Отолит
Эта статья может быть слишком технической для понимания большинства читателей . ( Март 2021 г. ) |
| Отолит | |
|---|---|
 Отолитовые органы, показывающие детали маточки, отокония, эндолимфы, купулы, макулы, нитей волосковых клеток и мешотчатого нерва. | |
 Молодь сельди . Длина 30 мм; 3 месяца; все еще прозрачный; отолиты видны слева от глаза. | |
| Подробности | |
| Идентификаторы | |
| латинский | статоконий |
| ТА98 | А15.3.03.086 |
| ФМА | 77826 |
| Анатомическая терминология | |
Отолит отоконием ( греч . ὠτο- , ōto- ear + λῐ́θος , líthos , камень), также называемый статоконием , . или статолитом , представляет собой структуру карбоната кальция в мешочке или матке внутреннего уха , особенно в вестибулярной системе позвоночных животных . Мешочек и маточка, в свою очередь, вместе образуют отолитовые органы . Именно эти органы позволяют организму, включая человека, воспринимать линейное ускорение , как горизонтальное, так и вертикальное (гравитация). Они были идентифицированы как у вымерших, так и у современных позвоночных. [1]
Подсчет годовых колец на отолитах — распространенный метод оценки возраста рыб . [2]
Описание
[ редактировать ]Эндолимфатические наполнения, такие как отолиты, представляют собой структуры в мешочке и матке внутреннего уха , особенно в вестибулярном лабиринте всех позвоночных животных (рыб, амфибий, рептилий, млекопитающих и птиц). У позвоночных мешочек и маточка вместе образуют отолитовые органы . И статоконии, и отолиты используются в качестве индикаторов силы тяжести, равновесия, движения и направления у всех позвоночных и имеют второстепенную функцию при обнаружении звука у высших водных и наземных позвоночных. [3] [4] Они чувствительны к гравитации и линейному ускорению . Из-за своей ориентации в голове маточка чувствительна к изменению горизонтального движения, а мешочек дает информацию о вертикальном ускорении (например, в лифте ) .
Подобные рецепторы баланса, называемые статоцистами, можно найти у многих групп беспозвоночных , но они не содержатся в структуре внутреннего уха. Статоцисты моллюсков имеют морфологию , аналогичную чувствительным к смещению органам позвоночных животных; [5] однако функция статоциста моллюска ограничивается обнаружением силы тяжести и, возможно, некоторым обнаружением углового момента. [6] Это аналогичные структуры , имеющие схожую форму и функции, но не произошедшие от общей структуры .
Статоконии (также называемые отокониями) представляют собой многочисленные зерна, часто сферической формы, размером от 1 до 50 мкм ; коллективно. [ нужна ссылка ] Статоконии также иногда называют статоцистами. Отолиты (также называемые статолитами) представляют собой агглютинированные кристаллы или кристаллы, осаждающиеся вокруг ядра, с четко определенной морфологией, и все вместе их можно назвать эндолимфатическими наполнителями. [1] [7] [8]
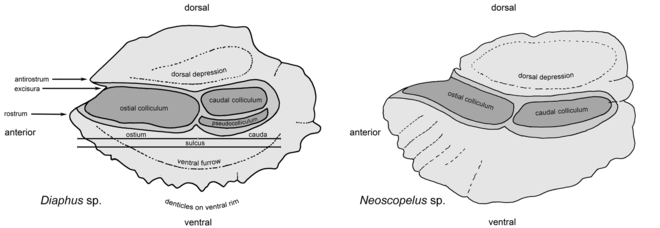
и неоскопелид отолиты ( Neoscopelus , правая сторона) [9]
Механизм
[ редактировать ]Полукружные каналы и мешочки у всех позвоночных прикрепляются к эндолимфатическим протокам, которые у некоторых групп (например, у акул ) заканчиваются небольшими отверстиями, называемыми эндолимфатическими порами, на дорсальной поверхности головы. [1] Через эти отверстия могут проникнуть посторонние зерна, обычно диаметром менее миллиметра. Размер поступающего материала ограничен частицами размером с песок, а в случае акул он связан эндогенной органической матрицей, которую выделяет животное.
