Абсцизовая кислота
 | |
| Имена | |
|---|---|
| Предпочтительное название ИЮПАК (2 Z ,4 E )-5-[(1 S )-1-Гидрокси-2,6,6-триметил-4-оксоциклогекс-2-ен-1-ил]-3-метилпента-2,4-диеновая кислота кислота [3] | |
| Другие имена (2 Z ,4 E )-( S )-5-(1-Гидрокси-2,6,6-триметил-4-оксо-2-циклогексен-1-ил)-3-метил-2,4-пентандиеновая кислота ; Дорминовая кислота; [ нужна ссылка ] они спят [1] [2] | |
| Идентификаторы | |
3D model ( JSmol ) | |
| 3DMeet | |
| Сокращения | АБА |
| 2698956 | |
| ЧЭБИ | |
| ЧЕМБЛ | |
| ХимическийПаук | |
| Информационная карта ECHA | 100.040.275 |
| Номер ЕС |
|
| МеШ | Абсцизовая+кислота |
ПабХим CID | |
| номер РТЭКС |
|
| НЕКОТОРЫЙ | |
Панель управления CompTox ( EPA ) | |
| Характеристики | |
| С 15 Н 20 О 4 | |
| Молярная масса | 264.321 g·mol −1 |
| Появление | Бесцветные кристаллы |
| Плотность | 1193 г/мл |
| Температура плавления | 163 ° С (325 ° F; 436 К) [4] |
| войти P | 1.896 |
| Кислотность ( pKa ) | 4.868 |
| Основность (p K b ) | 9.129 |
| Опасности | |
| СГС Маркировка : | |
 | |
| Предупреждение | |
| Х315 , Х319 , Х335 | |
| P261 , P264 , P271 , P280 , P302+P352 , P304+P340 , P305+P351+P338 , P312 , P321 , P332+P313, P337+P313 P362 , P403 , P301 , P403+P233 , P405, P5013 +P233, P405, P501, P501, P501 , , P362 , P403+P233, P3 | |
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа). | |
Абсцизовая кислота ( АБК или абсцизин II) [5] ) — растительный гормон . АБК участвует во многих процессах развития растений, включая покой семян и почек , контроль размера органов и закрытие устьиц . Это особенно важно для растений в ответ на стрессы окружающей среды , включая засуху , засоление почвы , устойчивость к холоду, устойчивость к заморозкам , тепловой стресс и устойчивость к ионам тяжелых металлов . [6]
Открытие
[ редактировать ]В 1940-х годах Торстен Хемберг, работая в Стокгольмском университете, обнаружил доказательства существования положительной корреляции между периодом покоя и наличием в клубнях картофеля растворимого в эфире кислотного ингибитора роста . [7] [8]
В 1963 году абсцизовая кислота была впервые идентифицирована и охарактеризована как растительный гормон Фредериком Т. Аддикоттом и Ларри А. Дэвисом. Они изучали соединения, вызывающие опадение (осыпание) хлопчатника плодов (коробочек) . Два соединения были выделены и названы абсцизином I и абсцизином II. Абсцизин II в настоящее время называют абсцизовой кислотой (АБК). [5]
В растениях
[ редактировать ]Функция
[ редактировать ]Первоначально считалось, что ABA участвует в отпадении , отсюда и название. Сейчас известно, что это происходит лишь у небольшого числа растений. Передача сигналов, опосредованная АБК, также играет важную роль в реакции растений на стресс окружающей среды и патогены растений. [9] [10] Растительные гены биосинтеза АБК и последовательность этого пути выяснены. [11] [12] АБК также продуцируется некоторыми фитопатогенными грибами путем биосинтеза, отличным от биосинтеза АБК в растениях. [13]
При подготовке к зиме АБК вырабатывается в верхушечных почках . [14] Это замедляет рост растений и побуждает зачатки листьев образовывать чешуйки для защиты спящих почек в холодное время года. АБК также ингибирует деление клеток сосудистого камбия , приспосабливаясь к холодным условиям зимой, приостанавливая первичный и вторичный рост.
