцы-сгибатели пластинчатой и голеевой пластинок, а в черепе возникает еще один слезный сгибатель, но ни пластинчатая пластинка, ни голея не имеют мышц-разгибателей. На ножках щупика имеются мышцы-леватора и депрессора, причем каждый сегмент щупика имеет по одной мышце, вызывающей сгибание следующего сегмента. [ 26 ]
В ротовом аппарате нижней челюсти губа представляет собой четвероногое образование, хотя она образована двумя сросшимися вторичными максиллами. Его можно охарактеризовать как дно рта. С помощью верхних челюстей он помогает манипулировать пищей во время жевания или жевания или, в необычном случае нимфы стрекозы , вытягивается, чтобы подхватить добычу обратно к голове, где челюсти могут ее съесть. Губа по строению аналогична верхней челюсти , но придатки двух сторон срослись по средней линии и образуют срединную пластинку. Базальная часть губы, эквивалентная верхнечелюстным кардиналам и, возможно, включающая часть грудины губного сегмента, называется постментумом. Его можно разделить на проксимальный субментум и дистальный ментум. Дистальнее постментума и эквивалентно сросшимся верхнечелюстным ножкам находится прементум. Прементум закрывает предротовую полость сзади. На конце он имеет четыре доли, две внутренние язычки и две наружные параглоссы, которые вместе известны как язычка. Одна или обе пары долей могут отсутствовать или они могут сливаться, образуя единый срединный отросток. С каждой стороны прементума отходят щупики, часто трехчлениковые. [ 26 ]
Гипофаринкс — срединная доля непосредственно позади рта, выступающая вперед из задней части предротовой полости; это доля неопределенного происхождения, но, возможно, связанная с нижнечелюстным сегментом; [ 26 ] у аптеригот, уховерток и нимфальных поденок гипофаринкс несет пару боковых долей, суперлингвов (единственное число: суперлингва). Он делит полость на спинной пищевой мешок, или цибарий, и вентральный слюнный пузырь, в который открывается слюнный проток. [ 1 ] : 22–24 Обычно его можно найти слитым с либием. [ 27 ] Большая часть гипофаринкса перепончатая, но адоральная поверхность дистально склеротизирована и проксимально содержит пару подвесных склеритов, идущих вверх и заканчивающихся в латеральной стенке стомодеума. К этим склеритам прикрепляются мышцы, возникающие на лбу, которые дистально шарнирно прикреплены к паре язычных склеритов. Они, в свою очередь, прикрепили к ним антагонистические пары мышц, возникающие на намете и губах. Различные мышцы служат для поворота гипоглотки вперед и назад, а у таракана еще две мышцы пересекают гортаноглотку и расширяют слюнное отверстие и расширяют слюну. [ 26 ]
- Разнообразие нижних челюстей
-

-

-

-

-

-

-








Пирсинг-сосание
[ редактировать ]Ротовой аппарат может выполнять несколько функций. Некоторые насекомые сочетают колющие части с губчатыми, которые затем используются для прокалывания тканей растений и животных. Самки комаров питаются кровью ( гемофагами ), что делает их переносчиками болезней. Ротовой аппарат комара состоит из хоботка, парных челюстей и максилл. Верхняя челюсть образует игольчатые структуры, называемые стилетами , которые окружены губами. При укусе комара верхние челюсти проникают в кожу и фиксируют ротовой аппарат, позволяя таким образом вставить другие части. Похожая на ножны губа отодвигается назад, а оставшиеся ротовые части проходят через ее кончик в ткани. Затем через гортаноглотку комар впрыскивает слюну , содержащую антикоагулянты, препятствующие свертыванию крови. И, наконец, верхняя губа (верхняя губа) используется для всасывания крови. Для видов рода Anopheles характерны длинные щупики (две части с расширяющимся концом), почти достигающие конца верхней губы. [ 29 ]
- Разнообразие ротовых аппаратов для пирсинга
-

-

-

-
 слепень (самка)
слепень (самка) -






перекачивание
[ редактировать ]Хоботок и представляет собой образован из верхнечелюстных косточек приспособление некоторых насекомых для сосания. [ 30 ] Мышцы кибария или глотки сильно развиты и образуют насос. У полужесткокрылых и многих двукрылых, которые питаются жидкостями растений или животных, некоторые компоненты ротового аппарата модифицированы для прокалывания, а удлиненные структуры называются стилетами. Комбинированные трубчатые структуры называются хоботками, хотя в некоторых группах используется специализированная терминология.
У видов чешуекрылых он состоит из двух трубок, скрепленных крючками и отделяемых для чистки. Каждая трубка вогнута внутрь, образуя центральную трубку, через которую всасывается влага. На всасывание влияет сокращение и расширение мешка в голове. [ 31 ] Хоботок свернут под головой, когда насекомое находится в состоянии покоя, и выдвигается только во время кормления. [ 30 ] Верхнечелюстные щупики редуцированы или даже рудиментарны. [ 32 ] В некоторых базальных семействах они заметны, пятичлениковые и часто складчатые. [ 8 ] Форма и размеры хоботка изменились, чтобы обеспечить различным видам более широкий и, следовательно, более выгодный рацион. [ 30 ] Существует аллометрическая зависимость между массой тела чешуекрылых и длиной хоботка. [ 33 ] интересным адаптивным отклонением от него является необычно длинноязычный бражник Xanthopan morganii praedicta . Чарльз Дарвин предсказал существование и длину хоботка этой бабочки до ее открытия, основываясь на своих знаниях о мадагаскарской звездчатой орхидее Angraecum sesquipedale с длинными шпорами . [ 34 ]
- Сифонирующий ротовой аппарат
-

-

-

-





Обтирание
[ редактировать ]Ротовой аппарат насекомых, питающихся жидкостью, видоизменяется различными способами, образуя трубку, через которую в рот может втягиваться жидкость, и обычно другую, через которую проходит слюна. Мышцы кибария или глотки сильно развиты и образуют насос. [ 26 ] У некусающих мух нижние челюсти отсутствуют, а другие структуры редуцированы; губные щупики видоизменились и образовали губу, а верхнечелюстные щупики присутствуют, хотя иногда и короткие. У Brachycera губка особенно заметна и используется для протирания жидкой или полужидкой пищи. [ 35 ] Лабеллы представляют собой сложную структуру, состоящую из множества борозд, называемых псевдотрахеями, которые впитывают жидкость. Слюнные выделения из лабеллы помогают растворять и собирать частицы пищи, чтобы они могли легче поглощаться псевдотрахеями или откладывать яйца в подходящую среду; Считается, что это происходит за счет капиллярного действия. Жидкая пища затем выводится из псевдотрахеи через пищевой канал в пищевод . [ 36 ]
Ротовой аппарат пчел жевательного и прижимно-сосущего типа. Притирание — это способ кормления, при котором жидкая или полужидкая пища, прилипшая к выступающему органу или «языку», переносится из субстрата в рот. У медоносной пчелы ( Hymenoptera : Apidae : Apis mellifera ) удлиненные и сросшиеся губные язычки образуют волосатый язык, который окружен верхнечелюстными щитками и губными щупиками, образуя трубчатый хоботок, содержащий пищевой канал. При кормлении язык окунают в нектар или мед, который прилипает к волоскам, а затем втягивают, так что приставшая жидкость переносится в пространство между галеями и губными щупиками. Это глоссальное движение вперед и назад происходит неоднократно. Движение жидкости ко рту происходит в результате действия сибариального насоса, чему способствует каждое втягивание языка, выталкивающее жидкость вверх по пищевому каналу либо для удовлетворения потребностей в питании, либо для получения подходящей среды для откладки яиц. [ 1 ] : 22–24
- Обтирание ротового аппарата
-

-

-




грудная клетка
[ редактировать ]насекомых Грудная клетка состоит из трех сегментов: переднегруди , среднегруди и заднегруди . Передний ; сегмент, ближайший к голове, — переднегруди его основными особенностями являются первая пара ног и переднеспинка . Средний сегмент — среднегруди; его основными особенностями являются вторая пара ног и передние крылья, если таковые имеются. Третий, задний , грудной сегмент, примыкающий к брюшку, — заднегруди, несущий третью пару ног и задние крылья. Каждый сегмент ограничен межсегментарным швом. Каждый сегмент имеет четыре основных региона. Дорсальная поверхность называется тергумом (или нотумом, в отличие от терги брюшка). [ 1 ] : 22–24 Две латеральные области называются плеврой (единственное число: плеврон), а вентральная часть называется грудиной. В свою очередь, нотум переднегруди называется переднеспинкой, нотум среднегруди — среднегруди, а нотум заднегруди — заднеспинкой. Продолжая эту логику, существуют также мезоплевры и метаплевры, а также мезостернум и заднегруди. [ 8 ]
Тергальные пластинки грудной клетки представляют собой простые структуры у аптеригот и многих неполовозрелых насекомых, но у крылатых взрослых особей они модифицированы по-разному. Каждая крылоторакальная нота состоит из двух основных отделов: переднего, несущего крылья, алинотума и заднего, несущего фрагму, постнотума. Фрагматы (единственное число: фрагма) представляют собой пластинчатые аподемы, которые простираются внутрь ниже антекостальных швов, отмечая первичные межсегментарные складки между сегментами; phragmata обеспечивают прикрепление мышц продольного полета. Каждый алинотум (иногда ошибочно называемый «нотум») может быть пересечен швами, которые отмечают положение внутренних укрепляющих гребней и обычно делят пластинку на три области: переднюю прескутум, щиток и меньший задний щиток. Считается, что латеральные плевральные склериты происходят из субкоксального сегмента ноги предкового насекомого. Эти склериты могут быть отдельными, как у чешуйницы, или слитыми в почти сплошной склеротический участок, как у большинства крылатых насекомых. [ 1 ] : 22–24
Переднегрудь
[ редактировать ]Переднеспинка переднегруди может быть простой по строению и небольшой по сравнению с остальными нотами, но у жуков, богомолов, многих клопов и некоторых прямокрылых переднеспинка расширена, а у тараканов она образует щиток, закрывающий часть голова и среднегруди. [ 8 ] [ 1 ] : 22–24
Птероторакс
[ редактировать ]Поскольку среднегруди и заднегруди удерживают крылья, они имеют общее название птерорторакс (pteron = крыло). Переднее крыло, которое в разных отрядах носит разные названия (например, надкрылья у прямокрылых и надкрылья у жесткокрылых), возникает между мезонотумом и мезоплеврой, а заднее крыло сочленяется между заднеспинкой и метаплеврой. Ножки возникают из мезоплевр и метаплевр. Среднегруди и заднегруди имеют плевральный шов (мезоплевральный и метаплевральный швы), который проходит от основания крыла до тазика ноги. Склерит перед плевральным швом называется эпистернумом (последовательно мезэпистернум и метэпистернум). Склерит позади шва называется эпимироном (последовательно мезепимирон и метепимирон). Дыхальца, внешние органы дыхательной системы, расположены на птеротораксе, обычно между про- и мезоплеврой, а также между мезо- и метаплеврами. [ 8 ]
Вентральный вид или грудина следует тому же соглашению: переднегруди находится под переднегруди, среднегруди под среднегруди и заднегруди под заднегруди. Нотум, плевра и грудина каждого сегмента имеют множество различных склеритов и швов, сильно различающихся от порядка к порядку, и они не будут подробно обсуждаться в этом разделе. [ 8 ]
Крылья
[ редактировать ]Большинство филогенетически продвинутых насекомых имеют две пары крыльев , расположенные на втором и третьем грудных сегментах. [ 1 ] : 22–24 Насекомые — единственные беспозвоночные , развившие способность к полету, и это сыграло важную роль в их успехе. Полет насекомых не очень хорошо изучен, поскольку он основан на турбулентных аэродинамических эффектах. Примитивные группы насекомых используют мышцы, действующие непосредственно на строение крыла. У более продвинутых групп, составляющих Neoptera , крылья складные, а их мышцы воздействуют на стенку грудной клетки и косвенно приводят в движение крылья. [ 1 ] : 22–24 Эти мышцы могут сокращаться несколько раз для каждого отдельного нервного импульса, позволяя крыльям биться быстрее, чем это обычно возможно.
Полет насекомых может быть быстрым, маневренным и универсальным, возможно, благодаря изменяющейся форме, исключительному контролю и переменному движению крыльев насекомых. Отряды насекомых используют разные механизмы полета; например, полет бабочки можно объяснить с помощью устойчивой, непереходной аэродинамики и тонкого профиля теории .