У млекопитающих отолиты представляют собой мелкие частицы, состоящие из сочетания студенистого матрикса и карбоната кальция в вязкой жидкости мешочка и матрикса. Вес и инерция этих маленьких частиц заставляют их стимулировать волосковые клетки при движении головы. Волосковые клетки состоят из 40–70 стереоцилий и одной киноцилии , которая связана с афферентным нервом. Волосковые клетки посылают сигналы по сенсорным нервным волокнам , которые интерпретируются мозгом как движение. Помимо определения ускорения головы, отолиты могут помочь определить ориентацию посредством воздействия на них силы тяжести. Когда голова находится в нормальном вертикальном положении, отолит давит на сенсорные рецепторы волосковых клеток. Это толкает отростки волосковых клеток вниз и предотвращает их перемещение из стороны в сторону. Однако когда голова наклонена, сила тяжести отолитов смещает отростки волосковых клеток в сторону, искажая их и посылая в центральную нервную систему сигнал о том, что голова наклонена.
Имеются данные о том, что вестибулярная система млекопитающих сохранила часть своей наследственной акустической чувствительности и что эта чувствительность опосредована отолитическими органами (вероятнее всего, саккулюсом , в силу его анатомического расположения). У мышей, у которых отсутствуют отоконии маточки и мешочка, сохраненная акустическая чувствительность теряется. [4] У человека вестибулярные вызванные миогенные потенциалы возникают в ответ на громкую низкочастотную акустическую стимуляцию у пациентов с нейросенсорной тугоухостью. [3] Также предполагалось, что вестибулярная чувствительность к ультразвуковым звукам участвует в восприятии речи, представленной на искусственно высоких частотах, превышающих диапазон улитки человека (~ 18 кГц). [10] Было продемонстрировано, что у мышей ощущение акустической информации через вестибулярную систему оказывает поведенчески значимый эффект; Реакция на вызванный акустический рефлекс испуга сильнее при наличии громких низкочастотных звуков, которые ниже порога для улитки мыши (~ 4 Гц), что повышает вероятность того, что акустическая чувствительность вестибулярной системы может расширить диапазон слышимости мелкие млекопитающие. [4]
Палеонтология
[ редактировать ]После смерти и разложения рыбы отолиты могут сохраняться внутри тела организма или рассеиваться до захоронения и окаменения . Рассеянные отолиты — одни из многих микроокаменелостей , которые можно обнаружить с помощью микропалеонтологического анализа мелких отложений. Их стратиграфическое значение минимально, но все же может быть использовано для характеристики уровня или интервала. Ископаемые отолиты редко встречаются in situ (на останках животного), вероятно, потому, что они не распознаются отдельно от окружающей матрицы породы. В некоторых случаях их можно идентифицировать из-за различий в цвете, размере зерен или отличительной форме. Эти редкие случаи имеют особое значение, поскольку наличие, состав и морфология материала могут уточнить родство видов и групп. В случае с примитивными рыбами различные ископаемые материалы показывают, что эндолимфатические заполнения по элементному составу были похожи на матрицу породы, но были ограничены крупнозернистым материалом, который, по-видимому, лучше подходит для обнаружения гравитации, смещения и звука. Наличие этих внешних зерен в остеостраканс , хондрихтианы и акантодии указывают на общую физиологию внутреннего уха и наличие открытых эндолимфатических протоков. [1]
Неклассифицированная окаменелость под названием Gluteus minimus считалась возможными отолитами, но до сих пор неизвестно, какому животному они могли принадлежать.
