Ушастая сова
Эта статья может оказаться слишком длинной для удобного чтения и навигации . ( январь 2022 г. ) |
| Ушастая сова | |
|---|---|

| |
| Ушастая сова в Венгрии. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Птицы |
| Заказ: | Стригиформы |
| Семья: | Стригиды |
| Род: | Добавьте это |
| Разновидность: | А. отус
|
| Биномиальное имя | |
| Дело в том, что это существо | |
| |
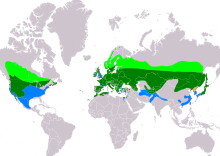
| Ареал A. otus Разведение Резидент Неразмножение Сохранившиеся (сезонность неясна)
| |
| Синонимы | |
| |

Ушастая сова ( Asio otus ), также известная как северная ушастая сова. [ 3 ] или, более неофициально, как малая рогатая сова или кошачья сова , [ 4 ] среднего размера — вид совы с обширным ареалом размножения. Название рода Asio означает в переводе с латыни «рогатая сова», а видовой эпитет otus происходит от греческого и относится к маленькой ушастой сове. [ 5 ] Этот вид гнездится во многих районах Европы и Палеарктики , а также в Северной Америке . Этот вид является частью более крупной группы сов, известной как типичные совы , семейства Strigidae , которое содержит большинство существующих видов сов. [ 6 ] [ 7 ] [ 8 ]
Эта сова проявляет пристрастие к полуоткрытым местам обитания, особенно к опушкам лесов , поскольку они предпочитают ночевать и гнездиться в густых зарослях леса , но предпочитают охотиться на открытой местности . [ 8 ] [ 9 ] Ушастая сова — несколько специализированный хищник, почти полностью сосредотачивающий свой рацион на мелких грызунах , особенно на полевках , которые нередко составляют большую часть их рациона. [ 4 ] [ 8 ] При некоторых обстоятельствах, таких как популяционные циклы их постоянной добычи, засушливые или островные региональные среды обитания или урбанизация , этот вид может довольно хорошо адаптироваться к разнообразию добычи, включая птиц и насекомых . [ 4 ] [ 10 ] [ 11 ] [ 12 ] Не все совы строят свои гнезда. Что касается ушастой совы, она обычно использует гнезда, построенные другими животными, причем во многих регионах предпочтение отдается гнездам, построенным врановыми . [ 13 ] [ 14 ] Успех размножения этого вида во многом коррелирует с популяцией добычи и риском нападения хищников. [ 4 ] [ 13 ] [ 14 ] В отличие от многих сов, ушастые совы не ведут строго территориального или оседлого образа жизни. Они являются частично мигрирующими , и, хотя совы, по-видимому, обычно ежегодно используют одни и те же маршруты миграции и места зимовки, они могут иметь тенденцию появляться настолько беспорядочно, что их иногда называют «кочующими». [ 15 ] Еще одной довольно уникальной характеристикой этого вида является его пристрастие к регулярным насестам, которые часто делят сразу несколько ушастых сов. [ 16 ] [ 17 ] Ушастая сова является одним из наиболее широко распространенных и самых многочисленных видов сов в мире, и из-за очень широкого ареала и численности ее видом, вызывающим наименьшее беспокойство считает МСОП . Тем не менее, в нескольких частях ареала было обнаружено сильное снижение численности этой совы. [ 1 ] [ 18 ]
Таксономия
[ редактировать ]Ушастая сова была официально описана шведским натуралистом Карлом Линнеем в 1758 году в десятом издании его «Системы природы» под биномиальным названием Strix otus . [ 19 ] Эта сова теперь отнесена к роду Asio , который был введен французским зоологом Матюреном Жаком Бриссоном в 1760 году. [ 20 ] [ 21 ]
Совы рода Asio иногда называют ушастыми совами. Несмотря на обширное распространение, считается, что существует только восемь современных видов. Четыре вида встречаются как в Евразии и Африке , так и в Америке , включая ушастую сову и ушастую сову. [ 21 ] [ 22 ] было обнаружено Несмотря на сходство и то, что они относятся к одному и тому же роду, в исследовании с использованием электрофореза , что генетическое расстояние между ушастыми совами и короткоухими совами было необычно большим для видов одного и того же рода. [ 23 ] Несмотря на то, что записи окаменелостей азиатских видов показывают их присутствие в доисторические времена в таких местах, как Канзас и Айдахо ( Asio brevipes ) и Калифорния ( Asio priscus ), точная область эволюционного происхождения ушастой совы неизвестна и вряд ли когда-либо будет известна. [ 4 ] [ 24 ] По крайней мере, три современных вида представляют собой родственные производные, возможно, с ушастыми совами как паравидами или как часть комплекса видов , который потенциально имеет общего базального предка . [ 8 ] [ 22 ] [ 23 ] Во всех трех случаях родственные совы, очевидно, имеют более тропическое распространение и адаптированы к более влажным условиям, имеют более темное оперение и более крупные тела с явно более сильными ногами и более развитыми когтями, возможно, используя относительно незанятую экологическую нишу против конкурирующих сов. [ 8 ] [ 22 ] Одной из этих трех является Стигийская сова , которая является самым темным производным из всех, и в течение некоторого времени было известно, что она отличается от других. [ 4 ] [ 6 ] [ 8 ] Два других, немного более крупных тропических вида, возможно, входящих в видовой комплекс с ушастой совой, одно время считались частью вида ушастой совы. Одна из них — мадагаскарская сова ( Asio madagascariensis ), очевидно, эндемичная для одноименного острова, а другая — абиссинская сова ( Asio abyssinicus ), обитающая в Восточной Африке , особенно в северных регионах, таких как Эфиопия . [ 6 ] [ 21 ] [ 25 ] [ 26 ] [ 27 ] В то время как болотная сова Африки азиатских внешне очень похожа на ушастую сову и, вероятно, тесно связана с ней, полосатая сова ( азиатский моллюск ) представляет собой своего рода особняк среди ныне живущих видов и имеет загадочное происхождение. Несмотря на то, что он генетически связан с другими ныне живущими видами Азии , он не кажется близким родственником. [ 8 ] [ 22 ] [ 23 ] Исследования митохондриального генома показали, что род Asio , а, следовательно, и ушастая сова, совсем недавно отделился среди ныне живущих групп сов от рода Otus или сплюшки , с более отдаленным ответвлением от рода Strix . [ 28 ] Исследование генетической однородности ушастых сов на одном насесте показало, что она несколько выше, чем между разными насестами. Однако эта однородность в целом относительно невелика для общей птицы, сидящей на ночлеге. [ 29 ]
Подвиды
[ редактировать ]Выделено четыре подвида ушастой совы: [ 21 ] [ 30 ] [ 31 ]
- А. о. otus ( Linnaeus , 1758 ) – это номинальный подвид, распространенный по всему ареалу вида в Палеарктике . Его можно встретить на западе, вплоть до Азорских островов , северо-западной Африки , Пиренейского полуострова и Британских островов , а также на востоке, вплоть до Сахалина , Японии и северного Китая . Некоторые популяции этой расы могут зимовать даже на юге, в Египте , Пакистане , северной Индии и южном Китае . Хорда крыла номинального подвида колеблется от 263 до 313 мм (от 10,4 до 12,3 дюйма), а длина хвоста может составлять от 132 до 165 мм (от 5,2 до 6,5 дюйма). Размер, по-видимому, немного увеличивается с запада на восток: совы в Китае примерно на 4% крупнее крылатых, чем совы из Европы. Несмотря на отсутствие известных опубликованных данных о весе восточных/китайских ушастых сов, они, похоже, имеют самые большие размеры среди представителей этого вида. [ 4 ] [ 8 ] У этого подвида лицевой диск имеет тенденцию быть бледно-охристо-коричневым, с черной каймой и относительно короткими бровями с беловатыми отметинами или вообще без отметин. Эректильные пучки ушей хорошо заметны, преимущественно черновато-коричневого цвета с желтовато-коричневыми краями. Верхняя часть тела охристо-желтая, с мелкими темными пятнами и черноватыми полосами на серой «вууле», а макушка от мелких до темных пятен. На затылке и задней части шеи имеются темные стержневые полосы, а внешние опахала перьев лопаток беловатые, образующие ряд через плечо. Первичные первичные волокна в основании однородные охристо-желтые, со светлой и темной дистальной полосой, а второстепенные - охристо-темные с полосой. Хвост обычно рыжевато-коричневый с сероватым оттенком, покрытый 6-8 очень узкими темно-коричневыми полосами. Нижняя часть тела имеет основной цвет охры, передняя часть шеи и верхняя часть груди отмечены черновато-коричневыми полосами; внизу они становятся бледнее и отмечены темными полосами-стержнями и узкими перекладинами. На нижнем крыле имеется характерная полоса и темные отметки в виде запятых на запястье (которые явно отсутствуют на перекрывающихся евразийских короткоухие совы ). Глаза обычно от желтовато-оранжевого до оранжевого цвета, но иногда могут быть хромово-желтыми . Восковица . буровато- мясистая , клюв серый, когти черновато-серые У этого подвида пушистый птенец беловатый с розовой кожей, а мезоптильное оперение от пушистого от сероватого до коричневато-белого с нечеткими полосами и темными маховыми и рулевыми перьями, они похожи на взрослых особей, но с еще не развитыми ушными пучками. [ 4 ] [ 8 ] [ 32 ]
- А. о. canariensis ( Madarász , 1901) — этот подвид является эндемиком Канарских островов . При длине хорды крыла от 257 до 284 мм (от 10,1 до 11,2 дюйма) это, по-видимому, самый маленький подвид ушастой совы. [ 4 ] [ 8 ] Эта раса в среднем темнее, чем большинство ушастых сов номинального подвида, и в целом имеет более тяжелые и резкие темные отметины. Кроме того, ушастые совы Канарских островов, как правило, имеют более яркие красновато-оранжевые глаза. [ 4 ] [ 8 ] [ 33 ]
- А. о. wilsonianus (Lesson, 1830). Этот подвид встречается от юго-центральной и юго-восточной Канады (от Манитобы до Новой Шотландии) до юга США (северная Оклахома и Вирджиния). [ 4 ] [ 8 ] Хорда крыла обычно составляет от 284 до 305 мм (от 11,2 до 12,0 дюймов). В целом американские ушастые совы отмечены более ярко, чем многие евразийские популяции. Лицевой диск ярко-рыжий, с сильным черноватым краем и обширной белой поверхностью вокруг диска. Глаза обычно темно-желтые. Между тем, отметины на нижней стороне обычно довольно черноватые и заметные с отчетливыми поперечными полосами.
- А. о. туфтси Годфри, 1948 г. - Ареал простирается от западной Канады (южный Юкон до юго-западной Манитобы) до юго-центральной части США (западный Техас). Он отличается от А. о. wilsonianus имеет более светлое оперение. Коричневые крапинки на верхней части тела более светлые и ограниченные. [ 34 ] Он слабо дифференцирован и может включать клинальные вариации, обусловленные регионом и средой обитания, а не подвидовыми различиями. [ 8 ] [ 32 ] [ 25 ]
Описание
[ редактировать ]
Этот вид представляет собой довольно стройную и длиннокрылую сову с обычно выступающими эректильными пучками ушей, которые расположены ближе к центру головы, чем у многих других видов сов. Назначение пучков ушей точно не известно, они присутствуют примерно у половины ныне живущих сов. [ 4 ] Вероятно, самая популярная теория среди биологов и орнитологов заключается в том, что пучки ушей могут быть средством внутривидовой коммуникации намерений и настроений. [ 35 ] В общей окраске ушастой совы часто считают оттенок охристо - рыжего с переменно проявляющимся сероватым или коричневатым отливом. На основной цвет обычно накладываются различные черноватые вертикальные полосы (а иногда и пятна), которые обычно более заметны на крыльях и спине. Лопатки , обычно имеют беловатую окраску что обеспечивает дополнительный контраст на фоне основного цвета и черноватых отметин. Темные пятна на запястьях крыльев также могут иметь широкие полосы желтовато-желтого или почти оранжевого цвета на крыльях у основания основных перьев, что представляет собой более ярко выраженную версию рисунка, характерного для других сов, которые, как правило, являются специалистами по охоте на полевок , например, короткоствольных сов. ушастые совы ( Asio flammeus ) и большая серая неясыть ( Strix nebulosa ). Нижняя сторона тела имеет тенденцию быть несколько более бледной охристо-коричневой по сравнению с верхней стороной. Ушастые совы, как правило, имеют темные полосы на верхней части груди, ниже которых они могут быть сильно отмечены рисунком елочки (который создается темными полосами на стержне и перекладинами на этих перьях). [ 3 ] [ 4 ] [ 8 ] [ 9 ] [ 36 ] Существует много индивидуальных и региональных различий в отметках: совы, обитающие в более обширных лесных районах, имеют тенденцию иметь более темный оттенок, часто настолько густо размытые сверху, что кажутся в основном темно-коричневыми на спине, а нижняя сторона в основном покрыта более яркими темно-черными отметинами. . Между тем, в некоторых пустынных регионах оперение может иметь несколько более размытый вид, иногда становясь кремовым или желтоватым , с более редкими и более светлыми темными отметинами в целом. [ 4 ] [ 37 ] Лицевой диск у этого вида хорошо развит и имеет различную окраску (см. подвиды), с темной каймой, часто с белыми полосами, спускающимися по центру через клюв, а иногда белые линии образуют «усы» и/или простираются внутрь. края лицевого диска. Пучки ушей обычно темные спереди и более бледно-желтые сзади. Ушастая сова имеет черноватый цвет клюва, глаза могут варьироваться от желтовато-оранжевого до оранжево-красного, лапки и пальцы ног оперены. [ 4 ] [ 8 ] [ 36 ]
Ушастая сова — это сова среднего размера, общая длина которой составляет от 31 до 40 см (от 12 до 16 дюймов). [ 37 ] [ 38 ] Размах их крыльев относительно велик для их размера: от 86 до 102 см (от 2 футов 10 дюймов до 3 футов 4 дюймов). [ 39 ] [ 40 ] Однако по сравнению с другими широко распространёнными совами, считающимися средними по размеру и с которыми они могут казаться во многом схожими по размеру, такими как сипуха ( Tyto alba ), ушастые совы и неясыти ( Strix aluco ), ушастая сова довольно немного легче и стройнее, взрослый вес составляет около половины веса коричневых сов, что не является редкостью. [ 4 ] [ 41 ] [ 42 ] Как и ожидалось у сов и хищных птиц в целом, ушастые совы демонстрируют обратный половой диморфизм , при котором самки обычно немного крупнее самцов (в отличие от большинства нехищных птиц). Кроме того, самцы могут иметь несколько более бледное оперение, чем самки. [ 9 ] [ 41 ] [ 42 ] В Финляндии одно исследование массы тела взрослых птиц показало, что 22 самца весили в среднем 288 г (10,2 унции), а 20 самок — 327 г (11,5 унции). [ 9 ] [ 37 ] Согласно исследованию, по массе тела европейские ушастые совы противоречат правилу Бергмана (что широко распространенные животные должны быть меньше по размеру ближе к экватору ), поскольку масса тела, по-видимому, увеличивается южнее, будучи самой легкой в Швеции , где в среднем насчитывалось 37 самцов. 197 г (6,9 унции), а 24 самки в среднем весили 225 г (7,9 унции), средний вес в Дании и самый тяжелый в Нидерландах , где средний вес составлял 21 самец. 256 г (9,0 унций), а 24 самки в среднем весили 308 г (10,9 унций). [ 43 ] У перелетных сов, в основном из скандинавских летних угодий в Англии , средний вес 8 самцов составлял 263,6 г (9,30 унции), а средний вес 28 самок - 294,7 г (10,40 унции). [ 44 ] Между тем, в различных исследованиях в Северной Америке 38 мужчин (в Монтане ) имели средний вес 245,3 г (8,65 унции), 55 мужчин (также в Монтане в другом исследовании) - в среднем 261 г (9,2 унции) и 15 мужчин (из Айдахо) ) 232 г (8,2 унции). Согласно тем же исследованиям, соответственно, 28 самок в среднем весили 279,4 г (9,86 унции), 49 самок - в среднем 337 г (11,9 унции) и 19 самок - в среднем 288 г (10,2 унции). [ 41 ] [ 42 ] [ 32 ] [ 45 ] Было обнаружено, что музейные экземпляры в Северной Америке весят в среднем 245 г (8,6 унции) у 38 мужчин и 279 г (9,8 унции) у 28 женщин, а у 520 мигрирующих взрослых в Дулуте , штат Миннесота, средний вес составляет 281,1 г (9,92 унции). [ 32 ] [ 46 ] В целом вес самцов ушастой совы может варьироваться от 160 до 330 г (от 5,6 до 11,6 унций), а самок - от 180 до 435 г (от 6,3 до 15,3 унций). [ 9 ] [ 43 ] [ 32 ]

ушастых сов варьируется При стандартных измерениях длина хорды крыльев от 262 до 315 мм (от 10,3 до 12,4 дюйма) у самцов, при этом 883 в Северной Америке составляют в среднем 285,1 мм (11,22 дюйма) и от 255 до 332 мм (от 10,0 до 13,1 дюйма). ) у женщин, из них 520 американских имеют средний размер 286,2 мм. (11,27 дюйма). Хвост . может иметь длину от 121,5 до 161 мм (от 4,78 до 6,34 дюйма) у самцов и от 143,5 до 165 мм (от 5,65 до 6,50 дюйма) у самок, при этом у 1408 сов средний размер хвоста составляет 146,3 мм (5,76 дюйма) [ 8 ] [ 9 ] [ 32 ] [ 47 ] Менее широко измеряются длина клюва , которая в среднем составляла 15,7 мм (0,62 дюйма) у самцов и 16,2 мм (0,64 дюйма) у самок из Северной Америки, а также длина предплюсны , которая в среднем составляла 38,2 мм (1,50 дюйма) у 20 самцов и 39,9 мм (1,57 дюйма). в) для 16 женщин из Европы с диапазоном у обоих полов от 36,9 до 42,3 мм (от 1,45 до 1,67 дюйма). [ 46 ] [ 48 ] [ 49 ] Что касается строения скелета, у него относительно широкий череп, но относительно маленькие глаза и орбиты, а клюв относительно удлиненный, но слабый. [ 4 ] [ 47 ] Ноги относительно длинные и тонкие, хотя когти острые, как иглы, относительно тонкие, а ступни относительно слабые для совы. Тем не менее, когти по-прежнему способны вытягивать кровь при контакте с кожей человека. [ 4 ] [ 47 ] [ 50 ] [ 51 ]
Идентификация
[ редактировать ]
При хорошем осмотре опытный наблюдатель обычно может отличить ушастую сову по сочетанию ее полевых отметин, размера и окраски. Однако некоторые потенциальные виды сов могут их спутать. Стигийская сова ( Asio stygius ) (которая едва перекрывается, возможно, в северной Мексике) крупнее, с частично голыми пальцами ног и, как правило, темнее, с чернильным оперением и более ярким рисунком, причем часто почти внутренняя лицевая маска выглядит не совсем черной. [ 8 ] [ 52 ] Желто-коричневые совы , которые в Евразии сосуществуют с ушастыми совами, вряд ли ошибутся, учитывая, что они обычно кажутся значительно круглее и громоздче в целом (и действительно немного крупнее и намного тяжелее) и обладают гораздо более широкой и округлой головой. . У коричневой совы нет пучков ушей, глаза черновато-коричневого цвета и сравнительно более короткие крылья. В полете у желто-коричневых сов маховые перья с хорошо развитыми пальцами (с пятью видимыми выемками) в отличие от квадратных крыльев ушастой совы. [ 8 ] [ 37 ] [ 53 ] Евразийский филин ( Bubo bubo ) намного крупнее и массивнее ушастой совы, с заметно более развитыми и мощными на вид ступнями и когтями, а также огромной, квадратной на вид головой с кисточками ушей, расположенными ближе к край. У филина часто более густой рисунок на макушке и спине с тяжелыми черноватыми отметинами, но у него менее четко очерченный (и более мелкий) лицевой диск по сравнению с ушастой совой. [ 8 ] [ 37 ] В некоторых частях зимовочного ареала другие филины едва ли могут соприкасаться с широким ареалом ушастых сов Евразии (и, возможно, северо-западной Африки ), но их обычно можно отличить (так же, как и евразийских видов) по размерным различиям, характеристикам оперения и окраске. , иногда, предпочтения среды обитания. [ 6 ] Азиатские рыбные совы , которые по сути являются подвидом филинов, обычно намного крупнее ушастых сов, с взлохмаченными кисточками на ушах, менее разнообразной окраской и часто оперенными только на части лапок. [ 8 ] В Северной Америке большие рогатые совы ( Bubo Virginianus ), еще один тип филин во всем, кроме названия, имеют квадратную голову и более широко расставленные пучки ушей. Как и другие виды бубонов , большие рогатые совы также заметно крупнее и массивнее любой ушастой совы (несмотря на то, что они меньше евразийского филина). Большие рогатые совы также обычно имеют сильно полосатую нижнюю часть тела, а не полосатую. [ 8 ] [ 9 ] Сплюшки и визгливые совы намного меньше ушастых сов, а также имеют различную окраску (часто с более индивидуально меняющейся общей окраской, например, от серой до коричневой и рыжей ) и обычно имеют довольно короткие пучки ушей. [ 8 ] Болотная сова ( Asio capensis ) (редкое совпадение, возможно, в северном Марокко ) обычно коричневого цвета с совсем другим видом мелких пятен или перемычек внизу, имеет карие глаза и крошечные пучки ушей. [ 8 ]

На большей части своего ареала ушастые совы встречаются с родственной короткоухой совой , причем последний вид в среднем немного крупнее. О различении их в полевых условиях написано много, хотя, если внимательно рассмотреть, сходство между видами не особенно сильное. Кроме того, эти два вида различаются по предпочтениям среды обитания: ушастая сова часто предпочитает (там, где это возможно) любые полностью безлесные, открытые места обитания (включая множество водно-болотных угодий и арктическую тундру , а также степи , прерии и обширные луга ), часто избегая крайние места обитания, излюбленные ушастой совой. [ 4 ] [ 8 ] [ 53 ] Однако при тусклом свете, на расстоянии или в полете замешательство, безусловно, возможно. [ 53 ] [ 54 ] Как у ушастых, так и у короткоухих сов стиль полета имеет характерные, беспорядочные и плавучие взмахи, которые многие орнитологи считают напоминающими мотылька . [ 55 ] В состоянии покоя пучки ушей ушастой совы позволяют легко отличить их друг от друга (хотя ушастые совы иногда могут держать пучки ушей расслабленными). Цвет радужной оболочки различен: желтый у короткоухих и часто оранжевый у длинноухих. Кроме того, черный цвет вокруг глаз вертикальный и легкий у длинноухих и горизонтальный и гораздо более заметный у короткоухих. В целом ушастая сова, как правило, более бледная и песчаная на вид птица, чем длинноухая, без более темных и обширных отметин, как у последней. [ 8 ] [ 53 ] [ 54 ] Есть ряд других различий между этими двумя видами, которые лучше всего заметны, когда они летают. У ушастых сов часто имеется широкая белая полоса по заднему краю крыла, которой нет у ушастых сов. На верхнем крыле основные пятна ушастой совы обычно более бледные и заметные. Полоса на верхней стороне хвоста ушастой совы обычно более жирная, чем у длинноухой. Самые внутренние второстепенные маховые крылья короткоухих часто имеют темные пятна, контрастирующие с остальной частью нижнего крыла. У ушастой совы полосатость проходит по всей нижней части тела, а у короткоухой они заканчиваются на груди. Темные отметины на нижней стороне кончиков самых длинных маховых перьев у короткоухих сов более заметные. У короткоухих сов верхняя часть тела в грубых пятнах, а у длинноухих они более мелкие. Ушастая сова также структурно отличается от длинноухой, имея более длинные и тонкие крылья. Более короткие и широкие крылья в сочетании с более квадратным хвостом у ушастой совы создают пропорции, более напоминающие канюк, чем ушастая сова. [ 8 ] [ 9 ] [ 36 ] [ 53 ] [ 54 ] [ 56 ] [ 57 ] Однако при изучении по остеологическим особенностям различить ушастых и короткоухих сов трудно. [ 58 ]
Вокализации и морфология ушей
[ редактировать ]
