Гаплогруппа R1a
| Гаплогруппа R1a | |
|---|---|
 | |
| Возможное время происхождения | 22,000 [ 1 ] до 25 000 [ 2 ] много лет назад |
| Возможное место происхождения | Евразия |
| Предок | Гаплогруппа R1 |
| Потомки | R1a-Z282, R1a-Z93 |
| Определение мутаций |
|
| Самые высокие частоты | См. Список частот R1a по населению. |

Гаплогруппа R1a , или гаплогруппа R-M420 , представляет собой гаплогруппу ДНК Y-хромосомы человека , которая распространена в большом регионе Евразии , простирающемся от Скандинавии и Центральной Европы до Центральной Азии , Южной Сибири и Южной Азии . [ 3 ] [ 2 ]
Хотя одно генетическое исследование показывает, что R1a возник 25 000 [ 2 ] лет назад его субклад M417 (R1a1a1) диверсифицировал ок. 5800 лет назад. [ 4 ] Место происхождения субклада играет роль в споре о происхождении протоиндоевропейцев .
Мутация SNP R-M420 была обнаружена после R-M17 (R1a1a), что привело к реорганизации линии, в частности к созданию новой парагруппы (обозначенной R-M420*) для относительно редких линий, не входящих в R-SRY10831. .2 (R1a1) ответвление, ведущее к R-M17.
Происхождение
[ редактировать ]Происхождение R1a
[ редактировать ]По оценкам, генетическая дивергенция R1a (M420) произошла 25 000 лет назад. [ 2 ] лет назад, во время последнего ледникового максимума . Исследование Питера А. Андерхилла и др., проведенное в 2014 году с участием 16 244 человек из более чем 126 популяций со всей Евразии, пришло к выводу, что существует «убедительные доказательства того, что Ближний Восток, возможно, недалеко от современного Ирана, является географическим источником hg R1a». ". [ 2 ] Древние записи ДНК показали, что первый R1a появился во время мезолита у восточных охотников-собирателей (из Восточной Европы, около 13 000 лет назад). [ 5 ] [ 6 ] и самый ранний случай R* среди верхнего палеолита древних северных евразийцев , [ 7 ] от которого преимущественно ведут свое происхождение восточные охотники-собиратели. [ 8 ]
Диверсификация R1a1a1 (M417) и древние миграции
[ редактировать ]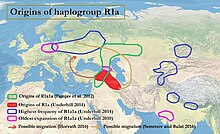
По данным Андерхилла и др. (2014) , нижележащий субклад R1a-M417 разделился на Z282 и Z93 около 5800 лет назад «в окрестностях Ирана и Восточной Турции». [ 4 ] [ примечание 1 ] Несмотря на то, что R1a встречается как гаплогруппа Y-хромосомы в различных языках, таких как славянский и индоиранский , вопрос о происхождении R1a1a имеет отношение к продолжающимся дебатам относительно урхеймата протоиндоевропейских народов , а также может быть имеет отношение к истокам цивилизации долины Инда . R1a показывает сильную корреляцию с индоевропейскими языками Южной Западной и Азии , Центральной и Восточной Европы и Скандинавии. [ 10 ] [ 3 ] наиболее распространен в Восточной Европе , Западной Азии и Южной Азии . В Европе особенно распространен Z282, а в Азии доминирует Z93. Связь между Y-ДНК R-M17 и распространением индоевропейских языков впервые была отмечена Т. Зерджалом и коллегами в 1999 году. [ 11 ]
Индоевропейские отношения
[ редактировать ]Предлагаемое распространение R1a1a в степи
[ редактировать ]Семино и др. (2000) предположили украинское происхождение и постледниковое распространение гаплогруппы R1a1 во время позднеледникового максимума , которое впоследствии усилилось за счет расширения курганной культуры в Европу и на восток. [ 12 ] Спенсер Уэллс предполагает центральноазиатское происхождение, предполагая, что распространение и возраст R1a1 указывают на древнюю миграцию, соответствующую распространению курганцев при их экспансии из евразийской степи . [ 13 ] По данным Памджава и др. (2012) , R1a1a распространена в Евразийских степях или в регионе Ближнего Востока и Кавказа:
Внутренняя и Центральная Азия представляют собой зону перекрытия линий R1a1-Z280 и R1a1-Z93, [что] подразумевает, что зона ранней дифференциации R1a1-M198 предположительно произошла где-то в Евразийских степях или в регионе Ближнего Востока и Кавказа, поскольку они лежат между Южной и Центральной Азией. Азия и Центральная и Восточная Европа. [ 14 ]
Три генетических исследования, проведенные в 2015 году, подтвердили курганскую теорию Гимбутаса относительно индоевропейского урхеймата . Согласно этим исследованиям, гаплогруппы R1b и R1a, наиболее распространенные сейчас в Европе (R1a также распространена в Южной Азии), должны были распространиться из Понтийско-Каспийских степей вместе с индоевропейскими языками; они также обнаружили аутосомный компонент, присутствующий у современных европейцев, которого не было у европейцев эпохи неолита, который должен был быть введен с отцовскими линиями R1b и R1a, а также с индоевропейскими языками. [ 15 ] [ 16 ] [ 17 ]
Сильва и др. (2017) отметили, что R1a в Южной Азии, скорее всего, «распространяется из одного пула источников в Центральной Азии , по-видимому, действительно существует по крайней мере три, а возможно, и больше клад-основателей R1a ; на Индийском субконтиненте , что соответствует множественным волнам прибытия». [ 18 ] По словам Мартина П. Ричардса, соавтора Silva et al. (2017) , распространенность R1a в Индии была «очень убедительным доказательством значительной миграции бронзового века из Центральной Азии, которая, скорее всего, привела в Индию носителей индоевропейского языка». [ 19 ] [ примечание 2 ]
Возможное происхождение ямной или шнуровой керамики
[ редактировать ]

Дэвид Энтони считает ямную культуру урхейматом индоевропейским . [ 20 ] [ 21 ] По данным Хаака и др. (2015) , произошла массовая миграция ямной культуры на север ок. 2500 г. до н.э., что составляет 75% генетического происхождения культуры шнуровой керамики , при этом отмечается, что R1a и R1b могли «распространиться в Европу с Востока после 3000 г. до н.э.». [ 22 ] Однако все семь образцов Ямной принадлежали к субкладу R1b-M269 , [ 22 ] но в их образцах из Ямной не обнаружено R1a1a. Возникает вопрос, откуда взялась R1a1a в культуре шнуровой керамики, если не из ямной культуры. [ 23 ]
По мнению Марка Хабера, отсутствие гаплогруппы R1a-M458 в Афганистане не подтверждает происхождение линий R1a в современных популяциях Центральной Азии из Понтийско-Каспийской степи. [ 24 ]
По мнению Лео Клейна , отсутствие гаплогруппы R1a в остатках Ямной (несмотря на ее присутствие в энеолитической Самаре и популяциях восточных охотников-собирателей ) делает маловероятным, что европейцы унаследовали гаплогруппу R1a от Ямной. [ 25 ]
Археолог Барри Канлифф сказал, что отсутствие гаплогруппы R1a в образцах из Ямной является основным недостатком предположения Хаака о том, что R1a имеет ямное происхождение. [ 26 ]
Семенов и Булат (2016) действительно приводят доводы в пользу ямного происхождения R1a1a в культуре шнуровой керамики, отмечая, что несколько публикаций указывают на присутствие R1a1 в культуре гребенчатой керамики . [ 27 ] [ примечание 3 ]
Предполагаемое южноазиатское происхождение
[ редактировать ]Кивисилд и др. (2003) предложили либо Южную, либо Западную Азию , [ 28 ] [ примечание 4 ] в то время как Мирабал и др. (2009) видят поддержку как Южной, так и Центральной Азии. [ 10 ] Сенгупта и др. (2006) предположили индийское происхождение. [ 29 ] Тансим и др. (2006) предложили либо Южную, либо Центральную Азию. [ 30 ] Саху и др. (2006) предложили либо Южную, либо Западную Азию. [ 31 ] Тангарадж и др. (2010) также предположили южноазиатское происхождение. [ 32 ] Шарма и др. (2009) предполагают существование R1a в Индии от 18 000 до, возможно, 44 000 лет происхождения. [ 1 ]
Ряд исследований, проведенных с 2006 по 2010 год, пришли к выводу, что популяции Южной Азии имеют самое высокое разнообразие STR в пределах R1a1a, [ 33 ] [ 34 ] [ 10 ] [ 3 ] [ 1 ] [ 35 ] и последующие более старые датировки TMRCA . [ примечание 5 ] R1a1a присутствует как среди высших ( браминских ) каст, так и среди низших каст, и хотя частота выше среди каст браминов, самые старые датировки TMRCA гаплогруппы R1a происходят в племени Сахария , зарегистрированной касте региона Бунделькханд в Центральной Индии . [ 1 ] [ 35 ]
На основании этих результатов некоторые исследователи предположили, что R1a1a возник в Южной Азии. [ 34 ] [ 1 ] [ примечание 6 ] исключая более поздний, хотя и незначительный, генетический приток индоевропейских мигрантов в северо-западных регионах, таких как Афганистан, Белуджистан, Пенджаб и Кашмир. [ 34 ] [ 33 ] [ 3 ] [ примечание 7 ]
Вывод о том, что R1a возник в Индии, был поставлен под сомнение недавними исследованиями. [ 18 ] [ 37 ] [ примечание 8 ] предлагая доказательства того, что R1a прибыл в Индию с многочисленными волнами миграции. [ 18 ] [ 38 ]
Предполагаемое происхождение Закавказья и Западной Азии и возможное влияние на цивилизацию долины Инда
[ редактировать ]Хаак и др. (2015) обнаружили, что часть предков ямной культуры произошла с Ближнего Востока и что методы неолита, вероятно, пришли в ямную культуру с Балкан . [ примечание 9 ] Культура Рёссен (4600–4300 гг. до н.э.), которая располагалась в Германии и предшествовала культуре шнуровой керамики, старый субклад R1a, а именно L664, все еще можно найти. [ примечание 10 ]
Часть южноазиатского генетического происхождения происходит от популяций Западной Евразии, и некоторые исследователи предполагают, что Z93 мог прийти в Индию через Иран. [ 40 ] и расширился там во времена цивилизации долины Инда . [ 2 ] [ 41 ]
Маскареньяс и др. (2015) предположили, что корни Z93 лежат в Западной Азии, и предположили, что «Z93 и L342.2 распространились в юго-восточном направлении от Закавказья в Южную Азию », [ 40 ] отмечая, что такая экспансия совместима с «археологическими записями экспансии населения Западной Азии на восток в 4-м тысячелетии до нашей эры, кульминацией которой стали так называемые миграции Кура-Аракс в период после Урука IV ». [ 40 ] Тем не менее, Лазаридис отметил, что образец I1635 Lazaridis et al. (2016) , их армянский образец Кура-Араксес нес Y-гаплогруппу R1 b1 -M415(xM269); [ примечание 11 ] (также называемый R1b1a1b-CTS3187). [ 42 ] [ ненадежный источник? ]
По данным Андерхилла и др. (2014) диверсификация Z93 и «ранняя урбанизация в долине Инда… произошла [5600 лет назад] и географическое распространение R1a-M780 (рис. 3d [ примечание 12 ] ) может отражать это». [ 2 ] [ примечание 13 ] Позник и др. (2016) отмечают, что «поразительные расширения» произошли внутри R1a-Z93 ок. 4500–4000 лет назад, что «на несколько столетий предшествует краху цивилизации долины Инда». [ 41 ] [ примечание 14 ]
Однако, по данным Нарасимхана и др. (2018) , степные скотоводы являются вероятным источником R1a в Индии. [ 44 ] [ примечание 15 ]
Филогения
[ редактировать ]Генеалогическое древо R1a теперь имеет три основных уровня ветвления, с наибольшим количеством определенных субкладов в пределах доминирующей и самой известной ветви R1a1a (которая будет встречаться под разными названиями, такими как «R1a1», в относительно недавней, но не в последней литературе). .
