Таламус
| Таламус | |
|---|---|
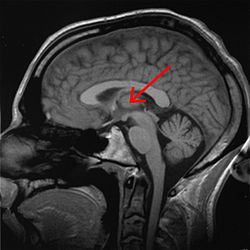 Таламус отмечен ( МРТ ) поперечное сечение | |
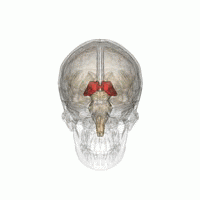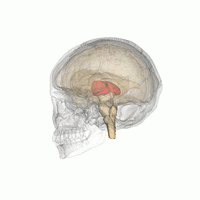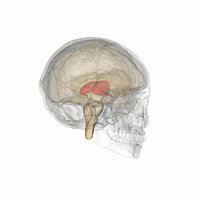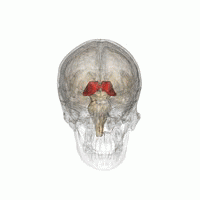 Таламус при вращении на 360°. | |
| Подробности | |
| Часть | Промежуточный мозг |
| Части | См. Список ядер таламуса. |
| Артерия | Задняя мозговая артерия и ее ветви |
| Идентификаторы | |
| латинский | таламус спинной |
| МеШ | D013788 |
| Нейроимена | 300 |
| НейроЛекс ID | бирнлекс_954 |
| ТА98 | А14.1.08.101 А14.1.08.601 |
| ТА2 | 5678 |
| ТО | E5.14.3.4.2.1.8 |
| ФМА | 62007 |
| Анатомические термины нейроанатомии | |
Таламус талам ( мн.: ; от греческого θάλαμος , «камера») [1] представляет собой большую массу серого вещества на боковых стенках третьего желудочка, образующую дорсальную часть промежуточного мозга (отдел переднего мозга ). Нервные волокна выходят из таламуса в кору головного мозга во всех направлениях, известные как таламокортикальное излучение , обеспечивая в виде концентратора обмен информацией . Он выполняет несколько функций, таких как передача сенсорных и моторных сигналов в кору головного мозга. [2] [3] и регуляция сознания , сна и бдительности . [4]
Анатомически это парамедиальная симметричная структура из двух половин (левой и правой) внутри мозга позвоночных , расположенная между корой головного мозга и средним мозгом . Он формируется во время эмбрионального развития как основной продукт промежуточного мозга , как впервые признал швейцарский эмбриолог и анатом Вильгельм Хис-старший в 1893 году. [5]
Анатомия
[ редактировать ]Таламус представляет собой парную структуру серого вещества длиной около четырех сантиметров, расположенную в переднем мозге , который превосходит средний мозг , недалеко от центра мозга, причем нервные волокна отходят к коре головного мозга во всех направлениях. Медиальная поверхность таламуса составляет верхнюю часть латеральной стенки третьего желудочка и соединяется с соответствующей поверхностью противоположного таламуса уплощенной серой полосой — межталамической спайкой . Латеральная часть таламуса является филогенетически новейшей частью таламуса (неоталамус) и включает латеральные ядра, пульвинарное , медиальное и латеральное коленчатые ядра . [6] [7] В таламусе имеются области белого вещества , включая поясной слой, покрывающий дорсальную поверхность, а также наружную и внутреннюю мозговые пластинки . Наружная пластинка покрывает латеральную поверхность, а внутренняя пластинка делит ядра на переднюю, медиальную и латеральную группы. [8]
Кровоснабжение
[ редактировать ]Кровоснабжение таламуса осуществляется из ряда артерий: полярной артерии ( задней соединительной артерии ), парамедиальных таламо-субталамических артерий, нижнелатеральных (таламогенных) артерий и задних (медиальных и латеральных) хориоидальных артерий . [9] Все это ветви задней мозговой артерии . [10]
У некоторых людей имеется артерия Першерона , которая представляет собой редкий анатомический вариант, при котором один артериальный ствол отходит от задней мозговой артерии и снабжает обе части таламуса.
