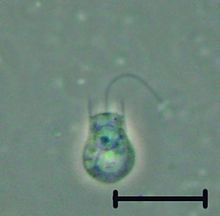
Хуанофлагелляты которые представляют собой группу свободноживущих одноклеточных и колониальных флагуальных эукариот, считаются самыми близкими живыми родственниками животных . Choanoflagellates представляют собой жгутики с воротником, имеющие воротник в форме воронки с взаимосвязанными микроворками у основания жгутика . Чонофлагелляты способны как на асексуальное , так и сексуальное размножение. [ 9 ] Они имеют отличительную клеточную морфологию , характеризующуюся яйцевидной или сферической клеточной телом диаметром 3–10 мкм с одним апикальным жгутиком, окруженным воротником 30–40 микроворсинок (см. Рисунок). Движение жгутика создает водные течения, которые могут выдвигать свободно плавающего хоанофлагелляция через толщу воды и улавливать бактерии и детрит против воротника микроворсинга, где эти продукты питания охватываются. Это кормление обеспечивает критическую связь в глобальном углеродном цикле , связывая трофические уровни . В дополнение к их критической экологической роли, хонофлагелляты представляют особый интерес к эволюционным биологам, изучающим происхождение многоклеточности у животных. Как самые близкие живые родственники животных, хонофлагелляты служат полезной моделью для реконструкций последнего одноклеточного предка животных. Согласно исследованию 2021 года, Crown Group Craspedids (и, возможно, Crown Group Choanoflagellates, если Acanthoecida возникла в рамках Craspedida [ 10 ] ) появился 422,78 миллионов лет назад, [ 11 ] Хотя предыдущее исследование 2017 года восстановило дивергенцию группы Crown Choanoflagellates (Craspedids) на 786,62 миллиона лет. [ 12 ]
Каждый хоанофлагеллят имеет единый жгутичок , окруженный кольцом актина , заполненных протруйками, называемыми микроворками , образуя цилиндрический или конический воротник ( choanos в греческом языке). Движение жгутика протягивает воду через воротник, а бактерии и детрит захватываются микроворками и проглатываются. [ 13 ] Водные токи, генерируемые жгутиком, также подталкивают клетки свободного плавания, как в животных сперме . Напротив, большинство других жгутиков тянутся их жгутиками. [ Цитация необходима ]
В дополнение к одному апикальному жгутике, окруженному актиновыми микроворителями, которые характеризуют хонофлагелляты, внутренняя организация органеллов в цитоплазме является постоянной. [ 14 ] Бламенное базальное тело сидит у основания апикального жгутика, а второе нефлагеллярное базальное тело лежит под прямым углом к основанию жгутиков. Ядро пищевые занимает апикальное к центру положения в клетке, а вакуоли расположены в базальной области цитоплазмы. [ 14 ] [ 15 ] Кроме того, клеточный организм многих хоанофлагеллятов окружен отличительным внеклеточным матриксом или перипластом . Эти клеточные покрытия сильно различаются по структуре и составу и используются таксономистами в целях классификации. Многие хонофлагелляты строят сложные «дома в форме корзины», называемые Лорикой , из нескольких полос кремнезема, укрепленных вместе. [ 14 ] Функциональная значимость перипласта неизвестна, но у сидячих организмов считается, что помогает привязанности к субстрату. В планктонных организмах есть предположения о том, что перипласт увеличивает сопротивление, тем самым противодействуя силе, генерируемой жгутиком и повышением эффективности кормления. [ 16 ]
Чоанофлагелляты либо свободно плавают в толще воды, либо сидячими , придерживаясь подложки напрямую, либо через перипласт или тонкую цветоножку. [ 17 ] Хотя считается, что хонофлагелляты строго свободно живут и гетеротрофны , ряд хоанофлагеллятных родственников, таких как члены Ichthyosporea или мезомицетозои , следуют паразитному или патогенному образу жизни. [ 18 ] Истории жизни хонофлагеллятов плохо изучены. Считается, что многие виды одиноки; Тем не менее, колониальность, по -видимому, возникала независимо несколько раз в группе, и колониальные виды все еще сохраняют одиночную стадию. [ 17 ]
Более 125 существующих видов choanoflagellates [ 13 ] известны, распределены во всем мире в морской , солоноватой и пресной среде от арктической до тропиков, занимая пелагические и бентические зоны. Хотя большая часть выборки хоанофлагеллатов произошла от 0 до 25 м (0 и 82 фута), они были извлечены из глубины до 300 м (980 футов) в открытой воде [ 19 ] и 100 м (330 футов) под антарктическими ледяными щитами. [ 20 ] Предполагается, что многие виды являются космополитическими в глобальном масштабе [например, Diaphanoeca Grandis сообщалось из Северной Америки , Европы и Австралии (OBIS)], в то время как другие виды имеют ограниченное региональное распределение. [ 21 ] Совместные виды хоанофлагелляции могут занимать совершенно разные микроокружения, но в целом факторы, которые влияют на распределение и дисперсию хоанофлагеллятов, еще предстоит выяснить. [ Цитация необходима ]