Топология
[ редактировать ]Топология R1a следующая (коды [в скобках] не-isogg-коды): [ 9 ] [ 45 ] [ нужна проверка ] [ 46 ] [ 2 ] [ 47 ] Татьяна и др. (2014) «Процесс быстрой диверсификации K-M526 , вероятно, произошел в Юго-Восточной Азии с последующей экспансией на запад предков гаплогрупп R и Q ». [ 48 ]
- P P295/PF5866/S8 (также известный как K2b2 ).
- Р (Р-М207) [ 46 ] [ 9 ]
- Р*
- Р1 (Р-М173)
- Р1* [ 46 ]
- Р1а (М420) [ 46 ] (Восточная Европа, Азия) [ 2 ]
- Р1а* [ 9 ]
- R1a1 [ 46 ] (М459/PF6235, [ 46 ] SRY1532.2/SRY10831.2 [ 46 ] )
- Р1а1 (М459) [ 46 ] [ 9 ]
- Р1а1а (М17, М198) [ 46 ]
- R1a1a1 ( M417 , стр.7) [ 46 ]
- Р1а1а1а (CTS7083/L664/S298) [ 46 ]
- R1a1a1b (S224/Z645, S441/Z647) [ 46 ]
- R1a1a1b1 (PF6217/S339/Z283) [ 46 ]
- R1a1a1b1a ( Z282 ) [ 46 ] [R1a1a1a*] (Z282) [ 49 ] (Восточная Европа)
- R1a1a1b1a1 [ 46 ] [Старый топологический код — R1a1a1b*, он устарел и может привести к некоторой путанице.] [ 49 ] (М458) [ 46 ] [ 49 ] [R1a1a1g] (M458) [ 47 ]
- R1a1a1b1a2 [ 46 ] (С466/З280, С204/З91) [ 46 ]
- R1a1a1b1a2a [ 46 ]
- R1a1a1b1a2b (CTS1211) [ 46 ] [R1a1a1c*] (M558) [ 49 ] [R-CTS1211] (V2803/CTS3607/S3363/M558, CTS1211/S3357, Y34/FGC36457) [ 9 ]
- R1a1a1b1a2b3* (M417+, Z645+, Z283+, Z282+, Z280+, CTS1211+, CTS3402, Y33+, CTS3318+, Y2613+) (кластер K Гвоздзя) [ 45 ] [ нужна проверка ]
- R1a1a1b1a2b3a (L365/S468) [ 46 ]
- R1a1a1b1a3 (Z284) [ 46 ] [R1a1a1a1] (Z284) [ 49 ]
- R1a1a1b1a ( Z282 ) [ 46 ] [R1a1a1a*] (Z282) [ 49 ] (Восточная Европа)
- R1a1a1b2 (F992/S202/ Z93 ) [ 46 ] [R1a1a2*] (Z93, M746) [ 49 ] (Центральная Азия, Южная Азия и Западная Азия)
- R1a1a1b1 (PF6217/S339/Z283) [ 46 ]
- [R1a1a1c] (M64.2, M87, M204) [ 47 ]
- [R1a1a1d] (P98) [ 47 ]
- [R1a1a1d2a] [ 50 ]
- [R1a1a1e] (ПК5) [ 47 ]
- R1a1a1 ( M417 , стр.7) [ 46 ]
- R1b (M343) (Западная Европа)
- R2 (Индия)
Гаплогруппа R
[ редактировать ]| Филогения гаплогруппы R |
Р-М173 (Р1)
[ редактировать ]R1a отличается несколькими уникальными маркерами, включая мутацию M420. Это субклад гаплогруппы R-M173 (ранее называвшейся R1). R1a имеет сестринские субклады гаплогруппы R1b -M343 и парагруппу R-M173*.
Р-М420 (Р1а)
[ редактировать ]R-M420, определяемый мутацией M420, имеет две ветви: R-SRY1532.2, определяемый мутацией SRY1532.2, которая составляет подавляющее большинство; и R-M420*, парагруппа , определенная как M420-положительная, но SRY1532.2-отрицательная. (В схеме 2002 года это отрицательное меньшинство SRY1532.2 входило в состав относительно редкой группы, классифицированной как парагруппа R1*.) Мутации, которые считаются эквивалентными M420, включают M449, M511, M513, L62 и L63. [ 3 ] [ 51 ]
Лишь отдельные образцы новой парагруппы R-M420* были обнаружены Андерхиллом в 2009 году, в основном на Ближнем Востоке и на Кавказе : 1/121 оманец , 2/150 иранцев , 1/164 в Объединенных Арабских Эмиратах и 3/612 в Турции. . Тестирование еще 7224 мужчин из 73 других евразийских популяций не выявило признаков этой категории. [ 3 ]
Эта парагруппа теперь известна как R1a2 (R-YP4141). Затем у него есть две ветви R1a2a (R-YP5018) и R1a2b (R-YP4132).
Р-SRY1532.2 (R1a1)
[ редактировать ]R1a1 определяется SRY1532.2 или SRY10831.2 (подразумевается, что он всегда включает SRY10831.2, M448, L122, M459 и M516). [ 3 ] [ 52 ] ). В этом семействе линий преобладают M17 и M198. Напротив, в парагруппе R-SRY1532.2* отсутствуют маркеры M17 или M198.
Парагруппа R-SRY1532.2*, по-видимому, менее редка, чем R1*, но все же относительно необычна, хотя она была проверена более чем в одном исследовании. Андерхилл и др. (2009) сообщили о 1/51 в Норвегии , 3/305 в Швеции , 1/57 греко-македонцах , 1/150 иранцах, 2/734 этнических армянах и 1/141 кабардинце . [ 3 ] Саху и др. (2006) сообщили о R-SRY1532.2* для 1/15 образцов штата Химачал-Прадеш- Раджпут. [ 34 ]
Р-М17/М198 (Р1а1а)
[ редактировать ]Следующие SNP связаны с R1a1a:
| СНП | Мутация | Y-положение (NCBI36) | Y-позиция (ГРЧ37) | Идентификатор RefSNP |
|---|---|---|---|---|
| М17 | ИНС Г | 20192556 | 21733168 | rs3908 |
| М198 | С->Т | 13540146 | 15030752 | rs2020857 |
| М512 | С->Т | 14824547 | 16315153 | rs17222146 |
| М514 | С->Т | 17884688 | 19375294 | rs17315926 |
| М515 | Т->А | 12564623 | 14054623 | rs17221601 |
| Л168 | А->Г | 14711571 | 16202177 | - |
| Л449 | С->Т | 21376144 | 22966756 | - |
| Л457 | Г->А | 14946266 | 16436872 | rs113195541 |
| Л566 | С->Т | - | - | - |
Р-М417 (Р1а1а1)
[ редактировать ]R1a1a1 (R-M417) является наиболее широко встречающимся субкладом, в двух вариациях, которые встречаются соответственно в Европе (R1a1a1b1 (R-Z282) ([R1a1a1a*] (R-Z282) (Underhill 2014) [ 2 ] ) и Центральной и Южной Азии (R1a1a1b2 (R-Z93) ([R1a1a2*] (R-Z93) Underhill 2014) [ 2 ] ).
R-Z282 (R1a1a1b1a) (Восточная Европа)
[ редактировать ]Этот большой субклад, по-видимому, охватывает большую часть R1a1a, обнаруженного в Европе. [ 14 ]
- R1a1a1b1a [R1a1a1a* (Underhill (2014))] (R-Z282*) встречается на севере Украины, в Беларуси и России с частотой c. 20%. [ 2 ]
- R1a1a1b1a3 [R1a1a1a1 (Underhill (2014))] (R-Z284) встречается в Северо-Западной Европе и достигает максимума при c. 20% в Норвегии. [ 2 ]
- R1a1a1c (M64.2, M87, M204), по-видимому, редок: он был обнаружен у 1 из 117 мужчин, типированных на юге Ирана. [ 53 ]
Р-М458 (Р1а1а1б1а1)
[ редактировать ]
R-M458 представляет собой преимущественно славянский SNP, характеризующийся собственной мутацией и впервые названный кластером N. Underhill et al. (2009) обнаружили, что он присутствует в современных европейских популяциях примерно между водосбором Рейна и Уральскими горами , и связали это с «эффектом основателя, который ... приходится на период раннего голоцена, 7,9 ± 2,6 тысяч лет назад». (Животовские скорости, завышены в 3 раза) [ 3 ] M458 был найден в одном скелете из могильного поля XIV века в Узедоме , Мекленбург-Передняя Померания, Германия. [ 54 ] Статья Underhill et al. (2009) также сообщает об удивительно высокой частоте M458 в некоторых популяциях Северного Кавказа (18% среди акногайцев , [ 55 ] 7,8% среди караногайцев и 3,4% среди абазин ). [ 56 ]
Р-Л260 (Р1а1а1б1а1а)
[ редактировать ]R1a1a1b1a1a (R-L260), обычно называемый западнославянским или польским , является субкладом более крупной родительской группы R-M458 и впервые был идентифицирован как кластер STR Pawlowski et al. 2002 . В 2010 году было подтверждено, что это гаплогруппа, идентифицированная по собственной мутации (SNP). [ 57 ] По-видимому, он составляет около 8% польских мужчин, что делает его наиболее распространенным субкладом в Польше. За пределами Польши это менее распространено. [ 58 ] Помимо Польши, он в основном встречается в Чехии и Словакии и считается «явно западнославянским». По оценкам, предок-основатель R-L260 жил между 2000 и 3000 лет назад, то есть во время железного века , со значительным ростом населения менее 1500 лет назад. [ 59 ]
Р-М334
[ редактировать ]Р-М334 ([R1a1a1g1], [ 47 ] субклад [R1a1a1g] (M458) [ 47 ] cq R1a1a1b1a1 (M458) [ 46 ] ) был обнаружен Underhill et al. (2009) только у одного эстонца и может определять совсем недавно основанную небольшую кладу. [ 3 ]
R1a1a1b1a2 (S466/Z280, S204/Z91)
[ редактировать ]R1a1a1b1a2b3* (кластер К Гвоздзя)
[ редактировать ]R1a1a1b1a2b3* (M417+, Z645+, Z283+, Z282+, Z280+, CTS1211+, CTS3402, Y33+, CTS3318+, Y2613+) (кластер K Гвоздзя) [ 45 ] [ нужна проверка ] представляет собой группу на основе STR, то есть R-M17(xM458). Этот кластер распространен в Польше, но не только в Польше. [ 59 ]
R1a1a1b1a2b3a (R-L365)
[ редактировать ]R1a1a1b1a2b3a (R-L365) [ 46 ] ранее назывался кластером G. [ нужна ссылка ]
R1a1a1b2 (R-Z93) (Азия)
[ редактировать ]| Область | Люди | Н | Р-М17 | Р-М434 | ||
|---|---|---|---|---|---|---|
| Число | Частота. (%) | Число | Частота. (%) | |||
| Пакистан | Белудж | 60 | 9 | 15% | 5 | 8% |
| Пакистан | Макрани | 60 | 15 | 25% | 4 | 7% |
| Средний Восток | Мой собственный | 121 | 11 | 9% | 3 | 2.5% |
| Пакистан | Синдхи | 134 | 65 | 49% | 2 | 1.5% |
| В таблице показаны только положительные наборы из N = 3667, полученные из выборки 60 популяций Евразии. [ 3 ] | ||||||
Этот большой субклад, по-видимому, охватывает большую часть R1a1a, обнаруженных в Азии, и связан с индоевропейскими миграциями (включая скифов , индоарийских миграций и так далее). [ 14 ]
- R-Z93* или R1a1a1b2* (R1a1a2* в Underhill (2014)) наиболее распространен (>30%) в Южно-Сибирском Алтайском регионе России, встречается в Кыргызстане (6%) и во всех иранских популяциях (1-8). %). [ 2 ]
- R-Z2125 наиболее часто встречается в Кыргызстане и у афганских пуштунов (>40%). С частотой >10% он также наблюдается у других афганских этнических групп и у некоторых популяций на Кавказе и в Иране. [ 2 ]
- R-M560 очень редок и наблюдался только в четырех образцах: двух говорящих на бурушаски (северный Пакистан), одном хазарейском (Афганистан) и одном иранском азербайджанце. [ 2 ]