Внутренний
[ редактировать ]Каждое из крыльев состоит из тонкой перепонки, поддерживаемой системой жилок. Мембрана образована двумя плотно прилегающими слоями покровов, в то время как вены образуются там, где два слоя остаются отдельными, а кутикула может быть более толстой и более сильно склеротизированной. Внутри каждой из крупных вен имеется нерв и трахея, и, поскольку полости вен соединены с гемоцелем , гемолимфа может оттекать в крылья. [ 26 ] По мере развития крыла дорсальный и вентральный покровные слои на большей части своей площади плотно прилегают друг к другу, образуя перепонку крыла. Остальные участки образуют каналы, будущие вены, в которых могут располагаться нервы и трахеи. Кутикула, окружающая жилки, утолщается и сильно склеротизируется, что придает крылу прочность и жесткость. На крыльях могут встречаться волоски двух типов: микротрихии, мелкие и неравномерно разбросанные, и макротрихии, более крупные, гнездовые, могут ограничиваться жилками. Чешуя Lepidoptera и Trichoptera представляет собой сильно модифицированные макротрихии. [ 27 ]
Вены
[ редактировать ]
У некоторых мелких насекомых жилкование может быть редуцировано. Например, у хальцидоидных ос имеются только подкоста и часть лучевой кости. И наоборот, увеличение жилкования может происходить за счет разветвления существующих жилок с образованием добавочных жилок или за счет развития дополнительных вставочных жилок между исходными, как в крыльях прямокрылых (кузнечиков и сверчков). У некоторых насекомых имеется большое количество поперечных жилок, которые могут образовывать сетку, как на крыльях Odonata (стрекозы и стрекозы), так и в основании передних крыльев Tettigonioidea и Acridoidea (катидиды и кузнечики соответственно). [ 26 ]
Архедиктион — название гипотетической схемы жилкования крыльев, предложенной для самого первого крылатого насекомого. Он основан на сочетании предположений и ископаемых данных. Поскольку считается, что все крылатые насекомые произошли от общего предка, архедиктия представляет собой «шаблон», который изменялся (и оптимизировался) естественным отбором в течение 200 миллионов лет. Согласно нынешним догматам, архедиктион содержал от шести до восьми продольных жилок. Эти вены (и их ответвления) названы в соответствии с системой, разработанной Джоном Комстоком и Джорджем Нидхэмом, — системой Комстока-Нидхэма: [ 37 ]
- Коста (C) – передняя кромка крыла
- Subcosta (Sc) - вторая продольная жилка (за ребром), обычно неразветвленная.
- Радиус (R) – третья продольная жилка, до края крыла доходят от одной до пяти ветвей.
- Медиа (М) - четвертая продольная жилка, до края крыла доходят от одной до четырех ветвей.
- Cubitus (Cu) - пятая продольная жилка, до края крыла доходит от одной до трех ветвей.
- Анальные вены (А1, А2, А3) – неразветвленные вены позади локтевой кости.
Коста (С) — ведущая краевая жилка у большинства насекомых, хотя над ребром иногда встречается небольшая жилка — прекоста. Практически у всех современных насекомых [ 1 ] : 41–42 прекоста слита с костой; ребра редко разветвляются, поскольку находятся на переднем крае, который у основания связан с плечевой пластинкой. Трахея реберной вены, возможно, является ветвью подреберной трахеи. После ребра расположена третья вена, подкоста, которая разветвляется на две отдельные вены: переднюю и заднюю. Основание подкосты связано с дистальным концом шейки первой подмышечной кости. Четвертая жилка — лучевая кость, которая разветвляется на пять отдельных жилок. Радиус обычно является самой сильной жилкой крыла. Ближе к середине крыла он раздваивается на первую неделимую ветвь (R1) и вторую ветвь, называемую радиальным сектором (Ra), которая дихотомически подразделяется на четыре дистальные ветви (R2, R3, R4, R5). В основании лучевая кость гибко соединена с передним концом второй подмышечной кости (2Ах). [ 38 ]
Пятая жилка крыла — СМИ. В архетипическом паттерне (А) медиа разветвляется на две основные ветви: переднюю медиану (МА), которая делится на две дистальные ветви (МА1, МА2), и срединный сектор, или медиа-заднюю (МП), которая имеет четыре концевые ветви (М1, М2, М3, М4). У большинства современных насекомых передняя медиа утеряна, и привычной «медией» является четырехветвистая медиа-задняя с общим базальным стеблем. У Ephemerida, согласно современным интерпретациям жилкования крыльев, сохраняются обе ветви сред, тогда как у Odonata сохранившимися средами являются примитивная передняя ветвь. Ствол медиальной кости часто соединяется с лучевой костью, но когда она встречается в виде отдельной вены, ее основание связано с дистальной срединной пластинкой (m') или непрерывно склеротизировано с последней. Кубитус, шестая жилка крыла, преимущественно двуветвистый. Первичное разветвление происходит возле основания крыла, образуя две основные ветви (Cu1, Cu2). Передняя ветвь может распадаться на несколько второстепенных ветвей, но обычно она раздваивается на две дистальные ветви. Вторая ветвь кубитуса (Cu2) у перепончатокрылых, трихоптер и чешуекрылых была ошибочно принята Комстоком и Нидхэмом за первый анальный. Проксимально основной стебель кубитуса связан с дистальной срединной пластинкой (m') основания крыла. [ 38 ]
Посткубитус (Pcu) — первый анальный элемент системы Комстока и Нидэма. Посткубитус, однако, имеет статус самостоятельной жилки крыла и должен быть признан таковым. В нимфальных крыльях ее трахея возникает между кубитальной трахеей и группой ванальных трахей. В зрелых крыльях более генерализованных насекомых посткубитус всегда связан проксимально с кубитом и никогда не связан тесно со склеритом-сгибателем (3Ax) основания крыла. У Neuroptera, Mecoptera и Trichoptera посткубитус может быть более тесно связан с ванальными жилками, но его основание всегда свободно от последних. Посткубит обычно неразветвленный; примитивно он двуветвистый. Ваннальные вены (от lV до nV) представляют собой анальные вены, непосредственно связанные с третьей подмышечной веной, и на них напрямую влияет движение этого склерита, вызывающее сгибание крыльев. Число ваннанных жилок варьирует от одной до 12 в зависимости от расширения ванальной области крыла. Ваннальные трахеи обычно возникают из общего ствола трахеи у нимфальных насекомых, а вены рассматриваются как ветви одной анальной вены. Дистально ванальные вены либо простые, либо разветвленные. Скуловая вена (J) скуловой доли крыла часто занята сетью неправильных жилок или может быть целиком перепончатой; иногда он содержит одну или две отдельные мелкие вены: первую скуловую вену, или дуговую вену, и вторую скуловую вену, или кардинальную вену (2J). [ 38 ]
- Перекрестные вены C-Sc - проходят между ребром и подкостой.
- R поперечные жилки – проходят между соседними ветвями лучевой кости.
- Поперечные вены RM – проходят между лучевой и медиальной поверхностью.
- Поперечные жилки M-Cu - проходят между медией и локтевой пластинкой.
Все жилки крыла подвержены вторичному разветвлению и слиянию поперечными жилками. У некоторых отрядов насекомых поперечные жилки столь многочисленны, что весь жилковый рисунок представляет собой плотную сеть ветвящихся жилок и поперечных жилок. Однако обычно встречается определенное количество поперечных жилок, имеющих определенное расположение. Более постоянными поперечными венами являются плечевая поперечная вена (h) между ребром и подкостой, радиальная поперечная вена (r) между R и первой вилкой Rs, секториальная поперечная вена (s) между двумя развилками. R8, срединная поперечная вена (мм) между M2 и M3 и медиально-кубитальная поперечная вена (m-cu) между медией и локтевой пластинкой. [ 38 ]
Жилки крыльев насекомых характеризуются выпукло-вогнутым расположением, как у подёнок (т. е. вогнутость — «вниз», выпуклость — «вверх»), которые регулярно чередуются и разветвляются; всякий раз, когда вена разветвляется, между двумя ветвями всегда имеется интерполированная жилка противоположного положения. Вогнутая жилка разветвляется на две вогнутые жилки (при этом интерполированная жилка становится выпуклой), при этом правильная смена жилок сохраняется. [ 39 ] Жилки крыла имеют волнообразную форму в зависимости от того, имеют ли они тенденцию складываться вверх или вниз, когда крыло расслаблено. Базальные стволы вен выпуклые, но каждая вена дистально разветвляется на переднюю выпуклую ветвь и заднюю вогнутую ветвь. Таким образом, ребра и подкосты рассматриваются как выпуклые и вогнутые ветви первичной первой вены, Rs — вогнутая ветвь лучевой кости, posterior media — вогнутая ветвь средней жилки, Cu1 и Cu2 — соответственно выпуклая и вогнутая, а примитивные postcubitus и первый vannal имеют переднюю выпуклую ветвь и заднюю вогнутую ветвь. Выпуклая или вогнутая природа жилок использовалась в качестве доказательства при определении идентичности сохранившихся дистальных ветвей жилок современных насекомых, но не было продемонстрировано, что она одинакова для всех крыльев. [ 26 ] [ 38 ]
Поля
[ редактировать ]
Области крыльев разграничены и разделены линиями сгиба , по которым крылья могут складываться, и линиями сгиба , которые сгибаются во время полета. Между линиями сгиба и линиями сгиба фундаментальное различие часто размыто, поскольку линии сгиба могут обеспечивать некоторую гибкость или наоборот. Двумя константами, обнаруженными почти у всех крыльев насекомых, являются ключичная (линия сгибания) и скуловая складки (или линия сгиба), образующие изменчивые и неудовлетворительные границы. Складывание крыльев может быть очень сложным: поперечная складчатость наблюдается на задних крыльях Dermaptera и Coleoptera, а у некоторых насекомых анальная область может быть сложена веером. [ 1 ] : 41–42 Четыре различных поля, обнаруженные на крыльях насекомых:
- Гребля
- Анальная область (ваннус)
- Югальная область
- Подмышечная область
- Они непростые
Большинство вен и поперечных вен встречается в передней области ремигиума , который отвечает за большую часть полета, приводимого в движение грудными мышцами. Заднюю часть ремигия иногда называют клавусом; два других задних поля — это анальная и скуловая области . [ 1 ] : 41–42 Когда ваннанная складка имеет обычное положение кпереди от группы анальных вен, в ремигиуме присутствуют реберные, подреберные, радиальные, медиальные, кубитальные и посткубитальные вены. В согнутом крыле ремигий поворачивается кзади на гибком базальном соединении лучевой кости со второй подмышечной, а основание медио-кубитального поля сложено медиально на подмышечной области по plica basalis (bf) между срединными пластинками (m, м') основания крыла. [ 38 ]
Ваннус ограничен ванальной складкой, которая обычно возникает между посткубитусом и первой ванальной жилкой. У прямокрылых он обычно занимает такое положение. Однако у переднего крыла Blattidae единственная складка в этой части крыла лежит непосредственно перед посткубитусом. У Plecoptera ванна складка находится позади посткубитуса, но проксимальнее пересекает основание первой ванальной жилки. У цикады ваннанная складка лежит сразу за первой ванальной жилкой (lV). Эти небольшие изменения в фактическом положении ванальной складки, однако, не влияют на единство действия ваннанных жилок, контролируемых сгибателем склерита (3Ах), при сгибании крыла. На задних крыльях большинства прямокрылых вторичная разделяющая вена образует ребро в ванальной складке. Ваннус обычно имеет треугольную форму, а его вены обычно расходятся от третьей подмышечной вены, как веерные ребра. Некоторые из венальных вен могут быть разветвленными, а вторичные жилки могут чередоваться с первичными. Ваннальная область обычно лучше всего развита в заднем крыле, где она может быть увеличена, образуя опорную поверхность, как у Plecoptera и Orthoptera. Большие веерообразные расширения задних крыльев Acrididae явно представляют собой ваннальные области, поскольку все их жилки опираются на третьи подмышечные склериты оснований крыльев, хотя Мартынов (1925) относит большую часть веерных областей у Acrididae к скуловым областям. крыльев. Истинная югума крыла саранчовых представлена лишь небольшой перепончатой (Ju) месадой последней ваннанной жилки. Югум более развит у некоторых других прямокрылых, например у Mantidae. У большинства высших насекомых с узкими крыльями ванна редуцируется и ванна складка теряется, но даже в таких случаях согнутое крыло может изгибаться по линии между послекубитусом и первой ванальной жилкой. [ 38 ]
Скуловая область, или neala, представляет собой область крыла, которая обычно представляет собой небольшую перепончатую область проксимальнее основания ваннуса, укрепленную несколькими небольшими неравномерными венозными утолщениями; но когда он хорошо развит, он представляет собой отдельную часть крыла и может содержать одну или две скуловые жилки. Когда скуловая область переднего крыла развивается как свободная лопасть, она выступает под плечевой угол заднего крыла и, таким образом, служит для соединения двух крыльев вместе. В группе Jugatae чешуекрылых он имеет длинную пальцеобразную лопасть. Скуловую область назвали неалой («новое крыло»), потому что это второстепенная и недавно развившаяся часть крыла. [ 38 ]