Абсцизовая кислота также вырабатывается в корнях в ответ на снижение водного потенциала почвы (что связано с сухой почвой) и в других ситуациях, в которых растение может находиться в состоянии стресса. Затем АБК перемещается в листья, где быстро изменяет осмотический потенциал замыкающих клеток устьиц , вызывая их сморщивание и устьиц закрытие . Закрытие устьиц, вызванное АБК, снижает транспирацию (испарение воды из устьиц), тем самым предотвращая дальнейшую потерю воды листьями в периоды нехватки воды. Была обнаружена тесная линейная корреляция между содержанием АБК в листьях и их проводимостью (сопротивлением устьиц) в зависимости от площади листа. [15]
Прорастание семян ингибируется АБК в антагонизме с гиббереллином . АБК также предотвращает потерю покоя семян . [ нужна ссылка ]
Несколько , мутантных растений Arabidopsis thaliana по АБК, были идентифицированы и доступны в Ноттингемском фондовом центре арабидопсиса - как растения с дефицитом продукции АБК, так и растения с измененной чувствительностью к ее действию. Растения, которые сверхчувствительны или нечувствительны к АБК, демонстрируют фенотипы в состоянии покоя семян , прорастания , регуляции устьиц , а у некоторых мутантов наблюдается задержка роста и коричневые/желтые листья. Эти мутанты отражают важность АБК для прорастания семян и раннего развития эмбрионов. [ нужна ссылка ]
Пирабактин ( пиридил -содержащий активатор АБК) представляет собой нафталинсульфонамидный ингибитор размножения гипокотилей клеток, который является агонистом сигнального пути АБК семян. [16] Это первый агонист пути АБК, который структурно не связан с АБК. [ нужна ссылка ]
Гомеостаз
[ редактировать ]Биосинтез
[ редактировать ]Абсцизовая кислота (АБК) представляет собой изопреноидный растительный гормон, который синтезируется по пластидному 2- C -метил-D-эритрит-4-фосфатному (MEP) пути ; В отличие от структурно родственных сесквитерпенов , которые образуются из (FDP), производного мевалоновой кислоты предшественника фарнезилдифосфата , основная цепь C 15 АБК образуется после расщепления C 40 каротиноидов в MEP. Зеаксантин является первым выделенным предшественником АБК; серия катализируемых ферментами эпоксидаций и изомеризаций через виолаксантин и окончательное расщепление C 40 каротиноида реакцией диоксигенирования дает проксимальный предшественник АБК, ксантоксин , который затем дополнительно окисляется до АБК. через абсцизовый альдегид . [11]

Абамин был спроектирован, синтезирован, разработан и затем запатентован как первый специфический ингибитор биосинтеза АБК, позволяющий регулировать эндогенные уровни АБК. [17]
Места и время биосинтеза АБК
[ редактировать ]- Синтезируется почти во всех тканях растений, например, в корнях, цветах, листьях и стеблях.
- Хранится в клетках мезофилла ( хлоренхимы ), где он конъюгируется с глюкозой посредством уридиндифосфат-глюкозилтрансферазы, образуя инактивированную форму АБК-эфира глюкозы. [18]
- Активируется и высвобождается из хлоренхимы в ответ на стресс окружающей среды , такой как тепловой стресс, водный стресс, солевой стресс. [18]
- Высвобождается при высыхании вегетативных тканей и при уплотнении корней почвы . [19]
- Синтезируется в зеленых плодах в начале зимнего периода.
- Синтезируется в созревающих семенах , обеспечивая период покоя.
- Подвижен внутри листа и может быстро перемещаться из листьев к корням (вопреки предыдущему мнению) по флоэме.
- Накопление в корнях изменяет развитие боковых корней, улучшая реакцию на стресс.
- АБК синтезируется практически во всех клетках, содержащих хлоропласты или амилопласты.
Инактивация
[ редактировать ]АБК может катаболизироваться до фазовой кислоты посредством CYP707A (группа ферментов P450 ) или инактивироваться путем конъюгации глюкозы (АБК-эфир глюкозы) с помощью фермента уридиндифосфат-глюкозилтрансферазы (UDP-глюкозилтрансфераза). Катаболизм посредством CYP707A очень важен для гомеостаза АБК, и мутанты по этим генам обычно накапливают более высокие уровни АБК, чем линии, сверхэкспрессирующие гены биосинтеза АБК. [20] об альтернативном пути катаболизма почвенных бактерий, ведущем к образованию дегидровомифолиола посредством фермента вомифолиолдегидрогеназы Сообщалось .