- R-M780 часто встречается в Южной Азии: Индии, Пакистане, Афганистане и Гималаях. В Турции доля сублиний R1a (12,1%). [ 60 ] Цыгане из Словакии имеют 3% в R1a [ 61 ] Эта группа также встречается в >3% в некоторых иранских популяциях и присутствует в >30% среди цыган из Хорватии и Венгрии. [ 2 ]
Географическое распространение R1a1a
[ редактировать ]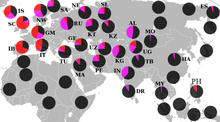
Доисторический
[ редактировать ]В мезолитической Европе R1a характерен для восточных охотников-собирателей (EHG). [ 62 ] Мужской ЭХГ веретской культуры, захороненный в Песчанице возле озера Лача в Архангельской области , Россия ок. 10700 г. до н.э. было обнаружено, что он является носителем отцовской гаплогруппы R1a5-YP1301 и материнской гаплогруппы U4a . [ 63 ] [ 64 ] [ 62 ] Было обнаружено , что самец по имени PES001 из Песчаницы на северо-западе России является носителем R1a5 и датируется по меньшей мере 10 600 лет назад. [ 5 ] Дополнительные примеры включают самцов Минино II (V) и Минино II (I/1), причем первый несет R1a1, а второй R1a соответственно, причем возраст первого составляет 10 600 лет, а второго - не менее 10 400 лет соответственно, оба из Минино на северо-западе России. [ 65 ] Мезолитический мужчина из Карелии ок. Было обнаружено, что период с 8800 по 7950 год до н.э. несет гаплогруппу R1a. [ 66 ] Мужчина эпохи мезолита, похороненный в Дереевке ок. С 7000 г. до н. э. по 6700 г. до н. э. несли отцовскую гаплогруппу R1a и материнскую гаплогруппу U5a2a . [ 17 ] Еще один самец из Карелии из ок. 5500–5000 гг. до н. э., считавшийся ЭХГ, носил гаплогруппу R1a. [ 15 ] Мужчина из культуры гребенчатой керамики в Кудрукюла ок. Было установлено, что в период с 5900 г. до н.э. по 3800 г. до н.э. он был носителем R1a и материнского U2e1 . [ 67 ] По словам археолога Дэвида Энтони, отцовский R1a-Z93 был найден на реке Оскол возле уже не существующего колхоза «Александрия», Украина c. 4000 г. до н. э., «самый ранний известный образец, демонстрирующий генетическую адаптацию к персистенции лактазы (13910-T)». [ 68 ] R1a был обнаружен в культуре шнуровой керамики . [ 69 ] [ 70 ] в котором оно преобладает. [ 71 ] эпохи бронзы Исследованные мужчины фатьяновской культуры полностью относятся к R1a, а именно к субкладу R1a-Z93. [ 62 ] [ 63 ] [ 72 ]
Гаплогруппа R1a позже была обнаружена в древних окаменелостях, связанных с культурой полей урн ; [ 73 ] а также захоронение останков Синташты , [ 16 ] Андроново , [ 74 ] Пазырык Pazyryk [ 75 ] Хэштеги , [ 74 ] Таштык , [ 74 ] и срубная культуры, жители древнего Танаиса , [ 76 ] в Таримских мумиях , [ 77 ] и аристократия хунну . [ 78 ] Скелетные останки отца и двух его сыновей из археологического памятника, обнаруженного в 2005 году недалеко от Эйлау (в Саксонии-Анхальт , Германия ) и датированного примерно 2600 годом до нашей эры, дали положительный результат на маркер Y-SNP SRY10831.2. Номер Ysearch останков Юлау — 2C46S. Таким образом, древняя клада присутствовала в Европе по крайней мере 4600 лет назад в связи с одним местом широко распространенной культуры шнуровой керамики . [ 69 ]
Европа
[ редактировать ]В Европе субклад R1a1 встречается на самом высоком уровне среди народов центрально- и восточноевропейского происхождения, с результатами от 35% до 65% среди чехов , венгров , поляков , словаков , западных украинцев , русинов , белорусов , молдаван и Россияне . [ 79 ] [ 80 ] [ 12 ] В странах Балтии частота R1a1a снижается от Литвы (45%) до Эстонии (около 30%). [ 81 ] [ 82 ] [ 83 ] [ 12 ] [ 84 ]
Значительное присутствие наблюдается у людей скандинавского происхождения, с самыми высокими уровнями в Норвегии и Исландии , где от 20 до 30% мужчин имеют R1a1a. [ 85 ] [ 86 ] Викинги и норманны, возможно, также распространили линию R1a1a дальше, составляя, по крайней мере, часть небольшого присутствия на Британских островах , Канарских островах и Сицилии . [ 87 ] [ 88 ] В Восточной Германии гаплогруппа R1a1a составляет в среднем от 20 до 30%, с пиком в Ростоке на уровне 31,3%. [ 89 ]
В Южной Европе R1a1a не распространен, но значительные уровни были обнаружены в отдельных районах, например, в долине Пас на севере Испании , в районах Венеции и Калабрии в Италии . [ 90 ] [ нужен лучший источник ] наблюдаются На Балканах большие различия между регионами со значительными уровнями R1a1a, например, 36–39% в Словении , [ 91 ] 27–34% в Хорватии , [ 82 ] [ 92 ] [ 93 ] [ 94 ] [ 95 ] и более 30% в греческой Македонии , но менее 10% в Албании , Косово и некоторых частях Греции к югу от ущелья Олимп. [ 96 ] [ 83 ] [ 12 ]
R1a практически состоит только из субклада Z284 в Скандинавии . В Словении основным субкладом является Z282 (Z280 и M458), хотя субклад Z284 был обнаружен в одном образце словенца. представленность Z93 незначительна В Турции , 12,1%. [ 60 ] [ 2 ] Для западных славян и венгров характерна высокая частота субклада М458 и низкая Z92, субклада Z280. У сотен словенских образцов и чехов отсутствует субклад Z92 от Z280, в то время как у поляков, словаков, хорватов и венгров наблюдается только очень низкая частота Z92. [ 2 ] , У балтов восточных славян , сербов , македонцев , болгар и румын наблюдается соотношение Z280>М458 и высокая, вплоть до преобладающей доли Z92. [ 2 ] Балты и восточные славяне имеют одинаковые субклады и схожие частоты в более детальной филогении субкладов. [ 97 ] [ 98 ] Русский генетик Олег Балановский предположил, что в генетике восточных и западнославянских популяций существует преобладание ассимилированного дославянского субстрата. По его мнению, общая генетическая структура, отличающая восточных славян и балтов от других популяций, может служить объяснением того, что праславянский субстрат восточных и западных славян в значительной степени состоял из балтоязычных народов, которые, в какой-то момент предшествовали славянам в культурах евразийской степи . согласно археологическим и топонимическим данным, [ примечание 16 ]
Азия
[ редактировать ]Центральная Азия
[ редактировать ]Зерджал и др. ) обнаружили R1a1a у 64% выборки таджиков Таджикистана и 63 % выборки кыргызов Кыргызстана ( 2002 . [ 99 ]
Хабер и др. (2012) обнаружили R1a1a-M17 в 26,0% (53/204) выборки из Афганистана , включая 60% (3/5) выборки нуристанцев , 51,0% (25/49) выборки пуштунов, 30,4% (17/56) выборки таджиков, 17,6% (3/17) выборки узбеков, 6,7% (4/60) выборки хазарейцев и у единственного в выборке туркменского человека. [ 100 ]
Ди Кристофаро и др. (2013) обнаружили R1a1a-M198/M17 у 56,3% (49/87) парных выборок пуштунов из Афганистана (включая 20/34 или 58,8% выборки пуштунов из Баглана и 29/53 или 54,7% выборки пуштунов из Баглана). выборка пуштунов из Кундуза ), 29,1% (37/127) пула выборок узбеков из Афганистана (в том числе 28/94 или 29,8% выборки узбеков из Джаузджана , 8/28 или 28,6% выборки узбеков из Сари Пола и 1/5 или 20% выборки узбеков из Балха ), 27,5% (39/142) пула выборок таджиков из Афганистана (в том числе 22/54 или 40,7% выборки Таджики из Балха , 9/35 или 25,7% выборки таджиков из Тахара , 4/16 или 25,0% выборки таджиков из Самангана , и 4/37 или 10,8% выборки таджиков из Бадахшана ), 16,2% (12/74) выборки туркмен из Джаузджана , и 9,1% (7/77) пары выборок хазарейцев из Афганистана (в том числе 7/69 или 10,1% выборки хазарейцев из Бамиана и 0/8 или 0% выборки хазарейцев из Балха ). [ 101 ]
По данным Малярчука и др. (2013) обнаружили R1a1-SRY10831.2 у 30,0% (12/40) выборки таджиков из Таджикистана. [ 102 ]
Аширбеков и др. (2017) обнаружили R1a-M198 в 6,03% (78/1294) выборки казахов из Казахстана . R1a-M198 наблюдался с частотой выше средней в выборках исследования следующих казахских племен: 13/41 = 31,7% выборки Суан, 8/29 = 27,6% выборки Ошакты, 6/30 = 20,0% выборки Кожа, 4/29 = 13,8% выборки Кыпшак, 1/8 = 12,5% выборки Торе, 9/86 = 10,5% выборки Джетыру, 4/50 = 8,0% выборки проба Аргын, 1/13 = 7,7% выборки Шанышклы, 8/122 = 6,6% выборки Алимулы, 3/46 = 6,5% выборки Албан. R1a-M198 также наблюдался у 5/42 = 11,9% выборки казахов незарегистрированной племенной принадлежности. [ 103 ]
Южная Азия
[ редактировать ]В Южной Азии R1a1a часто наблюдался в ряде демографических групп. [ 34 ] [ 33 ]
В Индии высокие частоты этой гаплогруппы наблюдаются у западнобенгальских брахманов (72%). [ 33 ] на востоке, Гуджарат Лоханас (60%) на западе, [ 3 ] Пенджаб/Харьяна Хатрис (67%) и Ахирс (63%) на севере, [ 3 ] и Карнатака Медарс (39%) [ 104 ] на юге. Он также был обнаружен у нескольких южноиндийских дравидийском , говорящих на адиваси языке , включая ченчу (26%) в Андхра-Прадеше и кота в Андхра-Прадеше (22,58%). [ 105 ] и Каллар из Тамилнада, что позволяет предположить, что R1a1a широко распространен среди племенных южных индейцев. [ 28 ]
Помимо этого, исследования показывают высокий процент в различных по региону группах, таких как манипури (50%) [ 3 ] на крайний северо-восток и среди пенджабцев (47%) [ 28 ] до крайнего Северо-Запада.