У ушастой совы относительно большие ушные прорези, расположенные асимметрично по бокам головы, как и у большинства сов, причем левое ухо выше, а правое ниже, чтобы они могли поглощать звук как сверху, так и снизу. [ 4 ] [ 59 ] Ушная щель занимает почти всю высоту черепа, имеет длину около 38 мм (1,5 дюйма) и покрыта подвижными кожными лоскутами. [ 4 ] [ 60 ] [ 61 ] Правое ухо примерно на 13% больше по сравнению с только что умершими совами. [ 4 ] [ 59 ] [ 62 ] Благодаря своему строению ушей слух ушастой совы примерно в десять раз лучше слуха на высоких и средних тонах , чем у человека. [ 63 ] Сипуха и бореальная сова ( Aegolius funereus ) имеют (в результате конвергентной эволюции ) примерно схожие структуры ушей, при этом относительный размер структуры уха и лицевого диска у сов обычно указывает на уровень важности острого слуха для их жизненного цикла. Совы с относительно меньшими ушными прорезями и более мелкими или рудиментарными лицевыми дисками склонны к более сумеречному или частично дневному поведению, тогда как совы, такие как ушастые совы, более или менее полностью ведут ночной образ жизни. Хорошо известно, что большинство сов могут охотиться в темноте благодаря своему необычайному слуху, который позволяет им определять местонахождение добычи, но они также могут использовать свой слух для отслеживания внутривидовых криков и действий и предотвращения риска нападения хищников. [ 4 ] [ 62 ] [ 63 ]
Вокализации этого вида весьма разнообразны. Среди сов всех возрастов было зарегистрировано, что ушастые совы в Мичигане издавали 23 различных вокала. Карел Воус считал их, вероятно, самыми разнообразными вокалистами из всех видов сов Северного полушария . [ 4 ] [ 64 ] Песня самца ушастой совы представляет собой глубокий крик , который повторяется с интервалом в несколько секунд. Он начинается с нескольких гудков на немного более низком тоне, прежде чем достигнет полной громкости и качества. В спокойные ночи эта песня может распространяться на расстояние от 1 до 2 км (от 0,62 до 1,24 мили) (по крайней мере, для слухового восприятия человека). Песня самца около 400 герц . [ 8 ] [ 9 ] В Северной Америке некоторые наблюдатели считают пение самцов аналогом глубокого воркования полосатых голубей ( Patagioenas fasciata ). [ 4 ] Самки издают более слабую, менее четкую и гораздо более высокую по тону песню гнусавого характера. Почти у всех видов сов самки, несмотря на то, что они обычно более крупные, имеют меньший сиринкс, чем самцы, и поэтому, как правило, имеют менее сильный голос. [ 4 ] [ 8 ] [ 61 ] Крик самки порой сравнивают со слабым оловянным свистком и слышен только на близком расстоянии, будучи примерно на 4-5 полутонов выше пения самца. [ 4 ] [ 8 ] Самки обычно кричат только в дуэте с самцом во время ухаживания, а также при выборе гнезда и в начале инкубации (вероятно, в сочетании с попрошайничеством). Фактически, тщательное исследование показало, что крик самки может происходить каждые 2–8 секунд в ночное время между выбором гнезда и откладкой яиц. [ 8 ] [ 9 ] [ 60 ] [ 64 ] Оба пола издают кошачьи , несколько хриплые ноты джайоу или высокие ноты тявканье-тявканье , причем последние напоминают крик сипух . Когда их потревожат возле гнезда, в котором находятся птенцы, оба родителя могут издать серию жестяных звуков: ватт-ватт-ватт-ватт . [ 4 ] [ 8 ] [ 36 ] В период ухаживания самец летает и машет крылом, издавая хлопающий звук. Во время показательного полета самец может сделать до 20 хлопков в ладоши. [ 8 ] [ 36 ] Как и многие совы, все возрасты могут издавать шипящие звуки и щелкать клювами, когда они чувствуют угрозу, особенно в контексте гнездования. [ 8 ] [ 36 ] [ 65 ] Крик оперившегося детеныша состоит из высоких, протяжных нот, которые по-разному транскрибируются как feek , peeyee и pzeei , и часто сравниваются со звуком ворот , раскачивающихся на ржавых петлях . [ 8 ] [ 36 ]
Распространение и среда обитания
[ редактировать ]

Ушастая сова имеет чрезвычайно большой ареал распространения. В Евразии они распространены с Пиренейского полуострова и Британских островов (включая почти всю Ирландию ), и там и там встречаются несколько пятнисто, но довольно широко, особенно для совы. От западной Франции на восток до остальной Европы они встречаются почти повсюду. Хотя они все еще довольно распространены в этих областях, есть небольшие места, где они обычно не встречаются в Италии , Австрии и Юго-Восточной Европе . [ 1 ] [ 8 ] [ 37 ] В Скандинавии они встречаются только в качестве гнездящегося вида примерно на южных двух третях Норвегии , Швеции и Финляндии , в то время как они обычно сохраняются круглый год в убежищах на южных оконечностях Норвегии и Швеции соответственно, а также во всей Дании . В прибрежной Норвегии они встречаются на северной границе мира в качестве гнездящихся птиц, а ушастые совы гнездятся даже на севере, например, в субарктической зоне Тромса . [ 1 ] [ 8 ] [ 37 ] В широтном диапазоне они встречаются на юге Азорских и Канарских островов , тогда как их ограниченный ареал размножения в Северной Африке простирается от Марокко до Туниса , а также, по-видимому, в самом северном Алжире . [ 1 ] [ 37 ] [ 66 ] [ 67 ] За пределами Европы они встречаются очень редко в качестве селекционеров в Турции , на севере Сирии , Израиле и Ливане . [ 1 ] [ 66 ] [ 67 ] Они довольно широко распространены в России , гнездятся примерно на южных двух третях страны (на север примерно до Чернышевского и Якутска ) и часто встречаются круглый год примерно в южной трети (к северу примерно от городов Пермь , Тюмень и Томск ) и на восток до Сибири , вплоть до Сахалина . Их ареал непрерывен от России до большей части Казахстана , Грузии , Киргизии , примерно половины Узбекистана и редко до северного Афганистана и Туркменистана . [ 1 ] [ 68 ] На востоке они распространены через большую часть Монголии (отсутствуют на юго-западе), а также через западную и восточную части северного Китая , с сезонно неопределенным статусом в Корее . Ушастые совы встречаются на всех островах Японии , но зимуют преимущественно только в точках южнее Осаки . [ 1 ] [ 69 ] Ушастая сова встречается, очевидно, только зимой в небольших местах на юге Франции , на юге Греции , на северо-западе Египта , на севере Ирана , на юге Туркменистана , в основном на большей части территории Афганистана , Пакистана и северной Индии (например , в Катче , Пенджабе , Кашмире ). а также на востоке в Бутане , южном Китае , Тайване и большей части Южной Кореи . [ 1 ] [ 4 ] [ 8 ] [ 67 ] [ 69 ] В результате набегов ушастых сов появились бродячие ушастые совы в различных местах, таких как Фарерские острова , Исландия и Мадейра , а также острова Рюкю на востоке. [ 4 ]
Этот вид широко распространен и в Северной Америке. Их северные пределы достигаются на большей части Британской Колумбии , хотя они в основном отсутствуют в западной и прибрежной части, при этом ареал размножения почти не распространяется на южную часть Северо-Западных территорий . Как и в большинстве стран Евразии, они обычно встречаются на высоте до 50 градусов северной широты . Ушастые совы также гнездятся на большей части территории Альберты , кроме северной Манитобы и южного Онтарио и Квебека, и только до самой южной части Гудзонова залива . Однако, за исключением внутренних районов южной Британской Колумбии, южной Альберты и южных частей Онтарио и Квебека, а также изолированной популяции в Ньюфаундленде , ушастые совы обычно покидают свой канадский ареал зимой. [ 1 ] [ 8 ] [ 9 ] [ 70 ] Ареал вида как размножающегося вида гораздо более обширен на западе, чем на востоке Соединенных Штатов . [ 1 ] [ 9 ] Они размножаются в Вашингтоне , Монтане и Северной Дакоте, в основном непрерывно, на большей части территории Калифорнии , где есть подходящая среда обитания, Аризоне , западном Колорадо и западном Нью-Мексико , а также, в меньшей степени, в Южной Дакоте и Айове . Хотя они отсутствуют на тихоокеанском побережье в Вашингтоне и Орегоне , их можно встретить гнездящимися вдоль Тихого океана в южной Калифорнии и даже в Нижней Калифорнии в Мексике . [ 1 ] [ 9 ] [ 70 ] Первая запись о размножении на материковой части Мексики была зафиксирована в случайно наблюдаемом гнезде, построенном совами в биосферном заповеднике Янош в Чиуауа . [ 71 ] Они также размножаются и встречаются круглый год на большей части территории Миннесоты , Висконсина и Мичигана . Гнездование и/или круглогодичное появление очень редко на востоке США, есть несколько записей о их гнездовании в штатах Мэн , Вирджиния и Западная Вирджиния . [ 1 ] [ 9 ] [ 70 ] [ 72 ] Ушастая сова встречается гораздо шире в Северной Америке в период отсутствия размножения и может быть найдена практически по всему Среднему Западу , Техасу и на юге Мексики, например, в Колиме , Веракрусе и северной Оахаке . Этот вид также встречается в период отсутствия размножения в Луизиане (кроме юго-востока) и на большей части северной Миссисипи , Алабамы , Джорджии и Южной Каролины к северу от Иллинойса , Индианы , Огайо и южной Пенсильвании . [ 1 ] [ 8 ] [ 9 ] [ 70 ] Очень редко эти птицы появлялись во Флориде (во времена исключительных нашествий) и в качестве залета даже на Бермудских островах . [ 4 ] [ 8 ] Они также встречаются зимой и во время миграции на большей части восточного побережья Соединенных Штатов , от Внешних берегов Северной Каролины , широко в восточной Пенсильвании и почти везде в Делавэре или Нью-Джерси , юго-восточном Нью-Йорке (включая Нью-Йорк ). и на север до большей части южной Новой Англии, включая почти весь Коннектикут , Массачусетс и Род-Айленд, а также южный Нью-Гэмпшир . [ 1 ] [ 8 ] [ 9 ] [ 70 ]
среда обитания
[ редактировать ]Оптимальная среда обитания предполагает доступ к открытым пространствам с короткой растительностью и обильной добычей, а также к лесистому покрову для ночевок и гнездования. [ 9 ] [ 73 ] Что касается диапазона отношений, этот вид может обитать на многих высотах без каких-либо сильных высотных предпочтений, хотя они, как правило, отсутствуют выше линии горных деревьев . Было зарегистрировано исключительное гнездование этого вида на высоте 2700 м (8900 футов) над уровнем моря в Кашмире . [ 74 ] Ушастые совы обычно населяют довольно открытые ландшафты с группами деревьев, живыми изгородями или небольшими лесками , а также пастбища с рядами деревьев и кустарников , любой тип леса с вырубками , опушками , полуоткрытым таежным лесом , заболоченными участками. и болота , особенно с ивами , ольхой и тополями , сады со старыми фруктовыми деревьями , парки, кладбища с деревьями и кустами, даже сады и лесные массивы в деревнях , поселках и городах . [ 8 ] Во многих частях мира, включая Китай , Израиль и юго-запад Америки , ушастые совы продемонстрировали способность адаптироваться к пустыням , хотя чаще всего к полупустыням , и могут гнездиться и ночевать в доступных оазисах и приспосабливаться к охоте на добычу. открытая пустынная местность, будь то песчаная или каменистая . [ 75 ] [ 76 ] [ 77 ] [ 78 ] предпочтительная среда обитания в Великобритании Было обнаружено, что чаще всего (среди 200 гнезд) состоит из небольших древесных насаждений , перелесков или разбросанных деревьев на вересковых пустошах , вересковых пустошах или мхах (33%), за которыми следуют участки леса (24,5%), меньшие плантации, защитные полосы или живые изгороди в различных сельскохозяйственных районах (24%), а также кустарники или лесные массивы вблизи побережья и на водно-болотных угодьях (15%). [ 14 ] Все гнезда в Финляндии, участвовавшие в исследовании, находились на расстоянии не более 500 м (1600 футов) от обрабатываемых земель и встречались только на окраинах более крупных лесов или лесов. [ 79 ] Зоны экологической компенсации (т.е. места обитания диких животных на частных сельскохозяйственных угодьях) в Швейцарии действительно обеспечивали среду обитания для ушастых сов, но было обнаружено, что на полевок более активно охотились на скошенных участках земель, а не на участках с более густой растительностью, где полевки были наиболее многочисленны. обильный. Это указывает на то, что среда обитания (в частности, среда обитания в открытом грунте) более важна для хищников, чем плотность добычи, по крайней мере, локально. [ 80 ] В Испании , по сравнению с маленькой совой ( Athene noctua ), ушастые совы чаще встречаются в районах с низким уровнем беспокойства, где лес переходит в плантации, и в районы с относительно низким присутствием человека. [ 81 ] Предпочтение ушастых сов хвойным насаждениям отмечено во многих американских исследованиях. [ 9 ] [ 82 ] В Онтарио этот вид чаще всего гнездится в густых хвойных зарослях и лесовосстановительных рощах, которые часто несколько влажны, а затем реже в смешанных или лиственных районах. [ 83 ] Подобная ассоциация с хвойными деревьями наблюдалась сезонно в Мичигане . [ 82 ] В Северной Дакоте основным местом обитания этих сов были густые заросли небольших деревьев и заросшие кустарником опушки более обширных лесных массивов. [ 84 ] Однако в западных районах, где могут встречаться смешанные леса, лиственные насаждения могут привлечь зимующих сов, если у них есть густые заросли вьющихся виноградных лоз. [ 85 ] В Сьерра-Неваде ушастые совы часто встречаются в прибрежной зоне смешанных лесов вокруг дубов и сосен пондероза ( Pinus ponderosa ). [ 86 ] Анализ, проведенный в Орегоне, показал, что управление лесным хозяйством не оказало заметного влияния на ушастых сов, что указывает на то, что они на самом деле не лесные совы, а вырубка прибрежной растительности, преобразование районов кормления в сельскохозяйственные поля и восстановление лесов в открытых средах обитания действительно сократили местную численность. [ 87 ] Несмотря на то, что ушастая сова способна адаптироваться как к очень холодным районам, включая тайгу и почти субарктику , так и к довольно теплым районам, включая более засушливые и/или засушливые части субтропиков , ушастая сова в основном приурочена к умеренным зонам севера и менее климатична. адаптируется, чем ушастая сова, при этом последний вид адаптируется почти к любому климату и обитает как в Арктике , так и в тропиках, как влажных, так и сухих, при условии, что это открытые места обитания. доступны. [ 4 ] [ 6 ] [ 88 ] [ 89 ]
Поведение
[ редактировать ]
Ушастые совы ведут более или менее строго ночной образ жизни. Обычно активность этого вида начинается в сумерках. [ 8 ] После наступления темноты ушастые совы в Айдахо были наименее активны с 8-10 часов вечера и с 5-6 часов утра, тогда как часы около 22-12 часов и 3-5 часов утра часто были периодами пика активности. [ 90 ] Живя относительно близко к Арктике , ушастые совы могут быть вынуждены добывать корм в светлое время суток, поскольку летом не может быть полной ночи. [ 91 ] Во время дневного полета ушастые совы часто подвергаются нападкам дневных птиц, таких как врановые и другие хищные птицы . [ 8 ] Часто ушастые совы выбрасывают довольно большое количество погадок и бросают их ниже обычных дневных ночевок. В отличие от большинства других сов, этот вид не имеет территориального охотничьего угодья. [ 4 ] [ 36 ] В Швейцарии было обнаружено , что 14 ушастых сов, обследованных с помощью радиотелеметрии, имеют среднюю территорию обитания 980 га (3,8 квадратных миль). В ходе исследования им потребовались поля вдоль границ лесных массивов, избегая полностью безлесных территорий, а не преобладающих в окружающей среде. [ 92 ] В Ческе-Будеевице районе в Чехии было исследовано 9 радиометочных сов. Почти одинаковое количество было обнаружено в пригородных и городских районах, причем городские жители использовали развитые территории для более чем 50% своей ночной деятельности, тогда как пригородные использовали развитые территории для менее половины своей деятельности. Подобные среды обитания предпочитались как городским, так и пригородным совам, но городским совам приходилось расширять ареал, чтобы избежать интенсивной человеческой деятельности и получать доступ к городским паркам, поэтому их средний ареал был больше: 446 га (1,72 квадратных миль) против 56 га (0,22 квадратных миль), в то время как пригородные имели более легкий доступ к лугам и рощам. [ 93 ]
Миграция
[ редактировать ]
Из примерно 19 обычных видов сов в Северной Америке и 13 обычных видов сов в Европе ушастая сова на обоих континентах классифицируется как один из пяти действительно мигрирующих видов , ежегодно перемещающихся по крайней мере в некоторых областях, а в некоторых. цифры от лета к зиме и обратно, вне зависимости от того, буйный ли это год . [ 37 ] [ 70 ] [ 94 ] Северные популяции мигрируют и осенью проявляют сильную тенденцию кочевать на юг. Некоторые обычно молодые птицы из Центральной Европы мигрируют на юго-запад на расстояния более 2000 км (1200 миль). Взрослые особи Центральной Европы менее мигрируют и в лучшем случае просто кочуют зимой. [ 8 ] Ушастые совы, размножающиеся в Скандинавии, обычно мигрируют практически в любую точку Европы, от Великобритании до Юго-Восточной Европы , хотя некоторые из них могут расселиться до Северной Африки или Малой Азии . [ 37 ] [ 95 ] В Европе мужчины и женщины, похоже, немного различаются в миграционном поведении. ушастые совы, зимующие в Дании, Было обнаружено, что сильно предвзято относятся к самкам; также наблюдается предвзятое отношение к самкам в зимних исследованиях в других регионах, таких как южная Швеция . В 10 зимовках Европы самки встречались на 36% чаще, чем самцы. Гипотеза, выдвинутая теми, кто изучал сов в Дании, заключается в том, что самки подвергаются более высокому уровню нападения хищников со стороны более крупных хищных птиц и могут распространяться за пределы Фенноскандии , где плотность этих хищников высока, в районы с низкой плотностью этих хищников. Другая, неисключительная теория заключается в том, что они могут избегать участков с глубоким снегом, который может препятствовать захвату добычи. [ 43 ] Подтверждающие данные о том, что самцы ушастых сов зимуют дальше на север, чем самки, были собраны на юге Норвегии , где при восстановлении мертвых сов (в результате столкновений с автомобилями или линиями электропередачи ) этого вида, встречающегося поздней осенью в течение зимы, самцы были на 45% чаще, чем самки. . [ 96 ] По имеющимся данным, многие из женщин, покидающих Норвегию и даже Фенноскандию, приезжают зимовать в Великобританию (по данным одного исследования, женщины-мигранты здесь встречаются зимой в 3,5 раза чаще, чем мужчины-мигранты). [ 44 ] [ 96 ] В ходе исследований по кольцеванию, зарегистрированных в конце зимы в Германии, было замечено, что две птицы вернулись на лето в центральную Россию (около Ярославля , на расстоянии 2050 км (1270 миль)) и на восток России (около Казани , на расстоянии 2410 км (1500 миль)). ). [ 4 ] птицы, которые размножаются в Центральной Азии, Было зарегистрировано, что зимуют в самых разных местах, включая египетскую долину Нила , Пакистан , северную Индию и южный Китай . [ 8 ] В Северной Америке мигранты, как правило, прибывающие из Канады и Верхнего Среднего Запада, зимуют практически в любой точке оставшихся Соединенных Штатов , однако они будут внезапно мигрировать до Джорджии и некоторых районов Мексики и редко во Флориду . [ 8 ] [ 36 ] Обычно северная граница ареала зимовки приходится на долину Оканаган в Британской Колумбии , южные пределы Верхнего Среднего Запада и центральную часть Новой Англии . [ 4 ] [ 70 ] Весеннюю миграцию отслеживали вместе с северными точильщиками ( Aegolius acadius ), когда они мигрировали через округ Освего, штат Нью-Йорк , через туманные сети , при этом численность ушастых сов значительно превышала численность ушастых сов. Здесь миграция ушастой совы происходила с 21 марта по 14 апреля, и, в отличие от пил, на миграцию ушастой совы, по-видимому, не так сильно влияют погодные условия. [ 97 ] Ушастые совы составляют 19,5% всех сов (или 197 сов), зарегистрированных мигрирующими через мыс Кейп-Мэй осенью (против большинства, 60,6%, являющихся совами-точилками), при этом 26,1% особей видов, пойманных в туманные сети, будучи взрослыми. Более 90% ушастых сов мигрируют в период с середины октября по конец ноября, при этом неполовозрелые особи мигрируют раньше, 52,1% молодых особей прошли в октябре, тогда как только 9,4% взрослых особей мигрировали за месяц. Исследования в Кейп-Мэй также показали, что 58,87% ушастых сов были пойманы в темноте перед рассветом, а не в другое время ночи. [ 98 ] Судя по данным, полученным в Кейп-Мэй, мигрирующие ушастые совы, как правило, летают выше над землей, чем мигрирующие совы-пилы и сипухи , но не так высоко, как ушастая сова, причем последняя сова, часто может избегать туманных сетей по-видимому, . через высоту полета при движении. [ 55 ] Согласно исследованию, проведенному в Айдахо, семь сов с радиометками на самом деле мигрируют как на север, на 75–125 км (47–78 миль) к северу от мест их гнездования, так и на более высокие высоты после сезона размножения (во многих случаях, возможно, для эксплуатации перемещенных грызунов). в недавно зарегистрированных областях). [ 99 ]
Ушастая сова обладает своеобразной способностью увеличивать популяцию, а затем рассеиваться почти в разных направлениях в хорошие годы для увеличения количества добычи. Записи о полосах по Северной Америке показывают крайне нестабильные цифры и перемещения по континенту Северной Америки с непредсказуемыми пиковыми числами мигрантов в совершенно разные годы соответственно для штатов Висконсин , Мичиган , Нью-Йорк и Нью-Джерси . [ 100 ] Поэтому этот вид иногда считают «кочующим», несмотря на то, что многие популяции этого вида являются постоянными ежегодными мигрантами. Аналогичную тенденцию к так называемому «кочевничеству» разделяют и другие широко распространенные хищники, полуспециализированные на охоте на полевок на открытом грунте, например ушастые совы и луни ( Circus cyaneus ). [ 4 ] [ 18 ] [ 100 ] Эти беспорядочные перемещения, пики и отливы северных популяций позволили описать ушастую сову как « назойливую », однако в целом она перемещается совершенно иначе, чем совы, в основном эндемичные для тайги (или тундры ), которые более традиционно агрессивны. в том смысле, что они имеют тенденцию мигрировать незначительно или вообще не мигрировать, когда добыча остается в изобилии в их родном ареале, но затем массово перемещаются на юг, когда популяция добычи сокращается. Ушастые совы, в отличие от этих своенравных северных сов, часто мигрируют из северных районов независимо от условий. Однако, как и северные вспыльчивые совы, ушастые совы, как правило, появляются в беспрецедентных количествах на юге, когда за годом пика добычи следует зима, во время которой популяция добычи сокращается. В Северной Америке миграционные привычки ушастых сов во многом повторяют привычки северных точильщиков . [ 18 ] [ 101 ] [ 102 ] Изучение записей о кольцевании в Саскачеване показывает, что канадские популяции ушастой совы можно считать более агрессивным видом как в качестве гнездящегося, так и в качестве мигрирующего вида, поскольку они появляются в больших количествах только в годы пика полевок, причем большая численность - только в 4 из 44 лет кольцевания. За семь низких лет ушастые совы Саскачевана, казалось, вообще исчезли на большей части территории провинции. Годы пика также часто совпадали с пиками зайца-беляка ( Lepus americanus ), возможно, из-за уменьшения конкуренции (поскольку более крупные совы предпочитают добычу - на местном уровне заяц) и межвидового хищничества со стороны больших рогатых сов . [ 15 ] [ 100 ] Явления перемещений, которые по своей природе кажутся кочующими или предположительно вторженными, могут возникать и в Европе, хотя, учитывая более плотную популяцию этого вида в целом по сравнению с Северной Америкой, это может привести к менее заметным и широким различиям в численности. [ 4 ] [ 37 ] отмечаются годы, когда численность ушастых сов На Британских островах достигает пика, а затем возвращается в Скандинавию, в результате чего на острова попадает гораздо большее количество мигрирующих ушастых сов, чем обычно, а также одновременно большое количество короткоухих сов. ушастые совы и луни. [ 14 ] [ 103 ] На юге Финляндии в год пика добычи, за которым последовал спад добычи, было обнаружено очень большое количество ушастых сов, которые, вероятно, испытывали пищевой стресс, поскольку некоторые из них активно добывали пищу в дневное время, несмотря на продолжительные ночные часы в течение сезона. [ 104 ]