Эффекты
[ редактировать ]- Антитранспирант – вызывает закрытие устьиц , уменьшая транспирацию и предотвращая потерю воды. [21]
- Способствует росту корней в периоды низкой влажности. [22]
- Тормозит созревание плодов
- Отвечает за покой семян, подавляя рост клеток – подавляет прорастание семян.
- Ингибирует синтез кинетина нуклеотида [23]
- Снижает активность ферментов, необходимых для фотосинтеза . [24]
- Действует на эндодерму, предотвращая рост корней при воздействии соленой среды. [25]
- Повышение противовирусного иммунитета растений [26] [27]
Сигнальный каскад
[ редактировать ]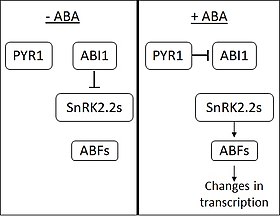
В отсутствие АБК фосфатаза ABI1-INSENSITIVE1 (ABI1) ингибирует действие SNF1-родственных протеинкиназ ( подсемейство 2) (SnRK2s). АБК воспринимается PYRABACTIN RESISTANCE 1 ( PYR1 ) и PYR1-подобными мембранными белками. При связывании ABA PYR1 связывается с ABI1 и ингибирует его. Когда SnRK2 освобождаются от ингибирования, они активируют несколько факторов транскрипции из семейства ABA RESPONSIVE ELEMENT-BINDING FACTOR (ABF). ABF затем вызывают изменения в экспрессии большого количества генов . [6] Считается, что около 10% генов растений регулируются АБК. [ нужна ссылка ]
В грибах
[ редактировать ]Как и растения, некоторые виды грибов (например, Cercospora rosicola , Botrytis cinerea [28] и Magnaporthe oryzae ) имеют эндогенный путь биосинтеза АБК. У грибов, по-видимому, преобладает путь биосинтеза MVA (а не путь MEP , который отвечает за биосинтез АБК в растениях). Одной из функций АБК, продуцируемой этими патогенами, по-видимому, является подавление иммунных реакций растений. [29]
У животных
[ редактировать ]Также было обнаружено, что АБК присутствует у многоклеточных животных , от губок до млекопитающих , включая человека. [30] В настоящее время его биосинтез и биологическая роль у животных изучены мало. АБК оказывает мощное противовоспалительное и антидиабетическое действие на мышиных моделях диабета/ожирения, воспалительных заболеваний кишечника, атеросклероза и гриппозной инфекции. [31] Многие биологические эффекты на животных были изучены с использованием АБК в качестве нутрицевтика или фармакогностического препарата, но АБК также генерируется эндогенно некоторыми клетками (например, макрофагами ) при их стимуляции. Существуют также противоречивые выводы из различных исследований: некоторые утверждают, что АБК необходима для провоспалительных реакций, тогда как другие демонстрируют противовоспалительный эффект. Как и многие природные вещества с медицинскими свойствами, АБК стала популярной и в натуропатии . Хотя АБК явно обладает полезной биологической активностью [ нужна ссылка ] и многие натуропатические средства содержат высокие уровни АБК (например, сок ростков пшеницы , фрукты и овощи), некоторые заявления о пользе для здоровья могут быть преувеличенными или чрезмерно оптимистичными. В клетках млекопитающих АБК нацелена на белок, известный как лантионинсинтетаза C-подобная 2 ( LANCL2 ), запуская альтернативный механизм активации гамма-рецептора, активируемого пролифератором пероксисомы (PPAR гамма) . [32] LANCL2 консервативен в растениях, и первоначально предполагалось, что он также является рецептором ABA у растений, что позже было подвергнуто сомнению. [33]
Измерение концентрации АБК
[ редактировать ]Несколько методов могут помочь количественно определить концентрацию абсцизовой кислоты в различных тканях растений. Используемые количественные методы основаны на ВЭЖХ и ИФА . Два независимых зонда FRET могут измерять внутриклеточные концентрации АБК в реальном времени in vivo. [34] [35]
Ссылки
[ редактировать ]- ^ О'Нил, Мариадель Дж; Хекельман, ЧП; Кох, CB; Роман, К.Дж. (2006). Индекс Merck, 14 место .