Таламические ядра
[ редактировать ]



Производные промежуточного мозга включают расположенный дорсально эпиталамус (по существу хабенулу и придатки) и периталамус (преталамус), содержащий неясную зону и ретикулярное ядро таламуса . В силу различного онтогенетического происхождения эпиталамус и периталамус формально выделяют из собственно таламуса. Металамус состоит из латерального и медиального коленчатых ядер.
Таламус состоит из системы пластинок (состоящих из миелинизированных волокон ), разделяющих различные части таламуса. Другие области определяются отдельными кластерами нейронов , такими как перивентрикулярное ядро , внутриламинарные элементы, «пограничное ядро» и другие. [11] Эти последние структуры, отличающиеся по строению от основной части таламуса, были сгруппированы в аллоталамус, а не в изоталамус . [12] Это различие упрощает глобальное описание таламуса.
Соединения
[ редактировать ]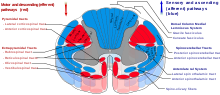
Таламус имеет множество связей с гиппокампом через маммиллоталамический тракт . Этот тракт включает маммиллярные тела и свод . [13]
Таламус связан с корой головного мозга посредством таламокортикальных излучений . [14]
Спиноталамический тракт — сенсорный путь, берущий начало в спинном мозге. Он передает в таламус информацию о боли, температуре, зуде и грубом прикосновении . Выделяют две основные части: латеральный спиноталамический тракт , который передает боль и температуру, и передний (или вентральный) спиноталамический тракт , который передает грубое прикосновение и давление.
Функция
[ редактировать ]Таламус выполняет множество функций и обычно считается, что он действует как ретрансляционная станция или концентратор , передавая информацию между различными подкорковыми областями и корой головного мозга. [15] В частности, каждая сенсорная система (за исключением обонятельной ) включает таламическое ядро, которое получает сенсорные сигналы и отправляет их в соответствующую первичную область коры. [ нужна ссылка ] Например, для зрительной системы сигналы от сетчатки передаются в латеральное коленчатое ядро таламуса, которое, в свою очередь, проецируется на зрительную кору в затылочной доле . [16] Считается, что таламус одновременно обрабатывает сенсорную информацию и передает ее — каждая из первичных сенсорных ретрансляционных областей получает сильные обратные связи от коры головного мозга. [17] Аналогичным образом, медиальное коленчатое ядро действует как ключевой слуховой ретранслятор между нижними бугорками мозга среднего и первичной слуховой корой . [ нужна ссылка ] Вентральное заднее ядро является ключевым соматосенсорным реле, которое отправляет сенсорную и проприоцептивную информацию в первичную соматосенсорную кору . [ нужна ссылка ]
Таламус также играет важную роль в регуляции состояний сна и бодрствования . [18] Ядра таламуса имеют прочные реципрокные связи с корой головного мозга, образуя таламо-кортико-таламические цепи , которые, как полагают, связаны с сознанием . [19] Таламус играет важную роль в регулировании возбуждения, уровня осознанности и активности. Повреждение таламуса может привести к постоянной коме . [20]
Роль таламуса в более передних паллидных и черных областях в нарушениях системы базальных ганглиев признана, но все еще плохо изучена. Вклад таламуса в вестибулярные и тектальные функции практически игнорируется. Таламус считался «реле», которое просто пересылает сигналы в кору головного мозга. Новые исследования показывают, что функция таламуса более избирательна. [21] Множество различных функций связаны с различными областями таламуса. Это справедливо для многих сенсорных систем (кроме обонятельной), таких как слуховая , соматическая , висцеральная , вкусовая и зрительная системы , где локализованные поражения провоцируют специфический сенсорный дефицит. Основная роль таламуса заключается в поддержке двигательной и языковой систем, и большая часть схем, задействованных в этих системах, является общей. Таламус функционально связан с гиппокампом. [22] как часть расширенной системы гиппокампа в передних ядрах таламуса [23] Что касается пространственной памяти и пространственных сенсорных данных, они имеют решающее значение для памяти эпизодических событий человека. [24] [25] Связь таламической области с средневисочной долей обеспечивает дифференциацию функционирования припоминательной и знакомой памяти. [13]