Экология
[ редактировать ]Состав
[ редактировать ]Состав отолитов рыб также оказался полезным для ученых-рыболовов. Карбонат кальция, из которого состоит отолит, в основном выделяется из воды. По мере роста отолита образуются новые кристаллы карбоната кальция. Как и в любой кристаллической структуре, во время образования кристаллов будут существовать вакансии в решетке, позволяющие микроэлементам из воды связываться с отолитом. Изучение микроэлементного состава или изотопных характеристик микроэлементов в отолитах рыб дает представление о водоемах, которые рыбы ранее занимали. [11] Отолиты рыб возрастом 172 миллиона лет использовались для изучения среды, в которой жили рыбы. [12] Роботизированные устройства микроизмельчения также использовались для восстановления записей истории жизни рыб с очень высоким разрешением, включая диету и температуру на протяжении всей жизни рыб, а также их натальное происхождение. [13]
Наиболее изученными следами и изотопными признаками являются стронций из-за того же заряда и ионного радиуса , что и кальций ; однако ученые могут изучить несколько микроэлементов в отолите, чтобы различить более конкретные признаки. Обычным инструментом, используемым для измерения микроэлементов в отолите, является масс-спектрометр с лазерной абляцией и индуктивно связанной плазмой . Этот инструмент может одновременно измерять различные микроэлементы. масс- спектрометр вторичных ионов Также можно использовать . Этот прибор может обеспечить более высокое химическое разрешение, но может измерять только один микроэлемент за раз. Целью этого исследования является предоставление учёным ценной информации о местах, где обитала рыба. В сочетании с отолитовыми кольцами ученые могут определить, сколько лет было рыбам, когда они путешествовали по разным водоемам. Эту информацию можно использовать для определения жизненного цикла рыб, чтобы учёные-рыболовы могли принимать более обоснованные решения относительно рыбных запасов.
Скорость роста и возраст
[ редактировать ]

Рыбы (класс Osteichthyes ) имеют три пары отолитов — сагитты (единственное число сагитта), лапилли (единственное число лапиллусов) и астериски (единственное число звездочек). Сагитты самые крупные, находятся сразу за глазами и примерно на одном уровне с ними по вертикали. Лапилли и астериски (самые маленькие из трех) расположены внутри полукружных каналов. Сагитты обычно состоят из арагонита (хотя могут встречаться и ватеритовые аномалии). [14] ), как и лапилли, а астериски обычно состоят из ватерита.
Форма и пропорциональные размеры отолитов варьируются в зависимости от вида рыб. В целом, рыбы из высокоструктурированных местообитаний, таких как рифы или каменистое дно (например, окуни , груперы , многие барабанщики и горбыли ), будут иметь более крупные отолиты, чем рыбы, которые проводят большую часть своего времени, плавая на высокой скорости по прямым линиям в открытом океане (например, тунец , скумбрия , дельфин ). У летающих рыб необычно большие отолиты, возможно, из-за того, что им необходимо сохранять равновесие, когда они выпрыгивают из воды, чтобы «летать» в воздухе. Часто вид рыб можно определить по различным морфологическим характеристикам изолированного отолита.
образуют слои карбоната кальция Отолиты рыб на протяжении всей жизни и студенистого матрикса. Скорость прироста варьируется в зависимости от роста рыбы – часто меньше роста зимой и больше летом – что приводит к появлению колец, напоминающих годичные кольца . Пересчитав кольца, можно определить возраст рыбы в годах. [15] Обычно используется сагитта, так как она самая большая. [16] но иногда применяют лапилли, если они имеют более удобную форму. Звездочка, самая маленькая из трех, редко используется в исследованиях возраста и роста.
Кроме того, у большинства видов нарастание карбоната кальция и студенистого матрикса чередуется в суточном цикле. Поэтому также возможно определить возраст рыбы в днях. [17] Последняя информация часто получается под микроскопом и предоставляет важные данные для исследований истории ранней жизни.
Предполагалось (по крайней мере, у некоторых видов), что измеряя толщину отдельных колец, можно оценить рост рыбы, поскольку рост рыбы прямо пропорционален росту отолитов. [18] Однако некоторые исследования опровергают прямую связь между ростом тела и ростом отолитов. В периоды низкого или нулевого роста тела отолит продолжает срастаться, что заставляет некоторых исследователей полагать, что прямая связь связана с метаболизмом, а не с ростом как таковым. Отолиты, в отличие от чешуи, не реабсорбируются в периоды снижения энергии, что делает их еще более полезным инструментом для старения рыбы. Рыба никогда полностью не перестает расти, хотя скорость роста взрослой рыбы снижается. В результате кольца, соответствующие более поздним этапам жизненного цикла, имеют тенденцию быть ближе друг к другу. Более того, небольшой процент отолитов у некоторых видов со временем деформируется. [19]
Исследования возраста и роста рыб важны для понимания таких вещей, как время и масштабы нереста, пополнение и использование среды обитания, продолжительность жизни личинок и молоди, а также возрастная структура популяции . Такие знания, в свою очередь, важны для разработки соответствующей рыболовством политики управления . Из-за большого количества человеческого труда, необходимого для определения возраста отолитов, ведутся активные исследования по автоматизации этого процесса. [20]
Исследование диеты
[ редактировать ]Поскольку соединения в отолитах рыб устойчивы к пищеварению , они обнаруживаются в пищеварительном тракте и помете морских птиц и рыбоядных морских млекопитающих , таких как дельфины , тюлени , морские львы и моржи . многих рыб можно определить Род и вид по отолитам. Таким образом, отолиты могут в некоторой степени использоваться для определения и реконструкции состава добычи морских млекопитающих и рационов морских птиц.