Вспомогательная область, содержащая аксиллярные склериты, имеет в целом форму разностороннего треугольника. Основание треугольника (ab) — шарнир крыла с корпусом; вершина (в) — дистальный конец третьего подмышечного склерита; более длинная сторона находится впереди вершины. Точка d на передней стороне треугольника отмечает место сочленения лучевой вены со вторым подмышечным склеритом. Линия между d и c представляет собой базальную складку (bf) или складку крыла у основания медиокубитального поля. [ 38 ]
В заднем углу основания крыла у некоторых двукрылых имеется пара перепончатых лопастей (чешуек или чашелистиков), известных как алулы. Алула у комнатной мухи хорошо развита. Наружная чешуя (в) возникает от основания крыла позади третьего подмышечного склерита (3Ах) и представляет собой скуловую долю других насекомых (А, D); более крупная внутренняя чешуя (d) начинается от заднего щиткового края тергума крылатого сегмента и образует защитный капюшонообразный навес над недоуздоком. В согнутом крыле наружная чешуя alula перевернута вверх дном над внутренней чешуей, причем последняя не затрагивается движением крыла. У многих двукрылых глубокий разрез анальной области перепонки крыла позади единственной ванальной вены выделяет проксимальную крыловую лопасть, дистальнее внешней чешуи алулы. [ 38 ]
Суставы
[ редактировать ]
Различные движения крыльев, особенно у насекомых, которые в состоянии покоя сгибают крылья горизонтально над спиной, требуют более сложной суставной структуры у основания крыла, чем простое соединение крыла с телом. Каждое крыло прикреплено к телу перепончатой базальной областью, но суставная перепонка содержит несколько небольших суставных склеритов, известных под общим названием птералии. Птералии включают переднюю плечевую пластинку у основания реберной вены, группу подмышечных желез (Ax), связанных с подреберной, радиальной и ванной венами, и две менее выраженные срединные пластинки (m, m') у основания реберной вены. медиокубитальная зона. Подмышечные клетки специально развиты только у насекомых, сгибающих крылья, где они составляют сгибательный механизм крыла, управляемый мышцей-сгибателем, возникающей на плевронах. Для основания крыла характерна также небольшая лопасть на переднем крае суставной области проксимальнее плечевой пластинки, которая у передних крыльев некоторых насекомых развита в крупный плоский чешуевидный лоскут — тегулу, перекрывающий друг друга. основание крыла. Сзади суставная перепонка часто образует обширную лопасть между крылом и телом, а ее край обычно утолщен и гофрирован, создавая вид связки, так называемого подмышечного тяжа, продолжающегося мезиально с задней краевой щитичной складкой тергальная пластинка, несущая крыло. [ 38 ]
Суставные склериты, или птералии основания крыльев крылатых насекомых и их взаимоотношения с телом и жилками крыла, показанные схематически, следующие:
- Плечевые пластины
- Первая подмышечная
- Вторая подмышечная
- Третья подмышечная
- Четвертая подмышечная
- Срединные пластины ( м , м' )
Плечевая пластинка обычно представляет собой небольшой склерит на переднем крае основания крыла, подвижный и сочлененный с основанием реберной вены. У Odonata плечевые пластинки сильно увеличены. [ 38 ] с двумя мышцами, отходящими от эпистерна, вставленными в плечевые пластинки, и двумя от края эпимерона, вставленными в подмышечную пластинку. [ 26 ]
Первый подмышечный склерит (lAx) — передняя шарнирная пластинка основания крыла. Его передняя часть опирается на передний нотальный крыльевой отросток тергума (ANP); его задняя часть сочленяется с тергальным краем. Передний конец склерита обычно представляет собой тонкое плечо, вершина которого (д) всегда связана с основанием подреберной вены (Sc), но не соединена с последней. Тело склерита латерально сочленяется со второй подмышечной впадиной. Второй аксиллярный склерит (2Ах) более изменчив по форме, чем первый, но его механические связи не менее определенны. Она наклонно шарнирно прикреплена к наружному краю тела первой подмышечной вены, а к ее переднему концу (г) всегда гибко прикрепляется лучевая вена (R). Вторая подмышечная мышца имеет как дорсальную, так и вентральную склеротизацию в основании крыла; его вентральная поверхность опирается на опорный крыльевой отросток плевры. Таким образом, вторая подмышечная кость представляет собой основной склерит основания крыла и конкретно управляет радиальной жилкой. [ 38 ]
Третий подмышечный склерит (3Ах) лежит в задней части суставной области крыла. Форма его весьма разнообразна и часто неправильная, но третья подмышечная кость представляет собой склерит, к которому прикрепляется мышца-сгибатель крыла (D). Мезально он сочленяется спереди (е) с задним концом второй подмышечной кости, а сзади (б) с задним крыльевым отростком тергума (ПНП) или с небольшой четвертой подмышечной веткой, если последняя имеется. Дистально третья подмышечная артерия удлинена, образуя отросток, всегда связанный с основаниями группы жилок анальной области крыла, называемой здесь ванальными жилками (V). Таким образом, третья подмышечная мышца обычно представляет собой заднюю шарнирную пластинку основания крыла и является активным склеритом сгибательного механизма, который непосредственно управляет ванальными венами. Сокращение мышцы-сгибателя (D) вращает третью подмышечную мышцу в ее мезальных сочленениях (b, f) и тем самым поднимает ее дистальное плечо; это движение вызывает сгибание крыла. Четвертый аксиллярный склерит не является постоянным элементом основания крыла. Если он присутствует, то обычно представляет собой небольшую пластинку, находящуюся между третьей подмышечной впадиной и задним отростком нотального крыла и, вероятно, представляет собой отдельную часть последнего. [ 38 ]
Срединные пластинки (m, m') также представляют собой склериты, которые не так четко дифференцированы как специфические пластинки, как три основные подмышечные кости, но являются важными элементами сгибательного аппарата. Они лежат в срединной области основания крыла дистальнее второй и третьей подмышечных костей и отделены друг от друга косой линией (bf), образующей при сгибании крыла выраженную выпуклую складку. Проксимальная пластинка (m) обычно прикрепляется к дистальному плечу третьей подмышечной впадины и, возможно, ее следует рассматривать как часть последней. Дистальная пластинка (m') менее постоянно представлена в виде отчетливого склерита и может быть представлена общей склеротизацией основания медио-кубитального поля крыла. Когда вены этой области различимы у основания, они связаны с наружной срединной пластинкой. [ 38 ]
Сцепление, складывание и другие функции
[ редактировать ]У многих видов насекомых переднее и заднее крыло соединены вместе, что повышает аэродинамическую эффективность полета. Наиболее распространенный механизм сцепления (например, у Hymenoptera и Trichoptera ) представляет собой ряд маленьких крючков на переднем крае заднего крыла, или «хамули», которые фиксируются на переднем крыле, удерживая их вместе (крючковидное соединение). У некоторых других видов насекомых (например, Mecoptera , Lepidoptera и некоторых Trichoptera ) скуловая лопасть переднего крыла покрывает часть заднего крыла (скуловое соединение), или края переднего и заднего крыла широко перекрываются (амплексиформное соединение), или щетинки заднего крыла, или уздечка, зацепляются под удерживающую структуру или сетчатку на переднем крыле. [ 1 ] : 43
В состоянии покоя у большинства насекомых крылья держат за спиной, что может включать продольное, а иногда и поперечное складывание перепонки крыла. Иногда по линиям сгибания может возникать складчатость. Хотя линии сгиба могут быть поперечными, как на задних крыльях жуков и уховерток, они обычно радиальны к основанию крыла, что позволяет сгибать соседние части крыла друг над другом или под друг другом. Наиболее распространенной линией сгиба является скуловая складка, расположенная сразу за третьей анальной веной. [ 27 ] хотя у большинства Neoptera есть скуловая складка сразу за жилкой 3А на передних крыльях. Иногда он присутствует и на задних крыльях. Там, где анальная область заднего крыла большая, как у прямокрылых и Blattodea, вся эта часть может загибаться под переднюю часть крыла по ванальной складке немного позади ключичной борозды. Кроме того, у прямокрылых и Blattodea анальное поле веерообразно сложено вдоль жилок, при этом анальные жилки выпуклые, на гребнях складок, а добавочные жилки вогнутые. Если ключичная борозда и скуловая складка, вероятно, гомологичны у разных видов, то положение ваннальной складки у разных таксонов различается. Складывание производится мышцей, возникающей на плевре и прикрепляющейся к третьему подмышечному склериту таким образом, что при его сокращении склерит поворачивается вокруг точек сочленения с задним нотальным отростком и вторым подмышечным склеритом. [ 26 ]
В результате дистальное плечо третьего подмышечного склерита вращается вверх и внутрь, так что, наконец, его положение полностью меняется на противоположное. Анальные жилки сочленены со склеритом таким образом, что при его движении они увлекаются за ним и изгибаются над спиной насекомого. Активность той же мышцы в полете влияет на выходную мощность крыла, поэтому она также важна для управления полетом. У ортоптероидных насекомых эластичность кутикулы приводит к сгибанию венальной области крыла вдоль жилок. Следовательно, энергия расходуется на раскрытие этой области, когда крылья перемещаются в полетное положение. В целом разгибание крыльев, вероятно, является результатом сокращения мышц, прикрепленных к базилярному склериту или, у некоторых насекомых, к субалярному склериту. [ 26 ]
Ноги
[ редактировать ]
Типичные и обычные сегменты ноги насекомых делятся на тазик, один вертел , бедренную кость, голень, лапку и предплюсну . Тазик в своей более симметричной форме имеет форму короткого цилиндра или усеченного конуса, хотя обычно он яйцевидный и может быть почти сферическим. Проксимальный конец тазика опоясан субмаргинальным базикостальным швом, который образует внутри гребень, или базикоста , и отграничивает краевой фланец, коксомаргинальный , или базикоксит . Базикоста ; укрепляет основание тазика и обычно расширяется на внешней стенке, чтобы обеспечить прикрепление мышц однако на мезальной половине тазика он обычно слабый и часто сливается с краем тазика. Вертельные мышцы, берущие начало в тазике, всегда прикрепляются к базикосте дистально. Тазик прикреплен к телу суставной перепонкой, кориумом тазика, которая окружает его основание. Эти два сочленения, возможно, являются основными дорсальной и вентральной суставными точками подтаксиально-тазового шарнира. Кроме того, тазик насекомых часто имеет переднее сочленение с передним, вентральным концом трохантина, однако вертельное сочленение не сосуществует с грудинным сочленением. Плевральная суставная поверхность тазика опирается на мезальный перегиб стенки тазика. Если тазик подвижен только на плевральном сочленении, суставная поверхность тазика обычно изогнута на достаточную глубину, чтобы обеспечить рычаг отводящих мышц, прикрепленных к внешнему краю основания тазика. Дистально тазик имеет переднее и заднее сочленение с вертелом. Наружная стенка тазика часто отмечена швом, идущим от основания до переднего вертельного сочленения. У некоторых насекомых коксальный шов совпадает с плевральным швом. В таких случаях тазик оказывается разделенным на две части, соответствующие эпистернуму и эпимерону плевры. Коксальный шов отсутствует у многих насекомых. [ 38 ] : 163–164
Перегиб стенки тазика, несущей плевральную суставную поверхность, делит латеральную стенку базоксита на предсуставную часть и послесуставную часть, причем эти две области часто выглядят как две краевые доли на основании тазика. Задняя доля обычно крупнее и называется мероном. Мерон может быть значительно увеличен за счет расширения дистально на задней стенке тазика; у Neuroptera, Mecoptera, Trichoptera и Lepidoptera мерон настолько велик, что тазик кажется разделенным на переднюю часть, так называемый «генуинный тазик», и мерон, но мерон никогда не включает в себя область задний вертельный сочленение, а ограничивающая его бороздка всегда входит в состав базикостального шва. Тазик с увеличенным мероном имеет внешний вид, похожий на тот, который разделен тазобедренным швом, совпадающим с плевральным швом, но эти два состояния принципиально совершенно различны, и их не следует путать. У двукрылых мерон доходит до крайней точки своего отклонения от обычного состояния. У некоторых более распространенных мух, например у Tipulidae, мерон средней ноги выглядит как большая лопасть тазика, выступающая вверх и назад от основания тазика; у высших представителей отряда он полностью отделяется от тазика и образует пластинку латеральной стенки среднегруди. [ 38 ] : 164