- ^ 21293-29-8
- ^ «Абсцизовая кислота - Краткое описание соединений» . Пабхим соединение . США: Национальный центр биотехнологической информации. 16 сентября 2004 г. Идентификация и соответствующие записи . Проверено 22 октября 2011 г.
- ^ «База данных ChemSpider — Абсцизовая кислота — Свойства» . Проверено 27 декабря 2012 г. Точка плавления определена по экспериментальным данным компании Tokyo Chemical Industry Ltd.
- ^ Jump up to: а б Дэвис, Луизиана; Аддикотт, FT (апрель 1972 г.). «Абсцизовая кислота: корреляция с опадением и развитием плодов хлопчатника» . Физиология растений . 49 (4): 644–648. дои : 10.1104/стр.49.4.644 . ISSN 0032-0889 . ПМК 366021 . ПМИД 16658017 .
- ^ Jump up to: а б Финкельштейн, Рут (1 ноября 2013 г.). «Синтез и реакция абсцизовой кислоты» . Книга «Арабидопсис» . 11 : e0166. дои : 10.1199/tab.0166 . ПМК 3833200 . ПМИД 24273463 .
- ^ Хемберг, Торстен (январь 1949 г.). «Значение ростингибирующих веществ и ауксинов для периода покоя клубня картофеля» . Физиология Плантарум . 2 (1): 24–36. дои : 10.1111/j.1399-3054.1949.tb07645.x . ISSN 0031-9317 .
- ^ Дёрффлинг, Карл (1 декабря 2015 г.). «Открытие абсцизовой кислоты: ретроспектива» . Журнал регулирования роста растений . 34 (4): 795–808. дои : 10.1007/s00344-015-9525-6 . ISSN 1435-8107 . S2CID 253856375 .
- ^ Чжу, Цзянь-Кан (2002). «Передача сигналов стресса от соли и засухи в растениях» . Ежегодный обзор биологии растений . 53 : 247–73. doi : 10.1146/annurev.arplant.53.091401.143329 . ПМК 3128348 . ПМИД 12221975 .
- ^ Со, М; Кошиба, Т (2002). «Комплексная регуляция биосинтеза АБК в растениях». Тенденции в науке о растениях . 7 (1): 41–8. дои : 10.1016/S1360-1385(01)02187-2 . ПМИД 11804826 .
- ^ Jump up to: а б Намбара, Эйдзи; Мэрион-Пол, Энни (2005). «Биосинтез и катаболизм абсцизовой кислоты». Ежегодный обзор биологии растений . 56 : 165–85. doi : 10.1146/annurev.arplant.56.032604.144046 . ПМИД 15862093 .
- ^ Милборроу, Б.В. (2001). «Путь биосинтеза абсцизовой кислоты в сосудистых растениях: обзор современного состояния знаний о биосинтезе АБК» . Журнал экспериментальной ботаники . 52 (359): 1145–64. дои : 10.1093/jexbot/52.359.1145 . ПМИД 11432933 .
- ^ Сиверс, В.; Смедсгаард, Дж.; Тудзинский, П. (2004). «Монооксигеназа P450 BcABA1 необходима для биосинтеза абсцизовой кислоты у Botrytis cinerea» . Прикладная и экологическая микробиология . 70 (7): 3868–76. Бибкод : 2004ApEnM..70.3868S . дои : 10.1128/АЕМ.70.7.3868-3876.2004 . ПМЦ 444755 . ПМИД 15240257 .
- ^ Ван, Дунлин; Гао, Чжэньчжэнь; Ду, Пейён; Сяо, Вэй; Тан, Цюпин; Чен, Сюде; Ли, Линг; Гао, Дуншэн (2016). «Экспрессия генов, связанных с метаболизмом АБК, предполагает сходство и различия между покоем семян и покоем почек персика (Prunus persica)» . Границы в науке о растениях . 6 : 1248. doi : 10.3389/fpls.2015.01248 . ISSN 1664-462X . ПМЦ 4707674 . ПМИД 26793222 .