Нейрональные информационные процессы, необходимые для двигательного контроля, были предложены как сеть, включающая таламус как подкорковый двигательный центр. [26] Путем исследований анатомии мозга приматов [27] Характер взаимосвязанных тканей мозжечка с множественной моторной корой предполагает, что таламус выполняет ключевую функцию, обеспечивая специфические каналы от базальных ганглиев и мозжечка к корковым моторным областям. [28] [29] При исследовании саккады и антисаккады [30] При двигательном ответе у трех обезьян обнаружено, что области таламуса участвуют в генерации антисаккадных движений глаз (т. е. в способности тормозить рефлекторное судорожное движение глаз в направлении предъявленного стимула). [31]
Недавние исследования показывают, что медиодорсальный таламус (МД) может играть более широкую роль в познании. В частности, медиодорсальный таламус может «усиливать связность (силу сигнала) только тех цепей в коре, которые подходят для текущего контекста, и тем самым способствовать гибкости (мозга млекопитающих) для принятия сложных решений, связывая множество ассоциаций, на основе которых принимаются решения». зависят от слабо связанных корковых цепей». [32] Исследователи обнаружили, что «усиление активности МД увеличивает способность мышей «думать». [32] снижая более чем на 25 процентов частоту ошибок при принятии решения, каким противоречивым сенсорным стимулам следует следовать, чтобы найти награду». [33]
Разработка
[ редактировать ]Таламический комплекс состоит из периталамуса (или преталамуса, ранее также известного как вентральный таламус), среднего диэнцефального организатора (который позже образует zona limitans интраталамус (ZLI)) и таламуса (дорсального таламуса). [34] [35] Развитие таламуса можно разделить на три этапа. [36] Таламус — крупнейшая структура, происходящая из эмбрионального промежуточного мозга , задней части переднего мозга, расположенной между средним и большим мозгом.
Раннее развитие мозга
[ редактировать ]После нейруляции внутри нервной трубки индуцируется закладка преталамуса и таламуса. Данные, полученные от различных модельных организмов позвоночных, подтверждают модель, в которой решающее значение имеет взаимодействие между двумя факторами транскрипции , Fez и Otx. Фес экспрессируется в преталамусе, и функциональные эксперименты показывают, что Фес необходим для формирования преталамуса. [37] [38] Сзади Otx1 и Otx2 примыкают к домену экспрессии Fez и необходимы для правильного развития таламуса. [39] [40]
Формирование доменов-предшественников
[ редактировать ]На ранних этапах развития таламуса формируются два домена-предшественника: каудальный домен и ростральный домен. Каудальный домен дает начало всем глутаматергическим нейронам таламуса взрослого человека, тогда как ростральный домен дает начало всем ГАМКергическим нейронам таламуса взрослого человека. [41]
Формирование среднедиэнцефального организатора (СДО)
[ редактировать ]индуцируется средний диэнцефальный организатор (MDO, также называемый организатором ZLI) На границе между доменами экспрессии Fez и Otx внутри таламического зачатка . MDO является центральным организатором сигнализации в таламусе. Отсутствие организатора приводит к отсутствию таламуса. В процессе развития MDO созревает от вентральной к дорсальной части. Члены семейства sonic hedgehog (SHH) и семейства Wnt являются основными основными сигналами, излучаемыми MDO.
Помимо своей важности как сигнального центра, организатор созревает в морфологическую структуру интраталамической зоны (ZLI).