Отолиты (сагитты) двусторонне симметричны , у каждой рыбы есть один правый и один левый. Таким образом, разделение восстановленных отолитов на правые и левые позволяет сделать вывод о минимальном количестве съеденных особей добычи для данного вида рыб. Размер отолита также пропорционален длине и весу рыбы. Таким образом, их можно использовать для обратного расчета размера и биомассы добычи , что полезно при попытке оценить потребление добычи морскими млекопитающими и потенциальное воздействие на рыбные запасы . [21]
Однако отолиты нельзя использовать отдельно для надежной оценки рациона китообразных или ластоногих . У них может возникнуть частичная или полная эрозия пищеварительного тракта, что приводит к искажению показателей количества и биомассы добычи . [22] Виды с хрупкими, легко перевариваемыми отолитами могут быть недооценены в рационе. Чтобы устранить эти предубеждения, в ходе экспериментов по кормлению в неволе были разработаны поправочные коэффициенты отолитов, в которых тюленей кормят рыбой известного размера, а степень эрозии отолитов количественно определяется для различных таксонов добычи . [23]
Включение рыбных позвонков , челюстных костей, зубов и других информативных элементов скелета улучшает идентификацию и количественную оценку добычи по сравнению с одним лишь анализом отолитов. [24] Это особенно справедливо для видов рыб с хрупкими отолитами, но другими отличительными костями, таких как атлантическая скумбрия ( Scombrus scombrus ) и атлантическая сельдь ( Clupea harengus ). [25]
Отолитовые украшения
[ редактировать ]Украшения «морские драгоценности» из отолитов рыб недавно появились на рынке Индии благодаря усилиям группы энтузиастов-рыбац из Вижинджама . ученые Центрального научно-исследовательского института морского рыболовства Этих женщин-рыболовов обучили (ЦМФРИ). Украшения из рыбьих отолитов, известные римлянам и египтянам как счастливые камни, продолжают использоваться в таких странах, как Бразилия и Фаэрер, а в Индии их собирают и продают организованным и устойчивым образом. [26]
См. также
[ редактировать ]- косточки
- Отолитовая мембрана
- Микрохимический анализ отолита
- Орбитальная лягушка-отолит , космическая миссия 1970 года.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д Сахни, Сарда; Уилсон, Марк В.Х. (2001). «Внешние заполнения лабиринта подразумевают открытые эндолимфатические протоки у остеостраканов нижнего девона, акантодов и предполагаемых хондрихтианов». Журнал палеонтологии позвоночных . 21 (4): 660–669. doi : 10.1671/0272-4634(2001)021[0660:ELIIOE]2.0.CO;2 . S2CID 86182956 .
- ^ Гордон, ДП (2009). Инвентаризация биоразнообразия Новой Зеландии: 1. Царство животных: Radiata, Lophotrochozoa, Deuterostomia . Крайстчерч: Издательство Кентерберийского университета. п. 459. ИСБН 978-1-877257-72-8 .
- ^ Перейти обратно: а б Шейхолеслами, Киануш; Кага, Кимитака (2002). «Отолитовый орган как рецептор вестибулярного слуха, выявленный с помощью вестибулярно-вызванных миогенных потенциалов у пациентов с аномалиями внутреннего уха». Исследование слуха . 165 (1–2): 62–67. дои : 10.1016/S0378-5955(02)00278-2 . ISSN 0378-5955 . ПМИД 12031516 . S2CID 23530754 .