Социальное и ночлежное поведение
[ редактировать ]
В дневное время ушастые совы обычно ночуют в вертикальном положении на ветке, нередко близко к стволу, часто в густой листве. Зимой ушастая сова часто остается рядом с одним и тем же деревом или рощей (например, в парках, больших садах или кладбищах и вокруг них). Обычно при приближении сова замирает, замерев в вертикальном положении, с закрытыми глазами в узкие щели и торчащими пучками ушей. Это называется «высоким-худым положением» и характерно как минимум для пары десятков видов типичных сов. Если подойти близко, совы поочередно откроют и закроют глаза (вероятно, пошевелившись, но пытаясь обмануть потенциальных хищников, заставив их думать, что сова все еще находится в состоянии покоя), наконец, опуская пучки ушей, взбивая оперение тела и перелетая на другой насест. [ 8 ] [ 37 ] В отличие от большинства сов, которые, когда это возможно, демонстрируют тенденцию к территориальному поведению на фиксированном расстоянии круглый год, ушастые совы в период отсутствия размножения могут образовывать скопления сов во время ночлега. Такие группы могут включать от 6 до 50 сов, при этом европейский рекорд составляет около 150 сов на одном насесте. [ 4 ] [ 36 ] [ 37 ] [ 85 ] Даже другие перелетные совы в умеренном поясе не переносят друг друга так близко, как ушастые совы: короткоухие совы, по-видимому, образуют скопления только тогда, когда запасы пищи исключительно высоки, в то время как социальное ночлег у ушастых сов, по-видимому, происходит независимо от этого. местной численности добычи. [ 37 ] [ 105 ] Ушастые совы, как правило, устраиваются на ночлег в глубине «самых темных деревьев», чтобы скрыть свое присутствие, хотя они предпочитают находиться рядом с опушкой леса, чтобы иметь доступ к охоте на более открытой местности. [ 36 ] Исследование, проведенное в районе Медоулендс в Нью-Джерси, показало, что совы, сидящие на ночлеге, очень привязаны к определенным деревьям, особенно к хвойным деревьям, таким как кедры , где основной грузовик скрыт из виду и встречается группа по крайней мере из 2-3 тесно сгруппированных деревьев. Высота насеста в исследовании в Нью-Джерси составляла от 3 до 15 м (от 9,8 до 49,2 футов), а иногда и выше. В Нью-Джерси каждый год предпочтение отдавалось разным местам обитания, и в местной сильно измененной среде совы частично приучаются к деятельности человека. Однако приближение на расстояние ближе 3–4 м (9,8–13,1 футов) обычно приводило к их смыванию. Вылет сов на ночную охоту обычно происходит через 40–49 минут после захода солнца. [ 17 ] Исследования Московского зимовок региона проводились в течение 10 лет, обнаружено 12 общинных и 14 одиночных гнездовий. На коллективных ночлегах за зиму насчитывалось до 16 особей, в среднем 9,9. В целом по Москве в среднем на одно место гнездования приходилось 2,1 совы. Численность полевок в предыдущем году, вероятно, была причиной численных изменений в годах здесь, в то время как избегание ветра и снега было ключевым моментом в характеристиках мест ночевок. [ 106 ] Чрезвычайно большие насесты были нормой в исследовании, проведенном в Ставрополе , Россия , где общая территория насеста могла принимать от 80 до 150 особей каждую зиму в течение 4 лет, при этом 93,7% насестов располагались на хвойных деревьях. [ 107 ] В Милане , Италия , на одном городском месте ночевки наблюдалось от 2 до 76 ушастых сов. Здесь ночные наблюдения показали, что совы по отдельности покидают насест в пик пика в самую темную часть ночи. Большинство миланских сов не летали в сторону городских районов, а летели в пригородные поля и леса, где легче встретить добычу. [ 16 ] Причиной ночлега в скоплениях, по-видимому, является, по крайней мере частично, снижение риска нападения хищников. [ 4 ] [ 17 ]
Диетическая биология
[ редактировать ]
Ушастые совы могут разделить свою охоту на этапы: первый прекращается около полуночи, второй начинается где-то после полуночи и заканчивается за час до восхода солнца. [ 4 ] [ 8 ] Во время охоты они, как правило, сосредотачиваются на опушках лесов, живых изгородях и открытых пространствах с грубыми лугами, а также на молодых деревьях и открытой местности всех видов. Они охотятся в основном на крыле, летая низко и довольно медленно, часто находясь довольно низко от земли, то есть всего около 150 см (59 дюймов). [ 4 ] [ 8 ] [ 36 ] Считается, что ушастые совы чаще всего на слух улавливают шорох намеченной добычи, чем визуально обнаруживая свою добычу. В лабораторных условиях ушастые совы были значительно более умелыми в поиске мышей по звуку в более или менее полной темноте, чем желто-коричневые и полосатые совы ( Strix varia ), а в некоторых американских экспериментах даже превосходили в этом отношении слуховых сипух . [ 108 ] [ 109 ] Как только добыча обнаружена, полет ушастой совы внезапно прекращается, затем они быстро падают с растопыренными когтями, чтобы наброситься на замеченную добычу или особенно на вылезшую на открытое пространство. [ 4 ] [ 8 ] Возможно, среда обитания играет даже более важную роль в выборе охотничьих угодий, чем количество добычи. Часто, когда численность добычи более обильна в лесных массивах рядом с насестом или в богатых болотистых районах поблизости, ушастые совы все равно часто приходят в более бедные добычей районы, такие как старые поля , чтобы они могли обнаружить добычу на открытой местности. [ 80 ] [ 110 ] Размах ног ушастой совы, включая когти, достигает в среднем 11,3 и 12,5 см (4,4 и 4,9 дюйма) у самцов и самок соответственно, что было бы большим для дневной хищной птицы , но довольно маленьким для совы его размер, учитывая физиологические различия в том, как разные хищные птицы склонны убивать свою добычу. В то время как хищники-акциптриды, как правило, убивают, пронзая когтями жизненно важные органы, совы с большей вероятностью задушат свою жертву до смерти, поэтому, как правило, у них пропорционально большие и крепкие ноги. [ 4 ] [ 108 ] [ 111 ] [ 112 ] У других сов среднего размера размах ног у коричневых сов и сипух соответственно составляет в среднем 13,4 и 13,2 см (5,3 и 5,2 дюйма) между полами (в частности, размах ног также является довольно надежным показателем массы тела совы). . [ 4 ] [ 108 ] Поимку добычи ушастыми совами изучали и сравнивали с желто-коричневыми совами в европейском эксперименте. Два вида сов ловили млекопитающих и жуков, пойманных одинаковыми способами, но ушастые совы избегали летающих воробьев и ловили воробьев только тогда, когда они садились на насест (хотя желтовато-коричневые не избегали летающих воробьев, они тоже ловили добычу только тогда, когда сидел). Ушастые совы либо игнорировали, либо убивали, но не ели земноводных , в то время как рыжие совы ловили и поедали земноводных, когда они были доступны. Иногда рыжие совы нападали на рыбу, а одна ушастая сова поймала, но не съела рыбу. Европейское исследование также показало, что несколько сов разных видов ели кусочки растений и, по-видимому, не делали этого из-за недостатка еды или голода. [ 113 ]

В целом, может показаться, что мировая популяция ушастых сов имеет очень разнообразный рацион. В одном исследовании была собрана информация из 312 исследований со всего ареала этого вида. Всего было обнаружено 478 описанных видов добычи, из которых 180 видов млекопитающих , 191 вид птиц , 83 вида беспозвоночных , 15 рептилий , 7 амфибий и, как сообщается, всего пара рыб видов . Всего было проверено около 813 033 объектов добычи. [ 114 ] Однако при ближайшем рассмотрении ушастая сова обычно оказывается чем-то вроде специалиста по питанию. он потребляет в основном, а часто почти полностью, мелких млекопитающих , например грызунов Обычно в качестве пищи почти во всех частях своего ареала . Обычно вырисовывается общая картина: от 80 до 99% рациона составляют млекопитающие, что в среднем составляет 94% по одной оценке для всей Европы. Однако в более теплых , изолированных или более урбанизированных условиях больший процент или, реже, даже большая часть рациона может локально составлять добычу, не являющуюся млекопитающими. [ 4 ] [ 8 ] [ 114 ] [ 115 ] Средние размеры добычи были тщательно изучены и почти всегда находятся в очень узком диапазоне. В Европе средний предполагаемый размер добычи составил 32,2 г (1,14 унции). [ 48 ] [ 115 ] Между тем, в Северной Америке средний размер добычи по двум оценкам варьировался от 30,7 до 37 г (от 1,08 до 1,31 унции). [ 108 ] [ 115 ] В целом, по всему ареалу средний размер добычи обычно составляет от 20 до 50 г (0,71–1,76 унции), обычно значительно меньше 40 г (1,4 унции), и только в тех случаях, когда ушастые совы, возможно, из-за уменьшенного конкуренция, имеет регулярный доступ к добыче весом от 60 до 100 г (от 2,1 до 3,5 унций) или более, средний размер добычи может варьироваться в пределах редко достигают 50–60 г (от 1,8 до 2,1 унции). [ 108 ] [ 114 ] [ 115 ] Случаи исключительно крупной добычи упоминаются ниже.
Млекопитающие
[ редактировать ]
Большую часть пищевой энергии ушастые совы получают от грызунов . Среди этого отряда их обычно относят к одной группе — полевки . История жизни ушастой совы неразрывно связана с полевок. Полёвки, представители семейства Cricetidae и подсемейства Arvicolinae , часто представляют собой многочисленные грызуны малого и среднего размера с относительно короткими хвостами. [ 116 ] [ 117 ] В Европе ушастую сову можно считать специализированным охотником на полевок. Из 86 исследований добычи на континенте примерно в 69% полевки составляли более половины добычи. [ 4 ] [ 114 ] В частности, в регионе Центральной Европы чуть более 82% из 57 500 объектов добычи составляли полевки. [ 4 ] [ 37 ] [ 118 ] Там, где в Европе доступны разнообразные полевки, ушастые совы отдают предпочтение наиболее общительной обыкновенной полевке ( Microtus arvalis ) менее общительной полевке ( Microtus agrestis ). В Центральной Европе 76% рациона составляли только виды обыкновенных полевок. [ 4 ] [ 118 ] [ 119 ] В частности, в крупнейшем известном исследовании, проведенном в Германии , из 45 439 объектов добычи в регионах Берлина и Нордгарца на долю обыкновенной полевки приходилось 72%, а на полевку-полевку и тундровую полевку ( Microtus oeconomus ) вместе взятых - еще 5,5%. [ 120 ] Еще одно большое представительство обыкновенной полевки было в Словакии , где они составляли 84,1% рациона (27 720 из 32 192 объектов добычи). В разные годы в Словакии доля обыкновенных полевок может колебаться от 92,4% до 57,2% в зависимости от численности полевок. [ 121 ] На местах, например, в бывшей Чехословакии и на западе Украины , около 94-95% рациона могут составлять одни только обыкновенные полевки (4153 и 5896 объектов добычи соответственно). [ 120 ] Диетическая связь с обыкновенной полевкой в целом сохраняется в большинстве районов европейской части России , например, в Москве . [ 122 ] [ 123 ] Иногда, как, например, в Молдове , ушастые совы способны уничтожать до 50% популяции обыкновенных полевок, и высказывалось мнение, что полевки легко могут стать опасными для человека, если их не контролировать естественным путем. [ 124 ] Как и многие полевки, обыкновенные полевки подвержены популяционным циклам . В соответствии с циклическим характером популяций полевок локальная численность ушастых сов может резко возрастать и падать. В годы низкого уровня полевок они, как правило, откладывают меньше яиц и выкармливают меньше детенышей и могут вообще не пытаться размножаться. В годы старшей полевки они обычно откладывают и высиживают больше яиц и выращивают больше детенышей. Хотя инкубация начинается с первого яйца, в годы скудного корма можно кормить только самых старших братьев и сестер. Обычно в бедные годы для обыкновенных полевок едят различные виды мышей , но они кажутся неэффективной заменой (по крайней мере, в более северных странах), поскольку у сов более низкие темпы размножения. [ 4 ] [ 36 ] [ 37 ] [ 118 ] По-видимому, обыкновенные полевки время от времени могут коллективно переходить от временной активности к более дневной , что может быть попыткой смягчить сильное хищничество сов (особенно потому, что практически все европейские совы склонны охотиться на них), хотя некоторые дневные хищники являются почти такими же специализированными хищниками, как и обычные полевки. их тоже. [ 125 ] Имеющиеся данные указывают на то, что обыкновенные полевки меняют свои жизненные циклы с неизвестными долгосрочными последствиями, вероятно, из-за глобального потепления . Вполне вероятно, что это повлияет на ушастых сов региона, но неясно, каким именно будет результат. [ 126 ]
В некоторых частях Европы обыкновенные полевки иногда не встречаются или локально встречаются нечасто или редко, особенно на крупных островах, в Скандинавии и в некоторых частях южных областей, таких как Пиренейский полуостров , Италия и Греция . Поэтому ушастые совы питаются в основном разными видами добычи. Дополнительной или, иногда, основной добычей, когда полевки менее распространены, являются грызуны-мюриды , особенно представители более обычных родов, такие как Apodemus или полевые мыши, Mus или домашние мыши и, иногда, Rattus или типичные крысы. [ 4 ] [ 37 ] В скандинавских исследованиях обыкновенные полевки по-прежнему были основной добычей в Финляндии , где они были распространены (и превосходили полевок в рационе совы в соотношении четыре к одному) и частично в Дании (где пища была разделена между двумя видами обычных полевок и полевыми мышами), но находятся не в Швеции и Норвегии . [ 37 ] [ 120 ] [ 127 ] В Швеции, где обыкновенные полевки не встречаются, основной пищей были полевки, составляя 65,2% из 13 917 объектов добычи, за которыми следовали виды полевых мышей Apodemus , которые составляли еще 25,3%. [ 128 ] В Норвегии 3431 объектом добычи были в основном полевки (42,75%), Apodemus виды (12,64%), тундровые полевки (12,35%) и рыжие полевки ( Myodes glareolus ) (12,06%). [ 120 ] На Британских островах основная добыча переключалась между полевками (46,6% из 1228 объектов добычи в Пик-Дистрикте и 79% в южной Шотландии ) и мышами Apodemus , особенно лесной мышью ( Apodemus sylvaticus ) (75,6% из 1772 объектов добычи). собраны из разных частей Англии, и 69,5% из 1373 объектов добычи находятся в Ирландии , где родной полевки полностью отсутствуют). [ 12 ] [ 120 ] [ 129 ] [ 130 ] В более теплых регионах рацион ушастых сов может различаться в зависимости от местного состава добычи. В Испании полевки, в том числе обыкновенные полевки, средиземноморские сосновые полевки ( Microtus duodecimcostatus ) и лузитанские сосновая полевка ( Microtus lusitanicus ), в совокупности составляли 76,4% из 6945 объектов добычи в центральной части страны, а на реке Эбро на севере Испании Алжирская мышь ( Mus spretus ) доминировала, составив 69,5% из 846 жертв. предметы. [ 131 ] [ 132 ] В целом, в 7 исследованиях, проведенных в различных местах Испании, лесные мыши и виды Mus были наиболее регулярной добычей (в совокупности они составляли чуть менее 60% от общего числа объектов добычи). [ 132 ] Многие исследования, проведенные в Италии, показывают, что этот вид не является специализированным кормовым объектом в стране со значительными региональными различиями в предпочтениях добычи. [ 133 ] В частности, ненастная погода , включая любые осадки или сильный ветер , по-видимому, заставляет итальянских ушастых сов увеличивать разнообразие добычи, которую обычно ловят. [ 134 ] В некоторых итальянских исследованиях сосновая полевка Сави ( Microtus savii ) была основным продуктом питания в Приньяно-Чиленто , где они составляли 60,4% по численности и 61,6% по биомассе . В других древесная мышь была основной пищей, например, у Кремоны , составлявшей 59,1% из 1482 объектов добычи. [ 135 ] [ 136 ] Необычная ассоциация с близкой добычей была отмечена в северной Италии , где доступ к свалкам позволил им получить доступ к исключительно крупной добыче, бурой крысе ( Rattus norvegicus ), с пойманными молодыми крысами, которые весили в среднем 140 г (4,9 унции), а иногда и до 243 г. г (8,6 унции), что составляет 20,5% по численности и 65,1% по биомассе, хотя самой многочисленной найденной добычей были лесные мыши. в гранулах. Из-за доступа к крысам средний размер добычи в североитальянском исследовании был исключительно высоким — 58 г (2,0 унции). [ 137 ] В целом, мыши Mus кажутся основным кормом для ушастых сов в Греции, особенно македонская мышь ( Mus macedonicus ), но нередко и южная полевка ( Microtus levis ) также играет важную роль в рационе там. [ 138 ] [ 139 ] На Канарских островах завезенная домовая мышь ( Mus musculus ) сегодня считалась главным кормом для сов, составляя 69,5% из 3628 объектов добычи, согласно крупнейшему известному исследованию. [ 11 ]
За пределами Европы, в пределах Евразии, пищевая ассоциация ушастых сов с полевками несколько ослабевает, но продолжается в прилегающей Турции , особенно южная полевка и полевки Гюнтера ( Microtus guentheri ) (последние, например, составляют до 78,7% от 5324 предметы добычи в Карапынаре ). [ 140 ] [ 141 ] [ 142 ] На Ближнем Востоке предпочтения добычи варьировались в зависимости от состава почвы на окраинах пустыни : израильские исследования показали, что первичный переход от Gerbillus видов песчанок Meriones к песчанкам и полевкам Гюнтера аналогичен результатам, полученным у зимующих сов в Иране. [ 77 ] [ 143 ] [ 144 ] относительно крупная добыча, индийские песчанки ( Tatera indica ) и короткохвостые крысы-бандикуты ( Nesokia indica Сообщалось, что у зимующих ушастых сов в Иране ), весит в среднем 163 г (5,7 унции) и 155 г (5,5 унции). соответственно, составляли значительную часть добычи (72,9% биомассы) и добывались почти в четных количествах с меньшими Виды гербилл . [ 144 ] Гораздо дальше на восток, в Китае и Монголии , ушастые совы часто питаются различными видами грызунов, особенно мелкими хомяками , которые также являются крицетидами, но не арвиколинами, такими как китайская полосатая ( Cricetulus barabensis ) и карликовые роборовские ( Phodopus roborovski ). с более крупными джирдами, такими как полуденные джирды ( Meriones meridianus ), а также с домашние мыши . [ 75 ] [ 145 ] [ 146 ] [ 147 ] В более северо-восточных регионах полевки по-прежнему имеют важное значение. В Западной Сибири тундровые полевки, узкоголовые полевки ( Microtus gregalis ), евроазиатская промысловая мышь ( Micromys minutus ) и степной лемминг ( Lagurus lagurus ). основной добычей грызунов были [ 148 ] В Японии рацион питания сильно смещен в сторону грызунов, таких как японская травяная полевка ( Microtus montebelli ) (84,2% корма в Ниигате на Хонсю ), серая красная полевка ( Myodes rufocanus ) (87,2% на Хоккайдо ) или домовая мышь (77,7%). в префектуре Эхимэ , Сикоку ). [ 149 ] [ 150 ] [ 151 ]
В Северной Америке ушастые совы также в основном полагаются на мелких грызунов в своем рационе, но их рацион несколько более разнообразен в зависимости от семейства грызунов и менее полностью зависит от полевок, чем у их евразийских собратьев. [ 115 ] В целом, чем севернее они обитают в Северной Америке, тем более ограниченной и основанной на полевках диета этого вида. Хотя диеты в Канаде детально не изучены, [ 152 ] в северных штатах Массачусетс , Мичиган , Миннесота , Огайо , Нью-Йорк , Висконсин и северный Орегон полевки легко становились основной добычей ушастых сов. [ 64 ] [ 110 ] [ 153 ] [ 154 ] [ 155 ] [ 156 ] [ 157 ] [ 158 ] В частности, луговая полевка ( Microtus pennsylvanicus ), как правило, является основным продуктом питания, как, например, в двух крупных американских исследованиях: в Мичигане, где они составляли 70,6% из 3269 объектов добычи, и в Висконсине, где они составляли 83,4% из 3273 объектов добычи. предметы. [ 64 ] [ 159 ] Еще одна полевка, регулярно включаемая в рацион питания в Америке, — это степная полевка ( Microtus ochrogaster ), но, судя по всему, мало других полевок, за исключением штата Орегон, где на местном уровне лидировали серохвостая полевка ( Microtus canicaudus ) и полевка Таунсенда ( Microtus Townsendii ). продукты. [ 156 ] [ 158 ] [ 160 ] В Северной Америке , не относящиеся к арвиколину, крицетидные грызуны такие как род Peromyscus , или оленьи мыши, и более мелкие Reithrodontomys , или мыши-промысловики, заполняют нишу мелких диких мышей и могут быть неотразимы для охоты на ушастых сов. [ 115 ] Мыши Peromyscus были основным кормом для этого вида в Колорадо , Иллинойсе и большинстве исследований в Айове (составляя до 59,1% местного рациона). [ 108 ] [ 161 ] [ 162 ] [ 163 ]
Во многих районах, особенно в засушливых районах, надсемейство Geomyoidea вытесняет крицетидных грызунов в качестве основного продукта питания, а именно карманных мышей , кенгуровых крыс и иногда карманных сусликов и прыгающих мышей . В частности, это справедливо на юго-западе Америки где в Аризоне , карманные мыши Perognathus составляли 61,3% рациона, в Нью-Мексико , где пары видов карманных мышей и крыс-кенгуру составляли 51,8% и 20,5% пищи соответственно. и в южной Калифорнии , где Perognathus составлял 51% пищевых продуктов, а кенгуровые крысы Dipodomys составляли еще один процент пищевых продуктов. 37,8% продуктов. [ 78 ] [ 164 ] [ 165 ] В пустыне Сонора в Мексике почти вся известная добыча была геомиоидами, в частности крыса-кенгуру Мерриама ( Dipodomys merriami ), которая сама по себе составляла 74,7% пищи. [ 166 ] В хорошо изученной популяции региона Снейк-Ривер на юго-востоке Айдахо , а также округа Оуихи в юго-западной части штата геомиоидные грызуны обычно являются наиболее заметной добычей, особенно карманная мышь Большого Бассейна ( Perognathus parvus ) и кенгуровая крыса Орда ( Perognathus parvus ). Dipodomys ordii ) (часто в больших количествах добавляют мышей Peromyscus ). Из-за относительно большого размера крыс-кенгуру в Айдахо средний размер добычи может достигать как минимум 41 г (1,4 унции). [ 167 ] [ 168 ] [ 169 ] [ 170 ] В некоторых частях Северной Америки более богатая биомасса вероятна, когда первичную позицию занимает более крупная добыча, например хлопковые крысы . Хлопковые крысы были основной добычей в биосферном заповеднике Янос в Мексике (43,2% по численности, 69,1% по биомассе) и в Техасе , в последнем немного превосходя по численности (36%) гораздо меньших по размеру промысловых мышей (23%). [ 171 ] [ 172 ] Поскольку средний размер тела пойманных хлопковых крыс ( Sigmodon hispidus ), по сообщениям, составляет около 100 г (3,5 унции), они, вероятно, представляют собой очень продуктивную добычу для ушастых сов. [ 172 ] [ 173 ] Аналогичным образом, исключительная крупная добыча была добыта на северо-востоке Орегона , где были пойманы как молодые особи весом около 30 г (1,1 унции), так и взрослые особи, весом около 90 г (3,2 унции), северных карманных сусликов ( Thomomys talpoides ), которые составляли 55,7% по численности. и 74,4% по биомассе рациона ушастой совы. В зависимости от обстоятельств средний размер северных карманных сусликов, пойманных в разных районах, может варьироваться от 41 до 100 г (от 1,4 до 3,5 унций) или выше, но ушастые совы обычно вылавливают молодь за пределами исследования в Орегоне (с весом на нижнем пределе). такого массового масштаба). [ 90 ] [ 169 ] [ 174 ] [ 175 ]