Созревание и парцелляция таламуса
[ редактировать ]После индукции MDO начинает управлять развитием таламической закладки путем высвобождения сигнальных молекул, таких как SHH. [42] У мышей функция передачи сигналов MDO не была затронута напрямую из-за полного отсутствия промежуточного мозга у SHH-мутантов. [43]
Исследования на цыплятах показали, что SHH необходим и достаточен для индукции таламических генов. [44] У рыбок данио было показано, что экспрессия двух генов SHH, SHH-a и SHH-b (ранее описанных как twhh), маркирует территорию MDO, и что передача сигналов SHH достаточна для молекулярной дифференциации как преталамуса, так и таламуса, но не требуется для их поддержания, а передача сигналов SHH от MDO/алярной пластинки достаточна для созревания преталамической и таламической территории, в то время как вентральные сигналы Shh необязательны. [45]
Воздействие SHH приводит к дифференцировке нейронов таламуса. Передача сигналов SHH от MDO индуцирует задне-переднюю волну экспрессии пронейрального гена Neurogenin1 в основной (каудальной) части таламуса и Ascl1 (ранее Mash1) в оставшейся узкой полосе ростральных клеток таламуса, непосредственно прилегающих к MDO. и в преталамусе. [46] [47]
Такая зональность экспрессии пронейральных генов приводит к дифференцировке глутаматергических релейных нейронов от предшественников Neurogenin1+ и ГАМКергических ингибирующих нейронов от предшественников Ascl1+. У рыб выбор этих альтернативных судеб нейротрансмиттеров контролируется динамической экспрессией Her6, гомолога HES1 . Экспрессия этого волосатоподобного транскрипционного фактора bHLH , который репрессирует нейрогенин, но необходим для Ascl1, постепенно теряется в каудальном таламусе, но сохраняется в преталамусе и в полоске ростральных клеток таламуса. Кроме того, исследования на курах и мышах показали, что блокирование пути Shh приводит к отсутствию рострального таламуса и существенному уменьшению каудального таламуса. Ростральный таламус даст начало ретикулярному ядру, в результате чего каудальный таламус сформирует релейный таламус и будет далее подразделяться на ядра таламуса . [36]
Было показано , что у людей общая генетическая вариация в промоторной области переносчика серотонина (аллель SERT-long и -short: 5-HTTLPR ) влияет на развитие нескольких областей таламуса у взрослых. Люди, унаследовавшие два коротких аллеля (SERT-ss), имеют больше нейронов и больший объем в пульвинарной и , возможно, лимбической областях таламуса. Увеличение таламуса обеспечивает анатомическую основу того, почему люди, унаследовавшие два аллеля SERT-ss, более уязвимы к большой депрессии , посттравматическому стрессовому расстройству и самоубийству. [48]
Клиническое значение
[ редактировать ]Таламус, поврежденный инсультом, может привести к таламическому болевому синдрому . [49] которое включает в себя одностороннее жжение или боль, часто сопровождающееся перепадами настроения . Двусторонняя ишемия области, снабжаемой парамедианной артерией, может вызвать серьезные проблемы, включая акинетический мутизм , и сопровождаться глазодвигательными проблемами. Родственное понятие – таламокортикальная аритмия . Окклюзия артерии Першерона может привести к двустороннему инфаркту таламуса.
Алкогольный синдром Корсакова возникает в результате повреждения маммиллярного тела , маммиллоталамического пучка или таламуса. [50] [51]
Фатальная семейная бессонница — наследственное прионное заболевание, при котором происходит дегенерация таламуса, в результате чего пациент постепенно теряет способность спать и прогрессирует до состояния тотальной бессонницы , которая неизменно приводит к смерти. Напротив, повреждение таламуса может привести к коме.
Атрофия таламуса является показателем рассеянного склероза . [52] [53]
Дополнительные изображения
[ редактировать ] Рассечение человеческого мозга, показывающее таламус.
Рассечение человеческого мозга, показывающее таламус. Таламус человека вместе с другими подкорковыми структурами в стеклянном мозге.
Таламус человека вместе с другими подкорковыми структурами в стеклянном мозге. Латеральная группа ядер таламуса.
Латеральная группа ядер таламуса. Медиальная группа таламических ядер.
Медиальная группа таламических ядер.




См. также
[ редактировать ]- 5-НТ7-рецептор
- Криста и Татьяна Хоган — сиамские близнецы со сросшимися таламами.
- Список регионов человеческого мозга
- Немоторная область вентральной ядерной группы таламуса.