- ^ Стойер, Барбара; Томас Штулфаут; Генрих П. Фок (1988). «Эффективность использования воды у растений, испытывающих водный стресс, повышается благодаря закрытию устьиц, индуцированному АБК». Исследования фотосинтеза . 18 (3): 327–336. Бибкод : 1988PhoRe..18..327S . дои : 10.1007/BF00034837 . ISSN 0166-8595 . ПМИД 24425243 . S2CID 30298332 . [ нужна ссылка ]
- ^ Пак, Сан-Юль; П. Фунг; Н. Нисимура; доктор Дженсен; Х. Фуиджи; Ю. Чжао, С. Лумба; и др. (май 2009 г.). «Абсцизовая кислота ингибирует протеинфосфатазы типа 2C через семейство белков START PYR/PYL» . Научная сигнализация . 324 (5930): 1068–1071. Бибкод : 2009Sci...324.1068P . дои : 10.1126/science.1173041 . ПМК 2827199 . ПМИД 19407142 .
- ^ US 7098365 , Yoshida, Shigeo & Asami, Tadao, «Ингибитор биосинтеза абсцизовой кислоты», опубликован 29 августа 2006 г., передан Riken.
- ^ Jump up to: а б Чжан, Юцинь; Киламби, Химабинду Васуки; Лю, Цзе; Бар, Хамутал; Лазари, Шани; Эгбария, Айман; Потрошитель, Дагмар; Шарье, Лоуренс; Белью, Зейну Мусса; Вульф, Николай; Дамодаран, Суреш; Нур-Элдин, Хусам Хасан; Ахарони, Асаф; Рагни, Лаура; Стрейдер, Люсия (22 октября 2021 г.). «Гомеостаз АБК и транслокация на большие расстояния чрезмерно регулируются импортерами АБК ABCG» . Достижения науки . 7 (43). дои : 10.1126/sciadv.abf6069 . ISSN 2375-2548 . ПМЦ 8528425 . ПМИД 34669479 .
- ^ ДеДжонг-Хьюз, Дж. и др. (2001) Уплотнение почвы: причины, последствия и контроль. Служба распространения знаний Университета Миннесоты
- ^ Финкельштейн, Рут (ноябрь 2013 г.). «Синтез и реакция абсцизовой кислоты» . Книга «Арабидопсис» . 11 : e0166. дои : 10.1199/tab.0166 . ПМК 3833200 . ПМИД 24273463 .
- ^ Чжан, Цзяньхуа; Шурр, У.; Дэвис, WJ (1987). «Контроль поведения устьиц с помощью абсцизовой кислоты, которая, по-видимому, возникает в корнях». Журнал экспериментальной ботаники . 38 (7): 1174–1181. дои : 10.1093/jxb/38.7.1174 .
- ^ Раллс, Эрик (27 июня 2023 г.). «В засушливые дни листья растений посылают своим корням сигналы, приказывая им продолжать копать глубже в поисках воды» . www.msn.com/ . Проверено 4 октября 2023 г.
- ^ Мерник, Ю.А. (1979). «Ингибирование абсцизовой кислотой образования нуклеотидов кинетина в прорастающих семенах салата». Физиология Плантарум . 45 : 63–6. дои : 10.1111/j.1399-3054.1979.tb01664.x .
- ^ Чендлер, премьер-министр; Робертсон, М. (1994). «Экспрессия генов, регулируемая абсцизовой кислотой, и ее связь с устойчивостью к стрессу». Ежегодный обзор физиологии растений и молекулярной биологии растений . 45 : 113–41. дои : 10.1146/annurev.pp.45.060194.000553 .
- ^ Дуан, Лина; Д. Дитрих; СН Нг; ПМИ Чан; Р. Бхалерао; М. Дж. Беннетт; Джей Ар Диннени. (январь 2013 г.). «Эндодермальная передача сигналов АБК способствует покою латерального корня во время солевого стресса у проростков арабидопсиса» . Растительная клетка . 25 (1): 324–341. дои : 10.1105/tpc.112.107227 . ПМЦ 3584545 . ПМИД 23341337 .
- ^ Пасин, Фабио; Шан, Хунъин; Гарсиа, Беатрис; Мюллер, Марен; Сан-Леон, Дэвид; Людман, Марта; Фресно, Дэвид Х.; Фатиол, Карой; Мунне-Бош, Сержи; Родриго, Гильермо; Гарсия, Хуан Антонио (14 сентября 2020 г.). «Абсцизовая кислота связывает передачу сигналов фитогормонов с метаболическими путями РНК и способствует противовирусному ответу, от которого уклоняется самоконтролируемый РНК-вирус» . Заводские коммуникации . 1 (5): 100099. doi : 10.1016/j.xplc.2020.100099 . ISSN 2590-3462 . ПМЦ 7518510 . ПМИД 32984814 .