Добыча из млекопитающих, не являющихся грызунами, редко имеет большое значение для ушастых сов, хотя на местном уровне они могут добывать некоторое количество других видов млекопитающих. Несмотря на заявления о том, что ушастые совы «избегают» землероек как добычи, вероятно, правильнее сказать, что они не ищут их почти так часто, как более социально настроенных и/или густонаселенных грызунов. Некоторые другие совы могут считаться обычными и обычными хищниками землероек, например, часто сипухи . В Европе общая картина выбора добычи показывает, что около 2% рациона ушастых сов составляют землероек. [ 4 ] [ 48 ] [ 115 ] На местном уровне относительно высокая численность землероек была зарегистрирована в Финляндии , где 10,7% из 3759 объектов добычи составляли обыкновенные землеройки ( Sorex araneus ), в южной Шотландии , где обыкновенная бурозубка составляла 17,3% из 514 объектов добычи, на северо-востоке Греции , где меньшие белозубая бурозубка ( Crocidura suaveolens ) составляла 19,3% из 311 объекты добычи и в Западной Сибири , где виды Sorex составляли 17,3% от 335 объектов добычи. [ 12 ] [ 127 ] [ 138 ] [ 148 ] Точно так же в Северной Америке землеройки редко являются чем-то большим, чем второстепенная добыча. [ 115 ] Исключительно высокая численность землероек отмечена в Огайо , где относительно крупная северная короткохвостая бурозубка ( Blarina brevicauda ) и относительно мелкая североамериканская землеройка ( Cryptotis parva ) составляли 10,46% и 10,22% рациона соответственно, а в Теннесси , где 12,85% рациона меньше всего составляли землероек. [ 156 ] [ 173 ] Самое маленькое млекопитающее на земле (и, следовательно, самая маленькая зарегистрированная добыча среди млекопитающих или позвоночных), этрусская землеройка весом 1,8 г (0,063 унции) ( Suncus etruscus ), редко становится объектом охоты ушастых сов. [ 176 ] Летучие мыши — еще один дополнительный тип добычи ушастых сов. Одно компиляционное исследование, основанное на 12 исследовательских участках в Средиземноморском регионе (в Испании , Италии , Греции , Словении , Румынии и Швейцарии ), показало, что до 2% останков добычи составляли летучие мыши. Размер летучих мышей, пойманных в Средиземноморском регионе, варьировался от усатой летучей мыши ( Myotis mystacinus ), вес которой оценивается всего в 4 г (0,14 унции), до европейской летучей мыши со свободным хвостом ( Tadarida teniotis ), вес которой, по оценкам, достигает 54 г. (1,9 унции). [ 177 ] Исключительно тесные хищнические отношения были отмечены между летучими мышами, особенно японскими домашними летучими мышами ( Pipistrellus abramus ), и ушастыми совами в районе Пекина в Китае , где летучие мыши составляли 28,6% из 3561 объекта добычи в целом и 56,6% рациона на местном уровне. в городских, а не пригородных поселениях. [ 178 ] Известно, что на других млекопитающих, не входящих в вышеупомянутые группы (например, крицетидных, мюридных и геомиоидных) грызунов, ведется охота, но они, как правило, составляют незначительную часть рациона ушастой совы, включая ежей , кротов , кроликов , зайцев и ласк, а также, в редких случаях, добыты грызуны, такие как сони , летяги и белки (в том числе бурундуки ). [ 4 ] [ 114 ] [ 115 ] [ 121 ] [ 131 ] [ 147 ] Хотя зайцеобразные, такие как зайцы и кролики, являются очень редкой добычей ушастых сов, иногда они могут поймать одну из них. По оценкам, такая добыча весила 271 г (9,6 унции) (для кроликов ) и 471 г (1,038 фунта) (для кроликов ) в Айдахо, что означает, что, скорее всего, будут пойманы очень маленькие молодые кролики и кролики. [ 167 ] [ 169 ] [ 170 ] [ 174 ] Однако иногда ушастым совам очень редко удается поймать исключительно крупных зайцеобразных. Добыча рекордного размера среди млекопитающих, пойманная ушастой совой, была зарегистрирована в случае нападения хищника на предположительно молодого чернохвостого кролика ( Lepus Californicus ), который весил примерно 800 г (1,8 фунта). [ 115 ] Два случая поедания падали были добычей зарегистрированы в Италии , первый известный случай этого для этого вида, где ушастые совы поедали части взрослого хохлатого дикобраза ( Hystrix cristata ) и взрослой европейской куницы ( Martes martes ), которые являются ушастые совы вряд ли могли убить. [ 179 ]
Птицы
[ редактировать ]
Ушастые совы вообще нечастые хищники птиц. Исследования пищевых продуктов в Евразии показывают, что это оппортунистический и случайный хищник птиц, в то время как в Северной Америке они, похоже, обычно не ловят большое количество птиц в каком-либо районе. Зимой иногда эти совы могут питаться в основном мелкими птицами, собравшимися в общих ночлегах, часто вблизи деревень или городов. В частности, снежный покров зимой может повлиять на то, что местные ушастые совы перейдут от млекопитающих к птицам. [ 180 ] Как и сипухи , ушастые совы, как известно, носятся вокруг насестов в кустах, пытаясь потревожить спящих птиц, что может спровоцировать добычу вылететь из своего убежища только для того, чтобы ее поймали. [ 4 ] [ 181 ] Ушастые совы, а также мигрирующие короткоухие совы , были замечены в Испании, охотясь на ночных воробьинообразных , которых привлекали искусственные источники света. [ 182 ] Убивая птиц, ушастые совы, скорее всего, будут клевать заднюю часть тела и голову и обезглавливать своих жертв, что приводит к большему повреждению скелета, чем это типично для других сов, и потенциально затрудняет идентификацию добычи. [ 183 ] Особенно часто ушастые совы в городских районах и/или на окраинах засушливых ареалов ловят домашних воробьев ( Passer Domesticus ), а иногда и евразийского древесного воробья ( Passer montanus ). Удивительно высокий баланс добычи для зимующих ушастых сов в пустынных районах приходится на птиц. Так было в Алжире , где 37,5% рациона и 40% биомассы составляли птицы, а наиболее идентифицируемым общим видом добычи были виды Passer с 20,7% по численности и 17% биомассы. [ 184 ] У зимующих сов в городе Иерусалиме 90,7% рациона (150 объектов добычи) составляли мелкие птицы во главе с домашним воробьем (22%) и черной шапкой ( Sylvia atricapilla ) (16,7%). [ 10 ] Дальнейшее изучение рациона ушастой совы в израильской пустыне Негев показало, что 28,3% из 3062 объектов добычи составляли птицы. [ 185 ] В Египте 24,6% объектов добычи составляли птицы, в том числе домашний воробей (15,4%) и европейский щегол ( Carduelis carduelis ) (2,4%). [ 186 ] В Европе птицы редко портят большую часть продуктов питания, но, тем не менее, в некоторых регионах их добывают в больших количествах. По оценкам, в Центральной Европе птицы составляют в среднем 8% рациона (из 52 видов). В годы пика численности полевок птицы могли составлять менее 2% пищи, в то время как в годы спада полевок птицы составляли до 33%. [ 118 ] Аналогичным образом, в Софийском парке ( Болгария ) количество птиц, составляющее в среднем 9,6% по численности и 7,9% по биомассе, варьировалось в зависимости от численности полевок, но было зарегистрировано большое количество - 44 вида птиц-жертв. [ 187 ] В Сонийском лесу , Бельгия , 38,3% из 355 объектов добычи составляли птицы, преимущественно различных видов воробьиных . [ 188 ] Исследование, проведенное в Баден-Вюртемберге , Германия, показало, что птицы подвергают риску 14,75% из 12 890 объектов добычи, что является относительно высоким балансом, причем наиболее распространенными идентифицированными видами птиц являются европейский щегол. [ 189 ] В целом британские исследования показали, что в выборке из 7161 объекта добычи 1161 были птицами (14,95%), и эти птицы присутствовали в 90% исследованных погадок. Из них 46,9% — воробей домашний, 7,5% — скворец обыкновенный ( Sturnus vulgaris ), 4,65% — дрозд обыкновенный ( Turdus merula ), 3,35% — зеленушка европейская ( Chloris chromis ), 2,92% — певчий дрозд ( Turdus philomelos ), 2,49 % составили евразийский жаворонок ( Alauda arvensis ) и 2,23% составляли коноплянка обыкновенная ( Linaria cannabina ). [ 190 ] В более локальном районе Пик-Дистрикт Англии птицы составляли 23% добычи по количеству и 31,3% по биомассе. Среди исследованных здесь птиц большинство идентифицированных составляли луговые коньки ( Anthus pratensis ), за которыми следовал твит ( Linaria flavirostris ), а идентифицированные птицы варьировались по размеру от вероятной лесной славки ( Phylloscopus sibilatrix ) (средний вес взрослой особи 9,2 г (0,32 унции) )) взрослой чибиси ( Vanellus vanellus ) (средняя масса взрослой особи 219 г (7,7 унция)). При этом 80% по численности и 11 из 25 видов хищных птиц были характерны для открытого местообитания. [ 42 ] [ 129 ]
Многие итальянские исследования показывают относительно большое количество охотящихся птиц, главным образом в периоды, когда они не размножаются. Одно из самых маленьких и крупных представлений находилось в Приньяно-Чиленто , где 13,85% составляли в основном неопознанные птицы. [ 135 ] В региональном парке Десима-Малафеде птицы составляли 31,1% рациона, 13,7% из которых составляли неопознанные воробьиные, в то время как европейские щеглы и европейская зеленушка составляли еще 7,1% и 4,6% объектов добычи. [ 191 ] В районе Венеции из 642 исследованных объектов добычи и общей массы добычи 15 038 г (33,153 фунта) птицы составляли 38,47% по численности и 41% биомассы. [ 192 ] На зимних ночлегах Империи 63,43% из 1020 объектов добычи составляли птицы и 36,57% — млекопитающие. Основной идентифицированной добычей была черная шапка (51,6%), тогда как зяблик ( Fringilla coelebs ) был второстепенным среди птиц (6,73%). [ 193 ] В румынских исследованиях птичья добыча также имела относительно важное значение. В Агигее 32,71% продуктов питания составляли птицы, при этом виды Carduelis в совокупности составляли 6,04%, а ласточки были второстепенными, например, ласточка обыкновенная ( Delichon urbicum ) (2,52%) и деревенская ласточка ( Hirundo Rustica ) (2,44%). [ 194 ] Птицы были основным кормом для зимующих ушастых сов в дельте Дуная в Румынии : птицы составляли 59,5% общей добычи по численности и 51,6% по биомассе из 948 объектов добычи против 40,7% по численности и 48,4% по биомассе млекопитающих. Здесь в основном были добыты многочисленные воробьинообразные, семейство вьюрковых (18,6%), семейство воробьиных (15,7%) и семейство синиц причем наиболее распространенными кормовыми семействами среди птиц были (12,7%). Средний размер добычи птицы составил 22,2 г (0,78 унции), тогда как средний размер добычи млекопитающих составил 24 г (0,85 унции). На долю домовых воробьев приходилось 14,3% биомассы, а на евразийских черных дроздов - 12,3% биомассы Дуная. [ 195 ] В то время как млекопитающие обычно доминируют в рационе ушастых сов в Испании, в заповеднике Альбуфера страны птицы составляли 53,5% по численности и 48,6% биомассы 864 объектов добычи. Всего отмечено 34 вида птиц, во главе с пеночкой обыкновенной ( Phylloscopus colybita ) (12,5% по численности, 4,8% по биомассе), домовым воробьем (8,2% по численности, 12,2% по биомассе), деревенской ласточкой (6,4% по биомассе). численности, 7% по биомассе) и ласточка ( Riparia riparia ) (3,2% по численности, 2,7% по биомассе). [ 196 ] Основным зарегистрированным отдельным видом добычи в Пекине был евразийский древесный воробей, составлявший 38% рациона, но другая птичья добыча здесь была незначительной. [ 178 ]
Хотя большинство воробьинообразных, на которых нападают ушастые совы, находятся в пределах типичных размеров добычи для этого вида, во многих сообщениях отмечается, что во время преследования птицы можно напасть на необычайно крупную добычу. птичья добыча весом около 100 г (3,5 унции) или несколько выше, например, более крупные обыкновенные дрозды или сойки Нередко добывается . Однако необычно крупная птичья добыча для ушастых сов в Европе включала в себя несколько видов как минимум в два раза более массивных, в том числе имаго следующих видов: чибисы , вяхирь обыкновенный ( Columbus palumbus ), со средней массой взрослой особи 490 г ( 1,08 фунта), камышницы обыкновенные ( Gallinula chromopus ), средний вес взрослой особи 343 г (12,1 унции), красноногие куропатки ( Alectoris rufa ), со средним весом взрослой особи 528 г (1,164 фунта), галки западные ( Corvus monedula ), со средней массой взрослой особи 246 г (8,7 унции) и евразийские сороки ( Pica pica ), со средней массой взрослой особи 230 г (8,1 унции). Большую часть самой крупной идентифицируемой и доказуемой добычи ушастых сов в Европе в целом оказались птицы. [ 4 ] [ 42 ] [ 114 ] [ 118 ] [ 190 ] [ 195 ] [ 197 ] Несмотря на относительно нехватку птичьей добычи в рационе Северной Америки, там также были зарегистрированы необычно крупные птичьи жертвы. В число таких жертв входили взрослые особи северо-западной вороны ( Corvus caurinus ), средний вес которых составлял 392 г (13,8 унции), два крупных взрослых тетерева ( Bonasa umbellus ), вес которых, по оценкам, составлял чуть более 600 г (1,3 фунта), и даже, очевидно, по крайней мере один раз в год. взрослый острохвостый тетерев ( Tympanuchus phasianellus ), средний вес которого относительно огромный — 885 г. (1,951 фунта) или в три раза тяжелее средней ушастой совы. [ 64 ] [ 198 ] [ 199 ] [ 200 ]
Другая добыча
[ редактировать ]За исключением млекопитающих, которые портят большую часть продуктов питания, и птиц, которые ставят под угрозу второстепенную, но локально важную часть продуктов питания, ушастые совы редко где-либо поедают другие виды добычи. Нечасто это рептилии , такие как несколько видов змей и ящериц , и еще меньше земноводных, таких как лягушки и жабы . Обычно эта добыча появляется более чем поодиночке, насколько известно, в слегка засушливых и теплых частях ареала вида, в основном на Канарских островах и иногда на юго-западе Америки . [ 13 ] [ 115 ] [ 165 ] [ 201 ] [ 202 ] Рыба почти никогда не встречается в рационе, всего в Европе зарегистрировано два вида кормовых рыб: карп и еще неопознанный карп. [ 203 ] Несмотря на довольно высокое разнообразие насекомых (и низкое разнообразие других беспозвоночных , таких как паукообразные ), собранных в целом, особенно в разных частях Евразии , они редко вносят значительный вклад в рацион ушастой совы. Обычно в Европе, если в гранулах обнаруживаются какие-либо насекомые, их доля в численности добычи обычно составляет менее 2%. [ 4 ] [ 118 ] [ 204 ] [ 205 ] Аналогичное присутствие, но небольшое количество насекомых было отмечено и в Израиле. [ 77 ] В некоторых исследованиях отмечен исключительно большой вклад насекомых, чаще всего различных видов или родов жуков , в число жертв, например, они составляют 6,6% рациона в региональном парке Децима-Малафеде в Италии, 17,5% в северо-восточной Греции и 13,3%. % в западной Испании . [ 139 ] [ 206 ] Рекордным вкладом насекомых для Европы, несомненно, было исследование в центральной Польше , где было обнаружено, что одиночный жук , обыкновенный майский жук ( Melolontha melolontha ), составляет 25% объектов добычи. [ 207 ] Согласно нескольким исследованиям, проведенным в Алжире , насекомые являются там обычной дополнительной пищей, составляя около 17,3% объектов добычи. [ 184 ] [ 208 ] На острове Тенерифе на Канарских островах исключительные 33% пищи составляли насекомые, причем как кустарниковые, так и полевые сверчки составляли по 14,8% каждый, хотя в более широких исследованиях на Канарских островах значимость насекомых снижается до 10,4%. . [ 11 ] [ 209 ] Максимальный известный вклад насекомых в рацион Северной Америки составлял всего лишь 4,3% пищи для совместного ночлега ушастых сов на юго-востоке Айдахо . [ 90 ]
Межвидовые хищнические отношения
[ редактировать ]
Ушастая сова встречается во многих конкурентных средах умеренной зоны вместе с другими хищными птицами . Особенно вероятно встретить большое разнообразие сов как с точки зрения общего ночного образа жизни , так и общего предпочтения в отношении добычи грызунов, которому отдают предпочтение около 75% сов, обитающих в Северной Америке, и около 85% сов, обитающих в Европе, а иногда и в Европе. его предпочитают почти все совы на обоих континентах. [ 4 ] [ 9 ] [ 37 ] Много исследований было посвящено сопоставлению экологии ушастых сов с другими совами, а также иногда с дневными хищными птицами, особенно с точки зрения различий в пищевых привычках и предпочтениях среды обитания. В Европе, возможно, самой густонаселенной совой является желто -коричневая сова , поэтому ее часто сравнивают с ушастыми совами. Хотя предпочтения добычи желтовато-коричневых сов совпадают с предпочтениями ушастой совы, они, как правило, демонстрируют большую гибкость в питании, внося больший вклад второстепенной добычи, такой как птицы, рептилии, амфибии и насекомые. Например, на юге Швеции ширина пищевой ниши коричневых сов была примерно в три раза больше, чем у ушастых сов. Обычно коричневые совы предпочитают более лесистую среду, чем ушастые совы, но могут адаптироваться так же, как и ушастые, к фрагментации лесов и пригородам , таким образом, часто конкурируя довольно напрямую за пищу. В то время как серая неясыть обычно выбирает немного крупную добычу в среднем, чем ушастые совы, средняя масса добычи часто превышает 30 г (1,1 унции), изученные размеры обыкновенных полевок, на которых охотятся в центральной Литве, показали, что как ушастые, так и серо-коричневые совы выбирали добычу крупнее, чем ушастые совы. средние полевки, средний пойманный вес оценивается в 21,45 г (0,757 унции) по сравнению со средним весом, исследованным людьми, в 16,42 г. (0,579 унции). Удивительно, но в этом исследовании, особенно в период отсутствия размножения, ушастые совы в Литве, как правило, в среднем выбирали более крупных полевок (21,56 г (0,761 унции)), чем серо-коричневые совы (19,56 г (0,690 унции)). [ 113 ] [ 210 ] [ 211 ] [ 212 ] Несмотря на потенциальную конкуренцию и смертность (для ушастых сов) в межвидовых отношениях между коричневыми и ушастыми совами, близость коричневых сов в исследовании, проведенном в Швейцарии, по-видимому, не оказала вредного влияния на размножение длинноухих сов. ушастый вид. [ 213 ] В нескольких исследованиях изучались повадки маленькой совы , которая, вероятно, является следующей по распространенности европейской совой и опережает ушастую сову на третьем месте, а также ушастую сову в районах, где их несколько перекрывающиеся предпочтения среды обитания приводят их в схожие районы. . Тем не менее, ушастая сова в среднем обычно ловит более крупную добычу, чем маленькие совы, поскольку она больше ориентируется на грызунов, а не на беспозвоночных, таких как насекомые и дождевые черви , и, особенно зимой, меньше меняет состав своей добычи, чем более мелкие виды. Кроме того, ушастой сове требуется несколько деревянных мест для ночлега, в то время как маленькие совы могут адаптироваться как к безлесным, так и к частично засаженным деревьями территориям, а также, по крайней мере в Испании , к более сильно измененным человеком территориям. [ 81 ] [ 146 ] [ 184 ] [ 214 ] Многие дневные хищники в Европе в значительной степени совпадают в пищевых привычках, в основном поедая полевок там, где они доступны, включая большинство видов луней , канюков и некоторых соколов , особенно обыкновенных пустельг ( Falcotinnunculus ). За исключением случайных хищнических взаимодействий, конкуренция с большинством этих дневных хищных птиц ограничена из-за временных различий в их привычках. [ 4 ] [ 37 ] [ 215 ] [ 216 ] В Европе несколько других сов, от гораздо меньших до нескольких видов, намного крупнее ушастой совы, предпочитают полевок и/или леммингов в качестве добычи, но часто значительно различаются по предпочтениям среды обитания, распространению, привычкам гнездования и/или охоте. привычки, поэтому ушастые совы в значительной степени естественным образом отделены от прямой конкуренции с ними. [ 4 ] [ 37 ]
В более широком масштабе и ушастые совы , и сипухи потенциально конкурируют за ресурсы с ушастыми совами. Несмотря на значительное совпадение как в ареале, так и в питании, ушастые совы в значительной степени защищены от серьезной конкуренции с короткоухими совами из-за предпочтений в среде обитания, поскольку ушастые совы всегда предпочитают более открытые места обитания, обычно устраивая ночевки и гнездясь в в высокой траве. высокой траве, а не древесину и редко, если вообще когда-либо, охотятся в тех же окраинных местообитаниях, что и ушастые виды. [ 4 ] [ 37 ] [ 141 ] На западе России ушастая сова по сравнению с ушастой совой проявляла меньшую склонность появляться в скоплениях, где была сосредоточена добыча, скорее всего, из-за различий в среде обитания. Как на западе России, так и на востоке Западной Сибири короткоухие, как правило, ловили больше тундровых полевок в дополнение к обыкновенным полевкам, в то время как ушастые совы больше ориентировались исключительно на обыкновенных полевок. [ 148 ] [ 217 ] Сравнение сипух и ушастых сов проводилось во многих районах, где ареал последнего вида менее обширен. Среды обитания, используемые сипухой и ушастой совой, не являются взаимоисключающими, как и виды их добычи. Несмотря на то, что сипухи являются похожими воздушными охотниками на открытых пространствах, их жизненный цикл во многом отличается от ушастых сов, отчасти тем, что они гнездятся в дуплах. Хотя их пищевые привычки могут показаться схожими и демонстрировать аналогичную зависимость от мелких млекопитающих, сипухи являются несколько более универсальными и католическими в плане кормления, с меньшей зависимостью от полевок. Совы-сипухи могут неплохо жить практически на любом скоплении мелких млекопитающих. В некоторых частях Европы ширина пищевой ниши этих двух видов сопоставима или даже немного выше у длинноухих. Однако в глобальном масштабе сипухи гораздо более широко распространены, более тропически в своем центральном ареале и в глобальном масштабе имеют гораздо более широкий спектр добычи, чем ушастые совы. Совы-сипухи также более адаптированы к изолированному образу жизни, чем ушастые совы, и способны умело питаться разнообразными классами добычи даже в условиях полного отсутствия мелких млекопитающих. [ 129 ] [ 138 ] [ 184 ] [ 185 ] [ 215 ] [ 218 ] [ 219 ] [ 220 ] Американская раса сипух крупнее, чем западная раса сипух из Европы и соответствующих частей Азии, и по массе тела, а также размеру ступней и когтей более сопоставима с желто-коричневой совой, в то время как западная раса в Европе занимает примерно промежуточное положение между желто-коричневой и длинной сипухой. Ушастые совы по размерам тела и размаху ног. Следовательно, в Америке сипухи, как правило, постоянно ловят немного более крупную добычу, чем ушастые совы, и имеют тенденцию получать доступ к более широкой общей кормовой нише . [ 4 ] [ 108 ] [ 164 ] [ 167 ] Несмотря на большую приспособляемость и ареал обитания сипух, исследование, проведенное во Франции, показало , что у них более высокая смертность от голода, чем у ушастых и желтовато-коричневых сов. у сипух, Это было связано с меньшими запасами липидного жира когда они пытаются пережить более прохладные сезоны в умеренном климате , поскольку они лучше приспособлены к выживанию в более теплом климате. [ 221 ]