- Nucleus ventralis posterior Lateris pars oralis (VPLo), область таламуса.
- Система базальных ганглиев приматов
- Таламический стимулятор
- Таламотомия
Ссылки
[ редактировать ]- ^ Харпер - индекс. Архивировано 12 мая 2015 г. на Wayback Machine и Вашингтонского университета. веб-сервере факультета Архивировано 2 марта 2015 г. на странице поиска Wayback Machine и поисковой системе. Архивировано 27 сентября 2020 г. на Wayback Machine + проекта Perseus пучки . .edu. Архивировано 5 марта 2021 г. на Wayback Machine. Проверено 9 февраля 2012 г.
- ^ Шерман, С. (2006). «Таламус» . Схоларпедия . 1 (9): 1583. Бибкод : 2006SchpJ...1.1583S . doi : 10.4249/scholarpedia.1583 .
- ^ Шерман, С. Мюррей; Гиллери, RW (2000). Исследование Таламуса . Академическая пресса. стр. 1–18. ISBN 978-0-12-305460-9 .
- ^ Горветт, Зария. «Чему можно научиться из необычных привычек Эйнштейна» . bbc.com .
- ^ Джонс, Эдвард Дж., изд. (1985). Таламус-Спрингер . дои : 10.1007/978-1-4615-1749-8 . ISBN 978-1-4613-5704-9 . S2CID 41337319 .
- ^ «Медицинское определение НЕОТАЛАМУСА» . www.merriam-webster.com .
- ^ «Неоталамус | Определение неоталамуса на английском языке в Оксфордских словарях» . Оксфордские словари | Английский . Архивировано из оригинала 27 мая 2018 года.
- ^ Тортора, Жерар; Анагностакос, Николас (1987). Принципы анатомии и физиологии (5-е издание Harper International Ed.). Нью-Йорк: Харпер и Роу. п. 314 . ISBN 978-0060466695 .
- ^ Першерон, Г. (1982). «Артериальное кровоснабжение таламуса». В Шальтенбранде; Уокер, А.Э. (ред.). Стереотаксия головного мозга человека . Штутгарт: Тиме. стр. 218–32.
- ^ Найп, Х. Джонс, Дж. и др. Таламус http://radiopaedia.org/articles/thalamus. Архивировано 17 сентября 2017 г. в Wayback Machine.
- ^ Джонс Эдвард Г. (2007) «Таламус» Кембриджского университета. Нажимать [ нужна страница ]
- ^ Першерон, Г. (2003). «Таламус». В Паксиносе, Г.; Мэй, Дж. (ред.). Нервная система человека (2-е изд.). Амстердам: Эльзевир. стр. 592–675.
- ^ Перейти обратно: а б Карлесимо, Джорджия; Ломбарди, МГ; Кальтаджироне, К. (2011). «Сосудистая таламическая амнезия: переоценка». Нейропсихология . 49 (5): 777–89. doi : 10.1016/j.neuropsychologia.2011.01.026 . ПМИД 21255590 . S2CID 22002872 .
- ^ Вашингтонский университет (1991). «Таламокортикальное излучение» . Вашингтон.edu.
- ^ Газзанига; Иври; Мангун, Майкл, С.; Ричард Б.; Джордж Р. (2014). Когнитивная нейронаука — Биология разума . Нью-Йорк: WW Нортон. стр. 45 . ISBN 978-0-393-91348-4 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Шин, Лиза М; Либерзон, Израиль (январь 2010 г.). «Нейросхема страха, стресса и тревожных расстройств» . Нейропсихофармакология . 35 (1): 169–191. дои : 10.1038/нпп.2009.83 . ПМК 3055419 . ПМИД 19625997 .
- ^ «Таламус, посредник мозга, становится сенсорным проводником» . Медицинский университет Чикаго . Проверено 10 сентября 2020 г.
- ^ Стериаде, Мирча; Ллинас, Родольфо Р. (1988). «Функциональные состояния таламуса и связанное с ним взаимодействие нейронов». Физиологические обзоры . 68 (3): 649–742. дои : 10.1152/physrev.1988.68.3.649 . ПМИД 2839857 .