Вертел — базальный сегмент телоподита; это всегда небольшой сегмент ноги насекомого, свободно перемещающийся за счет горизонтального шарнира на тазике, но более или менее прикрепленный к основанию бедренной кости. При движении на бедре вертельная бедренная петля обычно вертикальна или наклонена в вертикальной плоскости, что обеспечивает небольшое движение производства и сокращения в суставе, хотя присутствует только приводящая мышца. У Odonata, как у нимф, так и у взрослых, имеются два вертельных членика, но они неподвижны друг на друге; второй содержит мышцу-редуктор бедренной кости. Таким образом, обычный одиночный вертельный сегмент насекомых, вероятно, представляет собой два вертела других членистоногих, слившиеся в один видимый сегмент, поскольку маловероятно, что первичный тазобедренный шарнир утрачен на ноге. У некоторых перепончатокрылых базальный отдел бедренной кости имитирует второй вертел, но прикрепление мышцы-редуктора к его основанию свидетельствует о принадлежности его к бедренному сегменту, поскольку, как показано на зубчатой ноге, переходник берет свое начало в настоящий второй вертел. [ 38 ] : 165
Бедренная кость — третий сегмент ноги насекомого, обычно самая длинная и сильная часть конечности, но ее размер варьируется от огромного заднего бедра прыгающих прямокрылых до очень маленького сегмента, который присутствует у многих личиночных форм. Объем бедренной кости обычно коррелирует с размером содержащихся в ней большеберцовых мышц, но иногда ее увеличивают и изменяют форму для других целей, помимо размещения большеберцовых мышц. У взрослых насекомых голень представляет собой тонкий сегмент , лишь немного короче бедренной кости или совокупности бедренной кости и вертела. Его проксимальный конец образует более или менее отчетливую головку, наклоненную к бедренной кости, устройство, позволяющее сгибать большеберцовую кость вплотную к нижней поверхности бедренной кости. [ 38 ] : 165
Термины пробедренная кость, мезобедренная кость и метабедренная кость относятся к бедрам передних, средних и задних ног насекомого соответственно. [ 40 ] Точно так же протибии, мезотибии и метаберцы относятся к голеням передних, средних и задних ног. [ 41 ]
Лапка насекомых соответствует предпоследнему сегменту генерализованной конечности членистоногих, который у ракообразных называется проподитом. У взрослых насекомых он обычно подразделяется на 2—5 подсегментов, или члеников, но у протуры, некоторых коллембол и большинства личинок голометаболических насекомых он сохраняет примитивную форму простого сегмента. Подсегменты лапок взрослого насекомого обычно свободно перемещаются друг относительно друга за счет изогнутых соединительных перепонок, но собственных мышц лапка никогда не имеет. Лапка взрослых крыловидных насекомых, имеющая менее пяти подсегментов, вероятно, специализирована за счет потери одного или нескольких подсегментов или за счет слияния соседних подсегментов. У лапок Acrididae длинный базальный кусок состоит из трех соединенных члеников, оставляя четвертый и пятый. Базальный предплюсник иногда заметно увеличен и называется базитарзусом. На нижней поверхности подсегментов лапок у некоторых прямокрылых имеются небольшие подушечки — пульвиллы лапок, или эуплантулы. У личинок насекомых лапка иногда сливается с голенью, образуя большеберцовый сегмент; в некоторых случаях он оказывается ликвидированным или редуцированным до зачатка между большеберцовой костью и претарзусом. [ 38 ] : 165–166
По большей части бедренная кость и голень являются самыми длинными сегментами ног, но различия в длине и прочности каждого сегмента зависят от их функций. Например, насекомые грессориального и бегущего типа, или шагающего и бегущего типа соответственно, обычно имеют хорошо развитые бедра и голени на всех ногах, тогда как прыгающие (скакующие) насекомые, такие как кузнечики, имеют непропорционально развитые метабедренные и метаберцовые кости. У водных жуков ( Coleoptera ) и клопов ( Hemiptera ) голени и/или лапки одной или нескольких пар ног обычно приспособлены для плавания (водоплавающие) с бахромой из длинных тонких волосков. У многих наземных насекомых, таких как медведки (Orthoptera: Gryllotalpidae ), нимфальные цикады (Hemiptera: Cicadidae ) и жуки-скарабеи ( Scarabaeidae ), голени передних ног (protibiae) увеличены и модифицированы для рытья (ископаемые), тогда как передние лапки некоторых хищных насекомых, например богомолов златоглазок ( Neuroptera ) и богомолы ( Mantodea ), специализируются на захвате добычи, или хищные. Голени и базальные лапки каждой задней ноги медоносных пчел модифицированы для сбора и переноса пыльцы. [ 26 ] : 45
Живот
[ редактировать ]Нижний план брюшка взрослого насекомого обычно состоит из 11–12 сегментов и менее сильно склеротизирован, чем голова или грудь. Каждый сегмент брюшка представлен склеротизованной тергумом, грудиной и, возможно, плевритом. Терги отделены друг от друга, а также от прилежащей грудины или плевры перепонкой. Дыхальца расположены в плевральной области. Вариант этого плана включает слияние терги или терги и грудины с образованием непрерывных спинных или вентральных щитков или конической трубки. У некоторых насекомых в плевральной области имеется склерит, называемый латеротергитом. Вентральные склериты иногда называют латеростернитами . На эмбриональной стадии многих насекомых и постэмбриональной стадии примитивных насекомых имеется 11 брюшных сегментов. У современных насекомых наблюдается тенденция к уменьшению числа брюшных сегментов, но примитивное число — 11 — сохраняется в ходе эмбриогенеза. Вариабельность числа брюшных сегментов значительна. Если считать Apterygota ориентиром для крыловидных, то царит путаница: у взрослых Protura 12 сегментов, у Collembola - 6. У прямокрылых семейства Acrididae 11 сегментов, а у ископаемых экземпляров Zoraptera брюшко из 10 сегментов. [ 8 ]
Как правило, первые семь брюшных сегментов взрослых особей (прегенитальные сегменты) сходны по строению и не имеют придатков. Однако у аптеригот (щетинохвосты и чешуйницы) и у многих неполовозрелых водных насекомых имеются брюшные придатки. Аптериготы обладают парой стилей; рудиментарные придатки, серийно гомологичные дистальной части грудных ног. Мезально — одна или две пары выдающихся (или выступающих) пузырьков, по крайней мере, на некоторых брюшных сегментах. Эти пузырьки происходят из коксальных и вертельных эндитов (внутренних кольчатых долей) предковых брюшных придатков. Водные личинки и нимфы могут иметь жабры по бокам от некоторых до большинства брюшных сегментов. [ 1 ] : 49 Остальные брюшные сегменты состоят из репродуктивной и анальной частей.
Наружные половые органы
[ редактировать ]
Органы, отвечающие за спаривание и отложение яиц, известны под общим названием наружные половые органы, хотя они могут быть в основном внутренними. Компоненты наружных половых органов насекомых очень разнообразны по форме и часто имеют значительную таксономическую ценность, особенно среди видов, которые в других отношениях кажутся сходными по строению. Наружные гениталии самцов широко использовались для различения видов, тогда как наружные гениталии самок могут быть более простыми и менее разнообразными.
Терминалии взрослых самок насекомых включают внутренние структуры для приема копулятивного органа самца и его сперматозоидов, а также внешние структуры, используемые для откладки яиц (яйцекладка; раздел 5.8). Сегменты 8 и 9 несут гениталии; сегмент 10 виден как целый сегмент у многих «низших» насекомых, но всегда без придатков. У большинства самок насекомых есть яйцекладочная трубка или яйцеклад; он отсутствует у термитов, паразитических вшей, многих Plecoptera и большинства Ephemeroptera. Яйцеклады имеют две формы:
- Анально-генитальная часть живота. который состоит обычно из сегментов 8 или 9 до верхушки брюшка.
- замещающий, состоящий из растяжимых задних брюшных сегментов.
Другие придатки
[ редактировать ]Концевые брюшные сегменты у всех насекомых, помимо репродуктивной функции у взрослых, выполняют выделительную и сенсорную функции. [ 1 ] : 49 Небольшой сегмент 11 может быть представлен эпипроктом ( обычно спинной пластинкой или нитью над анусом некоторых насекомых); другие придатки включают:
- парапрокты : парные пластинчатые придатки , также происходящие из грудины сбоку от кончика брюшка, часто наиболее заметные у некоторых базальных отрядов, таких как Odonata ;
- церки ; : пара придатков, сочленяющихся латерально на 11-м сегменте обычно они кольчатые и нитевидные, но видоизмененные (например, щипцы уховерток ) или редуцированные у разных отрядов насекомых.
- центральная хвостовая нить , продолжение или срединный отросток dorsalis , который возникает из кончика эпипрокта у некоторых аптеригот , многих подёнок ( Ephemeroptera ) и некоторых ископаемых насекомых. [ 42 ] Подобная структура у нимфальных веснянок ( Plecoptera ) имеет неопределенную гомологию.
Внутренний
[ редактировать ]Нервная система
[ редактировать ]Нервную систему насекомого можно разделить на головной мозг и брюшной нервный канатик . Головная капсула состоит из шести сросшихся сегментов, каждый из которых имеет пару ганглиев или скопление нервных клеток за пределами мозга. насекомого Первые три пары ганглиев слиты с мозгом, а три последующие пары слиты в структуру из трех пар ганглиев под пищеводом , называемую подпищеводным ганглием . [ 1 ] : 57
Грудные сегменты имеют по одному ганглию с каждой стороны, которые соединены в пару, по одной паре на каждый сегмент. Такое расположение также наблюдается в брюшной полости, но только в первых восьми сегментах. У многих видов насекомых количество ганглиев уменьшено за счет слияния или редукции. [ 43 ] У некоторых тараканов в брюшной полости всего шесть ганглиев, тогда как у осы Vespa crabro их только два в грудной клетке и три в брюшной полости. У некоторых насекомых, например у комнатной мухи Musca Domestica , все ганглии тела слиты в один большой грудной ганглий.
По крайней мере, у некоторых насекомых есть ноцицепторы – клетки, которые обнаруживают и передают ощущения боли . [ 44 ] Это было обнаружено в 2003 г. при изучении различий в реакциях личинок обыкновенной плодовой мухи Drosophila на прикосновение нагретого и ненагретого зонда. Личинки реагировали на прикосновение нагретого зонда стереотипным поведением, которое не наблюдалось при прикосновении к личинкам ненагретым зондом. [ 45 ] Хотя ноцицепция была продемонстрирована у насекомых, не существует единого мнения о том, что насекомые чувствуют боль сознательно. [ 46 ]
Пищеварительная система
[ редактировать ]Насекомое использует свою пищеварительную систему для всех этапов обработки пищи: переваривания, всасывания, а также доставки и выведения фекалий. [ 47 ] [ 48 ] Большая часть этой пищи поступает в организм в виде макромолекул и других сложных веществ, таких как белки , полисахариды , жиры и нуклеиновые кислоты . Эти макромолекулы должны быть расщеплены катаболическими реакциями на более мелкие молекулы, такие как аминокислоты и простые сахара, прежде чем они будут использованы клетками организма для получения энергии, роста или размножения. Этот процесс распада известен как пищеварение. Основная структура пищеварительной системы насекомых представляет собой длинную закрытую трубку, называемую пищеварительным каналом (или кишечником), которая проходит вдоль тела. Пищеварительный тракт направляет пищу в одном направлении: ото рта к анусу . В кишечнике происходит почти все пищеварение насекомых. Ее можно разделить на три отдела — переднюю , среднюю и заднюю — каждый из которых выполняет свой процесс пищеварения. [ 49 ] Помимо пищеварительного канала, у насекомых имеются также парные слюнные железы и слюнные резервуары. Эти структуры обычно расположены в грудной клетке, рядом с передней кишкой. [ 1 ] : 70–77
Передняя кишка
[ редактировать ]
Первым отделом пищеварительного канала является передняя кишка (элемент 27 на нумерованной схеме), или стомодеум. Передняя кишка покрыта кутикулярной оболочкой из хитина и белков , защищающей от жесткой пищи. Передняя кишка включает в себя щечную полость (рот), глотку , пищевод , а также зоб и преджелудочек (любая часть может быть сильно изменена), которые одновременно хранят пищу и указывают, когда следует продолжать движение дальше в среднюю кишку. [ 1 ] : 70 Здесь пищеварение начинается, когда частично пережеванная пища расщепляется слюной из слюнных желез. Поскольку слюнные железы вырабатывают жидкость и ферменты, переваривающие углеводы (в основном амилазы ), сильные мышцы глотки перекачивают жидкость в полость рта, смазывая пищу, как это делает слюна, и помогая кровопитателям, а также питателям ксилемы и флоэмы.
Отсюда глотка передает пищу в пищевод, который может представлять собой простую трубку, передающую ее в зоб и преджелудочек, а затем в среднюю кишку, как у большинства насекомых. С другой стороны, передняя кишка может расширяться в очень увеличенный зоб и преджелудок, или зоб может представлять собой просто дивертикул или структуру, заполненную жидкостью, как у некоторых видов Diptera. [ 50 ] : 30–31