В Северной Америке обитает больше видов сов, чем в Европе, и ее можно считать более конкурентной средой для обитающих там ушастых сов. Однако, опять же, в большинстве случаев предпочтения среды обитания, небольшое разделение диетических предпочтений (которые могут касаться видов добычи или размеров тела выбранной добычи) и истории жизни обычно позволяют большинству видов сохраняться, даже когда они живут рядом друг с другом. [ 4 ] [ 9 ] Как и во всем своем ареале, ушастые совы, как правило, отличаются от большинства других североамериканских сов тем, что по своей природе они гораздо более мигрирующие. Следовательно, циклические изменения в питании влияют на них по-разному и с меньшей вероятностью станут прямой причиной зимней смертности, чем у сов, живущих постоянно. [ 32 ] [ 101 ] [ 222 ] Как ни странно, среди американских сов ушастые совы наиболее сильно повторяют гораздо меньшую по размеру северную точильную сову по распространению, миграционным привычкам и, в меньшей степени, пищевым привычкам. Основной пищей совы-пилы, как правило, являются мыши Peromyscus там, где они доступны, и, как и у большинства сов, их выбор пищи для грызунов может во многом совпадать с выбором ушастой совы. [ 97 ] [ 98 ] [ 115 ] [ 156 ] Было высказано предположение, что менее широкое распространение ушастых сов в целом по сравнению с их европейским ареалом отчасти связано с большей конкуренцией за ресурсы между ними и двумя широко распространенными визжащими совами , восточной ( Megascops asio ) и западной визжащими совами. ( Мегаскопс Кенникотти ). [ 4 ] Тем не менее, кормовая ниша у визговых сов, как правило, гораздо шире и более оппортунистична, поскольку они часто питаются разнообразными беспозвоночными (в основном насекомыми, но также и раками ) и альтернативной добычей (например, более высокий баланс птиц и лягушек). вряд ли сможет конкурентно ограничить численность грызунов в ущерб ушастым совам. [ 9 ] [ 32 ] [ 115 ] [ 170 ]
Наиболее легко наблюдаемая и самая драматичная часть межвидовой экологии сов и хищников — это межвидовое хищничество. В связи с этим ушастые совы гораздо чаще оказываются жертвами, а не хищниками. Напротив, их в целом успешная жизненная история часто делает ушастых сов, возможно, более уязвимыми, чем большинство других сов среднего размера, для хищников и даже, возможно, для многих более мелких видов сов. К ним относится гнездование в относительно открытых заброшенных птичьих гнездах, а не в труднодоступных полостях. Кроме того, ушастые совы склонны обитать на довольно открытых участках во время охоты в сочетании с громкими вокалами (включая просьбы их птенцов) и другими слуховыми проявлениями, которые могут привлечь хищников. [ 4 ] [ 37 ] [ 41 ] [ 79 ] [ 223 ] В Европе их наиболее серьезными хищниками, как правило, являются евразийский филин и северный ястреб-тетеревятник ( Accipiter gentilis ). В одном отчете зарегистрировано 768 случаев хищничества со стороны филинов и 317 случаев нападения ястребов-тетеревятников (или 55% добычи совы, зарегистрированной для ястребов-тетеревятников в Европе). [ 4 ] [ 37 ] [ 79 ] Ушастые совы входят в пятерку наиболее часто встречающихся видов птиц, являющихся хищниками филинов в Европе. [ 79 ] [ 224 ] [ 225 ] Некоторые биологи считают, что склонность ушастых сов избегать более богатых концентраций добычи в пользу более открытых мест обитания и использования пространства, особенно во время миграции и зимовки, частично продиктована обнаружением активности филина (и, возможно, ястреба-тетеревятника), поэтому орел -сова оказывает серьезное влияние на историю жизни ушастой совы. [ 4 ] [ 36 ] [ 43 ] В целом ушастые совы терпят нападения огромного количества дневных хищных птиц в Европе. Вполне вероятно, что хищники, охотящиеся в основном на окуня, такие как Бутео и большинство орлов, могут либо случайно наткнуться на насест совы во время охотничьего набега, либо встретить ушастую сову, возможно, во время перекрывающейся активности на рассвете или в сумерках, в то время как крупные соколы , скорее всего, нападут только на одну из них. вниз, когда сова краснеет в дневное время. Ястребы-тетеревятники и другие ястребы- ястребы представляют собой особенно смертельную угрозу среди дневных хищников из-за их интенсивных методов поисковой охоты и готовности нырять в лесные заросли, где обитают сови. [ 37 ] [ 79 ] [ 226 ] [ 227 ] Помимо ястребов-тетеревятников, дневные хищники в Европе, которые, как известно, являются хищниками ушастых сов потенциально любого возраста, включают беркута ( Aquila chrysaetos ), орла Бонелли ( Aquila fasciata ), восточного могильника ( Aquila heliaca ) и большого подорлика. ( Clanga clanga ), Малый подорлик ( Clanga pomarina ), черный коршун ( Milvus migrans ), красный коршун ( Milvus milvus ), орлан-белохвост ( Haliaeetus albicilla ), обыкновенный канюк , зимняк ( Buteo lagopus ), сапсан ( Falco peregrinus ), балобан ( Falco cherrug ) и даже (в двух случаях) немного меньший по размеру евразийский перепелятник ( Accipiter nisus ). [ 4 ] [ 79 ] [ 228 ] [ 229 ] [ 230 ] [ 231 ] [ 232 ] [ 233 ] [ 234 ] Помимо филинов, неясыти и уральские совы ( Strix uralensis ) регулярно убивают ушастых сов там, где встречаются их ареалы, хотя в некоторых случаях это может начинаться с территориальных атак совы Стрикс , более мелкие ушастые совы могут быть съедены независимо от этого. . [ 4 ] [ 79 ]
В Северной Америке ареал хищников ушастой совы не менее устрашающий. Несомненно, самым опасным американским хищником является большая рогатая сова . Как и его европейский аналог филин, рогатые совы убивают ушастых сов независимо от сезона и условий. Однако, несмотря на многочисленные сообщения о хищничестве, ушастые совы, как правило, не занимают столь заметное место в рационе больших рогатых сов, по крайней мере, насколько это документально подтверждено, как в рационе евразийских филинов. Полосатые совы также могут быть серьезными хищниками для ушастых сов. [ 85 ] [ 159 ] [ 175 ] [ 235 ] Сообщается, что более редкие нападения на ушастых сов в Северной Америке совершали пятнистые совы ( Strix occidentalis ) и даже их родственники, ушастая сова . [ 4 ] [ 236 ] Дневные хищники ушастых сов, включая некоторые виды, которые также охотятся на них в Европе, такие как беркуты, ястребы-тетеревятники и сапсаны, а также белоголовый орлан ( Haliaeetus leucocephalus ), ястреб Купера ( Accipiter cooperii ), краснохвостый ястреб ( Buteo jamaicensis ) и красноплечий ястреб ( Buteo lineatus ). [ 13 ] [ 32 ] [ 175 ] [ 237 ] [ 238 ] [ 239 ] [ 240 ] Хотя ушастые совы менее хорошо документированы на всем ареале, они также уязвимы для хищников-млекопитающих, в основном вблизи гнезда. Подозреваемыми или подтвержденными хищниками в Европе часто являются европейские лесные или каменные куницы ( Martes foina ), которые, скорее всего, нападут на птенцов, но также будут поедать яйца и взрослых особей, если им удастся устроить на них засаду. [ 213 ] [ 241 ] [ 242 ] Куницы также представляют потенциальную угрозу в Северной Америке, как и североамериканский дикобраз ( Erethizon dorsatum ), бычья змея ( Pituophis catenifer ) и особенно енот ( Procyon lotor ) (последний вид может полурегулярно убивать и поедать высиживающую взрослую самку). ушастые совы). [ 13 ] [ 32 ] [ 175 ] [ 240 ] [ 243 ] [ 244 ] Врановые , многие из которых строят гнезда, которые используют ушастые совы, такие как сороки и вороны , также полурегулярно совершают набеги на гнезда ушастой совы и поедают яйца или птенцов. [ 13 ] [ 37 ] [ 32 ] [ 241 ] С другой стороны уравнения, ушастые совы сами могут нечасто охотиться на более мелких сов. Известно, что этот вид охотится на восточных сов , маленьких сов , евразийских карликовых сов ( Glaucidium passerinum ) и бореальных сов ( Aegolius funereus ), а также на молодь обыкновенной пустельги . [ 4 ] [ 13 ] [ 79 ] [ 85 ] [ 190 ]
Разведение
[ редактировать ]
Ушастые совы, как правило, моногамны. Немигрирующие популяции обычно моногамны в течение всего года, связь пары возобновляется ежегодно. [ 8 ] [ 36 ] Исследование, проведенное в Айдахо, установило, что ушастые совы локально чрезвычайно моногамны, при этом у 59 птенцов из 12 гнезд не обнаружено никаких внепарных оплодотворений. [ 245 ] В Нидерландах зарегистрирована единственная запись о спаривании самца с двумя самками, что является весьма нетипичным случаем. [ 8 ] Аналогичным образом, доказательства полиандрии и алло-родительства были обнаружены в гнезде в западной Монтане, где у четырех потомков самки совы было два отца, один из которых был родственником самки. [ 246 ] Самцы заявляют права на свою территорию пением и демонстрируют полеты хлопаньем крыльев. [ 8 ] Пение обычно начинается в сумерках тихим вечером и может продолжаться всю ночь, предпочтительны ясные, безветренные, лунные ночи. Песня обычно раздается с насеста, чаще всего на средней высоте на деревьях или с верхней половины у крон, иногда в полете. [ 8 ] [ 9 ] Самцы реагируют на воспроизведение в период, предшествующий сезону размножения (особенно во время ухаживания), до такой степени, что 45% территорий были бы незамеченными в Испании, если бы самцы не реагировали на записи (в отличие от прослушивания спонтанных криков). . [ 247 ] Часто в разных частях ареала на типичном расстоянии 100 км регистрируется от 8 до 50 пар. 2 (39 квадратных миль). В Шотландии около 17% из 9-18 пар на 10 км. 2 (3,9 квадратных миль) не занимались разведением. [ 12 ] Типичные территории для пар составляют от 50 до 100 га (от 0,19 до 0,39 квадратных миль) в Финляндии, когда полевок много. Тем не менее несколько пар могут гнездиться довольно близко друг к другу. Минимальное расстояние между активными гнездами обычно составляет от 50 до 150 м (от 160 до 490 футов), но в Айдахо было зарегистрировано расстояние до ближайшего гнезда всего 16 м (52 фута). [ 8 ] [ 13 ] [ 36 ] При обилии пищи на площади до 100 км2 могут гнездиться около 10-12 пар, реже до 50. 2 (39 квадратных миль). Пары могут терпеть друг друга на одном участке деревьев во время гнездования, если запасы еды в изобилии. [ 8 ] [ 36 ] Исследование проведенное в провинции Пиза в Италии, выявило 10-15 пар на 100 км. 32 территорий, 2 (39 квадратных миль) со средним расстоянием между гнездами 1727 м (5666 футов). [ 233 ] В районе исследования на юге и центральной части Айдахо пара встречалась в среднем на расстоянии 0,65 км (0,40 мили) друг от друга. [ 248 ] Самая высокая плотность может быть в Центральной Европе . Средняя плотность в Центральной Европе обычно составляет около 10-12 пар на 100 км. 2 (39 квадратных миль). [ 48 ] Однако в Бранденбурге , Германия , в ходе 24-летнего исследования было обнаружено, что средняя плотность составила 72,7 пар на 100 км. 2 (39 квадратных миль). [ 249 ] По данным исследований в Мичигане и Вайоминге, от 10 до 100 пар на 100 км. 2 (39 квадратных миль), при этом средний ареал в прибрежной среде обитания Вайоминга составляет около 55 га (0,21 квадратных миль). [ 9 ] [ 13 ] [ 82 ] Плотность оказалась ниже в районе реки Снейк (0,28–0,42 пары на квадратный километр), чем в других местах южного Айдахо (0,64–1,55 пары на квадратный километр). [ 9 ] [ 13 ] Во всем мире многие пары ушастой совы занимают одну и ту же территорию круглый год, но большинство из них предпочитают каждый год использовать новое гнездо, даже если то же гнездо, что и в прошлом году, все еще находится в хорошем состоянии. Самки обычно берут на себя обязанность осматривать потенциальные места гнездования и дуэты со своим партнером; сидя в выбранном гнезде, она поет, чтобы связаться с самцом, позже голосовая активность ограничивается слабыми криками, слышимыми только на близком расстоянии. [ 8 ] [ 13 ] Во время демонстраций также было зарегистрировано, что самки хлопают крыльями, но гораздо менее интенсивно, чем самцы. [ 48 ] Копуляция обычно происходит недалеко от гнезда. В Германии было зарегистрировано, что самец предшествует совокуплению криками и демонстрационными полетами, за которыми следуют сильные взмахи крыльями и наклоны тела, когда они сидят рядом с самкой и / или гнездом. [ 37 ] Спаривание также наблюдалось как на земле, так и на деревьях, которому часто предшествовал дуэт, воздушное представление самца, которое заканчивалось спуском на землю и полетом самки. [ 13 ] [ 37 ] Пограничные драки между самцами у этого вида не зафиксированы. [ 9 ] Размножение происходит позже в году, чем у симпатрических видов, таких как коричневые совы и сипухи, возможно, из-за их более активного миграционного характера. В регионах с более прохладным умеренным климатом спаривание может происходить в редких случаях уже в феврале, но совокупление пар обычно происходит в марте или апреле. [ 4 ] были зарегистрированы новые птенцы в возрасте 14–18 дней Зимнее размножение выявлено по мере того, как в начале февраля в Словакии . Другие предыдущие сообщения о зимнем размножении известны в двух случаях из Италии, а также из Чехии. [ 250 ] [ 251 ]


Обычно гнездится в гнездах крупных птиц, например, Corvus , Pica , хищников и цапель Ardea . Среди других строителей гнезд в Европе часто могут быть обыкновенные лесные голуби и евразийские перепелятники . [ 8 ] [ 36 ] В Великобритании и Финляндии 84% из 239 и 85 гнезд были сделаны падалью ( Corvus corone ) или воронами ( Corvus cornix ) и евразийскими сороками . В Англии и Финляндии 77% и 66% гнезд приходилось на хвойные породы соответственно. [ 14 ] [ 37 ] В Бранденбурге, Германия, 90% гнезд было построено воронами-падальщиками, и большая часть гнезд принадлежала Pinus sylvestris . [ 249 ] В различных исследованиях Айдахо почти все известные гнезда находились в старых врановых гнездах (большинство из них - в можжевельнике ). В одном крупном исследовании высота гнезда, которая в среднем составляла 3,2 м (10 футов), и диаметр конструкции гнезда, который в среднем составлял 22,3 см (8,8 дюйма), были признаны наиболее важными критериями выбора гнезда. [ 13 ] [ 167 ] [ 248 ] В Онтарио обычно использовались хвойные породы, часто Pinus или Juniperus , в гнездах врановых птиц высотой от 2,5 до 18,5 м (от 8,2 до 60,7 футов), но в основном от 5,5 до 9 м (от 18 до 30 футов). [ 83 ] Высота гнезда в Великобритании составила в среднем 6,7 м (22 фута). [ 14 ] В Словении ушастые совы предпочитали для размножения хвойные деревья (почти исключительно в гнездах врановых), у тех, которые гнездились на лиственных деревьях, наблюдалась более высокая смертность в гнездах, особенно в начале сезона, из-за более высокой численности хищников. [ 241 ] Точно так же выбор места для гнезда в значительной степени коррелировал с риском нападения хищников в Испании, где покрытие плющом и древесным пологом менее важно, чем густой кустарник снизу, отчасти потому, что наземные хищники были более распространены в конкретной зоне исследования, чем воздушные. [ 252 ] На северо-востоке Швейцарии места гнездования выбраны с учетом особенностей защиты от хищников: среди 38 мест гнездования те, которые, по-видимому, предпочитали более густые опушки леса, больший полог и находились в хвойных породах в гораздо большей степени, чем это было распространено в окружающей среде в целом, в то время как окрестности построек на территории исследования избегали. [ 213 ] Необычный случай городского размножения ушастых сов наблюдался в Москве , Россия, где рыхлой колонии наблюдалось образование . Гнездование в Москве считалось результатом синантропизации сов, отчасти из-за более низкого риска нападения хищников в городских районах по сравнению с сельскими окрестностями (где уровень хищничества был на 6,6% выше). Среднее расстояние гнездования сов в Москве составляло 603 м (1978 футов), тогда как в близлежащих сельских районах оно более чем в два раза превышало расстояние. [ 253 ] Гнезда на деревьях обычно находятся на высоте менее 30 м (98 футов) над землей и иногда могут быть настолько маленькими, что снизу могут быть видны крылья и хвост насиживающей самки. [ 36 ] Реже, чем птичьи древесные гнезда, могут использоваться листовые гнезда древесных белок , гнезда ястребов в кактусах и скальные гнезда различных птиц. [ 13 ] [ 14 ] [ 32 ] [ 254 ] Хотя они обычно занимают уже заброшенные места гнездования, иногда ушастые совы способны прогнать предыдущего обитателя гнезда, даже включая других хищников (вплоть до свирепых ястребов , таких как перепелятники, остроногие ястребы и даже более крупные ястребы Купера), что указывает на их потенциал жестокости и упорства. [ 4 ] Помимо гнезд других птиц, использовались и альтернативные места гнездования, но они обычно встречаются редко или необычно. Среди них есть неглубокие углубления на земле. [ 8 ] [ 32 ] [ 85 ] Некоторые зарегистрированные места наземных гнезд (в Европе) включали среди вереска , папоротника и ежевики и даже на тростниковых зарослях кроличьи норы . Некоторые гнезда были зафиксированы в плетеных корзинах , поставленных на деревьях для уток . [ 4 ] [ 14 ] [ 255 ] [ 256 ] [ 257 ] В Северной Америке пары наземных гнезд были обнаружены в каждой западно-центральной Монтане и в Окавагане, Британская Колумбия, во всех случаях между корнями или землей, непосредственно прилегающей к основанию деревьев или кустов (с двумя другими историческими записями о наземных гнездах). гнездование в Северной Америке). [ 175 ] [ 258 ] На местном уровне также принимаются искусственные гнездовья для сов из веток. В заповеднике Вудвалтон-Фен на востоке Англии было построено 71 гнездо в плетеных корзинах, предназначенных для сов. [ 259 ] Также в Изреэле в Израиле 6 из 16 корзин-гнезд, подвешенных на эвкалипте , использовались ушастыми совами, и все они были заняты к февралю. На этой территории Израиля было установлено еще как минимум 72 корзины-гнезда для ушастых сов, чтобы поощрить птиц, контролирующих грызунов. [ 260 ] В исключительных случаях ушастые совы гнездятся в неглубоких дуплах, дуплах ив или дубов, пнях деревьев или дуплах скал, однако, как правило, они не гнездятся в дуплах. [ 4 ] [ 261 ] 6,5% из 153 гнезд в Великобритании находились на естественных поверхностях (в основном на земле), а не в гнездах животных. [ 14 ] Окрестности гнездования, где предыдущие попытки были успешными, с большей вероятностью будут повторно использованы, например, в Айдахо, где 48% предыдущих гнездовий были повторно использованы после успешных попыток. [ 9 ] [ 13 ]
Откладка яиц обычно происходит с конца марта по начало мая на большей части ареала. Даты кладки яиц на северо-востоке США , в выборке из 42 яиц, были между 14 марта и 30 мая. Дополнительные записи на юге Канады показывают, что кладка произошла уже 5 июня. [ 9 ] [ 85 ] В период с 19 марта по 24 мая в Онтарио было отложено 43 яйца, более половины - с 15 апреля по 5 мая. [ 83 ] В северной Италии средняя дата кладки яиц приходится на 27 марта. [ 233 ] Исключительные яйца были зарегистрированы еще с 31 декабря по 3 января в Испании. [ 262 ] Повторное гнездование может произойти примерно через 20 дней после потери кладки. [ 85 ] В Европе в периоды изобилия пищи успешно выращивали до двух выводков. [ 9 ] [ 37 ] Обычно самка откладывает 3–5 яиц (иногда и больше, если пищи необычайно много). Более крупные кладки типичны севернее ареала. [ 8 ] [ 9 ] Рекордные по размеру кладки были зарегистрированы для Европы (до 8 яиц) в Швеции в годы «чумы полевок», а рекордные по размеру кладки во всем мире были зарегистрированы в 10–11 яиц в Казанском районе России в аналогичных условиях. [ 4 ] [ 36 ] [ 68 ] В Великобритании средний размер кладки был зафиксирован как 3,9, в Германии - 5,5, а в Словении - 5,6. [ 14 ] [ 263 ] [ 264 ] В исследовании, проведенном в Монтане, средний размер кладки составил 5. [ 265 ] Чисто белые яйца имеют размеры 40,2 × 32,5 мм (1,58 × 1,28 дюйма) в среднем как в Северной Америке, так и в Центральной Европе, а вес около 23 г (0,81 унции). [ 8 ] [ 85 ] [ 266 ] С интервалом 1-5 дней (в среднем 2) яйца откладываются на дно гнездового участка. Кладка из 7 яиц откладывается от 10 до 11 дней. [ 8 ] [ 9 ] Инкубация начинается с первого яйца и продолжается около 27–28 дней (в крайних случаях примерно от 21 до 30 дней). [ 8 ] [ 9 ] [ 174 ] Насиживает только самка, а самец доставляет пищу, которую приносят прямо в гнездо. Она может рано покинуть гнездо, чтобы кормиться, но в период инкубации делает это гораздо реже. [ 8 ] [ 9 ] Исследование, проведенное в Монтане, показало, что уровень кортикостерона был значительно выше у взрослых особей обоих полов в период размножения, чем в период отсутствия размножения, что позволяет предположить, что сезон размножения является более стрессовым для сов. [ 267 ]
Молодь вылупляется с двухдневным интервалом в любой момент с конца апреля по июнь. [ 4 ] [ 174 ] Средний вес 52 птенцов в Монтане составил 18,4 г (0,65 унции). [ 265 ] Их глаза открываются на 5–7 день (в среднем 6,4 дня в Монтане), и мать высиживает их около 2 недель, часто пока самец сидит рядом и наблюдает за ними. Самка кормит птенцов одна. В Айдахо было зарегистрировано, что самец приносил в 2,5 раза больше добычи во время гнездования, чем самка. [ 8 ] [ 99 ] [ 265 ] Демонстрации имитации травм были зафиксированы на стадии птенцов в качестве меры борьбы с хищниками. [ 8 ] [ 9 ] [ 64 ] Обычно ушастые совы менее смело защищают гнездо, чем некоторые другие совы, Стрикс например, совы , но, тем не менее, они способны на яростную защитную атаку. Защита гнезда со стороны родителей усилилась в Италии в период размножения, причем старшие птенцы защищаются более энергично. Самки выполняют большую часть защиты гнезда. Пары сов, подвергающиеся более высокому уровню регулярного беспокойства, с большей вероятностью будут иметь более мягкую защиту гнезда. [ 4 ] [ 99 ] Во время защитной демонстрации родитель взъерошивает свое оперение и частично расправляет крылья до полуоткрытого состояния, переступая с одной ноги на другую, шипя и щелкая клювом, и в этой позе может выглядеть на удивление большим. Если предполагаемая угроза сохраняется в сторону сов, они могут вскочить и попытаться схватить угрозу когтями. Даже такие крупные животные, как человек, могут оказаться жертвой защитных атак ушастой совы, если они приблизится или, особенно, если они заберутся к гнезду. [ 9 ] [ 36 ] Птенцы покидают гнездо в возрасте 20–27 дней (в среднем около 22 дней), но изначально не летают и часто лазают по окружающим ветвям. На этом этапе их можно назвать «ветвями». [ 8 ] [ 264 ] [ 265 ] Довольно часто молодые падают на землю, но обычно им удается подняться обратно, используя когти и клюв и тяжелые взмахи крыльев. В сумерках ветки-попрошайки звонят своим родителям высокими нотами, чтобы указать свое местонахождение. Примерно в 35–37 дней они становятся полностью оперенными и могут хорошо летать, но часто следуют за своими родителями и кормятся ими примерно до 2 месяцев, продолжая издавать пронзительные крики. [ 8 ] [ 9 ] [ 265 ] В Словении независимость была получена примерно через 50–80 дней после вылупления, а в Айдахо — примерно через 45–56 дней. [ 99 ] [ 268 ]