- ^ Кома и расстройства сознания ISBN 978-1-447-12439-9 стр. 143
- ^ Неврология сознания: когнитивная нейронаука и невропатология ISBN 978-0-123-74168-4 стр. 10
- ^ Леонард, Эбигейл В. (17 августа 2006 г.). «Ваш мозг загружается как компьютер» . ЖиваяНаука .
- ^ Штейн, Тор; Мориц, Чад; Куигли, Мишель; Кордес, Дитмар; Хотон, Виктор; Мейеранд, Элизабет (2000). «Функциональная связь таламуса и гиппокампа, изученная с помощью функциональной магнитно-резонансной томографии» . Американский журнал нейрорадиологии . 21 (8): 1397–401. ПМЦ 7974059 . ПМИД 11003270 .
- ^ Агглтон, Джон П.; Браун, Малкольм В. (1999). «Эпизодическая память, амнезия и ось гиппокамп – передняя таламуса» (PDF) . Поведенческие и мозговые науки . 22 (3): 425–44, обсуждение 444–89. дои : 10.1017/S0140525X99002034 . ПМИД 11301518 . S2CID 11258997 .
- ^ Агглтон, Джон П.; О'Мара, Шейн М.; Ванн, Сералин Д.; Райт, Ник Ф.; Цанов, Мариан; Эриксен, Джонатан Т. (2010). «Гиппокампально-передние таламические пути памяти: раскрытие сети прямых и косвенных действий» . Европейский журнал неврологии . 31 (12): 2292–307. дои : 10.1111/j.1460-9568.2010.07251.x . ПМК 2936113 . ПМИД 20550571 .
- ^ Берджесс, Нил; Магуайр, Элеонора А; О'Киф, Джон (2002). «Человеческий гиппокамп и пространственная и эпизодическая память» . Нейрон . 35 (4): 625–41. дои : 10.1016/S0896-6273(02)00830-9 . ПМИД 12194864 . S2CID 11989085 .
- ^ Эвартс, Е.В.; Тах, WT (1969). «Двигатели ЦНС: церебро-церебеллярные взаимоотношения». Ежегодный обзор физиологии . 31 : 451–98. дои : 10.1146/annurev.ph.31.030169.002315 . ПМИД 4885774 .
- ^ Ориоли, П.Дж.; Стрик, PL (1989). «Связи мозжечка с моторной корой и дугообразной премоторной областью: анализ с использованием ретроградного транснейронального транспорта WGA-HRP». Журнал сравнительной неврологии . 288 (4): 612–26. дои : 10.1002/cne.902880408 . ПМИД 2478593 . S2CID 27155579 .
- ^ Асанума С., Тач В.Т., Джонс Э.Г. (май 1983 г.). «Цитоархитектоническое очертание вентрально-латеральной области таламуса у обезьяны». Исследования мозга . 286 (3): 219–35. дои : 10.1016/0165-0173(83)90014-0 . ПМИД 6850357 . S2CID 25013002 .
- ^ Курата, К. (2005). «Свойства активности и расположение нейронов моторного таламуса, которые проецируются в кортикальные моторные области обезьян». Журнал нейрофизиологии . 94 (1): 550–66. дои : 10.1152/jn.01034.2004 . ПМИД 15703228 .
- ^ «Антисаккада — обзор фундаментальных и клинических исследований» . Архивировано из оригинала 16 сентября 2017 г. Проверено 10 февраля 2012 г. [ нужна полная цитата ]
- ^ Кунимацу, Дж; Танака, М. (2010). «Роль моторного таламуса приматов в генерации антисаккад» (PDF) . Журнал неврологии . 30 (14): 5108–17. doi : 10.1523/JNEUROSCI.0406-10.2010 . ПМК 6632795 . ПМИД 20371831 .
- ^ Перейти обратно: а б «Открыта новая роль области мозга» . Новости неврологии . 03 мая 2017 г. Проверено 3 декабря 2017 г.