- ^ Алазем, Мазен; Линь, На-Шэн (2017). «Противовирусная роль абсцизовой кислоты в растениях» . Границы в науке о растениях . 8 : 1760. дои : 10.3389/fpls.2017.01760 . ISSN 1664-462X . ПМЦ 5641568 . ПМИД 29075279 .
- ^ Сиверс, Верена; Коккелинк, Леони; Смедсгаард, Йорн; Тудзинский, Пол (июль 2006 г.). «Идентификация кластера генов абсцизовой кислоты в серой плесени Botrytis cinerea» . Appl Environ Microbiol . 72 (7): 4619–4626. Бибкод : 2006ApEnM..72.4619S . дои : 10.1128/АЕМ.02919-05 . ПМЦ 1489360 . ПМИД 16820452 .
- ^ Ливенс, Лоренс; Поллиер, Джейкоб; Гуссенс, Ален; Бейерт, Руди; Сталь, Йенс (2017). «Абсцизовая кислота как эффектор патогена и регулятор иммунитета» . Границы в науке о растениях . 8 :587 дои : 10.3389/fpls.2017.00587 . ISSN 1664-462X . ПМК 5395610 . ПМИД 28469630 .
- ^ На-Ханг, Ли; Жуй-Лин, Хао; Шань-Шань, Ву; Пэн-Чэн, Го; Цань-Цзян, Чен; Ли-Пин, Пан; Он, Ни (2011). «Возникновение, функции и потенциальное медицинское применение фитогормона абсцизовой кислоты у животных и людей». Биохимическая фармакология . 82 (7): 701–712. дои : 10.1016/j.bcp.2011.06.042 . ПМИД 21763293 .
- ^ Бассаганья-Риера, Дж; Сконечка, Дж; Кингстон, генеральный директор; Кришнан, А; Мисяк, С.А.; Гури, Эй Джей; Перейра, А; Картер, AB; Минорский, П; Тумаркин Р.; Хонтесильяс, Р. (2010). «Механизмы действия и медицинское применение абсцизовой кислоты» . Современная медицинская химия . 17 (5): 467–78. дои : 10.2174/092986710790226110 . ПМИД 20015036 . Архивировано из оригинала 1 апреля 2012 г. Проверено 30 сентября 2018 г.
- ^ Бассаганья-Риера, Дж.; Гури, Эй Джей; Лу, П.; Климент, М.; Карбо, А.; Собрал, Б.В.; Хорн, штат Вашингтон; Льюис, С.Н.; Беван, ДР; Хонтесильяс, Р. (2010). «Абсцизовая кислота регулирует воспаление посредством независимой от лигандсвязывающего домена активации рецептора, активируемого пролифератором пероксисомы» . Журнал биологической химии . 286 (4): 2504–16. дои : 10.1074/jbc.M110.160077 . ПМК 3024745 . ПМИД 21088297 .
- ^ Чен, Дж. Г.; Эллис, Б.Э. (2008). «GCR2 является новым членом семейства C-подобных белков эукариотического компонента лантионинсинтетазы» . Поведение сигнала объекта . 3 (5): 307–10. Бибкод : 2008PlSiB...3..307C . дои : 10.4161/psb.3.5.5292 . ПМЦ 2634266 . ПМИД 19841654 .
- ^ Ваадт, Р; Хитоми, К; Нисимура, Н.; Хитоми, К; Адамс, СР; Гецофф, Эд ; Шредер, Дж.И. (2014). «Репортеры на основе FRET для прямой визуализации изменений концентрации и распределения абсцизовой кислоты в Arabidopsis» . электронная жизнь . 3 : e01739. doi : 10.7554/eLife.01739 . ПМЦ 3985518 . ПМИД 24737861 .
- ^ Джонс, AM; Дэниэлсон, Дж.А.; Манджокумар, С.Н.; Лакар, В; Гроссманн, Г; Фроммер, ВБ (2014). «Динамика абсцизовой кислоты в корнях, обнаруженная с помощью генетически закодированных датчиков FRET» . электронная жизнь . 3 : e01741. doi : 10.7554/eLife.01741 . ПМЦ 3985517 . ПМИД 24737862 .
| Базы данных органов управления : Национальные |
|---|