В Пакистане он встречается на уровне 71% среди племени моханна в провинции Синд на юге и 46% среди балти в Гилгит-Балтистане на севере. [ 3 ] Среди сингальцев Шри -Ланки 23% оказались положительными на R1a1a (R-SRY1532). [ 106 ] У индусов района Читван в Тераи регионе , Непал, этот показатель составляет 69%. [ 107 ]
Восточная Азия
[ редактировать ]Частота R1a1a сравнительно низка среди некоторых тюркоязычных групп, таких как якуты , однако уровни выше (от 19 до 28%) в некоторых тюркоязычных или монголоязычных группах Северо-Западного Китая , таких как Бонань , Дунсян , Салар и Уйгуры . [ 13 ] [ 108 ] [ 109 ]
Китайская статья, опубликованная в 2018 году, обнаружила R1a-Z94 у 38,5% (15/39) выборки кериялик-уйгуров из деревни Дарья Бойи / деревня Дарья Бой, уезд Ютянь , Синьцзян (поселок Дарья Бой, уезд Ютянь), R1a-Z93 у 28,9. % (22/76) выборки уйгуров Долан из поселка Хорикол, уезда Ават , Синьцзяна (город Улукиле, уезд Ават) и R1a-Z93 в 6,3% (4/64) выборки уйгуров Лоплика из Каркуги/Карчуги Деревня Юлийского уезда Синьцзяна R1a(xZ93) наблюдалась только у одного из 76 доланских уйгуров. [ 110 ] Отметим, что деревня Дарья Бойи расположена в отдаленном оазисе, образованном рекой Керия в пустыне Такла-Макан . Исследование Y-ДНК 2011 года обнаружило Y-ДНК R1a1 у 10% выборки южных хуэйцев из Юньнани, 1,6% выборки тибетцев из Сизанга ( Тибетский автономный район ), 1,6% выборки сибэ из Синьцзяна. , 3,2% выборки северных хуэй из Нинся , 9,4% выборки хазаков ( казахов ) из Синьцзяна и показатели 24,0%, 22,2%, 35,2%, 29,2% в 4 различных выборках уйгуров из Синьцзяна, 9,1% в выборке монголов из Внутренней Монголии . Другой субклад R1 также был обнаружен в 1,5% выборки северного Хуэй из Нинся. [ 111 ] в том же исследовании вообще не было обнаружено случаев R1a в 6 образцах ханьцев в провинции Юньнань , 1 образце хань в Гуанси , 5 образцах хань в Гуйчжоу , 2 образцах хань в Гуандуне , 2 образцах хань в провинции Фуцзянь , 2 образца хань в Чжэцзяне , 1 образец хань в Шанхае , 1 образец хань в Цзянси , 2 образца хань в провинции Хунань , 1 образец хань в Хубэе , 2 образца хань в Сычуани , 1 образец хань в Чунцине , 3 образцы хань в Шаньдуне , 5 образцов хань в Ганьсу , 3 образца хань в Цзилине и 2 образца хань в Хэйлунцзяне . [ 112 ] 40% саларов, 45,2% таджиков Синьцзяна , 54,3% Дунсяна, 60,6% татар и 68,9% киргизов в Синьцзяне на северо-западе Китая, протестированных в одной выборке, имели R1a1-M17. Баоань (Бонань) имел наибольшее разнообразие гаплогрупп - 0,8946 ± 0,0305, в то время как другие этнические меньшинства на северо-западе Китая имели высокое разнообразие гаплогрупп, как и жители Центральной Азии, - 0,7602 ± 0,0546. [ 113 ]
В Восточной Сибири R1a1a встречается среди некоторых коренных этнических групп, включая камчатцев и чукотцев , и достигает максимума у ительмана - 22%. [ 114 ]
Юго-Восточная Азия
[ редактировать ]Y-гаплогруппы R1a-M420 и R2-M479 встречаются у народов Эде (8,3% и 4,2%) и Гиараи (3,7% и 3,7%) во Вьетнаме . дополнительно У чамов есть гаплогруппы R-M17 (13,6%) и R-M124 (3,4%).
R1a1a1b2a2a (R-Z2123) и R1a1 встречаются у кхмерских народов Таиланда (3,4%) и Камбоджи (7,2%) соответственно. Гаплогруппа R1a1a1b2a1b (R-Y6) встречается также у куйских народов (5%).
По данным Чангмая и др. al (2022), эти частоты гаплогрупп происходят от выходцев из Южной Азии, которые оставили культурное и генетическое наследие в Юго-Восточной Азии с первого тысячелетия нашей эры. [ 115 ]
Западная Азия
[ редактировать ]R1a1a был обнаружен в различных формах в большинстве частей Западной Азии , в самых разных концентрациях: от почти полного отсутствия в таких регионах, как Иордания , до гораздо более высоких уровней в некоторых частях Кувейта и Ирана . Бедуинское племя Шимар ( Шаммар ) в Кувейте показывает самую высокую частоту на Ближнем Востоке - 43%. [ 116 ] [ 117 ] [ 118 ]
Уэллс 2001 отметил, что в западной части страны у иранцев наблюдаются низкие уровни R1a1a, в то время как мужчины в восточных частях Ирана несут до 35% R1a1a. Насидзе и др. В 2004 году R1a1a был обнаружен примерно у 20% иранских мужчин из городов Тегеран и Исфахан . Регейро в 2006 году в исследовании Ирана отметил гораздо более высокую частоту на юге, чем на севере.
Новое исследование выявило 20,3% R-M17* среди курдских проб, взятых в провинции Курдистан на западе Ирана, 19% среди азербайджанцев в Западном Азербайджане , 9,7% среди мазандаранцев в Северном Иране в провинции Мазандаран , 9,4% среди гилаков. в провинции Гилян , 12,8% среди персов и 17,6% среди зороастрийцев в Йезде , 18,2% среди персов в Исфахане , 20,3% среди персов в Хорасане , 16,7% афроиранцев, 18,4% среди кешми "гешми", 21,4% среди персидских бандари в Хормозгане и 25% среди белуджей в провинции Систан и Белуджистан . [ 119 ]
Ди Кристофаро и др. (2013) обнаружили гаплогруппу R1a в 9,68% (18/186) набора выборок из Ирана, хотя и с большой дисперсией от 0% (0/18) в выборке иранцев из Тегерана до 25% (5/20 ) в выборке иранцев из Хорасана и 27% (3/11) в выборке иранцев неизвестного происхождения. Все иранцы R1a несли мутации M198 и M17, за исключением одного человека в выборке иранцев из Гилана ( n = 27), который, как сообщалось, принадлежал к R1a-SRY1532.2(xM198, M17). [ 101 ]
Малярчук и др. (2013) обнаружили R1a1-SRY10831.2 у 20,8% (16/77) выборки персов, собранной в провинциях Хорасан и Керман на востоке Ирана, но не нашли ни одного члена этой гаплогруппы в выборке из 25 курдов. собраны в провинции Керманшах на западе Ирана. [ 102 ]
С другой стороны, к северу от этих регионов Западной Азии уровни R1a1a начинают повышаться на Кавказе , опять же неравномерно. В нескольких изученных популяциях не было обнаружено никаких признаков R1a1a, в то время как самые высокие уровни, обнаруженные в регионе, по-видимому, принадлежат носителям карачаево -балкарского языка, среди которых около четверти протестированных мужчин имеют гаплогруппу R1a1a. [ 3 ]
Историческое название R1a
[ редактировать ]Историческая система именования, обычно используемая для R1a, была противоречивой в разных опубликованных источниках, поскольку она часто менялась; это требует некоторого объяснения.
В 2002 году Консорциум Y-хромосомы (YCC) предложил новую систему наименования гаплогрупп ( YCC 2002 ), которая теперь стала стандартной. В этой системе имена в формате «R1» и «R1a» являются « филогенетическим » именем, предназначенным для обозначения позиций в генеалогическом древе. Названия мутаций SNP также можно использовать для обозначения клад или гаплогрупп. Например, поскольку M173 в настоящее время является определяющей мутацией R1, R1 также является R-M173, «мутационным» названием клады. Когда обнаруживается новое ветвление в дереве, некоторые филогенетические названия изменятся, но по определению все мутационные названия останутся прежними.
Широко распространенная гаплогруппа, определенная мутацией M17, была известна под разными названиями, например «Eu19», как это использовалось в ( Semino et al. 2000 ) в старых системах именования. Предложение YCC 2002 года присвоило имя R1a гаплогруппе, определенной мутацией SRY1532.2. Сюда входил Eu19 (т.е. R-M17) в качестве субклада, поэтому Eu19 был назван R1a1. Обратите внимание: SRY1532.2 также известен как SRY10831.2. [ нужна ссылка ] Открытие M420 в 2009 году привело к изменению этих филогенетических названий. ( Underhill et al. 2009 и ISOGG 2012 ) R1a теперь определяется мутацией M420: в этом обновленном дереве субклад, определенный SRY1532.2, переместился из R1a. к R1a1 и Eu19 (R-M17) от R1a1 до R1a1a.
Более поздние обновления, записанные на справочной веб-странице ISOGG, касаются ветвей R-M17, включая одну основную ветку, R-M417.
| Схема 2002 г., предложенная в ( YCC 2002 ). | Схема 2009 г. согласно ( Underhill et al. 2009 ) | Дерево ISOGG по состоянию на январь 2011 г. [ нужна ссылка ] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
См. также
[ редактировать ]Субклады Y-ДНК R-M207
[ редактировать ]Основополагающее дерево Y-ДНК
[ редактировать ]Примечания
[ редактировать ]- ^ Согласно Семейному древу, [ ВОЗ? ] они диверсифицировали c. 5000 лет назад. [ 9 ]
- ^ См. также: « Рассредоточение населения по Индийскому субконтиненту с «сильной гендерной предвзятостью» (Сильва и др., 2017)» . Блог Еврогенеса . 28 марта 2017 г. [ самостоятельно опубликованный источник? ]
- ^ Семенов и Булат (2016) ссылаются на следующие публикации:
- Хаак, Вольфганг (2015). «Массовая миграция из степи является источником индоевропейских языков в Европе» . Природа . 522 (7555): 207–211. arXiv : 1502.02783 . Бибкод : 2015Natur.522..207H . биоRxiv 10.1101/013433 . дои : 10.1038/NATURE14317 . ПМК 5048219 . ПМИД 25731166 .
- Мэтисон, Иэн (2015). «Восемь тысяч лет естественного отбора в Европе». биоRxiv 10.1101/016477 .
- Чекунова Е.М., Ярцева Н.В., Чекунов М.К., Мазуркевич А.Н. Первые результаты генотипирования аборигенов и человеческих костных остатков археологических памятников Верхнего Подвинья. // Археология озерных поселений IV—II тысячелетий до нашей эры: Хронология культур и природной среды и климатических ритмов. Материалы международной конференции, посвященной 50-летнему исследованию свайных поселений Северо-Запада России. Санкт-Петербург, 13–15 ноября 2014 г.
- Джонс, скорая помощь; Гонсалес-Фортес, Г; Коннелл, С; Сиска, В; Эрикссон, А; Мартиниано, Р; Маклафлин, РЛ; Гальего Льоренте, М; Кэссиди, LM; Гамба, К; Мешвелиани, Т; Бар-Йосеф, О; Мюллер, В; Белфер-Коэн, А; Мацкевич З.; Джейкли, Н; Хайэм, штат Техас; Куррат, М; Лордкипанидзе, Д; Хофрейтер, М; Маника, А; Пинхаси, Р; Брэдли, генеральный директор (2015). «Геномы верхнего палеолита раскрывают глубокие корни современных евразийцев» . Нат Коммун . 6 : 8912. Бибкод : 2015NatCo...6.8912J . дои : 10.1038/ncomms9912 . ПМК 4660371 . ПМИД 26567969 .
- ^ Кивисилд и др. (2003): «Гаплогруппа R1a, ранее связанная с предполагаемым индоарийским вторжением, была обнаружена с самой высокой частотой в Пенджабе, но также с относительно высокой частотой (26%) в племени Ченчу. Это открытие вместе с более высоким R1a Связанное с этим разнообразие коротких тандемных повторов в Индии и Иране по сравнению с Европой и Центральной Азией позволяет предположить, что источником этой гаплогруппы могла быть Южная и Западная Азия». [ 28 ]
- ^ Сенгупта (2006) : «Мы обнаружили, что влияние Центральной Азии на ранее существовавший генофонд было незначительным. Возраст накопленных микросателлитных вариаций в большинстве индийских гаплогрупп превышает 10 000–15 000 лет, что свидетельствует о древности региональных Таким образом, наши данные не поддерживают модели, которые ссылаются на явно выраженный недавний генетический вклад из Центральной Азии для объяснения наблюдаемых генетических вариаций в Южной Азии».