- ^ Шмитт, Л. Ян; Виммер, Ральф Д.; Накадзима, Михо; Хэпп, Майкл; Мофахам, Сима; Халасса, Майкл М. (май 2017 г.). «Таламусное усиление корковых связей поддерживает контроль внимания» . Природа . 545 (7653): 219–223. Бибкод : 2017Natur.545..219S . дои : 10.1038/nature22073 . ПМК 5570520 . ПМИД 28467827 .
- ^ Куленбек, Хартвиг (1937). «Онтогенетическое развитие диэнцефальных центров мозга птицы (цыпленка) и сравнение с промежуточным мозгом рептилий и млекопитающих». Журнал сравнительной неврологии . 66 : 23–75. дои : 10.1002/cne.900660103 . S2CID 86730019 .
- ^ Шимамура, К; Хартиган, диджей; Мартинес, С; Пуэльес, Л; Рубинштейн, Дж. Л. (1995). «Продольная организация передней нервной пластинки и нервной трубки». Разработка . 121 (12): 3923–33. дои : 10.1242/dev.121.12.3923 . ПМИД 8575293 .
- ^ Перейти обратно: а б Шольпп, Штеффен; Ламсден, Эндрю (2010). «Строительство свадебного чертога: Развитие таламуса» . Тенденции в нейронауках . 33 (8): 373–80. дои : 10.1016/j.tins.2010.05.003 . ПМЦ 2954313 . ПМИД 20541814 .
- ^ Хирата, Т.; Наказава, М; Мураока, О; Накаяма, Р.; Пот, Y; Хиби, М. (2006). «Гены цинковых пальцев Fez и Fez-подобные функции участвуют в формировании подразделений промежуточного мозга». Развитие 133 (20): 3993–4004. дои : 10.1242/dev.02585 . ПМИД 16971467 .
- ^ Чон, Ж.-Ю.; Эйнхорн, З.; Матур, П.; Чен, Л.; Ли, С.; Каваками, К.; Го, С. (2007). «Формирование промежуточного мозга рыбок данио с помощью консервативного белка цинковых пальцев Fezl» . Разработка . 134 (1): 127–36. дои : 10.1242/dev.02705 . ПМИД 17164418 .
- ^ Акампора, Д; Авантаджиато, В; Туорто, Ф; Симеоне, А (1997). «Генетический контроль морфогенеза головного мозга посредством требования дозировки гена Otx». Разработка . 124 (18): 3639–50. дои : 10.1242/dev.124.18.3639 . ПМИД 9342056 .
- ^ Шолпп, С.; Фуше, И.; Штаудт, Н.; Пойкерт, Д.; Ламсден, А.; Уарт, К. (2007). «Otx1l, Otx2 и Irx1b устанавливают и позиционируют ZLI в промежуточном мозге» . Разработка . 134 (17): 3167–76. дои : 10.1242/dev.001461 . ПМК 7116068 . ПМИД 17670791 .
- ^ Сон, Хобум; Ли, Бамви; Пюн, Дохун; Гимера, Хорди; Сын, Янгсук; Юн, Джесын; Пэк, Кванхи; Вурст, Вольфганг; Чон, Ёнсу (февраль 2015 г.). «Ascl1 и Helt действуют комбинаторно, определяя идентичность таламических нейронов, подавляя активацию Dlxs» . Биология развития . 398 (2): 280–291. дои : 10.1016/j.ydbio.2014.12.003 . ПМИД 25512300 .
- ^ Пуэльес, Л; Рубинштейн, Дж. Л. (2003). «Домены экспрессии генов переднего мозга и развивающаяся просомерная модель». Тенденции в нейронауках . 26 (9): 469–76. дои : 10.1016/S0166-2236(03)00234-0 . ПМИД 12948657 . S2CID 14658562 .
- ^ Ишибаси, М; МакМахон, AP (2002). «Звуковое сигнальное реле, зависимое от ежа, регулирует рост диэнцефальных и мезэнцефальных зачатков у ранних эмбрионов мышей». Разработка . 129 (20): 4807–19. дои : 10.1242/dev.129.20.4807 . ПМИД 12361972 .