Слюнные железы (элемент 30 на нумерованной диаграмме) во рту насекомого вырабатывают слюну. Слюнные протоки ведут от желез к резервуарам, а затем через голову к отверстию, называемому слюной, расположенному позади гортаноглотки. Двигая ротовым аппаратом (элемент 32 на нумерованной схеме), насекомое может смешивать пищу со слюной. Смесь слюны и пищи затем попадает через слюнные трубки в рот, где начинает расщепляться. [ 47 ] [ 51 ] Некоторые насекомые, например мухи , имеют внеротовое пищеварение . Насекомые, использующие внеротовое пищеварение, выделяют пищеварительные ферменты в пищу, чтобы расщепить ее. Эта стратегия позволяет насекомым извлекать значительную часть доступных питательных веществ из источника пищи. [ 50 ] : 31
Средняя кишка
[ редактировать ]Как только пища покидает зоб, она попадает в среднюю кишку (элемент 13 на нумерованной диаграмме), также известную как мезентерон, где происходит основная часть пищеварения. Микроскопические выступы на стенке средней кишки, называемые микроворсинками , увеличивают площадь поверхности стенки и позволяют усваивать больше питательных веществ; они, как правило, располагаются близко к началу средней кишки. У некоторых насекомых роль микроворсинок и место их расположения могут различаться. Например, специализированные микроворсинки, продуцирующие пищеварительные ферменты, с большей вероятностью могут располагаться ближе к концу средней кишки, а всасывание - ближе к началу или началу средней кишки. [ 50 ] : 32
У бескрылых (аптериготных) отрядов Archaeognatha и Zygentoma (и шестиногих Entognatha ) эпителий средней кишки полностью происходит из клеток желтка. У большинства летающих насекомых ( Neoptera ) оно происходит от биполярного образования. Palaeoptera (подёнки и стрекозы) демонстрируют переход между аптериготами и неоптеранами, где средняя часть эпителия средней кишки происходит из клеток желтка, а передняя и задняя части формируются посредством биполярного образования. [ 52 ]
Задняя кишка
[ редактировать ]В задней кишке (элемент 16 на нумерованной диаграмме), или проктодеуме, непереваренные частицы пищи соединяются мочевой кислотой с образованием фекальных шариков. Прямая кишка поглощает 90% воды, содержащейся в фекальных гранулах, а затем сухие гранулы выводятся через задний проход (элемент 17), завершая процесс пищеварения. Мочевая кислота образуется из продуктов жизнедеятельности гемолимфы, диффундирующих из мальпигиевых канальцев (элемент 20). Затем он опорожняется непосредственно в пищеварительный тракт, на стыке средней и задней кишки. Количество мальпигиевых трубочек, которыми обладает одно и то же насекомое, варьируется в зависимости от вида: от всего двух трубочек у некоторых насекомых до более 100 трубочек у других. [ 1 ] : 71–72, 78–80
Дыхательные системы
[ редактировать ]Дыхание насекомых осуществляется без легких . Вместо этого дыхательная система насекомых использует систему внутренних трубок и мешочков, через которые газы либо диффундируют, либо активно перекачиваются, доставляя кислород непосредственно к тканям, которые в нем нуждаются, через трахею (элемент 8 на пронумерованной диаграмме). Поскольку кислород доставляется напрямую, система кровообращения не используется для переноса кислорода и, следовательно, значительно сокращается. Кровеносная система насекомых не имеет вен или артерий и вместо этого состоит из одной перфорированной спинной трубки, которая перистальтически пульсирует . По направлению к грудной клетке спинная трубка (элемент 14) разделяется на камеры и действует как сердце насекомого. Противоположный конец спинной трубки похож на аорту насекомого, циркулирующего гемолимфы членистоногих . , жидкого аналога крови внутри полости тела [ 1 ] : 61–65 [ 53 ] Воздух поступает через отверстия по бокам живота, называемые дыхалецами .
Существует множество различных закономерностей газообмена , демонстрируемых разными группами насекомых. Характер газообмена у насекомых может варьироваться от непрерывной и диффузной вентиляции до прерывистого газообмена . [ 1 ] : 65–68 В ходе непрерывного газообмена кислорода происходит захват углекислого газа и выделение в непрерывном цикле. Однако при прерывистом газообмене насекомое поглощает кислород, пока оно активно, и выделяет небольшое количество углекислого газа, когда насекомое находится в состоянии покоя. [ 54 ] Диффузная вентиляция — это просто форма непрерывного газообмена, который происходит за счет диффузии, а не физического поглощения кислорода. Некоторые виды насекомых, живущих под водой, также имеют приспособления, способствующие дыханию. В личиночном состоянии многие насекомые имеют жабры, способные извлекать кислород, растворенный в воде, в то время как другим необходимо подниматься к поверхности воды для пополнения запасов воздуха, который может удерживаться или задерживаться в специальных структурах. [ 55 ] [ 56 ]
Кровеносная система
[ редактировать ]Основная функция крови насекомых или гемолимфы — транспортная, и она омывает органы тела насекомого. Составляя обычно менее 25% массы тела насекомого, он переносит гормоны , питательные вещества и отходы и играет роль в осморегуляции, контроле температуры, иммунитете , хранении (воды, углеводов и жиров) и функциях скелета. Он также играет важную роль в процессе линьки. [ 57 ] [ 58 ] Дополнительная роль гемолимфы в некоторых отрядах может заключаться в защите от хищников. Он может содержать неприятные на вкус и зловонные химические вещества, которые будут отпугивать хищников. [ 1 ] Гемолимфа содержит молекулы, ионы и клетки; [ 1 ] Регулируя химический обмен между тканями , гемолимфа заключена в полости тела насекомого или гемоцеле . [ 1 ] [ 59 ] Он транспортируется по телу за счет комбинированной пульсации сердца (задней части) и аорты (передней части), которые расположены дорсально под поверхностью тела. [ 1 ] [ 57 ] [ 58 ] Она отличается от крови позвоночных тем, что не содержит эритроцитов и, следовательно, не обладает высокой способностью переносить кислород и больше похожа на лимфу , обнаруженную у позвоночных. [ 1 ] [ 59 ]
Жидкости организма поступают через односторонние клапанные отверстия, которые представляют собой отверстия, расположенные по всей длине объединенной аорты и органа сердца. Перекачивание гемолимфы происходит за счет волн перистальтического сокращения, берущих начало в заднем конце тела, перекачивающихся вперед в спинной сосуд, наружу через аорту и затем в голову, где она вытекает в гемоцель. [ 1 ] [ 59 ] Гемолимфа циркулирует к придаткам однонаправленно с помощью мышечных насосов или вспомогательных пульсирующих органов, обычно находящихся у основания усиков или крыльев, а иногда и в ногах. [ 1 ] при этом скорость откачки увеличивается с периодами повышенной активности. [ 58 ] Движение гемолимфы особенно важно для терморегуляции у таких отрядов, как Odonata , Lepidoptera , Hymenoptera и Diptera . [ 1 ]
Эндокринная система
[ редактировать ]Эти железы являются частью эндокринной системы:
1. Нейросекреторные клетки
2. Сердечный корпус
4. Тела привезли [ 60 ] [ 61 ]
Репродуктивная система
[ редактировать ]Женский
[ редактировать ]Самки насекомых способны откладывать яйца, получать и хранить сперму, манипулировать спермой разных самцов и откладывать яйца. Их репродуктивная система состоит из пары яичников , добавочных желез, одной или нескольких сперматек и протоков, соединяющих эти части. Яичники вырабатывают яйца, а добавочные железы производят вещества, помогающие упаковывать и откладывать яйца. Сперматека хранит сперму в течение разных периодов времени и вместе с частями яйцеводов может контролировать использование спермы. Протоки и сперматеки выстланы кутикулой. [ 8 ] : 880
Яичники состоят из ряда яйцевых трубок, называемых овариолами , размер и количество которых различаются в зависимости от вида. Количество яиц, которые способно отложить насекомое, зависит от количества овариол, причем на скорость развития яиц также влияет конструкция овариол. В мероистических яичниках будущие яйцеклетки делятся неоднократно, и большинство дочерних клеток становятся вспомогательными клетками для одного ооцита в кластере. В паноистических яичниках каждая яйцеклетка, вырабатываемая стволовыми зародышевыми клетками, развивается в ооцит; клеток-помощников зародышевой линии нет. Производство яиц паноистическими яичниками обычно происходит медленнее, чем мероистическими яичниками. [ 8 ] : 880
Добавочные железы или железистые части яйцеводов производят различные вещества для поддержания, транспорта и оплодотворения сперматозоидов, а также для защиты яйцеклеток. Они могут производить клей и защитные вещества для покрытия яиц или прочные покрытия для яиц, называемые оотеками . Сперматеки — это трубки или мешочки, в которых может храниться сперма между моментом спаривания и моментом оплодотворения яйцеклетки. Тестирование насекомых на отцовство показало, что некоторые, а возможно и многие, самки насекомых используют сперматеку и различные протоки для контроля или смещения используемой спермы в пользу одних самцов по сравнению с другими. [ 8 ] : 880
Мужской