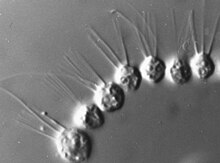
Ряд видов , таких как род Proterospongia , образуют простые колонии , [ 13 ] планктонные комки, напоминающие миниатюрный кластер винограда, в котором каждая ячейка в колонии жгутична или кластеры клеток на одном стебле. [ 14 ] [ 22 ] В октябре 2019 года ученые обнаружили новое поведение Choanoflagellates: они, очевидно, могут координировать, чтобы ответить на свет. [ 23 ]
Гомеостаз кальция современных сперматозоидов (б) выглядит очень похоже на гомех древнего хоанофлагеллята (а). Фарнесол очень древний по эволюции, и его использование восходит, по крайней мере, до того, как хонофлагелляты, которые предшествовали животным. [ 26 ] [ нужно разъяснения ]
Чонофлагелляты растут вегетативно, с несколькими видами, подвергающимися продольному делению; [ 15 ] Тем не менее, репродуктивный жизненный цикл хонофлагеллятов еще предстоит выяснить. Документ, опубликованный в августе 2017 года, показал, что изменения окружающей среды, включая наличие определенных бактерий, вызывают рой и последующее сексуальное воспроизведение хоанофлагеллятов. [ 9 ] Плоидский ; уровень неизвестен [ 27 ] Однако открытие как ретротранспозонов, так и ключевых генов, участвующих в мейозе [ 28 ] Ранее предположили, что они использовали сексуальное размножение как часть своего жизненного цикла. Некоторые хоанофлагелляты могут подвергаться окружению, которое включает в себя втягивание жгутикового и воротника и оборота в электронном плотной фибриллярной стенке. При переводе в свежие носители происходит экслан; хотя это еще предстоит непосредственно наблюдать. [ 29 ]
Доказательства сексуального размножения были зарегистрированы в видах хоанофлагелляции Salpingoeca Rosetta . [ 30 ] [ 31 ] Также сообщалось о наличии консервативных мейотических генов в хоанофлагеллятах моносиги Brevicollis и Monosiga Ovata . [ 32 ]
Чонофлагелляты Acanthoecid производят внеклеточную структуру корзины, известную как лорика. Лорика состоит из отдельных костюмов, изготовленных из биокомпозита кремнезема. Каждая каждая полоса образуется в ячейке хоанофлагелляции, а затем секретируется на клеточную поверхность. В орехообразных хоанофлагеллятах сборка Лорики происходит с использованием ряда щупальц, как только было получено достаточное количество костюмов, включающих полную лорику. В тектоформных хоанофлагеллятах костюмы накапливаются в установленном расположении ниже воротника. Во время клеточного деления новая ячейка принимает эти костюмы в рамках цитокинеза и собирает свою собственную Lorica, используя только эти ранее произведенные полосы. [ 33 ]
Биосилицификация хоанофлагелляции требует концентрации кремниковой кислоты в клетке. Это осуществляется белками транспортера кремния (SIT). Анализ SCITS Choanoflagellate показывает, что они похожи на кремниевые переносчики типа диатомовых и других формирующих кремнезема . Семейство SIT Gene показывает мало или вообще не является гомологией для любых других генов, даже для генов в неязных хоанофлагеллятах или StrameNopiles. Это говорит о том, что семейство SIT генов развивалось с помощью бокового события переноса генов между акантоцидами и стризенопилами. Это замечательный случай горизонтального переноса генов между двумя отдаленно связанными эукариотическими группами и дал подсказки для биохимии и кремниевых взаимодействий с уникальным семейством генов. [ 34 ]
Феликс Дюджардин , французский биолог, заинтересованный в эволюции простейших, зафиксировал морфологические сходства хоанофлагеллятов и губчатых хоаноцитов и предложил возможность тесных отношений еще в 1841 году. [ 16 ] За последнее десятилетие эта гипотетическая взаимосвязь между хоанофлагеллятами и животными была поддержана независимым анализом множественных неопределенных генетических последовательностей: 18S рДНК, гены кодирования ядерного белка и геномы митохондриальных (Steenkamp, et al., 2006; Burger, et al. , 2003; [ 18 ] Wainright, et al., 1993). Важно отметить, что сравнения последовательностей митохондриального генома из хоанофлагеллята и трех губок подтверждают размещение хоанофлагеллятов в качестве внешней группы к метазоа (животных, также известных как Animalia ) и отрицают возможность того, что хонофлагелляты эволюционировали из метазоян (Lavrov, et al., 2005). Наконец, исследование генов в 2001 году, экспрессируемых в хонофлагеллятах, показало, что хонофлагелляции синтезируют гомологи передачи сигналов и адгезии клеток метазоина. [ 35 ] Секвенирование генома показывает, что среди живых организмов хонофлагелляты наиболее тесно связаны с животными. [ 13 ]