- ^ Южноазиатское происхождение:
* Саху и др. (2006) : «…следует ожидать наблюдения резко меньшей генетической изменчивости среди индийских линий Rla. На самом деле всё наоборот: разнообразие гаплотипов STR на фоне R1a в Центральной Азии (а также в Восточной Европе) уже было показано, что она ниже, чем в Индии (6). Скорее, высокая распространенность R1* и Rla среди популяций Центральной Азии и Европы (без R2 и R* в большинстве случаев) более экономно объясняется потоком генов в противоположном направлении. , возможно, с ранним эффектом основателя в Южной или Западной Азии. [ 36 ]
* Шарма и др. (2009) : «Своеобразное наблюдение самой высокой частоты (до 72,22%) Y-гаплогруппы R1a1* у брахманов намекало на ее присутствие в качестве линии-основателя этой кастовой группы. Кроме того, наблюдение R1a1* в различных племенных группах населения , существование Y-гаплогруппы R1a* у предков и расширенный филогенетический анализ объединенного набора данных 530 индийцев, 224 пакистанцев и 276 выходцев из Центральной Азии и Евразии, несущих гаплогруппу R1a1*, подтвердили автохтонное происхождение линии R1a1 в Индии и племенную связь с индийцами. Однако важно обнаружить новый бинарный маркер(ы) Y-хромосомы для более высокого разрешения R1a1* и подтвердить сделанные выводы». - ^ Хотя Сенгупта (2006) признал, что «[R1a1 и R2] могли действительно прибыть в южную Индию из исходного региона на юго-западе Азии несколько раз». Полностью: «Широкое географическое распространение HG R1a1-M17 по всей Евразии и отсутствие в настоящее время информативных подразделений, определяемых бинарными маркерами, оставляют неясным географическое происхождение HG R1a1-M17. Однако контурная карта дисперсии R1a1-M17 показывает самый высокий вариация в северо-западном регионе Индии... Остается вопрос, насколько различна история L1 по отношению к некоторым или всем представителям R1a1 и R2. Эта неопределенность нейтрализует предыдущие выводы о вторжении HGs R1a1 и R2 с северо-запада в дравидский период. -говорящие южные племена связаны с одним недавним событием [R1a1 и R2], которое могло фактически прибыть в южную Индию из исходного региона юго-западной Азии несколько раз, причем некоторые эпизоды существуют значительно раньше, чем другие. Существуют значительные археологические свидетельства присутствия мезолита. народы Индии (Кеннеди, 2000), некоторые из которых могли проникнуть на субконтинент с северо-запада в эпоху позднего плейстоцена. Высокая дисперсия R1a1 в Индии (таблица 12), пространственное распределение частот микросателлитных дисперсионных клинов R1a1 (рис. 4) и время расширения (таблица 11) подтверждают эту точку зрения». [ 33 ]
- ^ Лалуэса-Фокс: «Несколько лет назад местные ученые поддержали точку зрения, что существование Y-хромосомы R1a не связано с потоком чужеродных генов, а вместо этого связано с тем, что эта линия возникла на субконтиненте и распространилась оттуда. Но филогенетическая реконструкция этой гаплогруппы не подтвердили эту точку зрения». [ 37 ]
- ^ Тем не менее, Хаак и др. также прямо заявляют: «тип ближневосточного происхождения, отличный от того, который был привнесен ранними фермерами». [ нужны разъяснения ] [ 39 ]
- ^ Согласно ДНК Генеалогического древа, L664 образовался 4700 лн, то есть 2700 г. до н.э. [ 9 ]
- ↑ Лазаридис, Твиттер, 18 июня 2016 г .: «I1635 (Armenia_EBA) — это R1b1-M415(xM269). Мы обязательно включим в редакцию. Спасибо человеку, который заметил! #ILovePreprints». [ ненадежный источник? ]
См. также «Большое дело 2016 года: территория современного Ирана не может быть индоевропейской родиной» . Блог Еврогенеса . 26 ноября 2016 г., [ ненадежный источник? ] для обсуждения той же темы. - ^ См. карту распространения M780 в блоге Dieneke's Anthropology, основная новая статья о глубоком происхождении Y-гаплогруппы R1a (Underhill et al. 2014). [ 43 ]
- ↑ Согласно генеалогическому древу ДНК, M780 образовался 4700 лн. [ 9 ] Эта датировка совпадает с перемещением на восток между 2800 и 2600 гг. до н. э. ямной культуры в область полтавской культуры , предшественницы синташтинской культуры , от которой произошли индоиранцы. М780 сосредоточен в долине Ганга, месте проживания классического ведического общества .
- ^ Позник и др. (2016) рассчитывают со временем генерации 30 лет; время генерации в 20 лет дает другие результаты.
- ^ «Доказательства того, что кластер Steppe_MLBA [средний и поздний бронзовый век] является вероятным источником степного происхождения в Южной Азии, также подтверждается данными Y-хромосомы, поскольку гаплогруппа R1a относится к подтипу Z93, распространенному сегодня в Южной Азии [Андерхилл et al. (2014), Silva et al. (2017)] была высокой частотой в Steppe_MLBA (68%) (16), но редко в Steppe_EMBA [от раннего до среднего бронзового века] (отсутствует в наших данных)». [ 44 ]
- ^ Балановский (2015) , p. 208 (in Russian) Прежде всего, это преобладание в славянских популяциях дославянского субстрата — двух ассимилированных ими генетических компонентов – восточноевропейского для западных и восточных славян и южноевропейского для южных славян...Можно с осторожностью предположить, что ассимилированный субстратмог быть представлен по преимуществу балтоязычными популяциями. Действительно, археологические данные указыва ют на очень широкое распространение балтских групп перед началом расселения славян. Балтскийсубстрату славян (правда, наряду с финно-угорским) выявляли и антропологи. Полученные нами генетические данные — и на графиках генетических взаимоотношений, и по доле общих фрагментов генома — указывают, что современные балтские народы являются ближайшими генетически ми соседями восточных славян. При этом балты являются и лингвистически ближайшими род ственниками славян. И можно полагать, что к моменту ассимиляции их генофонд не так сильно отличался от генофонда начавших свое широкое расселение славян. Поэтому если предположить,что расселяющиеся на восток славяне ассимилировали по преимуществу балтов, это может объяснить и сходство современных славянских и балтских народов друг с другом, и их отличия от окружающих их не балто-славянских групп Европы...В работе высказывается осторожное предположение, что ассимилированный субстрат мог быть представлен по преимуществу балтоязычными популяциями. Действительно, археологические данные указывают на очень широкое распространение балтских групп перед началом расселения славян. Балтский субстрат у славян (правда, наряду с финно-угорским) выявляли и антропологи. Полученные в этой работе генетические данные — и на графиках генетических взаимоотношений, и по доле общих фрагментов генома — указывают, что современные балтские народы являются ближайшими генетическими соседями восточных славян.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и Шарма и др. 2009 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в Андерхилл и др. 2014 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р Андерхилл и др. 2009 .
- ^ Перейти обратно: а б Андерхилл и др. 2014 , с. 130.
- ^ Перейти обратно: а б Сааг, Лехти; Васильев Сергей Владимирович; Варул, Лийви; Косорукова Наталья Владимировна; Герасимов Дмитрий В.; Ошибкина Светлана Владимировна; Гриффит, Сэмюэл Дж.; Сольник, Ану; Сааг, Лаури; Д'Атанасио, Евгения; Мецпалу, Эне (январь 2021 г.). «Изменения генетического происхождения при переходе от каменного века к бронзовому веку на восточноевропейской равнине» . Достижения науки . 7 (4): eabd6535. Бибкод : 2021SciA....7.6535S . дои : 10.1126/sciadv.abd6535 . ПМЦ 7817100 . ПМИД 33523926 .
- ^ Хаак, Вольфганг; Лазаридис, Иосиф; Паттерсон, Ник; Роланд, Надин; Маллик, Свапан; Пламя, Бастьен; Брандт, Гвидо; Норденфилд, Сюзанна; Харни, Эдаоин; Стюардсон, Кристина; Фу, Цяомэй (10 февраля 2015 г.). «Массовая миграция из степей — источник индоевропейских языков в Европе » bioRxiv : 013433. arXiv : 1502.02783 . дои : 10.1101/013433 . S2CID 196643946 . Архивировано из оригинала 23 декабря . Получено 8 февраля ,
- ^ Рагхаван, Маанаса; Скоглунд, Понт; Граф, Келли Э.; Мецпалу, Мейт; Альбрехцен, Андерс; Мольтке, Ида; Расмуссен, Саймон; Стаффорд-младший, Томас В.; Орландо, Людовик; Мецпалу, Эне; Кармин, Моника (январь 2014 г.). «Геном верхнепалеолита Сибири свидетельствует о двойном происхождении коренных американцев» . Природа . 505 (7481): 87–91. Бибкод : 2014Natur.505...87R . дои : 10.1038/nature12736 . ПМК 4105016 . ПМИД 24256729 .
- ^ Нарасимхан, Вагиш М.; Паттерсон, Ник; Мурджани, Прия; Роланд, Надин; Бернар, Ребекка; Маллик, Свапан; Лазаридис, Иосиф; Накацука, Натан; Олальде, Иниго; Липсон, Марк; Ким, Александр М. (6 сентября 2019 г.). «Формирование человеческих популяций в Южной и Центральной Азии » Наука 365 (6457): eaat7487. дои : 10.1126/science.aat7487 . ПМЦ 6822619 . ПМИД 31488661 .
Типы гаплогрупп Y-хромосомы R1b или R1a в этот период не представлены в Иране и Туране.
- ^ Перейти обратно: а б с д и ж г час я дж «Дерево R1a» . YПолный . Архивировано из оригинала 19 августа 2016 года . Проверено 15 июля 2016 г.
- ^ Перейти обратно: а б с Мирабал и др. 2009 .
- ^ Зержал, Т.; и др. (1999). «Использование вариаций Y-хромосомной ДНК для изучения истории населения: недавнее распространение самцов в Азии и Европе». В Папихе, СС; Дека Р. и Чакраборти Р. (ред.). Геномное разнообразие: применение в популяционной генетике человека . Нью-Йорк: Издательство Kluwer Academic/Plenum. стр. 91–101. ISBN 978-0-3064-6295-5 .
- ^ Перейти обратно: а б с д Семино и др. 2000 .
- ^ Перейти обратно: а б Уэллс 2001 .
- ^ Перейти обратно: а б с Памджав и др. 2012 .
- ^ Перейти обратно: а б Хаак и др. 2015 .
- ^ Перейти обратно: а б Allentoft et al. 2015Аллентофт и др. 2015
- ^ Перейти обратно: а б Мэтисон и др. 2015 .
- ^ Перейти обратно: а б с Сильва и др. 2017 .
- ^ Джозеф, Тони (16 июня 2017 г.). «Как генетика разрешает дебаты об арийской миграции» . Индус . Архивировано из оригинала 4 октября 2023 года . Проверено 2 июня 2019 г.
- ^ Энтони 2007 .
- ^ Энтони и Ринге, 2015 .
- ^ Перейти обратно: а б Хаак и др. 2015 , с. 5.
- ^ Semenov & Bulat 2016 .
- ^ Хабер и др. 2012 «R1a1a7-M458 отсутствовал в Афганистане, что позволяет предположить, что R1a1a-M17 не поддерживает, как считалось ранее [47], экспансию из Понтийской степи [3], приносящую индоевропейские языки в Среднюю Азию и Индию».
- ^ Клейн, Лео С. (22 апреля 2017 г.). «Степная гипотеза индоевропейского происхождения еще не доказана» . Акта Археологика . 88 (1): 193–204. дои : 10.1111/j.1600-0390.2017.12184.x . ISSN 0065-101X . Архивировано из оригинала 25 декабря 2022 года . Проверено 23 ноября 2022 г. «Что касается Y-хромосомы, то уже отмечалось в Хааке, Лазаридисе и др. (2015), что ямная из Самары имела Y-хромосомы, которые принадлежали R-M269, но не принадлежали к кладе, распространенной в Западной Европе (p .46 приложения). Кроме того, в Ямной нет ни одного R1a, в отличие от шнуровой керамики (с преобладанием R1a)».
- ^ Кох, Джон Т.; Канлифф, Барри (2016). Кельтский с Запада 3: Атлантическая Европа в эпоху металла . Книги Оксбоу. п. 634. ИСБН 978-1-78570-228-0 . Архивировано из оригинала 23 ноября 2022 года . Проверено 23 ноября 2022 г.
- ^ Semenov & Bulat 2016 , p. 41.
- ^ Перейти обратно: а б с д Кивисилд и др. 2003 .