[ редактировать ]Основным компонентом мужской репродуктивной системы являются семенники , подвешенные в полости тела с помощью трахей и жирового тела . У более примитивных аптериготных насекомых имеется один семенник, а у некоторых чешуекрылых два созревающих семенника вторично сливаются в одну структуру на более поздних стадиях личиночного развития, хотя ведущие от них протоки остаются отдельными. Однако у большинства самцов насекомых есть пара семенников, внутри которых находятся сперматозоиды или фолликулы, заключенные в перепончатый мешок. Фолликулы соединяются с семявыносящими протоками с помощью выносящих протоков, а два трубчатых семявыносящих протока соединяются со срединным семявыбрасывающим протоком, ведущим наружу. Часть семявыносящего протока часто увеличивается, образуя семенной пузырек, который хранит сперму до того, как она попадет в организм самки. Семенные пузырьки имеют железистую выстилку, которая выделяет питательные вещества для питания и поддержания сперматозоидов. Семявыделительный проток образуется в результате инвагинации эпидермальных клеток во время развития и в результате имеет кутикулярную выстилку. Конечная часть семявыбрасывающего протока может быть склеротизирована с образованием интромиттирующего органа, эдеагус . Остальная часть мужской репродуктивной системы происходит из эмбриональной мезодермы, за исключением зародышевых клеток, или сперматогониев , которые происходят от клеток примордиального полюса очень рано во время эмбриогенеза. [ 8 ] : 885 Эдеагус может быть достаточно выраженным или минимальным . Основанием эдеагуса может быть частично склеротизованная фаллотека, также называемая фаллосомой или текой. У некоторых видов фаллотека содержит пространство, называемое эндосомой (внутренний мешочек), в которое может втягиваться (втягиваться) кончик эдеагуса. Семявыносящий проток иногда втягивается (сворачивается) в фаллотеку вместе с семенным пузырьком. [ 62 ] [ 63 ]
Внутренняя морфология разных таксонов
[ редактировать ]Блаттодея
[ редактировать ]Тараканы наиболее распространены в тропическом и субтропическом климате. Некоторые виды тесно связаны с жилищами человека и широко встречаются возле мусора или на кухне. Тараканы, как правило, всеядны , за исключением видов, питающихся древесиной, таких как Cryptocercus ; эти тараканы не способны сами переваривать целлюлозу , но имеют симбиотические отношения с различными простейшими и бактериями , которые переваривают целлюлозу, что позволяет им извлекать питательные вещества. Сходство этих симбионтов рода Cryptocercus с симбионтами термитов таково, что было высказано предположение, что они более тесно связаны с термитами, чем с другими тараканами. [ 64 ] и текущие исследования решительно поддерживают эту гипотезу взаимоотношений. [ 65 ] Все изученные на данный момент виды несут облигатную мутуалистическую эндосимбионтную бактерию Blattabacterium , за исключением Nocticola australiensis , австралийского пещерного вида без глаз, пигмента и крыльев, который, как показывают недавние генетические исследования, является очень примитивными тараканами. [ 66 ] [ 67 ]
Тараканы, как и все насекомые, дышат через систему трубок, называемых трахеями . К дыхалецам насекомых прикрепляются трахеи , исключая голову. Таким образом, тараканы, как и все насекомые, не зависят от рта и трахеи при дыхании. Створки открываются, когда уровень CO 2 в насекомом поднимается до высокого уровня; затем CO 2 диффундирует из трахеи наружу, а свежий O 2 диффундирует внутрь. В отличие от позвоночных зависит от крови , у которых транспорт O 2 и CO 2 , трахеальная система доставляет воздух непосредственно к клеткам , трахеальные трубки постоянно разветвляются, как дерево до тех пор, пока их тончайшие подразделения, трахеолы, не будут связаны с каждой клеткой, позволяя газообразному кислороду растворяться в цитоплазме, лежащей через тонкую кутикулу, выстилающую трахеолу. CO 2 диффундирует из клетки в трахеолу. Хотя у тараканов нет легких и, следовательно, они не дышат активно, как легкие позвоночных, у некоторых очень крупных видов мускулатура тела может ритмично сокращаться, чтобы принудительно выталкивать воздух наружу и в дыхальца; это можно считать формой дыхания. [ 68 ]
Жесткокрылые
[ редактировать ]Пищеварительная система жуков в основном основана на растениях, которыми они по большей части питаются, причем пищеварение осуществляет преимущественно передняя средняя кишка . Однако у хищных видов (например, Carabidae ) большая часть пищеварения происходит в зобе с помощью ферментов средней кишки. У видов Elateridae хищные личинки испражняются ферментами на свою жертву, при этом пищеварение осуществляется внеротовым путем. [ 8 ] Пищеварительный канал в основном состоит из короткой узкой глотки , расширенного расширения, зева и слабо развитого желудка . Далее идет средняя кишка, размеры которой различаются у разных видов, с большим количеством слепой кишки , с шарнирной кишкой и различной длины. Обычно имеется от четырех до шести мальпигиевых канальцев . [ 69 ]
Нервная система жуков содержит все типы нервов насекомых, различающиеся у разных видов. От трех грудных и семи-восьми брюшных ганглиев можно отличить тот, у которого все грудные и брюшные ганглии слиты, образуя сложную структуру. Кислород получают через трахеальную систему . Воздух поступает в ряд трубок вдоль тела через отверстия, называемые дыхальцами , а затем попадает во все более тонкие волокна. [ 8 ] Насосные движения тела проталкивают воздух через систему. Некоторые виды жуков-плавунчиков ( Dytiscidae ) несут с собой пузырь воздуха всякий раз, когда ныряют под поверхность воды. Этот пузырь можно удерживать под надкрыльями или прижимать к телу специальными волосками. Пузырь обычно покрывает одно или несколько дыхалец, поэтому насекомое может дышать воздухом из пузыря, находясь под водой. Воздушный пузырь обеспечивает насекомому лишь кратковременный запас кислорода, но благодаря своим уникальным физическим свойствам кислород диффундирует в пузырь и вытесняет азот, что называется пассивной диффузией , однако объем пузырька со временем уменьшается, и жуку придется вернуться на поверхность. [ 70 ]
Как и у других видов насекомых, у жуков имеется гемолимфа вместо крови . Открытая система кровообращения жука приводится в движение трубчатым сердцем, прикрепленным к верхней внутренней части грудной клетки.
Разные железы специализируются на разных феромонах, вырабатываемых для поиска партнера. Феромоны видов Rutelinae производятся из эпителиальных клеток, выстилающих внутреннюю поверхность апикальных сегментов брюшка, или феромоны на основе аминокислот Melolonthinae из выворачивающихся желез на вершине брюшка. Другие виды производят разные типы феромонов. Дерместиды производят сложные эфиры , а виды Elateridae производят жирных кислот альдегиды и ацетаты . [ 8 ] Светлячки (Lampyridae) также использовали модифицированные жировые клетки тела с прозрачными поверхностями, покрытыми отражающими кристаллами мочевой кислоты, для биосинтетического производства света или биолюминесценции . Производство света очень эффективно, так как оно производится путем окисления люциферина ферментами люциферазой в присутствии АТФ (аденозинтрифосфата) и кислорода с образованием оксилюциферина , углекислого газа и света. [ 8 ]
У значительного числа видов развились специальные железы, вырабатывающие химические вещества для отпугивания хищников (см. Защита и хищничество ). ( Защитные железы жужелиц Carabidae), расположенные сзади, производят различные углеводороды , альдегиды , фенолы , хиноны , сложные эфиры и кислоты , выделяющиеся из отверстия в конце брюшка. В то время как африканские жужелицы (например, Anthia , некоторые из которых раньше входили в род Thermophilum ) используют те же химические вещества, что и муравьи: муравьиную кислоту . [ 31 ] При этом у жуков-бомбардиров , как и у других жужелиц, хорошо развиты пигидиальные железы, которые опорожняются с боковых краев межсегментных перепонок между седьмым и восьмым брюшными сегментами. Железа состоит из двух содержащих камер. Первый содержит гидрохиноны и перекись водорода , а второй содержит только перекись водорода плюс каталазы . Эти химические вещества смешиваются и приводят к взрывному выбросу, образуя температуру около 100 C, с распадом гидрохинона до H 2 + O 2 + хинон, при этом O 2 ускоряет выведение. [ 8 ]
Тимпанальные органы – это органы слуха. Такой орган обычно представляет собой мембрану (барабанную полость), натянутую поперек каркаса, поддерживаемого воздушным мешком и связанными с ним сенсорными нейронами. В отряде Coleoptera тимпанальные органы описаны как минимум у двух семейств. [ 30 ] У некоторых видов рода Cicindela семейства Cicindelidae уши расположены на дорсальной поверхности первого сегмента брюшка под крылом; У двух триб семейства Dynastinae ( Scarabaeidae ) уши находятся прямо под щитом переднеспинки или шейной перепонкой. Уши обеих семей воспринимают ультразвуковые частоты, и есть убедительные доказательства того, что они способны обнаруживать присутствие летучих мышей с помощью ультразвуковой эхолокации. Несмотря на то, что жуки составляют большой отряд и живут в различных нишах , у некоторых видов удивительно мало примеров слуха, хотя вполне вероятно, что большинство из них просто не обнаружены. [ 8 ]
Дермаптера
[ редактировать ]Нейроэндокринная система типична для насекомых. Имеется головной мозг, подпищеводный ганглий , три грудных ганглия и шесть брюшных ганглиев. Сильные нейронные связи соединяют нейрогемальные сердечные тела с мозгом и лобным ганглием, где близкородственное срединное аллатумное тело вырабатывает ювенильный гормон III в непосредственной близости от нейрогемальной дорсальной аорты. Пищеварительная система уховерток такая же, как и у всех других насекомых, и состоит из передней, средней и задней кишки, но у уховерток отсутствуют желудочные стволы, которые специализированы для пищеварения у многих видов насекомых. Длинные, тонкие (экстраторные) мальпигиевы канальцы можно найти между местом соединения средней и задней кишки. [ 8 ]
Половая система самок состоит из парных яичников , боковых яйцеводов , сперматеки и половой камеры . Боковые протоки — это то место, где яйцеклетки покидают тело, а сперматека — место хранения спермы. В отличие от других насекомых, гонопор , или половое отверстие, находится позади седьмого сегмента брюшка. Яичники примитивны в том смысле, что они политрофны ( питающие клетки и ооциты чередуются по длине овариолы ) . У некоторых видов эти длинные яичники отходят от латерального протока, а у других вокруг протока появляются короткие яичники. [ 8 ]
двукрылые