Поскольку чонофлагелляты и метазои тесно связаны, сравнения между двумя группами обещают дать представление о биологии их последнего общего предка и самых ранних событий в эволюции метазоя . Хуаноциты . (также известные как «клетки с воротниками») губок (рассматриваемых среди наиболее базальных метазоа) имеют такую же основную структуру, что и хонофлагелляты Клетки с воротниками обнаружены в других группах животных, таких как ленточные черви , [ 36 ] предполагая, что это была морфология их последнего общего предка. Последний общий предок животных и хоанофлагеллятов был одноклеточным, возможно, образуя простые колонии; Напротив, последним общим предком всех эйметазоических животных был многоклеточный организм, с дифференцированными тканями, определенным «планом тела» и эмбриональным развитием (включая гаструляцию). [ 13 ] Время расщепления этих линий трудно ограничить, но, вероятно, было в позднем докембрийском,> 600 миллионах лет назад . [ 13 ]
Чонофлагелляты были включены в Chrysophyceae до Hibberd, 1975. [ 38 ] Недавняя молекулярная филогенетическая реконструкция внутренних взаимосвязей хоанофлагеллятов позволяет поляризировать эволюцию характера в кладе. Большие фрагменты ядерной РНК рибосомальной белка , альфа -тубулин и 90 тепло-шока 90, использовались для разрешения внутренних отношений и полярности характера в хонофлагеллятах. [ 22 ] Каждый из четырех генов показал сходные результаты независимо независимо, и анализ комбинированного набора данных ( конкатенированный ) вместе с последовательностями из других близкородственных видов ( животных и грибов ) демонстрируют, что хонофлагелляты сильно подтверждаются как монофилетические и подтверждают их положение как наиболее близкое известное одноцветичное. родственник животных.
Ранее Choanoflagellida был разделен на эти три семейства на основе композиции и структуры их перипластов: Codonosigidae, Salpingoecidae и Acanthoecidae. Члены семейства Codonosigidae, по -видимому, не имеют перипласта при исследовании с помощью световой микроскопии, но может иметь тонкий внешний слой, видимый только с помощью электронной микроскопии . Семейство SalpingOecidae состоит из видов, клетки которых заключены в твердую тека, которая видна как с помощью световой, так и электронной микроскопии. Theca представляет собой секретируемый охват преимущественно состоит из целлюлозы или других полисахаридов . [ 39 ] В настоящее время известно, что эти подразделения являются парафилетическими , с конвергентной эволюцией этих форм широко распространены. Третье семейство Choanoflagellates, Acanthoecidae, было поддержано как монофилетическая группа. Эта клада обладает синапоморфией клеток, обнаруживаемых в лорике, подобной корзине, предоставляя альтернативное название «Loricate Choanoflagellates». Acanthoecid Lorica состоит из ряда кремнистых костюмов, расположенных в видоспецифичной схеме Lorica ». [ 14 ] [ 16 ]
Хуанофлагеллятное дерево, основанное на молекулярной филогенетике, делится на три хорошо поддерживаемых клад . [ 22 ] CLADE 1 и CLADE 2 каждый состоит из комбинации видов, традиционно приписываемых Codonosigidae и SalpingOecidae, в то время как клада 3 содержит виды из группы, таксономически классифицированных как Acanthoecidae. [ 22 ] Картирование черт характера с этой филогении указывает на то, что последним общим предком Choanoflagellates был морской организм с дифференцированным жизненным циклом с сидячими и подвижными этапами. [ 22 ]
Геном моносиги Brevicollis , с 41,6 миллионами пар оснований, [ 13 ] по размеру схож с нитевидными грибами и другими свободными одноклеточными эукариотами, но гораздо меньше, чем у типичных животных. [ 13 ] В 2010 году филогеномное исследование показало, что несколько генов водорослей присутствуют в геноме моносиги Brevicollis . Это может быть связано с тем, что в ранней эволюционной истории хоанофлагелляты потребляли водоросли в качестве пищи посредством фагоцитоза . [ 40 ] Carr et al. (2010) [ 32 ] Провернул геном M. brevicollis для известных эукариотического генов мейоза. Из 19 известных эукариотических мейотических генов (включая 8, которые функционируют ни в каком другом процессе, кроме мейоза), 18 были идентифицированы у M. brevicollis . Наличие мейотических генов, включая гены, специфичные для мейоза, указывает на то, что мейоз, и, как следствие, пол в хонофлагеллятах присутствует .