- ^ Сенгупта С., Животовский Л.А., Кинг Р., Мехди С.К., Эдмондс К.А., Чоу К.Э. и др. (февраль 2006 г.). «Полярность и временность распределения Y-хромосомы в Индии с высоким разрешением определяют как местное, так и экзогенное распространение и выявляют незначительное генетическое влияние скотоводов Центральной Азии» . Американский журнал генетики человека . 78 (2): 202–221. дои : 10.1086/499411 . ПМЦ 1380230 . ПМИД 16400607 . «Хотя значительное культурное влияние на социальную иерархию и язык в Южной Азии связано с приходом кочевых скотоводов из Центральной Азии, генетические данные (митохондриальные и Y-хромосомные) привели к крайне противоречивым выводам о генетическом происхождении племен и каст Южной Азии. Мы стремились разрешить этот конфликт, используя данные высокого разрешения по 69 информативным бинарным маркерам Y-хромосомы и 10 микросателлитным маркерам из большого набора географически, социально и лингвистически репрезентативных этнических групп Южной Азии. ранее существовавший генофонд был незначительным. Возраст накопленных микросателлитных вариаций в большинстве индийских гаплогрупп превышает 10–15 тыс. лет, что свидетельствует о древности региональной дифференциации. Поэтому наши данные не подтверждают модели, предполагающие выраженную новейшую генетическую дифференциацию. данные из Центральной Азии для объяснения наблюдаемых генетических вариаций в Южной Азии. Гаплогруппы R1a1 и R2 указывают на демографическую сложность, несовместимую с недавней единой историей. ПОЛУОСТРОВ ИНДИЙСКИЙ РЕГИОН».
- ^ Тансим И., Тангарадж К., Чауби Дж., Сингх В.К., Бхаскар Л.В., Редди Б.М. и др. (август 2006 г.). «Генетическое сходство среди низших каст и племенных групп Индии: вывод на основе Y-хромосомы и митохондриальной ДНК» . БМК Генетика . 7:42 . дои : 10.1186/1471-2156-7-42 . ПМЦ 1569435 . ПМИД 16893451 .
- ^ Саху С., Сингх А., Химабинду Г., Банерджи Дж., Ситалакшими Т., Гайквад С. и др. (январь 2006 г.). «Предыстория индийских Y-хромосом: оценка сценариев демического распространения» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (4): 843–848. Бибкод : 2006PNAS..103..843S . дои : 10.1073/pnas.0507714103 . ПМК 1347984 . ПМИД 16415161 .
- ^ Тангарадж К., Найду Б.П., Кривелларо Ф., Таманг Р., Упадхьяй С., Шарма В.К. и др. (декабрь 2010 г.). Кордо Р. (ред.). «Влияние естественных барьеров на формирование генетической структуры популяций Махараштры» . ПЛОС ОДИН . 5 (12): e15283. Бибкод : 2010PLoSO...515283T . дои : 10.1371/journal.pone.0015283 . ПМК 3004917 . ПМИД 21187967 .
- ^ Перейти обратно: а б с д и Сенгупта 2006 .
- ^ Перейти обратно: а б с д и Саху и др. 2006 год .
- ^ Перейти обратно: а б Тангарадж и др. 2010 .
- ^ Саху и др. 2006 , с. 845-846.
- ^ Перейти обратно: а б Лалуэса-Фокс, К. (2022). Неравенство: генетическая история . МТИ Пресс. стр. 81–82. ISBN 978-0-262-04678-7 . Архивировано из оригинала 16 июля 2023 года . Проверено 16 июля 2023 г.
- ^ Нарасимхан и др. 2019 .
- ^ Хаак и др. 2015 , с. 4.
- ^ Перейти обратно: а б с Маскареньяс и др. 2015 , с. 9.
- ^ Перейти обратно: а б Позник и др. 2016 , с. 5.
- ↑ Английский блог Араме, Y-ДНК с древнего Ближнего Востока. Архивировано 27 ноября 2016 г., в Wayback Machine.
- ^ «Блог антропологии Диенекеса: большая новая статья о глубоком происхождении Y-гаплогруппы R1a (Underhill et al. 2014)» . 27 марта 2014 года. Архивировано из оригинала 20 декабря 2019 года . Проверено 20 декабря 2019 г. [ ненадежный источник? ]
- ^ Перейти обратно: а б Нарасимхан и др. 2018 .
- ^ Перейти обратно: а б с "О нас" . ДНК генеалогического древа . Архивировано из оригинала 15 августа 2019 года . Проверено 20 декабря 2019 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа «ISOGG 2017 Гаплогруппа R Y-ДНК» . isogg.org . Архивировано из оригинала 10 февраля 2007 года . Проверено 20 декабря 2019 г.
- ^ Перейти обратно: а б с д и ж г час я дж к «Гаплогруппа R (Y-ДНК) — SNPedia» . www.snpedia.com . Архивировано из оригинала 5 мая 2018 года . Проверено 20 декабря 2019 г.
- ^ Карафет и др. 2014 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м Андерхилл и др. 2014 , с. 125.
- ^ «R1a в Ямной» . Блог Еврогенеса . 21 марта 2016. Архивировано из оригинала 5 мая 2018 года . Проверено 20 декабря 2019 г.
- ^ «Гаплогруппа R Y-ДНК и ее субклады» . Международное общество генетической генеалогии (ISOGG). Архивировано из оригинала 30 марта 2019 года . Проверено 8 января 2011 г.
- ^ Кран, Томас. «Проект дерева Y-хромосомы» . ДНК генеалогического древа . Архивировано из оригинала 26 мая 2013 года . Проверено 7 декабря 2012 г.
- ^ Стрим 2006 .
- ^ Фредер, Джанин (2010). Средневековые скелеты Узедома: антропологическое исследование с особым учетом этнического происхождения [ Антропологическое исследование с учетом этнического происхождения ] (Диссертация) (на немецком языке). Свободный университет Берлина. п. 86. дои : 10.17169/refubium-8995 .
- ^ https://cyberleninka.ru/article/n/tyurki-kavkaza-sravnitelnyy-analiz-genofondov-po-dannym-o-y-hromosome Archived November 7, 2023, at the Wayback Machine "высокая частота R1a среди кубанских ногайцев (субветвь R1a1a1g-M458 забирает 18%"
- ^ Андерхилл, Пенсильвания; и др. (2009). «Разделение постледникового происхождения европейских и азиатских y-хромосом внутри гаплогруппы R1a» . Европейский журнал генетики человека . 18 (4): 479–484. дои : 10.1038/ejhg.2009.194 . ПМЦ 2987245 . ПМИД 19888303 .
- ^ Гвоздз, Питер (6 августа 2018 г.). «Польские клады Y-ДНК» . Архивировано из оригинала 15 июля 2016 года . Проверено 15 июля 2016 г.
- ^ Павловский и др. 2002 .
- ^ Перейти обратно: а б Гвоздь 2009 .
- ^ Перейти обратно: а б Карс, Мэн; Башак, АН; Онат, О.Э.; Билгувар, К.; Чой, Дж.; Итан, Ю.; Чаглар, К.; Палвадо, Р.; Казанова, Дж.Л.; Купер, Д.Н.; Стенсон, доктор медицинских наук; Явуз, А.; Булуш, Х.; Гюнель, М.; Фридман, Дж. М.; Озчелик, Т. (2021). «Генетическая структура турецкого населения демонстрирует высокий уровень изменчивости и примеси» . Труды Национальной академии наук Соединенных Штатов Америки . 118 (36): e2026076118. Бибкод : 2021PNAS..11826076K . дои : 10.1073/pnas.2026076118 . ПМЦ 8433500 . ПМИД 34426522 .
- ^ Петрейчикова, Ева; Сотак, Мирослав; Бернасовска, Ярмила; Бернасовский, Иван; Совикова, Адриана; Бозикова, Александра; Боронова, Ивета; Швикова, Петра; Габрикова, Дана; МаЦекова, Сона (2009). «Частоты Y-гаплогруппы в населении словацких цыган» . Антропологическая наука . 117 (2): 89–94. дои : 10.1537/ase.080422 .
- ^ Перейти обратно: а б с Сааг и др. 2020 , с. 5.
- ^ Перейти обратно: а б Сааг и др. 2020 , с. 29, таблица 1.
- ^ Сааг и др. 2020 , Дополнительные данные 2, строка
- ^ Пост, Козимо; Ю, Хэ; Галичи, Айшин; Ружье, Элен; Кревкер, Изабель; Хуан, Илей; Рингбауэр, Харальд; Рорлах, Адам Б.; Нэгеле, Катрин; Вильяльба-Моуко, Ванесса; Радзевичюте, Рита; Феррас, Тьяго; Стессель, Александр; Тухбатова, Резеда; Друкер, Дороти Г. (1 марта 2023 г.). «Палеогеномика от верхнего палеолита до неолита европейских охотников-собирателей» . Природа . 615 (7950): 117–126. дои : 10.1038/s41586-023-05726-0 . hdl : 10256/23099 . ISSN 1476-4687 .
- ^ Фу и др. 2016 .
- ^ Сааг и др. 2017 .
- ^ Энтони 2019 , стр. 16, 17.
- ^ Перейти обратно: а б Хаак и др. 2008 .
- ^ Брандит и др. 2013 .
- ^ Мальмстрем и др. 2019 , стр. 2.
- ^ Сааг и др. 2020 г. , Дополнительные данные 2, строки 5–49.
- ^ Швейцер, Д. (23 марта 2008 г.). «Анализ данных пещеры Лихтенштейн» (PDF) . dirkschweitzer.net. Архивировано из оригинала (PDF) 14 августа 2011 года. Краткое изложение на английском языке Schilz (2006) .
- ^ Перейти обратно: а б с Кейзер и др. 2009 .
- ^ Рико и др. 2004
- ^ Korniyenko, I. V.; Vodolazhsky D. I. "Использование нерекомбинантных маркеров Y-хромосомы в исследованиях древних популяций (на примере поселения Танаис)" [The use of non-recombinant markers of the Y-chromosome in the study of ancient populations (on the example of the settlement of Tanais)]. Материалы Донских антропологических чтений [Materials of the Don Anthropological Readings]. Rostov-on-Don: Rostov Research Institute of Oncology, 2013.
- ^ Чуньсян Ли и др. 2010 .
- ^ Ким и др. 2010 .
- ^ Балановский и др. 2008 год .
- ^ Бехар и др. 2003 .
- ^ Касперавичюте, Кучинскас и Стоункинг 2005 .
- ^ Перейти обратно: а б Батталья и др. 2008 год .
- ^ Перейти обратно: а б Россер и др. 2000 .
- ^ Тамбетс и др. 2004 .
- ^ Боуден и др. 2008 год .
- ^ Дюпюи и др. 2005
- ^ Пассарино и др. 2002 .
- ^ Капелли и др. 2003 .
- ^ Кайзер и др. 2005 .
- ^ Скоццари и др. 2001
- ^ Андерхилл, Питер А. (1 января 2015 г.). «Филогенетическая и географическая структура гаплогруппы R1a Y-хромосомы» . Европейский журнал генетики человека . 23 (1): 124–131. дои : 10.1038/ejhg.2014.50 . ПМК 4266736 . ПМИД 24667786 .
- ^ Л. Барах; и др. (2003). «Y-хромосомное наследие хорватского населения и его островных изолятов» (PDF) . Европейский журнал генетики человека . 11 (7): 535–42. дои : 10.1038/sj.ejhg.5200992 . ПМИД 12825075 . S2CID 15822710 . Архивировано из оригинала (PDF) 17 декабря 2012 года . Проверено 10 сентября 2009 г.
- ^ С. Роотси; и др. (2004). «Филогеография гаплогруппы I Y-хромосомы раскрывает различные области доисторического потока генов в Европе» (PDF) . Американский журнал генетики человека . 75 (1): 128–137. дои : 10.1086/422196 . ПМК 1181996 . ПМИД 15162323 . Архивировано из оригинала (PDF) 5 сентября 2020 г. Проверено 13 февраля 2021 г.
- ^ М. Перичич; и др. (2005). «Филогенетический анализ Юго-Восточной Европы с высоким разрешением прослеживает основные эпизоды потока отцовских генов среди славянских популяций» . Молекулярная биология и эволюция . 22 (10): 1964–75. дои : 10.1093/molbev/msi185 . ПМИД 15944443 .