[ редактировать ]Гениталии самок мух повернуты в разной степени по сравнению с положением других насекомых. У некоторых мух это временное вращение во время спаривания, а у других — постоянное перекручивание органов, происходящее на стадии куколки. Этот перекрут может привести к тому, что анус окажется ниже половых органов или, в случае перекрута на 360°, к тому, что семявыносящий проток обернется вокруг кишки, несмотря на то, что внешние органы находятся в своем обычном положении. Когда мухи спариваются, самец сначала летит сверху самки, лицом в том же направлении, но затем поворачивается лицом в противоположном направлении. Это заставляет самца лежать на спине, чтобы его гениталии оставались сцепленными с гениталиями самки, или скручивание мужских гениталий позволяет самцу спариваться, оставаясь в вертикальном положении. Это приводит к тому, что у мух больше способностей к размножению, чем у большинства насекомых, и они происходят гораздо быстрее. Мухи образуют большие популяции из-за их способности эффективно и за короткий период времени спариваться, особенно во время брачного сезона. [ 71 ]
Самка откладывает яйца как можно ближе к источнику пищи, и развитие происходит очень быстро, что позволяет личинке съесть как можно больше еды за короткий период времени, прежде чем превратиться во взрослую особь. Яйца вылупляются вскоре после откладки, или мухи являются яйцеживородящими , при этом личинка вылупляется внутри матери. [ 71 ] Личинки мух, или личинки , не имеют настоящих ног и небольших границ между грудной клеткой и брюшком ; у более производных видов голова неотличима от остального тела. Личинки лишены конечностей или имеют маленькие ложноногие . Глаза и усики редуцированы или отсутствуют, на брюшке также отсутствуют придатки, такие как церки . Это отсутствие особенностей является адаптацией к среде, богатой пищей, например, к гниющему органическому веществу или к эндопаразиту . [ 71 ] Куколки . принимают различные формы, а в некоторых случаях развиваются внутри шелкового кокона После выхода из куколки взрослая муха редко живет более нескольких дней и служит в основном для размножения и расселения в поисках новых источников пищи.
Чешуекрылые
[ редактировать ]В репродуктивной системе бабочек и мотыльков гениталии самца сложны и нечетки. У самок в зависимости от родственных таксонов выделяют три типа гениталий: монотризий, экзопорий и дитрезий. У монотризового типа на сросшихся сегментах 9 и 10 грудины имеются отверстия, выполняющие функции осеменения и откладки яиц. У экзопорового типа (у Hepialoidea и Mnesarchaeoidea ) имеются два отдельных места для осеменения и откладки яиц, причем оба расположены на той же грудине, что и монотризовый тип, 9/10. У большинства видов гениталии окружены двумя мягкими лопастями, хотя у некоторых видов они могут быть специализированы и склеротизированы для откладки яиц в таких местах, как щели и внутри растительной ткани. [ 69 ] Гормоны и железы, которые их производят, управляют развитием бабочек и мотыльков на протяжении их жизненного цикла, называемого эндокринной системой . Первый гормон насекомых ПТТГ (проторакотропный гормон) управляет жизненным циклом вида и диапаузой (см. раздел «Связано »). [ 72 ] Этот гормон вырабатывается аллата и сердечными телами , где он и хранится. Некоторые железы специализируются на выполнении определенных задач, таких как выработка шелка или выработка слюны в щупиках. [ 1 ] : 65, 75 В то время как сердечные тела вырабатывают ПТТГ, аллата также вырабатывает ювенильные гормоны, а переднегрудные железы вырабатывают гормоны линьки.
В пищеварительной системе передняя часть передней кишки была изменена и образовала глоточный насос, поскольку она необходима им для приема пищи, которая по большей части представляет собой жидкости. Пищевод . следует за задней частью глотки и ведет к ней, а у некоторых видов образует форму зоба Средняя кишка короткая и прямая, задняя длиннее и изогнута. [ 69 ] Предки видов чешуекрылых, происходящие от перепончатокрылых , имели стволы средней кишки, хотя у современных бабочек и мотыльков они утрачены. Вместо этого все пищеварительные ферменты, кроме начального пищеварения, иммобилизуются на поверхности клеток средней кишки. У личинок бокаловидные клетки с длинной шеей и ножкой обнаруживаются в переднем и заднем отделах средней кишки соответственно. У насекомых бокаловидные клетки выделяют положительные ионы калия , которые поглощаются из листьев, проглатываемых личинками. Большинство бабочек и мотыльков имеют обычный пищеварительный цикл, однако виды с другим рационом требуют адаптации для удовлетворения этих новых потребностей. [ 8 ] : 279
В кровообращения системе гемолимфа или кровь насекомых используется для циркуляции тепла в форме терморегуляции , когда сокращение мышц производит тепло, которое передается остальной части тела при неблагоприятных условиях. [ 73 ] У чешуекрылых гемолимфа циркулирует по венам крыльев с помощью какого-либо пульсирующего органа: сердца или за счет поступления воздуха в трахею . [ 1 ] : 69 Воздух поступает через дыхальца по бокам брюшка и грудной клетки, снабжая трахею кислородом, проходя через дыхательную систему чешуекрылых . Есть три разные трахеи, снабжающие кислород и распространяющие кислород по всему телу вида: дорсальная, вентральная и висцеральная. Спинные трахеи снабжают кислородом спинную мускулатуру и сосуды, вентральные трахеи снабжают вентральную мускулатуру и нервные стволы, а висцеральные трахеи снабжают кишечник, жировые тела и половые железы . [ 1 ] : 71, 72
См. также
[ редактировать ]- Морфология (биология)
- Физиология насекомых
- Внешняя морфология чешуекрылых
- Внешняя морфология Odonata
- Экология насекомых
- Полет насекомых
- Беспозвоночное
- Насекомое
- Энтомология
- Доисторическое насекомое
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и Гуллан, Пи Джей; PS Крэнстон (2005). Насекомые: Очерк энтомологии (3-е изд.). Оксфорд: Издательство Блэквелл. ISBN 1-4051-1113-5 .
- ^ «Зоопарк насекомых О. Оркина» . Кафедра энтомологии Университета Небраски. Архивировано из оригинала 2 июня 2009 г. Проверено 3 мая 2009 г.
- ^ Реш, Винсент Х.; Карде, Ринг Т. (2009). Энциклопедия насекомых (2-е изд.). Сан-Диего, Калифорния: Academic Press. п. 12.
- ^ Кэмпбелл, Северная Каролина (1996) Биология (4-е издание) Бенджамин Каммингс, Новая работа. п. 69 ISBN 0-8053-1957-3
- ^ Джин Крицкий . (2002). Обзор энтомологии . iUniverse. ISBN 978-0-595-22143-1 .
- ^ Jump up to: а б «Внешняя морфология насекомых» (PDF) . Архивировано из оригинала (PDF) 19 июля 2011 г. Проверено 20 марта 2011 г.
- ^ Jump up to: а б Ричардс, Огайо; Дэвис, Р.Г. (1977). Общий учебник энтомологии Иммса: Том 1: Структура, физиология и развитие. Том 2: Классификация и биология . Берлин: Шпрингер. ISBN 0-412-61390-5 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В Реш, Винсент Х.; Ринг Т. Карде (1 июля 2009 г.). Энциклопедия насекомых (2-е изд.). США: Академическая пресса. ISBN 978-0-12-374144-8 .
- ^ Смит, Джон Бернхард, Объяснение терминов, используемых в энтомологии. Издатель: Бруклинское энтомологическое общество, 1906 г. (можно загрузить по адресу: https://archive.org/details/explanationofter00smit )
- ^ Фокс, Ричард (6 октября 2006 г.). «Внешняя анатомия» . Ландерский университет. Архивировано из оригинала 14 марта 2011 г. Проверено 20 марта 2011 г.
- ^ Волкель, Р.; Эйснер, М.; Вейбл, К.Дж. «Миниатюрные системы визуализации» (PDF) . СУСС Микрооптика. Архивировано из оригинала (PDF) 1 октября 2008 г. Проверено 20 марта 2011 г.
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Мейер-Рохов, В.Б. (1974). «Строение и функции личиночного глаза пилильщика Перга ». Журнал физиологии насекомых . 20 (8): 1565–1591. дои : 10.1016/0022-1910(74)90087-0 . ПМИД 4854430 .
- ^ Фёлькель, Р.; Эйснер, М.; Вейбл, К.Дж. (2003). «Миниатюрные системы визуализации». Микроэлектронная инженерия . 67–68: 461–472. дои : 10.1016/S0167-9317(03)00102-3 . ISSN 0167-9317 .
- ^ Гейтен, Эдвард (1998). «Оптика и филогения: есть ли понимание? Эволюция суперпозиционных глаз у десятиногих (ракообразных)» . Вклад в зоологию . 67 (4): 223–236. дои : 10.1163/18759866-06704001 .
- ^ Ричи, Александр (1985). « Ainiktozoon loganense Scourfield, протохордовый? из силура Шотландии». Алчеринга . 9 (2): 137. Бибкод : 1985Alch....9..117R . дои : 10.1080/03115518508618961 .
- ^ Майер, Г. (2006). «Строение и развитие онихофорных глаз: какой предковый орган зрения у членистоногих?». Строение и развитие членистоногих . 35 (4): 231–245. Бибкод : 2006АртСД..35..231М . дои : 10.1016/j.asd.2006.06.003 . ПМИД 18089073 .
- ^ Jump up to: а б с д Чепмен, РФ (1998). Насекомые: структура и функции (4-е изд.). Кембридж, Великобритания: Издательство Кембриджского университета. ISBN 0-521-57048-4 .
- ^ Краузе, А.Ф.; Винклер, А.; Дюрр, В. (2013). «Центральный привод и проприоцептивный контроль движений усиков у палочника». Журнал физиологии, Париж . 107 (1–2): 116–129. doi : 10.1016/j.jphysparis.2012.06.001 . ПМИД 22728470 . S2CID 11851224 .
- ^ Окада, Дж; Тох, Ю (2001). «Периферическое изображение ориентации усиков на волосистой пластинке таракана Periplaneta americana». Журнал экспериментальной биологии . 204 (Часть 24): 4301–4309. дои : 10.1242/jeb.204.24.4301 . ПМИД 11815654 .
- ^ Штаудахер, Э.; Гебхардт, MJ; Дюрр, В. (2005). «Движения антенн и механорецепция: нейробиология активных тактильных сенсоров». Достижения физиологии насекомых . 32 : 49–205. дои : 10.1016/S0065-2806(05)32002-9 . ISBN 978-0-12-024232-0 .
- ^ Сервадей, А.; Зангери, С.; Масутти, Л. (1972). Общая энтомология в прикладном ПОНИМАТЬ. пп. 492–530.
- ^ Кэмпбелл, Фрэнк Л.; Пристли, Джун Д. (1970). «Жгутиковые кольца Blattella germanica (Dictyoptera: Blattellidae). Изменения их количества и размеров в ходе постэмбрионального развития». Анналы Энтомологического общества Америки . 63 : 81–88. дои : 10.1093/aesa/63.1.81 .