- ^ Перейти обратно: а б с Джонс, Гарет П.; Лукашкина Виктория Александровна; Рассел, Ян Дж.; Лукашкин, Андрей Н. (2010). «Вестибулярная система опосредует ощущение низкочастотных звуков у мышей» . Журнал Ассоциации исследований в области отоларингологии . 11 (4): 725–732. дои : 10.1007/s10162-010-0230-7 . ISSN 1525-3961 . ПМЦ 2975890 . ПМИД 20821033 .
- ^ Голди, RW (1993). «Полиморфная кристаллическая структура отолитов рыб». Журнал морфологии . 218 (1): 1–28. дои : 10.1002/jmor.1052180102 . ПМИД 29865482 . S2CID 46932904 .
- ^ Вольф, Хайнц Г. (1973). «Разнонаправленная чувствительность рецепторных клеток статоцистов заднежаберных брюхоногих моллюсков Aplysia limacina ». Морское поведение и физиология . 1 (1–4): 361–373. дои : 10.1080/10236247209386910 .
- ^ Нольф, Д. 1985. Otolithi Piscium; в Х.-П. Шульце (редактор), Справочник по палеоихтиологии, Том. 10. Густав Фишер Верлаг, Штутгарт, 145 стр.
- ^ Шульце, Х.-П. (1990). «Новый акантодий из пенсильвании штата Юта, США и распространение отолитов у челюстноротых». Журнал палеонтологии позвоночных . 10 (1): 49–58. дои : 10.1080/02724634.1990.10011789 .
- ^ Шваржанс, Вернер; Карневале, Джорджио (19 марта 2021 г.). «Рост доминирования рыб-фонарей (Teleostei: Myctophidae) в океанических экосистемах: палеонтологическая перспектива» . Палеобиология . 47 (3). Издательство Кембриджского университета (CUP): 446–463. дои : 10.1017/pab.2021.2 . ISSN 0094-8373 . S2CID 233678539 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Ленхардт, М.; Скеллетт, Р; Ван, П; Кларк, А. (1991). «Ультразвуковое восприятие речи человека». Наука . 253 (5015): 82–85. Бибкод : 1991Sci...253...82L . дои : 10.1126/science.2063208 . ISSN 0036-8075 . ПМИД 2063208 .
- ^ Паттерсон, Уильям П.; Смит, Джеральд Р.; Ломанн, Кайгер К. (1993). «Континентальная палеотермометрия и сезонность с использованием изотопного состава арагонитовых отолитов пресноводных рыб». Изменение климата в континентальных изотопных записях . Серия геофизических монографий. Том. 78. стр. 191–202. Бибкод : 1993GMS....78..191P . дои : 10.1029/GM078p0191 . ISBN 9781118664025 .
- ^ Паттерсон, Уильям П. (1999). «Старейшие отолиты рыб, охарактеризованные изотопами, дают представление о юрском континентальном климате Европы» . Геология . 27 (3): 199. Бибкод : 1999Geo....27..199P . doi : 10.1130/0091-7613(1999)027<0199:OICFOP>2.3.CO;2 .
- ^ Заццо, А.; Смит, Греция; Паттерсон, В.П.; Дюфур, Э. (2006). «Реконструкция истории жизни современной и ископаемых нерк ( Oncorhynchus nerka ) с помощью изотопного анализа кислорода отолитов, позвонков и зубов: значение для палеоэкологических реконструкций» . Письма о Земле и планетологии . 249 (3/4): 200–215. дои : 10.1016/j.epsl.2006.07.003 .
- ^ Реймер, Т.; Демпстер, Т.; Уоррен-Майерс, Ф.; Дженсен, Эй Джей; Свирер, SE (2016). «Высокая распространенность ватерита в сагиттальных отолитах вызывает ухудшение слуха у выращиваемой рыбы» . Научные отчеты . 6 : 25249. Бибкод : 2016NatSR...625249R . дои : 10.1038/srep25249 . ПМЦ 4848507 . ПМИД 27121086 .