- ^ М. Перичич; и др. (2005). «Обзор хорватского генетического наследия, выявленного с помощью митохондриальной ДНК и Y-хромосомных линий». Хорватский медицинский журнал . 46 (4): 502–513. ПМИД 16100752 .
- ^ Перичич и др. 2005
- ^ «Без названия» . pereformat.ru (на русском языке). Архивировано из оригинала 15 марта 2016 года . Проверено 29 мая 2017 г.
- ^ «Без названия» . www.rodstvo.ru . Архивировано из оригинала 16 сентября 2021 года . Проверено 29 мая 2017 г.
- ^ Зержал и др. 2002 .
- ^ Хабер и др. 2012 .
- ^ Перейти обратно: а б Ди Кристофаро и др. 2013 .
- ^ Перейти обратно: а б Малярчук и др. 2013 .
- ^ Аширбеков и др. 2017 .
- ^ Шах 2011 .
- ^ Арункумар 2012 .
- ^ Тоомас Кивисилд; Сийри Роотси; Мейт Метспалу; Эне Мецпалу; Юрий Парик; Катрин Калдма; Эсиен Усанга; Сарабджит Мастана; Суриндер С. Папиха; Ричард Виллемс. «Генетика языка и распространения сельского хозяйства в Индии» (PDF) . В П. Белввуде; К. Ренфрю (ред.). Исследование гипотезы расселения сельского хозяйства/языка . Монографии Института Макдональда. Кембриджский университет. стр. 215–222. Архивировано из оригинала (PDF) 19 февраля 2006 г. Проверено 20 декабря 2019 г.
- ^ Форнарино и др. 2009 .
- ^ Ван и др. 2003 .
- ^ Чжоу и др. 2007 .
- ^ Лю Шу-ху и др. 2018 .
- ^ Чжун и др. 2011 .
- ^ Чжун, Хуа; Ши, Хун; Ци, Сюэ-Бин; Дуань, Цзы-Юань; Тан, Пин-Пин; Джин, Ли; Су, Бинг; Ма, Ранлин З. (2011). «Расширенное исследование Y-хромосомы предполагает постледниковую миграцию современного человека в Восточную Азию по Северному маршруту» . Молекулярная биология и эволюция . 28 (1): 717–727. дои : 10.1093/molbev/msq247 . ПМИД 20837606 .
- ^ Шоу, Вэй-Хуа; Цяо, Внь-Фа; Вэй, Чуан-Юй; Донг, Юн-Ли; Тан, Си-Цзе; Ши, Хун; Тан, Вэнь-Ру; Сяо, Чун-Цзе (2010). «Распределение Y-хромосомы среди популяций Северо-Западного Китая указывает на значительный вклад скотоводов Центральной Азии и меньшее влияние западных евразийцев» . Журнал генетики человека . 55 (5): 314–322. дои : 10.1038/jhg.2010.30 . ПМИД 20414255 . S2CID 23002493 .
- ^ Лелль и др. 2002
- ^ Чангмай, Пия; Джайсамут, Китипонг; Кампуансай, Ятуполь; и др. (2022). «Индийское генетическое наследие в популяциях Юго-Восточной Азии» . ПЛОС Генетика . 18 (2): e1010036. дои : 10.1371/journal.pgen.1010036 . ПМЦ 8853555 . ПМИД 35176016 .
- ^ Мохаммад и др. 2009 .
- ^ Насидзе и др. 2004 .
- ^ Насидзе и др. 2005 .
- ^ Груни и др. 2012 .
Источники
[ редактировать ]- Аллентофт, Мортен Э.; Сикора, Мартин; Шегрен, Карл-Горан; Расмуссен, Саймон; Расмуссен, Мортен; Стендерап, Йеспер; Дамгаард, Питер Б.; Шредер, Ханнес; и др. (2015). «Популяционная геномика Евразии бронзового века» . Природа . 522 (7555): 167–172. Бибкод : 2015 Природа.522..167А . дои : 10.1038/nature14507 . ПМИД 26062507 . S2CID 4399103 . Архивировано из оригинала 13 июля 2019 года . Проверено 23 июля 2019 г.
- Энтони, Дэвид В. (2007), Лошадь, колесо и язык. Как всадники бронзового века из евразийских степей сформировали современный мир , Princeton University Press
- Энтони, Дэвид (весна – лето 2019 г.). «Археология, генетика и язык в степях: комментарий к Бомхарду» . Журнал индоевропейских исследований . 47 (1–2). Архивировано из оригинала 3 мая 2024 года . Проверено 9 января 2020 г.
- Энтони, Дэвид; Ринге, Дон (2015), «Индоевропейская родина с лингвистической и археологической точек зрения», Annual Review of Linguistics , 1 : 199–219, doi : 10.1146/annurev-linguist-030514-124812
- Шах, AM; Таманг, Р.; Мурджани, П.; Рани, Д.С.; Говиндарадж, П.; Кулкарни, Г.; Бхаттачарья, Т.; Мустак, М.С.; Бхаскар, ЛВКС; Редди, AG; Гадви, Д.; Гай, ПБ; Чаубей, Г.; Паттерсон, Н.; Райх, Д.; Тайлер-Смит, К.; Сингх, Л.; Тангарадж, К. (2011). «Индийские сидди: африканские потомки с индийской примесью» . Американский журнал генетики человека . 89 (1): 154–61. дои : 10.1016/j.ajhg.2011.05.030 . ПМК 3135801 . ПМИД 21741027 .
- Арун Кумар, Г; Сория-Эрнанц, Германия; Кавита, виджей; Арун, В.С.; Шьяма, А; Ашокан, Канзас (2012). «Дифференциация населения мужских родов Южной Индии коррелирует с сельскохозяйственной экспансией, предшествовавшей кастовой системе» . ПЛОС ОДИН . 7 (11): е50269. Бибкод : 2012PLoSO...750269A . дои : 10.1371/journal.pone.0050269 . ПМК 3508930 . ПМИД 23209694 .
- Аширбеков Э.Э.; и др. (2017). «Распространение гаплогрупп Y-хромосомы казахов Южно-Казахстанской, Жамбылской и Алматинской областей» (PDF) . Доклады Национальной академии наук Республики Казахстан . 6 (316): 85–95. Архивировано из оригинала (PDF) 23 сентября 2021 года . Проверено 4 июня 2020 г.
- Балановский О, Роотси С, Пшеничнов А, Кивисилд Т, Чурносов М, Евсеева И, Почешхова Е, Болдырева М и др. (2008). «Два источника русского отцовского наследия в евразийском контексте» . Американский журнал генетики человека . 82 (1): 236–250. дои : 10.1016/j.ajhg.2007.09.019 . ПМК 2253976 . ПМИД 18179905 .
- Балановский, О. П. (November 30, 2015). Генофонд Европы (in Russian). KMK Scientific Press. ISBN 9785990715707 . Архивировано из оригинала 3 мая 2024 года . Проверено 8 августа 2020 г.
- Батталья В., Форнарино С., Аль-Захери Н., Оливьери А., Пала М., Майрес Н.М., Кинг Р.Дж., Рутси С. и др. (2008). «Y-хромосомные свидетельства культурного распространения сельского хозяйства в Юго-Восточной Европе» . Европейский журнал генетики человека . 17 (6): 820–30. дои : 10.1038/ejhg.2008.249 . ПМК 2947100 . ПМИД 19107149 .
- Бехар Д., Томас М.Г., Скорецки К., Хаммер М.Ф., Булыгина Е., Розенгартен Д., Джонс А.Л., Хелд К. и др. (2003). «Множественное происхождение ашкеназских левитов: свидетельства Y-хромосомы как ближневосточного, так и европейского происхождения» (PDF) . Американский журнал генетики человека . 73 (4): 768–779. дои : 10.1086/378506 . ПМК 1180600 . ПМИД 13680527 . Архивировано (PDF) из оригинала 17 апреля 2007 г. Проверено 23 апреля 2007 г.
- Боуден Г.Р., Балареск П., Кинг Т.Э., Хансен З., Ли А.С., Пергл-Уилсон Г., Херли Э., Робертс С.Дж. и др. (2008). «Раскопки прошлых структур населения путем выборки на основе фамилий: генетическое наследие викингов в северо-западной Англии» . Молекулярная биология и эволюция . 25 (2): 301–309. дои : 10.1093/molbev/msm255 . ПМЦ 2628767 . ПМИД 18032405 .
- Брандит, Г.; и др. (Генографический консорциум) (2013). «Древняя ДНК раскрывает ключевые этапы формирования генетического разнообразия митохондрий Центральной Европы» . Наука . 342 (6155): 257–261. Бибкод : 2013Sci...342..257B . дои : 10.1126/science.1241844 . ПМК 4039305 . ПМИД 24115443 .
- Капелли С., Редхед Н., Абернети Дж.К., Гратрикс Ф., Уилсон Дж.Ф., Моен Т., Хервиг Т., Ричардс М. и др. (2003). «Перепись хромосом AY на Британских островах» (PDF) . Современная биология . 13 (11): 979–84. Бибкод : 2003CBio...13..979C . дои : 10.1016/S0960-9822(03)00373-7 . ПМИД 12781138 . S2CID 526263 . Архивировано (PDF) из оригинала 8 июля 2021 г. Проверено 8 августа 2020 г. также в «Университетский колледж Лондона» (PDF) . Архивировано (PDF) из оригинала 17 января 2017 г. Проверено 29 января 2007 г.
- Чуньсян Ли; Хунцзе Ли; Иньцю Цуй; Чэнчжи Се; Давэй Цай; Вэньин Ли; Виктор Х Майр; Чжи Сюй; и др. (2010). «Свидетельства того, что смешанное население Запада и Востока жило в Таримской котловине еще в раннем бронзовом веке» . БМК Биология . 8 (1): 15. дои : 10.1186/1741-7007-8-15 . ПМЦ 2838831 . ПМИД 20163704 .
- Ди Кристофаро Дж., Пеннарун Э., Мазьер С., Майрес Н.М., Лин А.А., Темори С.А., Метспалу М., Метспалу Е. и др. (2013). «Афганский Гиндукуш: место, где сходятся потоки генов Евразийского субконтинента» . ПЛОС ОДИН . 8 (10). е76748. Бибкод : 2013PLoSO...876748D . дои : 10.1371/journal.pone.0076748 . ПМЦ 3799995 . ПМИД 24204668 .
- Дюпюи Б.М., Стенерсен М., Лу Т.Т., Олайсен Б. (2005). «Географическая гетерогенность линий Y-хромосомы в Норвегии» (PDF) . Международная судебно-медицинская экспертиза . 164 (1): 10–19. doi : 10.1016/j.forsciint.2005.11.009 . ПМИД 16337760 . Архивировано (PDF) из оригинала 22 июля 2012 г. Проверено 18 ноября 2009 г.
- Форнарино, Симона; Пала, Мария; Батталья, Винченца; Маранта, Рамона; Ахилли, Алессандро; Модиано, Гвидо; Торрони, Антонио; Семино, Орнелла; и др. (2009). «Митохондриальное и Y-хромосомное разнообразие Таруса (Непал): резервуар генетических вариаций» . Эволюционная биология BMC . 9 (1): 154. Бибкод : 2009BMCEE...9..154F . дои : 10.1186/1471-2148-9-154 . ПМК 2720951 . ПМИД 19573232 .
- Фу, Цяомэй; и др. (2 мая 2016 г.). «Генетическая история Европы ледникового периода» . Природа . 534 (7606): 200–205. Бибкод : 2016Natur.534..200F . дои : 10.1038/nature17993 . hdl : 10211.3/198594 . ПМЦ 4943878 . ПМИД 27135931 .
- Гругни В., Батталья В., Кашани Б.Х., Пароло С., Аль-Захери Н., Ахилли А., Оливьери А., Гандини Ф., Хоушманд М., Санати М.Х., Торрони А., Семино О (2012). «Древние миграционные события на Ближнем Востоке: новые данные по Y-хромосомной вариации современных иранцев» . ПЛОС ОДИН . 7 (7). е41252. Бибкод : 2012PLoSO...741252G . дои : 10.1371/journal.pone.0041252 . ПМЦ 3399854 . ПМИД 22815981 .