- ^ «Усики насекомых» . Общество энтомологов-любителей . Проверено 21 марта 2011 г.
- ^ Jump up to: а б «Морфология насекомых» . Университет Миннесоты (факультет энтомологии). Архивировано из оригинала 3 марта 2011 г. Проверено 21 марта 2011 г.
- ^ Кирейчук, А.Г. (ноябрь 2002 г.). "Голова" . Жуки (Coleoptera) и колеоптеролог . зин.ру. Проверено 21 марта 2011 г.
- ^ Jump up to: а б с д и ж г час я дж к л м Чепмен, РФ (1998). Насекомые: структура и функции (4-е изд.). Кембридж, Нью-Йорк: Издательство Кембриджского университета. ISBN 0-521-57048-4 .
- ^ Jump up to: а б с д и Гиллиотт, Седрик (август 1995 г.). Энтомология (2-е изд.). Спрингер-Паблишерс Нью-Йорк, ООО. ISBN 0-306-44967-6 .
- ^ Капур, VCC (январь 1998 г.). Принципы и практика систематики животных . Том. 1 (1-е изд.). Научные издательства. п. 48. ИСБН 1-57808-024-Х .
- ^ «Комар кусает ротовой аппарат» . allmosquitos.com. 2011 . Проверено 17 апреля 2011 г.
- ^ Jump up to: а б с д Скобл, М.Дж. (1992). Чешуекрылые: форма, функции и разнообразие . Оксфордский университет. Нажимать. ISBN 978-1-4020-6242-1 .
- ^ Jump up to: а б Эванс, Артур В.; Беллами, Чарльз (апрель 2000 г.). Чрезмерная любовь к жукам . Издательство Калифорнийского университета. ISBN 0-520-22323-3 .
- ^ Хеппнер, Дж. Б. (2008). «Бабочки и мотыльки» . В Капинере, Джон Л. (ред.). Энциклопедия энтомологии . Виртуальная справочная библиотека Гейла. Том. 4 (2-е изд.). Справочник Спрингера. п. 4345. ИСБН 978-1-4020-6242-1 .
- ^ Агоста, Сальваторе Дж.; Янзен, Дэниел Х. (2004). «Распределение размеров тела крупных сухих лесных бабочек Коста-Рики и основная связь между морфологией растений и опылителей». Ойкос . 108 (1): 183–193. дои : 10.1111/j.0030-1299.2005.13504.x .
- ^ Кунте, Крушнамег (2007). «Аллометрия и функциональные ограничения длины хоботка у бабочек» . Функциональная экология . 21 (5): 982–987. Бибкод : 2007FuEco..21..982K . дои : 10.1111/j.1365-2435.2007.01299.x .
- ^ «Обтирание» . Университет Миннесоты. Архивировано из оригинала 27 сентября 2011 года . Проверено 17 апреля 2011 г.
- ^ «Ротовой аппарат мухи» . Ресурсы онлайн-обучения Школы биологических наук . Университет Сиднея. 4 февраля 2010 года. Архивировано из оригинала 9 апреля 2011 года . Проверено 17 апреля 2011 г.
- ^ Мейер, Джон Р. (5 января 2007 г.). «Внешняя анатомия: КРЫЛЬЯ» . Кафедра энтомологии Государственного университета Северной Каролины. Архивировано из оригинала 16 июля 2011 года . Проверено 21 марта 2011 г.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т Снодграсс, Р.Э. (декабрь 1993 г.). Основы морфологии насекомых . Корнельский университет Пресс. ISBN 0-8014-8125-2 .
- ^ Спит, ХТ (1932). «Новый метод изучения жилок крыльев подёнок и некоторые результаты (Эфемериды)» (PDF) . Энтомологические новости . Архивировано из оригинала (PDF) 30 сентября 2011 г.
- ^ «профемур, профемора – BugGuide.Net» . bugguide.net . Проверено 10 декабря 2016 г.
- ^ «Протибия – BugGuide.Net» . bugguide.net . Проверено 10 декабря 2016 г.
- ^ Общество энтомологов-любителей: определение эпипрокта
- ^ Шнайдерман, Ховард А. (1960). «Прерывистое дыхание у насекомых: роль дыхалец» . Биол. Бык . 119 (3): 494–528. дои : 10.2307/1539265 . JSTOR 1539265 .
- ^ Эйземанн, ВК; Йоргенсен, ВК; Мерритт, диджей; Райс, MJ; Крибб, BW; Уэбб, PD; Залуцкий, депутат; и др. (1984). «Чувствуют ли насекомые боль? — Биологический взгляд». Клеточные и молекулярные науки о жизни . 40 (2): 1420–1423. дои : 10.1007/BF01963580 . S2CID 3071 .
- ^ Трейси, Дж; Уилсон, Род-Айленд; Лоран, Дж; Бензер, С; и др. (18 апреля 2003 г.). «безболезненно, ген дрозофилы необходим для ноцицепции» . Клетка . 113 (2): 261–273. дои : 10.1016/S0092-8674(03)00272-1 . ПМИД 12705873 . S2CID 1424315 .
- ^ Сёмме, Л.С. (14 января 2005 г.). «Чувствительность и боль у беспозвоночных» . Норвежский научный комитет по безопасности пищевых продуктов . Проверено 30 сентября 2009 г. [ постоянная мертвая ссылка ]
- ^ Jump up to: а б «Общая энтомология – Пищеварительная и выделительная система» . Государственный университет Северной Каролины . Проверено 3 мая 2009 г.
- ^ ТЕРРА, Уолтер Р.; Феррейра, Клелия (2009), «Пищеварительная система» , Энциклопедия насекомых , Elsevier, стр. 273–281, doi : 10.1016/b978-0-12-374144-8.00083-7 , ISBN 978-0-12-374144-8 , получено 29 сентября 2020 г.
- ^ Терра, Уолтер Р.; Феррейра, Клелия (01 января 2009 г.), «Глава 74 - Пищеварительная система» , в Реше, Винсент Х.; Карде, Ринг Т. (ред.), Энциклопедия насекомых (второе издание) , Сан-Диего: Academic Press, стр. 273–281, doi : 10.1016/b978-0-12-374144-8.00083-7 , ISBN 978-0-12-374144-8 , получено 29 сентября 2020 г.
- ^ Jump up to: а б с Нация, Джеймс Л. (ноябрь 2001 г.). «15» . Физиология и биохимия насекомых (1-е изд.). ЦРК Пресс. стр. 496стр. ISBN 0-8493-1181-0 .
- ^ Дункан, Карл Д. (1939). Вклад в биологию североамериканских веспиновых ос (1-е изд.). Стэнфорд: Издательство Стэнфордского университета. стр. 24–29.
- ^ Мтоу, Сёдо; Цуцуми, Тадааки; Масумото, Мика; Мачида, Рюитиро (2023). «Возврат к формированию эпителия средней кишки у Zygentoma (Insecta) на основе исследования развития светляка Thermobia Domestica (Packard, 1873) (Lepismatidae)» . Строение и развитие членистоногих . 73 . Бибкод : 2023ArtSD..7301237M . дои : 10.1016/j.asd.2023.101237 . ПМИД 36796137 .
- ^ Мейер, Джон Р. (17 февраля 2006 г.). «Система кровообращения» . Государственный университет Северной Каролины: Кафедра энтомологии, Государственный университет Северной Каролины. п. 1. Архивировано из оригинала 27 сентября 2009 года . Проверено 11 октября 2009 г.
- ^ Чоун, СЛ; SW Николсон (2004). Физиологическая экология насекомых . Нью-Йорк: Издательство Оксфордского университета. ISBN 0-19-851549-9 .
- ^ Ричард В. Мерритт, Кеннет В. Камминс и Мартин Б. Берг (редакторы). Введение в водных насекомых Северной Америки (4-е изд.). Издательство Кендалла Ханта. ISBN 978-0-7575-5049-2 .
{{cite book}}:|author=имеет общее имя ( справка ) CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Мерритт, Р.В., К.В. Камминс и М.Б. Берг (2007). Знакомство с водными насекомыми Северной Америки . Издательство Кендалла Ханта. ISBN 978-0-7575-4128-5 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б МакГэвин, GC (2001). Основная энтомология; Введение по порядку . Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-850002-5 .
- ^ Jump up to: а б с Триплхорн, Калифорния; Джонсон, Н.Ф. (2005). Введение Боррора и Делонга в изучение насекомых . Брукс / Томсон Коул: Брукс / Томсон Коул.
- ^ Jump up to: а б с Эльзинга, Р.Дж. (2004). Основы энтомологии (6-е изд.). Нью-Джерси, США: Пирсон/Прентис Холл.
- ^ Триплхорн, Чарльз А.; Джонсон, Норман Ф (2005). Введение Боррора и Делонга в изучение насекомых (7-е изд.). Австралия: Томсон, Брукс/Коул. ISBN 978-0-03-096835-8 .
- ^ Гуллан, Пи Джей; PS Крэнстон (2005). Насекомые: Очерк энтомологии (3-е изд.). Оксфорд: Издательство Блэквелл. стр. 100-1 61–65 . ISBN 1-4051-1113-5 .
- ^ Де Карло; Дж. А. (1983). «Водные и полуводные полужесткокрылые. Исследование в группах частей мужского полового аппарата изучаемых видов с одинаковой функцией». Журнал Аргентинского энтомологического общества . 42 (1/4): 149–154.
- ^ Андерсен, Н. Мёллер (1991). «Морские насекомые: морфология гениталий, филогения и эволюция морских коньков, род Halobates (Hemiptera: Gerridae)». Зоологический журнал Линнеевского общества . 103 (1): 21–60. дои : 10.1111/j.1096-3642.1991.tb00896.x .
- ^ Эгглтон, П. (2001). «Термиты и деревья: обзор последних достижений в филогенетике термитов». Общество насекомых . 48 (3): 187–193. дои : 10.1007/pl00001766 . S2CID 20011989 .
- ^ Ло, Натан; Клаудио Банди; Хирофуми Ватанабэ; Кристина Налепа; Тициана Бенинат (2003). «Доказательства кокладогенеза между различными линиями двукрылых и их внутриклеточными эндосимбионтами» (PDF) . Молекулярная биология и эволюция . 20 (6): 907–913. дои : 10.1093/molbev/msg097 . ПМИД 12716997 .
- ^ Люн, Чи Чи Люн (22 марта 2007 г.). «Пещера может содержать недостающее звено» . theage.com.au . Проверено 7 декабря 2013 г.
- ^ Ло, Н; Бенинати, Т; Стоун, Ф; Уокер, Дж; Сакки, Л. (2007). «Тараканы, лишенные эндосимбионтов Blattabacterium : филогенетически дивергентный род Nocticola » . Письма по биологии . 3 (3): 327–30. дои : 10.1098/rsbl.2006.0614 . ПМК 2464682 . ПМИД 17376757 .
- ^ Кункель, Джозеф Г. «Как дышат тараканы?» . Часто задаваемые вопросы о тараканах . Массачусетский университет в Амхерсте . Проверено 7 декабря 2013 г.
- ^ Jump up to: а б с Жилло, К. (1995). «Бабочки и мотыльки». Энтомология (2-е изд.). Спрингер. стр. 246–266. ISBN 978-0-306-44967-3 . Проверено 14 ноября 2010 г.
- ^ Шмидт-Нильсен, Кнут (15 января 1997 г.). «Дыхание насекомых» . Физиология животных: адаптация и окружающая среда (5-е (иллюстрированное) изд.). Издательство Кембриджского университета. п. 55. ИСБН 0-521-57098-0 . Проверено 6 марта 2010 г.
- ^ Jump up to: а б с Хоэлл, Х.В., Дойен, Дж.Т. и Перселл, А.Х. (1998). Введение в биологию и разнообразие насекомых, 2-е изд . Издательство Оксфордского университета. стр. 493–499. ISBN 0-19-510033-6 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Уильямс, CM (1947). «Физиология диапаузы насекомых. II. Взаимодействие куколочного мозга и переднегрудных желез при метаморфозе гигантского тутового шелкопряда Platysamia cecropia » . Биол. Бык . 92 (2): 89–180. дои : 10.2307/1538279 . JSTOR 1538279 . ПМИД 20268135 .
- ^ Лайтон, JRB; Лавгроув, Б.Г. (1990). «Температурное переключение с диффузионной на конвективную вентиляцию у пчел» . Журнал экспериментальной биологии . 154 : 509–516. дои : 10.1242/jeb.154.1.509 .
Внешние ссылки
[ редактировать ]- Морфология насекомых. Обзор внешней и внутренней анатомии насекомых.
| Поля |
|  |
|---|---|---|
| Бактерии и грибы | ||
| Протисты | ||
| Растения | ||
| Беспозвоночные | ||
| Млекопитающие | ||
| Другие позвоночные | ||
| Глоссарии | ||
| Связанные темы | ||