Обычно в Северной Америке этот вид производит одну кладку в год, но в годы высоких полевок зарегистрировано 2 кладки в год. [ 8 ] В Айдахо был зарегистрирован исключительный двойной выводок из-за большого количества пищи, что позволило паре успешно вырастить всех 11 птенцов до птенцов, в то время как в том же сезоне 3 другие самки в той же роще смогли произвести в среднем 5,3 птенца в своих одиночных выводках. [ 269 ] Как и другие виды, использующие открытые гнезда, а не закрытые полости, этот вид имеет относительно короткий период оперения и быстро покидает опасное место гнездования. [ 36 ] В исследовании 112 гнезд в Айдахо средний успех гнездования составил 46% за два года исследования, при этом хищничество енотов считалось наиболее серьезной причиной неудачного гнездования. [ 13 ] Другое исследование 24 гнезд в Айдахо показало, что совы оперяются в среднем по 3,7 детенышей на гнездо. [ 248 ] В Монтане среднее число вылупившихся особей на одно гнездо составило 3,8, а среднее число оперившихся особей на одно гнездо составило примерно 2,2. [ 265 ] В Великобритании 59% из 78 попыток гнездования оказались неудачными, при этом средний размер кладки на успешную пару составлял от 3,91 до 4,53. 41 из 78 особей, находящихся под постоянным наблюдением в этом британском исследовательском гнезде, успешно произвели на свет 1 или более птенцов. [ 14 ] В другом британском исследовании из 58 пар, за которыми наблюдали в течение 4 лет, 83% отложили яйца, 63% вывели одного или нескольких детенышей и 57% оперили детенышей, при этом в среднем на одно успешное гнездо оперилось 3,2 детеныша. [ 12 ] В еще одном английском исследовании, на этот раз исключительно сов, использующих плетеные корзины, в 50,7% из 71 попытки оперились детеныши. Среди пользователей плетеных корзин те, кто раньше гнездился (т. е. в марте-начале апреля), выращивали более крупные выводки и имели больше птенцов, чем позже (конец апреля - начало мая), отчасти из-за повышения уровня грунтовых вод, что делало поимку добычи менее идеальной в конце сезона гнездования. [ 259 ] Для 6 пар ушастых сов, использующих плетеные корзины в Израиле, средний размер выводка составил 3,6, а среднее количество молодых сов, покинувших гнездо, составило 3. [ 260 ] На всех стадиях и во всех регионах воспроизводство имеет тенденцию быть более успешным, когда популяция жертв выше. [ 12 ] [ 14 ] В континентальной Европе условия лучше, чем в Англии или Финляндии, возможно, из-за большой популяции обыкновенных полевок, которых нет в более северных странах, а средний успех гнездования выше. [ 9 ] [ 14 ] [ 37 ] Однако успех размножения по-прежнему зависит от популяции добычи. В 15 км. 2 Площадь (5,8 квадратных миль) на юге Германии, один холодный источник с небольшим количеством полевок, гнездящихся пар не обнаружено. Год спустя, с теплой весной и большим количеством полевок, в районе исследований поселилось 19 гнездящихся пар. [ 263 ] Смертность ушастых сов в первый год жизни в Германии составила 52% и 31%. [ 73 ] В Бранденбурге, Германия, из 867 попыток размножения 36,6% (335) пар удалось вырастить 1468 детенышей, что соответствует 1,57 птенцам на все попытки размножения, 4,31 на успешную пару. Данные Бранденбурга показывают, что после 1990 года, когда началась инициатива по сохранению сельского хозяйства, численность значительно возросла. Также в Бранденбурге в одном случае были убиты две последовательные самки одного и того же самца, самец смог спариться с третьей самкой, что привело к позднему успешному окрылению (24 августа). [ 249 ] В Словении, согласно исследованиям, проведенным в период с 1984 по 1993 год, из 79 гнезд 32 (40%) дали молодняк, 37 (47%) полностью погибли и 10 (12%) погибли после вылупления. Среднее количество вылупившихся сов в Словении составило 2,4 на гнездо (5,3 на удачное гнездо), а среднее количество оперившихся сов составило 1,6 на гнездо (3,9 на удачное гнездо). [ 264 ] В Пизе , Италия , пары ушастых сов дали в среднем 0,95 на территориальную пару и 2,13 на успешную пару. [ 233 ] В центральной Словении 57 гнезд давали в среднем 5,7 яиц на кладку. Из 51 оперившейся совы 31 погибла в ходе исследования, 22 из которых произошли из-за хищничества млекопитающих и птиц, 6 из-за голода, 2 из-за дорожно-транспортных происшествий и 1 утонула в канаве. [ 264 ] В Великобритании наиболее распространенной диагностированной причиной разрушения гнезд была кража яиц людьми (28,2% из 46 разрушенных гнезд). [ 14 ] Исследования по кольцеванию показывают, что у ушастой совы обычно короткая продолжительность жизни: более 91% из 105 выловленных в Северной Америке сов определенного возраста составляют 4 года или младше. Самый старый из зарегистрированных в ходе этих усилий особей был окольцован в Нью-Йорке и обнаружен в Онтарио в возрасте 11 лет и 4 месяцев. Однако другому могло быть больше 15 лет. [ 100 ] Известен один исключительный единичный экземпляр европейской ушастой совы возрастом чуть менее 28 лет. [ 8 ] [ 48 ] Годовая выживаемость взрослых в Германии и Швейцарии составляет 69%. [ 73 ]

Статус
[ редактировать ]
Ушастая сова довольно обычна и широко распространена во многих регионах. Имея ареал в 80 миллионов квадратных километров, это одна из наиболее широко встречающихся сов. По оценкам МСОП , общая популяция составляет от 2 до 5,5 миллионов, что делает ее одной из самых многочисленных сов после широко распространенной сипухи и менее широко распространенной маленькой совы (вероятно, от 5 до 10 миллионов птиц в целом) и примерно равной численности Общая популяция больших рогатых сов. [ 1 ] [ 270 ] Ушастая сова более многочисленна, чем ее более редкая, но более широко распространенная родственница, короткоухая сова, при этом максимальное возможное количество короткоухих примерно эквивалентно минимальному количеству длинноухих. [ 1 ] Диапазон их размножения обычно находится в засушливом и жарком климате средиземноморского типа, где наземные насекомые и рептилии могут преобладать над мелкими общительными грызунами, ушастая неясыть отрезана, тогда как это пиковые районы для сипух . На севере она почти отсутствует в районах глубокого бореального леса и безлесных северных районах, таких как большие заболоченные болота или нижняя тундра , где ушастая сова имеет тенденцию вытеснять их. [ 4 ] [ 8 ] Плотность населения зависит от наличия пищи. Местные угрозы обычно представляют собой пестициды и преследования . Из-за мифов и невежества этот вид долгое время подвергался преследованиям со стороны людей. [ 8 ] [ 36 ] Во многих регионах некоторые люди могут неосторожно стрелять в любое похожее на ворону гнездо, чтобы уничтожить его содержимое, из-за негодования по поводу того, что ворона воспринимается как вредитель. [ 8 ] В Северной Америке охотники исторически часто отстреливали ушастых сов, поскольку все совы подвергались жестоким преследованиям. Несмотря на то, что в начале 20-го века считалось, что этот вид на самом деле полезен для интересов человека, многие охотники продолжали отстреливать их, утверждая, что они представляют собой молодых особей презираемой тогда на национальном уровне большой рогатой совы. [ 85 ] [ 159 ] [ 240 ] Во многом из-за преследований ушастые совы были зарегистрированы как многочисленные, а затем редкие в Северной Америке в начале 20 века. [ 271 ] Ушастые совы потенциально сталкиваются со смертельным загрязнением тяжелыми металлами, такими как ртуть , органическими биоцидами , включая инсектициды , фунгициды и родентициды , а также ПХД зарегистрированными . Пестициды, похоже, влияют на этот вид меньше, чем на хищников с более разнообразным рационом и на тех, кто питается падалью. [ 4 ] Высокие уровни загрязнения были обнаружены в пометах ушастой совы в Сербии , при этом концентрации загрязняющих веществ в добыче происходят в результате промышленной и сельскохозяйственной практики в этом регионе. [ 272 ] Как и многие птицы, они могут быть уязвимы для гельминтов , которые, вероятно, недооцениваются как потенциальный источник смертности. [ 273 ] [ 274 ] Другие паразиты и клещи могут ухудшить здоровье населения. [ 4 ] [ 275 ] Вирус Западного Нила и сальмонелла были зарегистрированными источниками смертности некоторых ушастых сов. [ 276 ] [ 277 ] Антикоагулянты также могут угрожать этому виду. [ 278 ] На Канарских островах его становится все меньше из-за утраты среды обитания . [ 4 ] Многие ушастые совы также погибают в результате дорожного движения . Высокая смертность сов была особенно отмечена вдоль дорог во Франции, особенно из-за большого количества обыкновенных полевок на придорожных территориях. Было высказано предположение, что рост растительности может снизить смертность в некоторых случаях. [ 279 ] В 1963–1995 гг. из 128 мертвых ушастых сов, обнаруженных в Англии, 89 были самками и 34 самцами; 61% обратившихся погибли в результате столкновений (40% из них с транспортными средствами). В этом исследовании, проведенном в Англии, высокие уровни ДДЕ (метаболита ДДТ ) и HEOD, обнаруженные у ушастых сов до 1977 года, уровни, аналогичные тем, которые наблюдались у соколов, снизились в более поздних пробах, но у двух птиц все еще был смертельный уровень загрязнения пестицидами. . [ 280 ] В таких регионах, как Швейцария, чтобы компенсировать преследования и повысить выживаемость, а также облегчить наблюдателям возможность наблюдать за ними, зимующим совам в городах и поселках ежедневно кормили белых лабораторных мышей. [ 48 ] Размножению могут способствовать искусственные площадки из веток кустов или деревьев. Кроме того, усиленное кормление может обеспечить выживание в зимние периоды с низким уровнем корма. [ 8 ] Попытка выпустить выращенных в неволе ушастых сов на волю в Италии оказалась в значительной степени неудачной. Из них 3 из 8 сов, похоже, успешно рассеялись, в то время как остальные умерли или исчезли, а 8 коричневых неясытей выпустили всех, кроме одной, похоже, успешно рассеялись. [ 281 ] Количество видов в Иране фактически увеличилось : с 25 записей за 12 десятилетий до 49 за два десятилетия (с конца 70-х по 1997 год). Гнездование персидской гнездящейся популяции было подтверждено в 12 регионах, а в период с 1997 по 2014 год было зарегистрировано 32 негнездовых и 17 гнездовых записей. [ 282 ]
Считается, что в южной Калифорнии ( округ Сан-Диего и округ Ориндж ) ушастые совы потеряли более 55% своего ареала из-за изменений среды обитания. [ 240 ] Тенденция к уменьшению численности зимующих сов наблюдалась в Нью-Джерси : 9 из 58 известных насестов были полностью уничтожены из-за освоения земель, а остальные 49 показали сокращение численности или больше не использовались в течение 30 лет. Изменения в землепользовании и разрушение среды обитания приводят к снижению качества среды обитания и снижению численности полевок, при этом более адаптируемые хищники, такие как большие рогатые совы и ястребы, эксплуатируют большую часть того, что осталось, в ущерб ушастым совам. [ 283 ] Аналогичное снижение было отмечено за 20 лет в Пенсильвании . [ 284 ] Более широкие исследования кольцевания по всей Канаде, проведенные в ходе долгосрочного мониторинга на основе ежегодных подсчетов птиц в 1966–1992 годах, показали, что численность ушастых сов относительно значительно сократилась. За время исследования было зафиксировано чистое общее снижение на 0,98%. Это считалось вторым по величине сокращением численности после ушастой совы и роющей совы ( Athene cuncularia ). Более того, среди 19 хищников, обследованных в Канаде, у этих трех сов наблюдалась наибольшая тенденция к снижению численности. Ушастая сова, по-видимому, была самой редкой из шести исследованных суббореальных видов сов. Подобная тенденция была обнаружена по всей Северной Америке с очень большим чистым сокращением на 1,6% в целом во время рождественского подсчета птиц (CBC), что снова делает это самое серьезное снижение для совы, уступая только ушастой сове и двум родственным видам сов. произошло самое серьезное сокращение численности среди всех 28 видов хищников, упомянутых в этих исследованиях CBC. [ 18 ]
В искусстве
[ редактировать ]Джон Джеймс Одюбон проиллюстрировал «Ушастую сову - Strix otus » на табличке 383 в книге «Птицы Америки» , изданной в Лондоне, 1827–1838 гг. Гравюра была выгравирована Робертом Хавеллом в 1837 году. Оригинальная акварель была куплена Нью-Йоркским историческим обществом у обездоленной вдовы Одюбона.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д BirdLife International (2018). « Асио отус » . Красный список исчезающих видов МСОП . 2018 : e.T22689507A131922722. doi : 10.2305/IUCN.UK.2018-2.RLTS.T22689507A131922722.en . Проверено 12 ноября 2021 г.
- ^ «Приложения | СИТЕС» . сайт цитирует . Проверено 14 января 2022 г.
- ^ Перейти обратно: а б Олсен, П.Д. и Маркс, Дж.С. (2019). Северная ушастая сова (Asio otus) . В: дель Ойо, Дж., Эллиот, А., Саргатал, Дж., Кристи, Д.А. и де Джоанна, Э. (ред.). Справочник птиц мира живых. Lynx Editions, Барселона.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак с как в В из хорошо топор является тот нет бб до нашей эры др. быть парень бг чб с минет БК с бм млрд быть б.п. БК бр бс БТ этот бв б бх к бз что CB Вус, К.Х. (1988). Совы Северного полушария . Массачусетский технологический институт Пресс, ISBN 0262220350 .
- ^ Джоблинг, Джеймс А. (2010). Словарь научных названий птиц Хелма . Лондон: Кристофер Хелм. стр. 57, 286. ISBN. 978-1-4081-2501-4 .
- ^ Перейти обратно: а б с д и Вейк, Фридхельм (2007). Совы (Strigiformes): аннотированный и иллюстрированный контрольный список . Спрингер. ISBN 978-3-540-39567-6 .
- ^ Совы мира: Фотопутеводитель Микколы, H. Firefly Books (2012), ISBN 9781770851368
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак с как в В из хорошо топор является тот нет бб до нашей эры др. быть парень бг чб с минет БК с бм млрд быть б.п. БК бр бс БТ этот бв б бх Король, Клаус; Вейк, Фридхельм (2008). Совы мира (2-е изд.). Лондон: Кристофер Хелм. ISBN 9781408108840 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также Джонсгард, Пенсильвания (1988). Североамериканские совы: биология и естественная история . Смитсоновский институт.
- ^ Перейти обратно: а б Киат Ю., Перлман Г., Балабан А., Лешем Ю., Ижаки И. и Чартер М. (2008). Специализация питания городских ушастых сов Asio otus (Linnaeus, 1758) в Иерусалиме, Израиль . Зоология на Ближнем Востоке, 43 (1), 49-54.
- ^ Перейти обратно: а б с Трухильо О., Диас Г. и Морено М. (1989). Кормление ушастой совы (Asio otus canariensis) на острове Гран-Канария (Канарские острова) . Ардеола, 36 (2), 193–231.
- ^ Перейти обратно: а б с д и ж Деревня, А. (1981). Рацион и размножение ушастой совы в зависимости от численности полевок . Исследование птиц, 28(3), 214–224.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п Маркс, Дж. С. (1986). Характеристики места гнездования и репродуктивный успех ушастых сов на юго-западе Айдахо . Бюллетень Уилсона, 547–560.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н Клей, DE (1977). Биология размножения ушастой совы . Британские птицы, 70 (8), 318–331.
- ^ Перейти обратно: а б Хьюстон, CS (1997). Кольцевание сов Азио в южно-центральном Саскачеване . В: Дункан, Джеймс Р.; Джонсон, Дэвид Х.; Николлс, Томас Х., ред. Биология и охрана сов Северного полушария: 2-й Международный симпозиум. Генерал Тех. Реп. NC-190. Сент-Пол, Миннесота: Министерство сельского хозяйства США, Лесная служба, Северо-Центральная лесная экспериментальная станция. 237-242. (Том 190).
- ^ Перейти обратно: а б Пировано А., Руболини Д. и де Микелис С. (2000). Заселение и поведение городских ушастых сов на зимних ночлегах при вечернем отлете . Итальянский журнал зоологии, 67 (1), 63-66.
- ^ Перейти обратно: а б с Босаковски, Т. (1984). Выбор места и поведение ушастой совы (Asio otus), зимующей в Нью-Джерси . Раптор Рес, 18(13), 7-142.
- ^ Перейти обратно: а б с д Кирк Д.А. и Хислоп К. (1998). Состояние популяции и последние тенденции среди канадских хищников: обзор . Биологическая охрана, 83 (1), 91–118.
- ^ Линней, Карл (1758). Система природы по трем царствам природы по классам, отрядам, родам, видам, с признаками, различиями, синонимами, местами (на латыни). Том. 1 (10-е изд.). Холмии (Стокгольм): Лоуренс Сальвиус. п. 92.
- ^ Бриссон, Матюрен Жак (1760). Орнитология, или Метод разделения птиц на отряды, секции, роды, виды и их разновидности (на французском и латинском языках). Полет. 1. Париж: Жан-Батист Бош. п. 28.
- ^ Перейти обратно: а б с д Джилл, Фрэнк ; Донскер, Дэвид; Расмуссен, Памела , ред. (январь 2021 г.). «Совы» . Всемирный список птиц МОК, версия 11.1 . Международный союз орнитологов . Проверено 27 мая 2021 г.
- ^ Перейти обратно: а б с д Уинк М., Эль-Сайед А.А., Зауэр-Гюрт Х. и Гонсалес Дж. (2009). Молекулярная филогения сов (Strigiformes) установлена на основе последовательностей ДНК митохондриального цитохрома b и ядерного гена RAG-1 . Ардеа, 97(4), 581-592.
- ^ Перейти обратно: а б с Рэнди Э., Фуско Г., Лоренцини Р. и Спина Ф. (1991). Дивергенция аллозимов и филогенетические взаимоотношения внутри Strigiformes . Кондор, 93 (2), 295–301.
- ^ Форд, Н.Л., и Мюррей, Б.Г. (1967). Ископаемые совы из местной фауны Хагермана (верхний плиоцен) Айдахо . Аук, 84 (1), 115–117.
- ^ Перейти обратно: а б Майр Э. и Шорт ЛЛ (1970). Видовые таксоны птиц Северной Америки: вклад в сравнительную систематику . Орнитологический клуб Наттолла.
- ^ Вус, К.Х. (1966). Распространение сов в Африке в связи с общими зоогеографическими проблемами . Страус, 37(суп1), 499-506.
- ^ Сноу, Д.В. (1978). Гнездо как фактор, определяющий размер кладки тропических птиц . Журнал орнитологии, 119 (2), 227–230.
- ^ Ли, М.Й., Ли, СМ, Чон, Х.С., Ли, Ш., Пак, Дж.И. и Ан, Дж. (2018). Полный митохондриальный геном северной ушастой совы (Asio otus Linnaeus, 1758), определенный с помощью секвенирования нового поколения . Митохондриальная ДНК, часть B, 3(2), 494-495.
- ^ Галеотти П., Пиластро А., Тавеккья Г., Бонетти А. и Конджиу Л. (1997). Генетическое сходство в общинных зимних убежищах ушастой совы: исследование по дактилоскопии ДНК . Молекулярная экология, 6(5), 429-435.
- ^ « Ушастая сова » . owlpages.com . Проверено 1 мая 2014 г.
- ^ « Ушастая сова » . АРКив . Дикий экран. Архивировано из оригинала 4 февраля 2013 г. Проверено 23 апреля 2013 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м Маркс, Дж. С., Д. Л. Эванс и Д. У. Холт (1994). Ушастая сова (Asio otus) , версия 2.0. В «Птицах Северной Америки» (А. Ф. Пул и Ф. Б. Гилл, редакторы). Корнеллская лаборатория орнитологии, Итака, Нью-Йорк, США.
- ^ Баэз, М. (1992). Зоогеография и эволюция орнитофауны Канарских островов . Серия научных статей Музея естественной истории округа Лос-Анджелес, 36, 425–431.
- ^ Годфри, В. Эрл (1947). «Новая ушастая сова» . Канадский полевой натуралист . 61 (6): 196–197. дои : 10.5962/стр.340926 . Хотя на титульном листе указан 1947 год, статья была опубликована только в 1948 году.
- ^ Штайнбах, Г. (1980). Мир сов. Гофман и Кампе.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В Хьюм, Р. (1991). Совы мира . Бегущая пресса, Филадельфия.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но Миккола, Х. (1983). Совы Европы . Т. и А.Д. Пойзер.
- ^ Хьюм Р., Стилл Р., Сваш А., Харроп Х. и Типлинг Д. (2016). Британские птицы: Путеводитель по птицам Британии и Ирландии . Издательство Принстонского университета.
- ^ Хищные птицы реки Снейк (1985). Бюро землеустройства США, Idaho Power Company, Айдахо. Отдел рыболовства и охоты, типография правительства США.
- ^ Спрант, А., и Мэй, Дж. Б. (1955). Хищные птицы Северной Америки . Опубликовано Харпером при спонсорской поддержке Национального общества Обубона.
- ^ Перейти обратно: а б с д Эрхарт, К.М., и Джонсон, Северная Каролина (1970). Размерный диморфизм и пищевые привычки североамериканских сов . Кондор, 72 (3), 251–264.
- ^ Перейти обратно: а б с д и Справочник CRC по массе тела птиц , автор Джон Б. Даннинг-младший (редактор). ЦРК Пресс (1992), ISBN 978-0-8493-4258-5 .
- ^ Перейти обратно: а б с д Эрритцо Дж. и Фуллер Р. (1999). Половые различия в зимнем распространении ушастой совы (Asio otus) в Дании и соседних странах . Фогельварте, 40, 80-87.
- ^ Перейти обратно: а б Харви П.В. и Риддифорд Н. (1990). Неравномерное соотношение полов у перелетных ушастых сов . Звон и миграция, 11 (3), 132–136.
- ^ Ульмшнайдер, HM (1992). Использование мест зимовки и гнездования ушастыми совами в районе хищных птиц реки Снейк, 1992 г. Проект исследования хищных птиц реки Снейк. Анну. Представитель Деп. США Интер., Бур. Land Management., Бойсе, штат Айдахо, 362-366.
- ^ Перейти обратно: а б Снайдер, Н. Ф., и Уайли, Дж. В. (1976). Половой размерный диморфизм у ястребов и сов Северной Америки (№ 20). Американский союз орнитологов .
- ^ Перейти обратно: а б с Риджуэй Р. и Фридман Х. (1914). Птицы Северной и Средней Америки: описательный каталог высших групп, родов, видов и подвидов птиц, которые, как известно, встречаются в Северной Америке, от арктических земель до Панамского перешейка, Вест-Индии и других островов Карибского моря. Море и Галапагосский архипелаг (Том 50). Типография правительства США.
- ^ Перейти обратно: а б с д и ж г Крамп, С.; Симмонс, КЕЛ (1980). Птицы Западной Палеарктики . Том. 2. Оксфорд: Издательство Оксфордского университета.
- ^ «Ушастая сова» . Все о птицах . Корнеллская лаборатория орнитологии .
- ^ Шнайдер, Х., и Нидл, В. (1968). Дружба с лесными животными .
- ^ Чермели Д., Росси О. и Наси Ф. (2012). Сравнение геометрических характеристик когтей хищных и нехищных птиц . Итальянский журнал зоологии, 79 (3), 410–433.
- ^ Хауэлл, С.Н., и Уэбб, С. (1995). Путеводитель по птицам Мексики и северной части Центральной Америки . Издательство Оксфордского университета.
- ^ Перейти обратно: а б с д и Дэвис А.Х. и Притерч Р. (1976). Полевая идентификация ушастой и ушастой совы . Британские птицы, 69, 281–287.
- ^ Перейти обратно: а б с Робертсон, Иэн С. (1982). Полевая идентификация ушастой и ушастой совы . Британские птицы 75 (5): 227-229.
- ^ Перейти обратно: а б Саттон, Пэт; Саттон, Клэй (2006). Птицы и наблюдение за птицами в Кейп-Мэй: что посмотреть, когда и куда идти . Книги Стэкпола. ISBN 978-0811731348 .
- ^ Харрис А., Такер Л. и Виникомб К. (1989). Полевой справочник Макмиллана по идентификации птиц . Макмиллан; стр. 147-149.
- ^ Кемп, Дж. Б. (1982). Длина хвоста ушастой и ушастой совы . Британские птицы, 75 (5): 230.
- ^ Эмсли, SD (1982). Остеологическая идентификация ушастой и ушастой совы . Американская античность, 47 (1), 155–157.
- ^ Перейти обратно: а б Норберг, Р.О. (1977). Возникновение и независимая эволюция двусторонней асимметрии ушей у сов и последствия для таксономии сов . Философские труды Лондонского королевского общества. Б. Биологические науки, 280(973), 375-408.
- ^ Перейти обратно: а б Коллетт, Р. (1881). Строение черепа и ушных отверстий североевропейских видов семейства Strigidae . Брёггер.
- ^ Перейти обратно: а б Шварцкопф, Дж. (1963). Морфофизиологические свойства слуховой системы птиц . Учеб. XIII Интер. Орнитол. Конгресс, 1059-1068.
- ^ Перейти обратно: а б Шварцкопф, Дж. (1962). К вопросу о направленном слухе у сов (Striges) . Журнал сравнительной физиологии, 45 (5), 570–580.
- ^ Перейти обратно: а б ван Дейк, Т. (1973). Сравнительное исследование слуха сов семейства Strigidae . Нет Дж. Зул; 23:131-167.