- ^ Кимура, Дэниел К.; Андерл, Делса М. (2005). «Контроль качества данных о возрасте в Научном центре рыболовства Аляски» (PDF) . Морские и пресноводные исследования . 56 (5): 783–789. дои : 10.1071/mf04141 .
- ^ Otoliths Возраст и рост рыбы с помощью Агентства ресурсов дикой природы штата Теннесси . Проверено 7 апреля 2007 г.
- ^ Бос, Арканзас (1999). «Приливный транспорт личинок камбалы ( Pleuronectes flesus ) по реке Эльбе, Германия» . Архив рыболовства и морских исследований . 47 (1): 47–60.
- ^ Джонс, Синтия М. (1992). «Разработка и применение методики приращения отолитов». В Д.К. Стивенсоне; SE Кампана (ред.). Исследование и анализ микроструктуры отолитов . Канадское специальное издание по рыболовству и водным наукам. Том. 117. стр. 1–11.
- ^ Лакманн, Алек Р.; Эндрюс, Аллен Х.; Батлер, Малкольм Г.; Беляк-Лакманн, Эвелина С.; Кларк, Марк Э. (23 мая 2019 г.). «Большеротый буйвол Ictiobus cyprinellus устанавливает рекорд пресноводной костистости, поскольку улучшенный возрастной анализ показывает столетнее долголетие» . Коммуникационная биология . 2 (1): 197. дои : 10.1038/s42003-019-0452-0 . ISSN 2399-3642 . ПМК 6533251 . ПМИД 31149641 .
- ^ Сигурдардоттир, Андреа Ракель; Сверриссон, Тор; Йонсдоттир, Адальбьёрг; Гудьонсдоттир, Мария; Эльварссон, Бьярки Тор; Эйнарссон, Хафштайн (01 сентября 2023 г.). «Определение возраста отолитов с помощью простого метода обучения за несколько кадров, основанного на компьютерном зрении» . Экологическая информатика . 76 : 102046. doi : 10.1016/j.ecoinf.2023.102046 . ISSN 1574-9541 .
- ^ Арим, Матиас; Ная, Дэниел Э. (2003). «Рацион ластоногих, полученный на основе помета: анализ предубеждений в появлении добычи». Канадский журнал зоологии . 81 (1): 67–73. дои : 10.1139/z02-221 .
- ^ Боуэн, WD (2000). «Реконструкция рациона ластоногих: учет полного переваривания отолитов и клювов головоногих». Канадский журнал рыболовства и водных наук . 57 (5): 898–905. дои : 10.1139/f00-032 .
- ^ Грелье, К.; Хаммонд, PS (2005). «Метод кормления влияет на пищеварение отолитов у содержащихся в неволе серых тюленей: значение для оценки состава рациона». Наука о морских млекопитающих . 21 (2): 296–306. дои : 10.1111/j.1748-7692.2005.tb01229.x .
- ^ Браун, Терпение; Лааке, Джеффри Л.; Делонг, Роберт Л. (2002). «Улучшение анализа рациона ластоногих путем выявления множественных скелетных структур в образцах фекалий» (PDF) . Рыболовный вестник . 100 (3): 423–433.
- ^ Оувеханд, Дж.; Леопольд, МФ; Кампейсен, CJ (2004). «Сравнительное исследование рациона кайр Uria aalge и Razorbills Alca torda , убитых во время инцидента с нефтью Tricolor в юго-восточной части Северного моря в январе 2003 года» (PDF) . Атлантические морские птицы . 6 (3): 147–163. Архивировано из оригинала (PDF) 6 августа 2020 г.
- ^ «ОРНАМЕНТЫ РЫБНЫЕ ОТОЛИТЫ ДЕБЮТ НА РЫНКЕ» . Универсальная группа учреждений . 31 марта 2024 г. Проверено 31 марта 2024 г.
Внешние ссылки
[ редактировать ]| О рыбе |  | ||||
|---|---|---|---|---|---|
| Анатомия и физиология | |||||
| сенсорный системы | |||||
| Воспроизведение | |||||
| Передвижение | |||||
| Другой поведение | |||||
| По среде обитания | |||||
| Другие типы | |||||
| Коммерция |
| ||||
| Основные группы | |||||
| Списки | |||||