Геном Salpingoeca Rosetta составляет 55 мегабаз размера. [ 41 ] Гомологи генов клеточной адгезии, нейропептида и гликосфинголипидов присутствуют в геноме.
S. rosetta имеет сексуальный жизненный цикл и переходы между гаплоидными и диплоидными этапами . [ 31 ] В ответ на ограничение питательных веществ гаплоидные культуры С. Розетты становятся диплоидными. Этот сдвиг плоиды совпадает с спариванием, в ходе которого небольшие жгутикованные ячейки сливаются с более крупными флаглельными клетками. Существуют также доказательства исторического спаривания и рекомбинации в С. Розетте .
S. rosetta заинтересована для сексуального размножения морской бактерий Vibrio fischeri . [ 30 ] Единый белок V. fischeri , эрос полностью повторяет афродизиак-подобную активность Live V. Fisheri .
Одноклеточные амплифицированные геномы четырех некультурных морских хонофлагеллятов, предварительно называемых UC1-UC4, были секвенированы в 2019 году. Геномы UC1 и UC4 являются относительно полными. [ 42 ]
Набор данных EST от Monosiga Ovata был опубликован в 2006 году. [ 43 ] Основным выводом этого транскриптома стал домен хоглета Hoanoflagellate и пролил свет на роль перетасовки домена в эволюции сигнального пути ежа . М. Овата имеет по крайней мере четыре эукариотических мейотических гена. [ 32 ]
Транскриптом Spehanoeca Diplocostata был опубликован в 2013 году. Этот первый транскриптом лориката хоанофлагеллата [ 34 ] привел к открытию переносчиков кремния хоанофлагеллата. Впоследствии аналогичные гены были идентифицированы у второго лорикатного вида, Diaphanoeca grandis . Анализ этих генов обнаружил, что переносчики кремния хоанофлагеллята демонстрируют гомологию к кремниевым транспортерам типа сидячих диатомов и развивались с помощью горизонтального переноса генов .
Еще 19 транскриптомов были опубликованы в 2018 году. Было обнаружено большое количество семейств генов, которые ранее считались только животными. [ 44 ]
^ Кавалер-Смит Т (1998). «Неомонада и происхождение животных и грибов». В Coombs GH, Vickerman K, Sleigh Ma, Warren A (Eds.). Эволюционные отношения между простейшими . Лондон: Kluwer. С. 375–407.
^ Jump up to: а беременный в дюймовый и Leadbeater BS, Thomsen H (2000). «Заказ choanoflagellida». Иллюстрированное руководство по простейшим, второе издание. Лоуренс: Общество простейших . 451 : 14–38.
^ Томсен Х (1982). Планктонные хоанофлагелляции из Диско Багта, Западная Гренландия, с обзором морского нанопланктона этого района . Meddelelser Om Gronland, Bioscience. Тол. 8. С. 3–63. ISBN 978-87-635-1149-0 .
Arc.Ask3.Ru Номер скриншота №: c1e60e7b5d32f2db007c5592a35ca574__1725726000 URL1:https://arc.ask3.ru/arc/aa/c1/74/c1e60e7b5d32f2db007c5592a35ca574.html Заголовок, (Title) документа по адресу, URL1: Choanoflagellate - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)