- ^ Перейти обратно: а б с д и ж Армстронг, штат Вашингтон (1958). Гнездование и пищевые привычки ушастой совы в Мичигане (Том 1) . Мичиганский государственный университет сельского хозяйства и прикладных наук.
- ^ Галеотти П., Тавеккья Г. и Бонетти А. (2000). Родительская защита у ушастых сов Asio otus: влияние стадии размножения, пола родителей и преследования со стороны человека . Журнал птичьей биологии, 31 (4), 431–440.
- ^ Перейти обратно: а б Жиге Ф. и Одевард А. (2017). Птицы Европы, Северной Африки и Ближнего Востока: Фотопутеводитель . Издательство Принстонского университета.
- ^ Перейти обратно: а б с Кейт, С. (1989). Птицы Ближнего Востока и Северной Африки: Путеводитель . Монографии Пойзера.
- ^ Перейти обратно: а б Дементьев Г.П., Гладков Н.А., Птушенко Е.С., Спангенберг Е.П., Судиловская А.М. (1966). Птицы Советского Союза, т. 1 . Израильская программа научных переводов, Иерусалим.
- ^ Перейти обратно: а б Бразилия, М. (2009). Птицы Восточной Азии: Китая, Тайваня, Кореи, Японии и России. А&С Черный.
- ^ Перейти обратно: а б с д и ж г Вайденсаул, С. (2015). Совы Северной Америки и Карибского бассейна . Хоутон Миффлин Харкорт.
- ^ Рувалькаба-Ортега, И., Салинас-Родригес, ММ, Крус-Ньето, Дж., и Гонсалес-Рохас, Дж.И. (2014). Первая запись о гнездовании ушастой совы (Asio otus) в Чиуауа, Мексика . Юго-западный натуралист, 59 (1), 135–139.
- ^ Саттлер, HR (1995). Книга североамериканских сов . Хоутон Миффлин Харкорт.
- ^ Перейти обратно: а б с Глуц фон Блотцхайм, ООН, Бауэр, К.М., и Беззель, Э. (1980). Справочник птиц Центральной Европы . Аудитория, Висбаден.
- ^ Шелли, СУМКА (1895). Гнездование ушастой совы (Asio otus) в Индии . Дж. Бомбей Нат. Хист. Сок., 10: 149.
- ^ Перейти обратно: а б Шао М. и Лю Н. (2006). Рацион ушастой совы Asio otus в пустыне северо-западного Китая . Журнал засушливых сред, 65 (4), 673-676.
- ^ Котлер, BP (1985). Хищничество сов на пустынных грызунах, различающихся по морфологии и поведению . Журнал маммологии, 66 (4), 824–828.
- ^ Перейти обратно: а б с Лидер З., Йом-Тов Ю. и Мотро У. (2008). Рацион ушастой совы в северной и центральной пустыне Негев, Израиль . Орнитологический журнал Уилсона, 120 (3), 641–646.
- ^ Перейти обратно: а б Стофлет, Джей-Джей (1959). Гнездовая концентрация ушастых сов в округе Кочиз, штат Аризона . Бюллетень Вильсона, 71 (1), 97–99.
- ^ Перейти обратно: а б с д и ж г час Миккола, Х. (1976). Совы убивают и убивают другие совы и хищники в Европе . Британские птицы, 69, 144–154.
- ^ Перейти обратно: а б Ашванден Дж., Биррер С. и Дженни Л. (2005). Являются ли экологически компенсационные территории привлекательными местами охоты для обыкновенной пустельги (Falcotinnunculus) и ушастой совы (Asio otus)? Журнал орнитологии, 146 (3), 279–286.
- ^ Перейти обратно: а б Мартинес, Х.А., и Зуберогойтия, И. (2004). Предпочтения среды обитания ушастых сов Asio otus и маленьких сов Athene noctua в полузасушливых условиях в трех пространственных масштабах . Исследование птиц, 51 (2), 163–169.
- ^ Перейти обратно: а б с Крейгхед, Джей-Джей, и Крейгхед, ФК (1969). Ястребы, совы и дикая природа . Книги Стэкпола.
- ^ Перейти обратно: а б с Пек, Г.К., и Джеймс, Р.Д. (1983). Гнездящиеся птицы Онтарио: нидиология и распространение (Том 1) . ПЗУ.
- ^ Стюарт, RE (1975). Гнездящиеся птицы Северной Дакоты . Центр экологических исследований Трехколледжа.
- ^ Перейти обратно: а б с д и ж г час я Бент, AC (1938). Истории жизни хищных птиц Северной Америки, часть 2 . Национальный муз. США. Булл, 170.
- ^ Вернер, Дж., и Босс, А.С. (1980). Дикая природа Калифорнии и места их обитания: западная Сьерра-Невада . Генерал Тех. Реп. PSW-GTR-37. Беркли, Калифорния: Опыт лесов и ареалов юго-западной части Тихого океана. Stn., Лесная служба Министерства сельского хозяйства США: 439 стр., 37.
- ^ Холт, Д.В. (1997). УШАЯ СОВА (ASIO OTUS) И ЛЕСНОЕ ХОЗЯЙСТВО . Дж. Раптор Рес, 31 (2), 175–186.
- ^ Нейман, В. (2005). Обзор хищных птиц и сов (Falconiformes и Strigiformes) на морских островах Явы: исправления и дополнения . Зоологический бюллетень Раффлза, 53 (2), 287–288.
- ^ Рид, Д.Г., Дойл, Ф.И., Кенни, Эй.Дж., и Кребс, СиДжей (2012). Некоторые наблюдения за ушастой совой, Asio flammeus, экология арктической тундры, Юкон, Канада . Канадский полевой натуралист, 125 (4), 307–315.
- ^ Перейти обратно: а б с Крейг, Э.Х., Крейг, Т.Х., и Пауэрс, Л.Р. (1988). Характер деятельности и использование гнездовых ушастых сов на домашних участках . Бюллетень Вильсона, 204–213.
- ^ Армстронг, Э.А. (1954). Поведение птиц при непрерывном дневном свете . Ибис, 96(1), 1-30.
- ^ Анриу, Ф. (2000). Ареал обитания и использование среды обитания ушастой совы на северо-западе Швейцарии . Журнал исследований хищников, 34(2), 93-101.
- ^ Лёви М. и Ригерт Дж. (2013). Ареал и землепользование городских ушастых сов . Кондор, 115 (3), 551–557.
- ^ Рассел, Р.В., Данн, П., Саттон, К., и Керлингер, П. (1991). Визуальное исследование мигрирующих сов в Кейп-Мей-Пойнт, штат Нью-Джерси . Кондор, 93 (1), 55–61.
- ^ Лундберг, А. (1979). Оседлость, миграция и компромисс: адаптация к нехватке мест гнездования и пищевая специализация у трех видов фенноскандинавских сов . Экология, 41(3), 273-281.
- ^ Перейти обратно: а б Оверскауг К. и Кристиансен Э. (1994). Соотношение полов случайно убитых ушастых сов Asio otus в Норвегии . Звонок и миграция, 15(2), 104–106.
- ^ Перейти обратно: а б Slack, RS, Slack, CB, Робертс, RN, и Эморд, DE (1987). Весенняя миграция ушастых сов и северных совы-пилы в районе Найн-Майл-Пойнт, штат Нью-Йорк . Бюллетень Уилсона, 480–485.
- ^ Перейти обратно: а б Даффи К. и Керлингер П. (1992). Осенняя миграция совы в Кейп-Мей-Пойнт, штат Нью-Джерси . Бюллетень Уилсона, 312–320.
- ^ Перейти обратно: а б с д Ульмшнайдер, Х. (1990). Постгнездовая экология ушастой совы (Asio otus) на юго-западе Айдахо .
- ^ Перейти обратно: а б с д Хьюстон, CS (2005). Ушастые совы, Asio otus: обзор кольцевания в Северной Америке . Канадский полевой натуралист, 119 (3), 395–402.
- ^ Перейти обратно: а б Шево М., Драпо П., Имбо Л. и Бержерон Ю. (2004). Зимние набеги сов как индикатор популяционных циклов мелких млекопитающих в бореальных лесах восточной части Северной Америки . Ойкос, 107(1), 190-198.
- ^ Ньютон, И. (2002). Ограничение численности голарктических сов . Экология и охрана сов, 3-29.
- ^ Давенпорт, DL (1982). Приток в Великобританию луней, ушастых и ушастых сов зимой 1978/79 г. Британские птицы, 75, 309–316.
- ^ Сойккели, М. (1964). Uber das Uberwinter und di Nahrung der valdohreule (Asio otus) на юго-западе Финляндии, 1962/63 год . Орнис Фенника, 31, 37–40.
- ^ Шнайдер, К.Дж. (2003). Статус и экология ушастой совы (Asio flammeus) в штате Нью-Йорк . Королевская птица, 53, 313–330.
- ^ Шариков А.В., Макарова Т.В., Ганова Е.В. (2014). Многолетняя динамика ушастой совы Asio otus на северных зимовках в европейской части России . Ардеа, 101(2), 171-177.
- ^ Макарова Т. и Шариков А. (2015). Выбор места зимнего ночлега ушастой совы в европейской части России . Дж. Раптор Рес, 49, 333–336.
- ^ Перейти обратно: а б с д и ж г Марти, компакт-диск (1974). Экология питания четырех симпатрических сов . Кондор, 76 (1), 45–61.
- ^ Дайс, LR (1945). Минимальная интенсивность освещения, при которой совы могут на глаз найти мертвую добычу . Американский натуралист, 79 (784), 385–416.
- ^ Перейти обратно: а б Гетц, Л. Л. (1961) Места охоты ушастой совы . Бюллетень Уилсона, 79–82.
- ^ Фаулер, Д.В., Фридман, Э.А., и Сканнелла, Дж.Б. (2009). Функциональная морфология хищников у хищников: межпальцевые различия в размере когтей связаны с техникой удержания добычи и иммобилизации . ПЛОС ОДИН, 4(11), е7999.
- ^ Уорд, AB, Вейгль, PD, и Конрой, RM (2002). Функциональная морфология задних конечностей хищников: значение для разделения ресурсов . Аук, 119 (4), 1052–1063.
- ^ Перейти обратно: а б Ребер, Х. (1949). Поведение содержащихся в неволе ушастых сов (Asio otus otus) и неясыти (Strix aluco aluco) по отношению к добыче . Поведение, 1-95.
- ^ Перейти обратно: а б с д и ж Биррер, С. (2009). Синтез 312 исследований по питанию ушастой совы Asio otus . Ардеа, 97(4), 615-625.
- ^ Перейти обратно: а б с д и ж г час я дж к л м Марти, компакт-диск (1976). Обзор выбора добычи ушастой совы . Кондор, 78 (3), 331–336.
- ^ Сунделл Дж., Уиту О., Хенттонен Х., Кайкусало А., Корпимяки Э., Пиетияйнен Х., Саурола П. и Хански И. (2004). Крупномасштабная пространственная динамика популяций полевок в Финляндии, выявленная успехом размножения птиц-хищников, питающихся полевками . Журнал экологии животных, 73 (1), 167–178.
- ^ Хафнер, DJ и Катцефлис, FM (2000). Североамериканские грызуны. Исследование состояния и План действий по сохранению . Дэвид Дж. Хафнер, Эрик Йенсен и Гордон Л. Киркланд (составители и редакторы); Группа специалистов МСОП/SSC по грызунам. МСОП, Гланд, Швейцария и Кембридж, Великобритания.
- ^ Перейти обратно: а б с д и ж Уттендорфер, О. (1952). Новые результаты по питанию хищных птиц и сов . Ойген Умер, Штутгарт, Германия.
- ^ Розенфельд Ф. и Добли А. (2000). Закапывание обыкновенных полевок (Microtus arvalis) в различные социальные среды . Поведение, 137 (11), 1443–1461.
- ^ Перейти обратно: а б с д и Шмидт, Э. (1975). Рацион ушастой совы (Asio otus) в Европе . Аквила, 80(81), 221-238.
- ^ Перейти обратно: а б Тулис Ф., Балаж М., Обуч Дж. и Шотнар К. (2015). Реакция рациона ушастой совы Asio otus и численности зимующих особей на изменение численности обыкновенной полевки Microtus arvalis . Биология, 70(5), 667-673.
- ^ Шариков А. и Макарова Т. (2014). Погодные условия объясняют разнообразие рациона ушастой совы на зимних ночлегах в центральной части Европейской России . Орнис Фенника, 91 (2).
- ^ Sharikov, A. V. (2006). Peculiarities of winter feeding in the Long-Eared Owl (Asio otus) in settlements of Stavropol Krai . Zoologicheskii Zhurnal, 85, 871-877.
- ^ Нистреану, В. (2007). Значение ушастой совы Asio otus otus (L.) в борьбе с грызунами . Вестник Университета сельскохозяйственных наук и ветеринарной медицины Клуж-Напока. Сельское хозяйство, 63.
- ^ Джейкоб Дж. И Браун Дж. С. (2000). Использование микросред обитания, отказ от плотности и временная активность как краткосрочное и долгосрочное поведение против хищников у обыкновенных полевок . Ойкос, 91(1), 131-138.
- ^ Гжендзицка, Э. (2014). Означает ли численность полевок Microtus spp. Все же определить численность зимующих ушастых сов Asio otus? Экология, 33(4), 354-364.
- ^ Перейти обратно: а б Корпимяки, Э. (1992). Состав рациона, выбор добычи и успех размножения ушастых сов: влияние многолетних колебаний изобилия пищи . Канадский журнал зоологии, 70 (12), 2373–2381.
- ^ Калландер, Х. (1977). Питание ушастой совы Asio otus в Швеции . Орнис Фенника, 54(2), 79–84.
- ^ Перейти обратно: а б с Ялден, Д.В. (1985). Диетическое разделение сов в Пик-Дистрикте . Исследование птиц, 32(2), 122–131.
- ^ Фэрли, Дж. С. (1967). Питание ушастых сов на северо-востоке Ирландии . Британские птицы, 60, 130–135.
- ^ Перейти обратно: а б Араужо Дж., Рей Дж. М., Ландин А. и Морено А. (1974). Вклад в изучение Бухо Чико (Asio otus) в Испании . Ардеола, 19, 397–428.
- ^ Перейти обратно: а б Эскала Дж., Алонсо Д., Масуэлас Д., Мендибуру А., Вилчес А., Арисага Дж. и Схема Р. (2009). Зимний рацион ушастой совы Asio otus в долине Эбро (северо-восток Иберии) . Каталонский орнитологический журнал, 25, 49–53.
- ^ Бертолино С., Гиберти Э. и Перроне А. (2001). Экология питания ушастой совы (Asio otus) на севере Италии: специалист-диетолог? Канадский журнал зоологии, 79 (12), 2192–2198.
- ^ Руболини Д., Пировано А. и Борги С. (2003). Влияние сезонности, температуры и количества осадков на зимний рацион ушастой совы Asio otus . FOLIA ZOOLOGICA-PRAHA-, 52(1), 67-76.
- ^ Перейти обратно: а б Сесере Ф. и Вичини Г. (2000). Микромлекопитающие в рационе ушастой совы (Asio otus) в оазисе Сан-Джулиано WWF (Матера, Южная Италия) . Хайстрикс, 11(2), 3-13.
- ^ Масутти Л., Паолуччи П. и Усберти А. (2008). Результаты осенне-зимнего рациона ушастой совы Asio otus (Linnaeus) в условиях равнины Ломбардии . УНИВЕРСИТЕТ ПАДУАНЫ.
- ^ Пировано А., Руболини Д., Брамбилла С. и Феррари Н. (2000). Зимний рацион городских ночевок Ушастые совы Asio otus в северной Италии: значение бурой крысы Rattus norvegicus . Исследование птиц, 47(2), 242–244.
- ^ Перейти обратно: а б с Аливизатос Х. и Гутнер В. (1999). Зимний рацион сипухи (Tyto alba) и ушастой совы (Asio otus) на северо-востоке Греции: сравнение . Журнал исследований хищников, 33 (2), 160–163.
- ^ Перейти обратно: а б Кафкалету-Диес А., Цахалидис Е.П. и Пуазидис К. (2008). Сезонные изменения в рационе ушастой совы (Asio otus) в северо-восточной сельскохозяйственной зоне Греции . Дж Биол Рес Салоник, 10, 181–9.
- ^ Сечкин С. и Джошкун Ю. (2005). Мелкие млекопитающие в рационе ушастой совы Asio otus из Диярбакыра, Турция . Зоология на Ближнем Востоке, 35 (1), 102–103.
- ^ Перейти обратно: а б Сельчук А.Ю., Банкоглу К. и Кефелиоглу Х. (2017). Сравнение зимнего рациона ушастой совы Asio otus (L., 1758) и ушастой совы Asio flammeus (Pontoppidan, 1763) (Aves: Strigidae) в Северной Турции . Acta Zoologica Bulgarica, 69(3), 345-348.
- ^ Хызал, Э. (2013). Рацион ушастой совы Asio otus в Центральной Анатолии (Aves: Strigidae) . Зоология на Ближнем Востоке, 59 (2), 118–122.
- ^ Чартер М., Ижаки И., Лешем Ю. и Рулен А. (2012). Рацион питания и успех размножения ушастых сов в полузасушливых условиях . Журнал засушливой среды, 85, 142–144.
- ^ Перейти обратно: а б Халегизаде А., Арбаби Т., Нури Г., Джавидкар М. и Шахриари А. (2009). Рацион зимующей ушастой совы Asio otus в Заболе, юго-восточный Иран . Ардеа, 97(4), 631-634.
- ^ Лю, Н.Ф., Чжао, JY, Чжао, В., Шао, MQ, и Сун, С. (2010). Сезонные изменения в рационе ушастой совы Asio otus в пустыне Северо-Западного Китая . Биология животных, 60 (2), 115–122.
- ^ Перейти обратно: а б Скотт, Д.М., Гладвин, К., и Бартон, Н. (2005). Сравнение рациона двух пустынных сов: ушастой совы (Asio otus) и маленькой совы (Athene noctua) из южной Монголии . Монгольский журнал биологических наук, 3 (1), 31-37.
- ^ Перейти обратно: а б Стуббе М., Бацайчан Н., Линдеке О., Самджаа Р. и Стуббе А. (2016). Новые данные по экологии питания Bubo bubo и Asio otus (Aves: Strigidae) в Монголии . Исследования биоразнообразия в Монголии, Vol. 13.
- ^ Перейти обратно: а б с Дюпал Т.А. и Чернышов В.М. (2013). Мелкие млекопитающие в рационе ушастой совы (Asio otus) и ушастой совы (A. flammeus) на юге Западной Сибири . Российский экологический журнал, 44(5), 397-401.
- ^ Чиба А., Онодзима М. и Киносита Т. (2005). Добыча ушастой совы Asio otus в пригороде города Ниигата в центральной Японии, как показал анализ погадок . Орнитологическая наука, 4 (2), 169–172.
- ^ Мацуока, С. (1974). Добыча, добытая ушастой совой Asio otus в период размножения на Хоккайдо . Журнал Института орнитологии имени Ямашина, 7(3), 324-329.
- ^ Кавагути С. и Ямамото Т. (2003). Пищевой продукт, обнаруженный в погадках ушастых сов, зимующих в Эхимэ, Япония . Японский журнал орнитологии, 52 (1), 29–31.
- ^ Манро, Дж. А. (1929). Заметки о пищевых привычках некоторых хищников Британской Колумбии и Альберты . Кондор, 31 (3), 112–116.
- ^ Холт, Д.В., и Чайлдс, Н.Н. (1991). Диета ушастых сов вне сезона размножения в Массачусетсе . Джей Раптор Рес, 25, 23-4.
- ^ Спайкер, CJ (1933). Анализ двухсот погадок ушастой совы . Бюллетень Вильсона, 45 (4), 198–198.
- ^ Кристенсон, Г., и Фуллер, MR (1975). Пищевые привычки двух семей ушастой совы в восточно-центральной Миннесоте . Лун, 47(2), 58-61.
- ^ Перейти обратно: а б с д Рэндл В. и Остин Р. (1952). Экологические заметки об ушастых и точильных совах на юго-западе Огайо . Экология, 33(3), 422-426.
- ^ Морган, С. и Спайс-младший, К. (1965). Зимний корм ушастой совы . Кингберд, 15: 222.
- ^ Перейти обратно: а б Рейнольдс, RT (1970). Наблюдения за гнездами ушастой совы (Asio otus) в округе Бентон, штат Орегон, с заметками об их пищевых привычках . Мюррелет. 51: 8-9., 51, 8-9.
- ^ Перейти обратно: а б с Эррингтон, Польша (1932). Пищевые привычки хищников южного Висконсина. Часть I. Совы . Кондор, 34 (4), 176–186.
- ^ Рейни, Д.Г., и Робинсон, Т.С. (1954). Еда ушастой совы в округе Дуглас, штат Канзас . Труды Канзасской академии наук, 57 (2), 206–207.
- ^ Кан, А.Р., и Кемп, Дж.Т. (1930). О еде некоторых сов в восточно-центральном Иллинойсе . Аука, 323–328.
- ^ Моррисси, TJ (1949). Зимний корм некоторых ушастых сов . Айова Берд Лайф, 19: 70–71.
- ^ Войт, Дж., и Гленн-Левин, округ Колумбия (1978). Доступность добычи и добыча ушастых сов в Айове . Американский натуралист из Мидленда, 162–171.
- ^ Перейти обратно: а б Бэрроуз, CW (1989). Рацион пяти видов пустынных сов . Западные птицы, 20 (1), 1–10.
- ^ Перейти обратно: а б Марти, К.Д., Маркс, Дж.С., Крейг, Т.Х., и Крейг, Э.Х. (1986). Диета ушастой совы на северо-западе Нью-Мексико . Юго-западный натуралист, 31 (3), 416–419.
- ^ Браун, Нидерланды (1995). Заметки о зимнем ночевке и питании ушастых сов в пустыне Сонора . Журнал исследований хищников, 29 (4), 277–279.
- ^ Перейти обратно: а б с д Маркс, Дж. С. и Марти, CD (1984). Экология питания симпатрических сипух и ушастых сов в Айдахо . Орнис скандинавский, 135–143.
- ^ Крейг, TH, Крейг, EH, и Пауэрс, LR (1985). Пищевые привычки ушастой совы (Asio otus) на коллективном ночлеге в период гнездования . Аук, 102 (1), 193–195.
- ^ Перейти обратно: а б с Маркс, Дж. С. (1984). Экология питания размножающихся ушастых сов на юго-западе Айдахо . Канадский журнал зоологии, 62 (8), 1528–1533.
- ^ Перейти обратно: а б с Маркс, Дж. С., и Маркс, В. А. (1981). Сравнительные пищевые привычки визжащей совы и ушастой совы на юго-западе Айдахо . Мюррелет, 62 (3), 80–82.
- ^ Гонсалес-Рохас, Дж.И., Падилья-Рангель, Х., Рувалькаба-Ортега, И., Крус-Ньето, Массачусетс, Каналес-дель-Кастильо, Р., и Гусман-Веласко, А. (2017). Зимний рацион ушастой совы Asio otus (Strigiformes: Strigidae) на лугах Яноса, Чиуауа, Мексика . Чилийский журнал естественной истории, 90 (1), 1.
- ^ Перейти обратно: а б Ноланд Р.Л., Максвелл Т.С. и Даулер Р.К. (2013). Пищевые привычки ушастой совы (Asio otus) на зимнем коллективном ночлеге в Техасе . Юго-западный натуралист, 58 (2), 245–248.
- ^ Перейти обратно: а б Клиппель, МЫ, и Пармали, П.В. (1982). Добыча зимующей ушастой совы в бассейне Нэшвилла, штат Теннесси . Журнал полевой орнитологии, 53 (4), 418–420.
- ^ Перейти обратно: а б с д Крейг, TH, и Трост, CH (1979). Биология и плотность гнездования гнездящихся американских пустельг и ушастых сов на реке Биг-Лост, юго-восточный Айдахо . Бюллетень Уилсона, 50–61.
- ^ Перейти обратно: а б с д и Булл, Э.Л., Райт, А.Л., и Хенджум, М.Г. (1989). Гнездование и рацион ушастой совы в хвойных лесах штата Орегон . Кондор, 91 (4), 908–912.
- ^ Том, Д. (1994). Состав рациона ушастой совы в центральной Словении: сезонные изменения в использовании добычи . Журнал исследований хищников, 28(4), 253-258.