- ^ Кикер, К; Ламсден, А. (2004). «Передача сигналов Hedgehog от ZLI регулирует диэнцефальную региональную идентичность». Природная неврология . 7 (11): 1242–9. дои : 10.1038/nn1338 . ПМИД 15494730 . S2CID 29863625 .
- ^ Шолпп, С.; Вольф, О; Бренд, М; Ламсден, А. (2006). «Передача сигналов ежей из zona limitans Intrathalamica управляет формированием паттерна промежуточного мозга рыбок данио» . Разработка . 133 (5): 855–64. дои : 10.1242/dev.02248 . ПМИД 16452095 .
- ^ Шолпп, С.; Делогу, А.; Гилторп, Дж.; Пойкерт, Д.; Шиндлер, С.; Ламсден, А. (2009). «Her6 регулирует нейрогенетический градиент и идентичность нейронов в таламусе» . Труды Национальной академии наук . 106 (47): 19895–900. Бибкод : 2009PNAS..10619895S . дои : 10.1073/pnas.0910894106 . ПМЦ 2775703 . ПМИД 19903880 .
- ^ Вуэ, Тоу Ия; Блуске, Криста; Алишахи, Амин; Ян, Линь Линь; Кояно-Накагава, Наоко; Нович, Беннетт; Накагава, Ясуси (2009). «Сигнализация Sonic Hedgehog контролирует идентичность таламических предшественников и спецификацию ядер у мышей» . Журнал неврологии . 29 (14): 4484–97. doi : 10.1523/JNEUROSCI.0656-09.2009 . ПМЦ 2718849 . ПМИД 19357274 .
- ^ Янг, Кейт А.; Холкомб, Ли А.; Бонкале, Вилли Л.; Хикс, Пол Б.; Яздани, Умар; Герман, Дуайт К. (2007). «Полиморфизм 5HTTLPR и увеличение пульвинара: открытие черного хода в лимбическую систему». Биологическая психиатрия . 61 (6): 813–8. doi : 10.1016/j.biopsych.2006.08.047 . ПМИД 17083920 . S2CID 2214561 .
- ^ Дежерин, Дж.; Русси, Г. (1906). «Таламический синдром». Неврологический обзор . 14 :521–32.
- ^ Копельман, доктор медицинских наук; Томсон, AD; Геррини, я; Маршалл, Э.Дж. (16 января 2009 г.). «Синдром Корсакова: клинические аспекты, психология и лечение». Алкоголь и алкоголизм . 44 (2): 148–54. дои : 10.1093/alcalc/agn118 . ПМИД 19151162 .
- ^ Раме, Р; Мусса, Р; Авада, А; Ибрагим, я; Али, Ю; Мааррави, Дж; Ризк, Т; Нора, Дж; Окайс, Н; Самаха, Э. (апрель 2007 г.). «Острый корсаковский амнестический синдром, возникающий в результате инфаркта левого таламуса после кровоизлияния в правый гиппокамп» . АДЖНР. Американский журнал нейрорадиологии . 28 (4): 759–60. ПМЦ 7977335 . ПМИД 17416834 .
- ^ Векслер, Мариса (20 октября 2023 г.). «Модели машинного обучения позволяют оценить, когда при рассеянном склерозе начинается атрофия мозга» . Новости рассеянного склероза сегодня .
- ^ Цен, Стивен; Гебрегзиабхер, Мулугета; Моазами, Саид; Азеведо, Кристина Дж.; Пеллетье, Даниэль (28 сентября 2023 г.). «На пути к прецизионной медицине с использованием подхода «цифрового двойника»: моделирование возникновения специфической атрофии головного мозга у людей с рассеянным склерозом» . Научные отчеты . 13 (1): rs.3.rs–2833532. Бибкод : 2023NatSR..1316279C . дои : 10.1038/s41598-023-43618-5 . ПМЦ 10187410 . ПМИД 37205476 .