- ^ Родригес, А. (2005). Хищничество ушастых сов на летучих мышах в Средиземноморье и регионах с умеренным климатом на юге Европы . Дж. Раптор Рес, 39 (4), 445–453.
- ^ Перейти обратно: а б Тянь Л., Чжоу X., Ши Ю., Го Ю. и Бао В. (2015). Летучие мыши как основная добыча зимующей ушастой совы (Asio otus) в Пекине: интеграция защиты биоразнообразия и городского управления . Интегративная зоология, 10 (2), 216–226.
- ^ Мори Э., Менчетти М. и Дартора Ф. (2014). Данные о поедании падали ушастой совы Asio otus (Linnaeus, 1758) (Aves: Strigiformes: Strigidae) . Итальянский журнал зоологии, 81 (3), 471–475.
- ^ Канова, Л. (1989). Влияние снежного покрова на выбор добычи ушастой совы Asio otus . Этология, экология и эволюция, 1 (4), 367–372.
- ^ Хартли, PHT (1947). Еда ушастой совы в Ираке . Ибис, 89(4), 566-569.
- ^ Канарио, Ф., Лейтан, А.Х., и Томе, Р. (2012). Попытки хищничества короткоухих и ушастых сов на перелетных певчих птиц привлекали искусственное освещение . Журнал исследований хищников, 46 (2), 232–235.
- ^ Боченски З.М. и Томек Т. (1994). Характер фрагментации костей птиц в погадках ушастой совы Asio otus и его тафономические последствия . Acta Zoologica Cracoviensia, 37(1).
- ^ Перейти обратно: а б с д Секур М., Базиз Б., Денис К., Думанджи С., Сутту К. и Гезуль О. (2010). Пищевой режим Cheveche d'Athena Athene noctua, de l'Effraie des Clochers Tyto alba, du Hibou moyen-duc Asio otus et du Grand-duc ascalaphe Bubo ascalaphus: заповедник природы Мергеба (Алжир) . Алауда, 78(2), 103-117.
- ^ Перейти обратно: а б Лидер З., Йом-Тов Ю. и Мотро У. (2010). Сравнение рациона двух симпатрических сов — Tyto alba и Asio otus — в пустыне Негев, Израиль . Израильский журнал экологии и эволюции, 56 (2), 207–216.
- ^ Хандверк, Дж. (1990). Ушастая сова (Asio otus) в Египте . Боннские зоологические статьи, 41, 171–179.
- ^ Мильчев Б., Боев З. и Тотева Т. (2003). Состав рациона ушастой совы (Asio otus) в осенне-зимний период в северном парке Софии . Годишник на Софийский университет «Св. Климент Охридский» Биологический факультет книги, 1, 93-94.
- ^ Де Ваврин, Х., Уолравенс, М., и Рабози, Д. (1991). Исключительные гнездования средней неясыти (Asio otus) и хохлатого сокола (Falcotinnunculus) в 1991 году в лесу Суань (Брабант) . Птицы, 28, 169–188.
- ^ Сметтан, Х. (1987). Результаты двенадцатилетних пищевых инспекций ушастой совы (Asio otus L.) в центральном Неккарланде/Баден-Вюртемберге с учетом сезонных изменений и динамики численности мелких млекопитающих . Орнитологические ежегодные журналы Баден-Вюртемберга. 3:1-52.
- ^ Перейти обратно: а б с Клей, DE (1972). Птичья добыча, добытая британскими совами . Исследование птиц, 19(2), 91–96.
- ^ Тротта, М. (2010). Первые данные о репродуктивной фенологии и питании ушастой совы Asio otus в заповеднике Децима-Малафеде (Рим) . Алула, 17 (1-2): 105-111.
- ^ Бон, М., Роккафорте, П., Боргони, Н., и Реджиани, П. (1998). Первые данные о питании ушастой совы Asio otus в провинции Венеции . Материалы 2-й Венецианской конференции фаунистов. Доп. Бык. гражданский Св. Нат. Венеция, 48, 186–189.
- ^ Галли, Л. (2015). Данные о рационе ушастой совы Asio otus на зимнем насесте в Империале (Западная Лигурия, Северная Италия) и заметки об их суточном цикле активности . Bollettino dei Musei e degli Istituti Biologici, 77.
- ^ Петреску, А. (1997). Остатки добычи Asio otus otus L. (Aves: Strigiformes), питающиеся летом в природном заповеднике Аджигея (Румыния) . Работы Национального музея естественной истории Григоре Антипа, 37, 305–317.
- ^ Перейти обратно: а б Шандор, А.Д., и Кисс, Б.Дж. (2008). Птицы в рационе зимующих ушастых сов (Asio otus) в дельте Дуная, Румыния . Журнал исследований хищников, 42 (4), 292–295.
- ^ Гарсиа А. и Сервера Ф. (2001). Заметки о сезонных и географических изменениях в рационе ушастой совы Asio otus . Ардеола, 48(1), 75-80.
- ^ Вейга, JP (1980). Кормовые и трофические взаимоотношения сипухи (Tyto alba) и ушастой совы (Asio otus) в горах Сьерра-де-Гуадаррама (Испания) . Ардеола, 25, 113–142.
- ^ Робинетт, Р. и Джеймс, К. (2001). Социальные и экологические факторы, влияющие на бдительность северо-западных ворон Corvus caurinus . Поведение животных, 62 (3): 447–452.
- ^ Саттон, GM (1926). Ушастая сова ловит рябчика . Аук, 43 (2), 236–237.
- ^ Коннелли, Дж.В., М.В. Гратсон и К.П. Риз (1998). Острохвостый тетерев (Tympanuchus phasianellus) , версия 2.0. В «Птицах Северной Америки» (А. Ф. Пул и Ф. Б. Гилл, редакторы). Корнеллская лаборатория орнитологии, Итака, Нью-Йорк, США.
- ^ Каррильо Дж., Ногалес М., Дельгадо Г. и Марреро М. (1989). Предварительные данные для сравнительного изучения особенностей питания Asio otus canariensis на Эль-Йерро и Гран-Канарии, Канарские острова . В Мейбурге Б.-У. И ред. Канцлера РД. Хищники в современном мире WWGBP: Берлин, Лондон и Париж.
- ^ Йосеф Р. и Мейром К. (2009). Далее о рационе зимующих ушастых сов Asio otus на севере Израиля . Рябок, 31: 106-108.
- ^ Шариков А., Шишкина Е.М. и Ковинка Т. (2018). Рыба в рационе ушастой совы Asio otus . Исследование птиц, 65 (2): 266–269.
- ^ Бохач Д. и Михалкова Д. (1970). Пища ушастого бычка (Asio otus) . Сильвия, 18 лет, 63–70.
- ^ Беззель, Э. (1972). Некоторые данные о питании верхнебаварской ушастой совы (Asio otus) . Нет. Орнитол. Гес Бавария, 11, 181–184.
- ^ Амат, Дж. А., и Соригер, Р.К. (1981). Сравнительный анализ рациона питания совы Tyto alba и неясыти Asio otus в Западной Испании . Алауда, 49(2), 112-120.
- ^ Сиах, М. (2006). Жук обыкновенный (Melolontha melolontha; Coleoptera: Scarabaeidae) в рационе ушастой совы Asio otus . Бутео, 15, 23-25.
- ^ Сутту, К., Манаа, А., Секур, М., Абабса, Л., Гезул, О., Бакрия, М., Думанджи, С. и Денис, К. (2015). Выбор добычи сипухой Tyto alba и ушастой совой Asio otus в сельскохозяйственных условиях в Эль-Маальбе (Джельфа, Алжир) . Ливанский научный журнал, 16 (2), 3–17.
- ^ Дельгадо, Г., Куилис, В., Мартин, А., и Эммерсон, К. (1986). Кормление ушастой совы (Asio otus) на острове Тенерифе и сравнительный анализ с рационом Tyto alba . Донана Acta Vertebrata, 13, 87–93.
- ^ Бальчаускас, Л., и Бальчаускене, Л. (2014). Избирательное хищничество серых неясытей и ушастых сов на обыкновенных полевок зимой и весной . Турецкий журнал зоологии, 38(2), 242-249.
- ^ Бальчаускене Л., Йовайшас А., Нарушявичюс В., Петрашка А. и Скуя С. (2006). Рацион неясыти (Strix aluco) и ушастой совы (Asio otus) в Литве, полученный из гранул . Acta Zoologica Lituanica, 16(1), 37-45.
- ^ Нильссон, Индиана (1984). Вес добычи, перекрытие пищи и репродуктивная способность потенциально конкурирующих ушастых и коричневых сов . Орнис Скандинавица, 176–182.
- ^ Перейти обратно: а б с Анриу, Ф. (2002). Выбор места гнездования ушастой совы Asio otus на северо-западе Швейцарии . Исследование птиц, 49(3), 250–257.
- ^ Романовский, Дж. (1988). Трофическая экология Asio otus (L.) и Athene noctua (Scop.) в пригороде Варшавы . Пол. Экол. Исследования, 14, 223–234.
- ^ Перейти обратно: а б Рулен, А. (1996). Зимнее питание сипухи (Tyto alba), ушастой совы (Asio otus), луня (Circus cyaneus) и обыкновенной пустельги (Falcotinnunculus) . Бюллетень Водуазского общества естественных наук, 84 (1), 19–32.
- ^ Лак, Д. (1946). Конкуренция хищных птиц за пищу . Журнал экологии животных, 123–129.
- ^ Волков С.В., Шариков А.В., Басова В.Б., Гринченко О.С. (2009). Влияние численности мелких млекопитающих на численность и выбор местообитаний ушастой (Asio otus) и короткоухой (Asio flammeus) сов . Энтомологическое обозрение, 88 (10), 1248–1257.
- ^ Капицци, Д., и Луиджи Кану, PV (1998). Пищевые привычки симпатрической ушастой совы Asio otus, неясытии Strix с тех пор и сипухи Tyto alba в прибрежных лесах Средиземноморья .
- ^ Китовски, И. (2013). Зимний рацион сипухи (Tyto alba) и ушастой совы (Asio otus) в Восточной Польше . Северо-западный зоологический журнал, 9 (1).
- ^ Петровичи М., Молнар П. и Сандор А.Д. (2013). Перекрытие трофических ниш двух симпатрических видов сов (Asio otus Linnaeus, 1758 и Tyto alba Scopoli, 1769) в северо-западной части Румынии . Северо-Западный зоологический журнал, 9 (2).
- ^ Массемин, С., и Хандрич, Ю. (1997). Более высокая зимняя смертность сипухи по сравнению с ушастой совой и неясытью: влияние липидных запасов и изоляции? Кондор, 99 (4), 969–971.
- ^ Кребс, CJ, Бутин, С. и Бунстра, Р. (2001). Экосистемная динамика бореального леса . Нью-Йорк, Проект Клуэйн.
- ^ Эррера, К.М., и Хиральдо, Ф. (1976). Пищево-нишевые и трофические взаимоотношения европейских сов . Орнис Сканд, 7(1), 29.
- ^ Бэйл П. и Шаулс Р. (2011). Биология четырех пар Великого Герцога Европы Бубо-бубо в Люксембурге . Бык. Соц. Нат. Люксембург, 112, 51.
- ^ Фёрстель, А. (1983). Увеличение популяции филина Bubo bubo в Баварии . Реклама Орнитологического общества Баварии, 22, 145–167.
- ^ Серхио Ф. и Хиральдо Ф. (2008). Внутригильдийное хищничество в сообществах хищников: обзор . Ибис, 150, 132-145.
- ^ Хаккарайнен, Х., и Корпимаки, Э. (1996). Конкурентные и хищнические взаимодействия среди хищников: наблюдательное и экспериментальное исследование . Экология, 77(4), 1134-1142.
- ^ Мэр-младший (2014). Исследование экологии питания орла Бонелли: влияние диеты на состояние тела, показатели жизнедеятельности и демографию . Университет Барселоны (Докторская диссертация)
- ^ Чавко Дж., Данко Ш., Обуч Дж. и Михок Дж. (2007). Питание орла-могильника (Aquila heliaca) в Словакии . Словацкий журнал Raptor, 1, 1-18.
- ^ Домбровский, В. (2010). Рацион большого подорлика (Aquila clanga) в Белорусском Полесье . Словацкий журнал Raptor, 4, 23–36.
- ^ Вяли, Ю. (2003). Малый подорлик и его охрана в Эстонии . Хирундо Саппл, 6(1), 66.
- ^ Ивановский, В.В. (2010). Орлан-белохвост Haliaeetus albicilla в Белорусском Поозерье: материалы по биологии вида в ареале . Русский орнитологический журнал, 19: 1876-1887.
- ^ Перейти обратно: а б с д Серджио Ф., Маркези Л. и Педрини П. (2008). Плотность, рацион и продуктивность ушастой совы Asio otus в итальянских Альпах: значение полевок Microtus . Исследование птиц, 55(3), 321–328.
- ^ Чавко Дж., Слободник Р., Дойчова Л., Липтак Дж., Михок Дж., Обух Дж. и Немчек В. (2014). Популяция, рацион и гнездовые ящики балобана (Falco cherrug) в Словакии: отчет проекта LIFE 2011–2014 . Словацкий журнал Raptor, 8(2), 73-86.
- ^ Холт, Д.В., и Биттер, К. (2007). Зимний рацион полосатой совы и размеры гранул в западной Монтане . Северо-западный натуралист, 88 (1), 7–12.
- ^ Форсман, ЭД, Энтони, Р.Г., Меслоу, ЕС, и Забель, CJ (2004). Рацион и пищевое поведение северных пятнистых сов в Орегоне . Дж. Раптор Рез. 38 (3): 214–230.
- ^ Шеррод, СК (1978). Рацион североамериканских соколообразных . Раптор Рес, 12(3/4), 49-121.
- ^ Олендорф, Р.Р. (1976). Пищевые привычки североамериканских беркутов . Американский натуралист из Мидленда, 231–236.
- ^ Палмер, Р.С. (Ред.). (1988). Справочник птиц Северной Америки, том VI: Дневные хищники (Часть 1) . Издательство Йельского университета.
- ^ Перейти обратно: а б с д Блум, PH (1994). Биология и современный статус ушастой совы в прибрежной южной Калифорнии . Бюллетень Академии наук Южной Калифорнии, 93 (1), 1–12.
- ^ Перейти обратно: а б с Том, Д. (2003). Выбор места гнездования и деспотическое распространение гнездящихся ушастых сов Asio otus, вызванное хищничеством . Журнал птичьей биологии, 34 (2), 150–154.
- ^ Шариков А.В., Волков С.В., Иванов М.Н. и Басова В.Б. (2010). Образование агрегированных поселений как выражение синантропизации ушастой совы (Asio otus L.) . Российский экологический журнал, 41(1), 44-50.
- ^ Хейворд, Г.Д., и Гартон, Э.О. (1984). Выбор места обитания тремя маленькими лесными совами . Бюллетень Уилсона, 96(4), 690-692.
- ^ Амструп, SC, и МакЭнини, TP (1980). Змея-бык убивает и пытается съесть птенцов ушастой совы . Бюллетень Уилсона, 92 (3), 402–402.
- ^ Маркс, Дж. С., Дикинсон, Дж. Л., и Хейдок, Дж. (1999). Генетическая моногамия у ушастых сов . Кондор, 101 (4), 854–859.
- ^ Маркс, Дж. С., Дикинсон, Дж. Л., и Хейдок, Дж. (2002). Серийная полиандрия и аллородительство у ушастой совы . Кондор, 104 (1), 202–204.
- ^ Мартинес, Х. А., Зуберогоития, И., Колас, Дж., и МА-СИА, Дж. (2002). Использование записывающих звонков для обнаружения ушастой совы Asio otus . Ардеола, 49(1), 97-101.
- ^ Перейти обратно: а б с Туроу, Т.Л., и Уайт, К.М. (1984). Успех гнездования и выбор добычи ушастых сов вдоль экотона можжевельника и полыни в южно-центральной части Айдахо . Мюррелет, 10–14.
- ^ Перейти обратно: а б с Блок, Б. (2009). Долгосрочные тенденции плотности популяции и репродуктивный успех ушастой совы Asio otus в Бранденбурге, Германия . Ардеа, 97(4), 439-444.
- ^ Нога, М. (2009). Зимнее размножение ушастой совы (Asio otus) в Юго-Западной Словакии . Словацкий журнал Raptor, 3, 61–62.
- ^ Гастин М., Провенца Н. и Сорас А. (2006). Первые упоминания о зимнем размножении ушастой совы в Италии . Журнал исследований хищников, 40 (3), 249–251.
- ^ Родригес А., Гарсия А.М., Сервера Ф. и Паласиос В. (2006). Ландшафт и факторы борьбы с хищничеством, определяющие выбор места гнезда, распределение гнезд и продуктивность средиземноморской популяции ушастых сов Asio otus . Ибис, 148(1), 133-145.
- ^ Шариков А.В., Волков С.В., Иванов М.Н. и Басова В.Б. (2010). Образование агрегированных поселений как выражение синантропизации ушастой совы (Asio otus L.) . Российский экологический журнал, 41(1), 44-50.
- ^ Миллсап, Бакалавр (1998). Ушастая сова (Asio otus) . В книге «Хищники Аризоны» под редакцией Р. Л. Глинского. Тусон: Univ. Аризона Пресс.
- ^ Грюнвальд, К. (1972). Ушастая сова (Asio otus) гнездится на земле . Орнитол. Середина 24: 80-82.
- ^ Мингоцци, Т. (1980). Наземное гнездование ушастой совы Asio otus в Пьемонте . Наши птицы 35: 369–371.
- ^ Хоскинг, Э.Дж., Ньюберри, К.В., и Смит, С.Г. (1945). Птицы Ночи . Коллинз.
- ^ Мэйплс, МТ, Холт, Д.В., и Кэмпбелл, Р.В. (1995). Наземно-гнездящиеся ушастые совы . Бюллетень Уилсона, 563–565.
- ^ Перейти обратно: а б Гарнер, диджей, и Милн, бакалавр наук (1998). Исследование ушастой совы Asio otus с использованием плетеных гнездовых корзин . Исследование птиц, 45(1), 62–67.
- ^ Перейти обратно: а б Чартер М., Лешем Ю. и Халеви С. (2009). Использование гнездовых корзин ушастыми совами Asio otus в Израиле . рябок, 31, 36-37.
- ^ Хавершмидт, Ф. (1946). Заметки о местах гнездования кулика-сорока и ушастой совы как нороковода . Британские птицы, 34, 334–336.
- ^ Коррал, Дж. Ф., Кортес, Дж. А., и Гил, Дж. М. (1979). Вклад в изучение питания Asio otus на юге Испании . Доньяна, Acta Vertebrata, 6 (2), 179–190.
- ^ Перейти обратно: а б Рокенбаух, Д. (1978). Биология размножения и факторы, контролирующие популяции коричневых сов (Strix aluco) и ушастых сов (Asio otus) в Швабском Альбе . Журнал орнитологии, 119 (4), 429–440.
- ^ Перейти обратно: а б с д Том, Д. (1997). Биология размножения ушастой совы (Asio otus) в центральной Словении . Folia Zoologica, 46(1), 43-48.
- ^ Перейти обратно: а б с д и ж Зайденстикер, М.Т., Флокхарт, ДТТ, Холт, Д.В., и Грей, К. (2006). Рост и развитие оперения птенцов ушастой совы . Кондор, 108 (4), 981–985.
- ^ Макач, В. (1976). Яйца птиц Европы . Лейпциг.
- ^ Ромеро, Л.М., Холт, Д.В., и Петерсен, Дж.Л. (2009). Эффекты приливов и сезонные изменения уровня кортикостерона у взрослых ушастых сов Asio otus . Ардеа, 97(4), 603-609.
- ^ Том, Д. (2011). Выживаемость после вылета и динамика расселения ушастой совы Asio otus . Исследование птиц, 58(2), 193–199.
- ^ Маркс, Дж. С., и Перкинс, А. Е. (1999). Двойное насиживание у ушастой совы . Бюллетень Вильсона, 273–276.
- ^ Зауэр, младший, Нивен Д.К., Хайнс Дж.Дж., Циолковски-младший DJ, Пардик К.Л., Фэллон Дж.Дж. и WA Link (2017). Североамериканское обследование гнездящихся птиц, результаты и анализ, 1966–2015 гг . Версия 2.07.2017. Центр исследования дикой природы USGS Патаксент, Лорел, Мэриленд, США.
- ^ Ветмор, А. (1935). Тенистые птицы ночи . National Geographic, 67 (2), 217–240.
- ^ Демайо, М.А., Цветичанин, Дж., Стоилькович, М., Трпков, Д., Андрич, В., Онья, А., и Нешкович, О. (2011). Обнаружение элементов и радиоактивности в погадках ушастой совы (Asio otus), населяющих город Белград (Сербия) . Химия и экология, 27(5), 393-400.
- ^ Бартлетт, CM, и Андерсон, RC (1987). Лемдана вернаарти н. сп. и другие филяриоидные нематоды из Bubo Virginianus и Asio otus (Strigiformes) в Онтарио, Канада, с пересмотренной версией Lemdana и ключом к родам птичьих филяриоидов . Канадский журнал зоологии, 65 (5), 1100–1109.
- ^ Феррер Д., Молина Р., Кастелла Дж. и Кинселла Дж. М. (2004). Паразитарные гельминты в пищеварительном тракте шести видов сов (Strigiformes) в Испании . Ветеринарный журнал, 167(2), 181-185.
- ^ Скорацкий М. и Бочков А.В. (2002). Новый вид гусиного клеща Bubophilus asiobius sp. n.(Acari: Syringophilidae) от ушастой совы Asio otus (Strigiformes: Strigidae) . Род, 13(1), 149-152.
- ^ Комар, Н. (2003). Вирус Западного Нила: эпидемиология и экология в Северной Америке. Достижения в исследованиях вирусов, 61, 185–234.
- ^ Молина-Лопес, Р.А., Вальверду, Н., Мартин, М., Матеу, Э., Обон, Э., Серда-Куэльяр, М., и Дарвич, Л. (2011). Дикие хищники как переносчики устойчивых к противомикробным препаратам штаммов Salmonella и Campylobacter . Ветеринарная запись, vetrecc7123.
- ^ Стоун, В.Б., Оконьевски, Дж.К., и Стеделин, младший (2003). Родентициды-антикоагулянты и хищники: недавние данные, полученные в Нью-Йорке, 1998–2001 гг . Бюллетень загрязнения окружающей среды и токсикологии, 70(1), 0034-0040.
- ^ Бодвен, Х. (1997). Смертность сипухи (Tyto alba) и ушастой совы (Asio otus) вдоль автомагистралей в Бургундии-Шампани: отчет и предложения . В: Дункан, Джеймс Р.; Джонсон, Дэвид Х.; Николлс, Томас Х., ред. Биология и охрана сов Северного полушария: 2-й Международный симпозиум. Генерал Тех. Реп. NC-190. Сент-Пол, Миннесота: Министерство сельского хозяйства США, Лесная служба, Северо-Центральная лесная экспериментальная станция. 58-61. (Том 190).
- ^ Уилли И., Дейл Л. и Ньютон И. (1996). Неодинаковое соотношение полов, причины смертности и остатки загрязняющих веществ у ушастых сов в Великобритании . Британские птицы, 89 (10), 429–436.
- ^ Чермели, Д. (2000). Поведение выращенных вручную осиротевших ушастых сов и неясытей после выпуска в дикую природу . Итальянский журнал зоологии, 67 (1), 57-62.
- ^ Халегизаде А., Тохидифар М., Мусави С.Б., Хашеми А., Хани А. и Омиди М. (2015). Увеличение популяции ушастой совы Asio otus (Linnaeus, 1758) в Иране (Aves: Strigidae) . Зоология на Ближнем Востоке, 61 (3), 215–219.
- ^ Босаковски Т., Кейн Р. и Смит Д.Г. (1989). Упадок ушастой совы в Нью-Джерси . Бюллетень Уилсона, 101(3), 481-485.
- ^ Смит, Д.Г. (1981). Верность зимнему месту ушастых сов в центральной Пенсильвании . Американские птицы, 35 (3), 1-339.
Внешние ссылки
[ редактировать ]- Ушастая сова на сайте owlpages.com включает фотографии и записи разговоров.
- Asio otus в Полевом справочнике: Птицы мира на Flickr
- «Асио отус» . Авибаза .
- Перья ушастой совы (Asio otus). Архивировано 3 декабря 2013 г. в Wayback Machine.
- «Северная ушастая сова медиа» . Интернет-коллекция птиц .
- Фотогалерея ушастой совы в VIREO (Университет Дрекселя)
| Базы данных органов управления : Национальные |
|---|