- Гвоздз (2009). «Горы Y-STR в гаплопространстве, Часть II: Приложение к общим польским кладам» (PDF) . Журнал генетической генеалогии . 5 (2). Архивировано (PDF) из оригинала 21 июля 2011 г. Проверено 28 ноября 2009 г.
- Хаак, В.; Брандт, Г.; Джонг, HN d.; Мейер, К.; Гансмейер, Р.; Хейд, В.; Хоксворт, К.; Пайк, AWG; и др. (2008). «Древняя ДНК, изотопы стронция и остеологический анализ проливают свет на социальную и родственную организацию позднего каменного века» . Труды Национальной академии наук . 105 (47): 18226–18231. Бибкод : 2008PNAS..10518226H . дои : 10.1073/pnas.0807592105 . ПМК 2587582 . ПМИД 19015520 .
- Хаак, Вольфганг; Лазаридис, Иосиф; Паттерсон, Ник; Роланд, Надин; Маллик, Свапан; Ламас, Бастьен; Брандт, Гвидо; Норденфельт, Сюзанна; и др. (2015). «Массовая миграция из степи является источником индоевропейских языков в Европе» . Природа . 522 (7555): 207–211. arXiv : 1502.02783 . Бибкод : 2015Natur.522..207H . биоRxiv 10.1101/013433 . дои : 10.1038/NATURE14317 . ПМК 5048219 . ПМИД 25731166 .
- Хабер М., Платт Д.Э., Ашрафиан Бонаб М., Юханна С.К., Сориа-Эрнанц Д.Ф., Мартинес-Крус Б., Дуайхи Б., Гассибе-Саббах М. и др. (2012). «Этнические группы Афганистана имеют общее Y-хромосомное наследие, структурированное историческими событиями» . ПЛОС ОДИН . 7 (3). е34288. Бибкод : 2012PLoSO...734288H . дои : 10.1371/journal.pone.0034288 . ПМЦ 3314501 . ПМИД 22470552 .
- Карафет Татьяна М.; Мендес, Фернандо Л.; Судойо, Херавати; Лансинг, Дж. Стивен; Хаммер, Майкл Ф. (2014). «Улучшенное филогенетическое разрешение и быстрая диверсификация гаплогруппы Y-хромосомы K-M526 в Юго-Восточной Азии» . Природа . 23 (3): 369–373. дои : 10.1038/ejhg.2014.106 . ПМЦ 4326703 . ПМИД 24896152 .
- Касперавичюте, Д.; Кучинскас, В.; Стоункинг, М. (2005). «Вариации Y-хромосомы и митохондриальной ДНК у литовцев». Анналы генетики человека . 68 (5): 438–452. дои : 10.1046/j.1529-8817.2003.00119.x . ПМИД 15469421 . S2CID 26562505 .
- Кайзер М., Лао О., Анслингер К., Августин С., Баргель Г., Эдельманн Дж., Элиас С., Генрих М. и др. (2005). «Значительная генетическая дифференциация между Польшей и Германией соответствует современным политическим границам, как показал анализ Y-хромосомы» (PDF) . Генетика человека . 117 (5): 428–443. дои : 10.1007/s00439-005-1333-9 . ПМИД 15959808 . S2CID 11066186 . Архивировано из оригинала (PDF) 4 марта 2009 г.
- Кейзер, Кристина; Буаказе, Кэролайн; Крубези, Эрик; Николаев Валерий Георгиевич; Монтаньон, Даниэль; Рейс, Татьяна; Людес, Бертран (2009). «Древняя ДНК дает новое понимание истории южносибирских курганцев». Генетика человека . 126 (3): 395–410. дои : 10.1007/s00439-009-0683-0 . ПМИД 19449030 . S2CID 21347353 .
- Ким, Киджон; Бреннер, Чарльз Х.; Майр, Виктор Х.; Ли, Кван-Хо; Ким, Джэ Хён; Гелегдорж, Эрегзен; Батболд, Нацаг; Сон, И-Чунг; и др. (2010). «Западно-евразийский мужчина найден на 2000-летнем элитном кладбище хунну на северо-востоке Монголии». Американский журнал физической антропологии . 142 (3): 429–440. дои : 10.1002/ajpa.21242 . ПМИД 20091844 .
- Кивисилд, Т; Рутси, С; Метспалу, М; Мастана, С; Калдма, К; Парик, Дж; Метспалу, Э; Адохаан, М; и др. (2003). «Генетическое наследие первых поселенцев сохраняется как в индийских племенных, так и в кастовых популяциях» . АДЖХГ . 72 (2): 313–32. дои : 10.1086/346068 . ПМК 379225 . ПМИД 12536373 .
- Лазаридис, Иосиф; и др. (2016). «Геномный взгляд на происхождение земледелия на древнем Ближнем Востоке» . Природа . 536 (7617): 419–424. Бибкод : 2016Natur.536..419L . дои : 10.1038/nature19310 . ПМЦ 5003663 . ПМИД 27459054 .
- Лелл Дж.Т., Сукерник Р.И., Стариковская Ю.Б., Су Б., Джин Л., Шурр Т.Г., Андерхилл П.А., Уоллес округ Колумбия (2002). «Двойное происхождение и сибирское сходство Y-хромосом коренных американцев» (PDF) . Американский журнал генетики человека . 70 (1): 192–206. дои : 10.1086/338457 . ПМЦ 384887 . ПМИД 11731934 . Архивировано из оригинала (PDF) 22 апреля 2003 г.
- Лю Шу-ху; Низам Йилихам; Ребиам Бэке; Абдукерам Бупатима; Долкун Мэтьюз (2018). «Исследование генетического разнообразия трех изолированных популяций в Синьцзяне с использованием Y-SNP». Acta Anthropologica Sinica 37 (1): 146–156.
- Карлос Килес (10 сентября 2018 г.). «Исследование генетического разнообразия трех изолированных популяций в Синьцзяне с использованием Y-SNP» . Индоевропейский . Архивировано из оригинала 20 сентября 2020 года . Проверено 24 июля 2020 г.
- Мальмстрем, Хелена; Гюнтер, Торстен; Свенссон, Эмма М.; Юрас, Анна; Фрейзер, Магдалена; Мунтерс, Ариэль Р.; Поспешный, Лукаш; Тырв, Мари; и др. (9 октября 2019 г.). «Геномное происхождение людей скандинавской культуры боевых топоров и их связь с более широким горизонтом шнуровой керамики» . Труды Королевского общества Б. 286 (1912). дои : 10.1098/rspb.2019.1528 . ПМК 6790770 . ПМИД 31594508 .
- Малярчук Борис; Деренко, Мирослава; Возняк, Марцин; Гжибовский, Томаш (2013). «Вариация Y-хромосомы у таджиков и иранцев». Анналы биологии человека . 40 (1): 48–54. дои : 10.3109/03014460.2012.747628 . ПМИД 23198991 . S2CID 2752490 .
- Маскареньяс, Десмонд Д.; Райна, Анупума; Астон, Кристофер Э.; Сангера, Дхарабир К. (2015). «Генетическая и культурная реконструкция миграции древней линии» . БиоМед Исследования Интернэшнл . 2015 : 651415. doi : 10.1155/2015/651415 . ПМК 4605215 . ПМИД 26491681 .
- Мэтисон, Иэн; Лазаридис, Иосиф; Роланд, Надин; Маллик, Свапан; Паттерсон, Ник; Алпаслан Руденберг, Сонгул; Харни, Эдаоин; Стюардсон, Кристин; и др. (2015). «Восемь тысяч лет естественного отбора в Европе». биоRxiv 10.1101/016477 .
- Мирабал, Шейла; Регейро, М; Каденас, AM; Кавалли-Сфорца, LL; Андерхилл, Пенсильвания; Вербенко Д.А.; Лимборска, ЮАР; Эррера, Р.Дж.; и др. (2009). «Распространение Y-хромосомы в геолингвистическом ландшафте северо-запада России» . Европейский журнал генетики человека . 17 (10): 1260–1273. дои : 10.1038/ejhg.2009.6 . ПМЦ 2986641 . ПМИД 19259129 .
- Мохаммад Т., Сюэ Ю., Эвисон М., Тайлер-Смит С. (2009). «Генетическая структура кочевых бедуинов из Кувейта» . Наследственность . 103 (5): 425–433. дои : 10.1038/hdy.2009.72 . ПМК 2869035 . ПМИД 19639002 .
- Нарасимхан, Вагиш М.; Энтони, Дэвид; Мэллори, Джеймс; Райх, Дэвид (2018). «Геномное формирование Южной и Центральной Азии». bioRxiv 10.1101/292581 .
- Нарасимхан, Вагиш М.; Паттерсон, Нью-Джерси; Мурджани, Прия; Роланд, Надин; и др. 365 Science, , PMID eaat7487 ( 6457): doi : 10.1126 , PMC 6822619 , science.aat7487 /
- Насидзе И., Линг Е.Ю., Куинке Д., Дюпанлуп И., Кордо Р., Рычков С., Наумова О., Жукова О. и др. (2004). «Митохондриальная ДНК и вариации Y-хромосомы на Кавказе» (PDF) . Анналы генетики человека . 68 (Часть 3): 205–221. дои : 10.1046/j.1529-8817.2004.00092.x . ПМИД 15180701 . S2CID 27204150 . Архивировано из оригинала (PDF) 30 октября 2004 г.
- Насидзе И., Куинке Д., Озтюрк М., Бендукидзе Н., Стоункинг М. (2005). «Вариации мтДНК и Y-хромосомы в курдских группах» (PDF ) Анналы генетики человека 69 (Часть 4): 401–412. дои : 10.1046/j.1529–8817.2005.00174.x . ПМИД 15996169 . S2CID 23771698 . Архивировано из оригинала (PDF) 23 августа.
- Памджав, Хоролма; Фехер, Тибор; Немет, Эндре; Падар, Жолт (2012), «Краткое сообщение: новые бинарные маркеры Y-хромосомы улучшают филогенетическое разрешение в гаплогруппе R1a1», Американский журнал физической антропологии , 149 (4): 611–615, doi : 10.1002/ajpa.22167 , PMID 23115110 , S2CID 4820868
- Пассарино Г., Каваллери Г.Л., Лин А.А., Кавалли-Сфорца Л.Л., Борресен-Дейл А.Л., Андерхилл (2002). «Различные генетические компоненты норвежской популяции, выявленные с помощью анализа полиморфизмов мтДНК и Y-хромосомы» . Европейский журнал генетики человека . 10 (9): 521–529. дои : 10.1038/sj.ejhg.5200834 . ПМИД 12173029 .
- Патхак, Аджай К.; Кадиан, Анураг; Кушнеревич Алена; Монтинаро, Франческо; Мондал, Маюх; Онгаро, Линда; Сингх, Манвендра; Кумар, Прамод; и др. (6 декабря 2018 г.). «Генетическое происхождение современного населения долины Инда из северо-западной Индии» . Американский журнал генетики человека . 103 (6): 918–929. дои : 10.1016/j.ajhg.2018.10.022 . ПМК 6288199 . ПМИД 30526867 .
- Павловский, Р; Детлафф-Какол, А; Макиеевска, А; Пашковска, Р; Райхерт, М; Езерски, Г. (2002). «Популяционная генетика 9 локусов STR Y-хромосомы в Северной Польше». Арх. Мед. Садова Крыминол . 52 (4): 261–277. ПМИД 14669672 .
- Перичич М., Лаук Л.Б., Кларич И.М., Роотси С., Яничиевич Б., Рудан И., Терзич Р., Чолак И. и др. (2005). «Филогенетический анализ Юго-Восточной Европы с высоким разрешением прослеживает основные эпизоды потока отцовских генов среди славянских популяций» . Мол. Биол. Эвол . 22 (10): 1964–75. дои : 10.1093/molbev/msi185 . ПМИД 15944443 .
- Позник Г.Д. и др. (2016). «Периодические всплески демографии мужского пола, выявленные на основе 1244 последовательностей Y-хромосомы по всему миру» . Природная генетика . 48 (6): 593–599. дои : 10.1038/ng.3559 . hdl : 11858/00-001M-0000-002A-F024-C . ПМЦ 4884158 . ПМИД 27111036 .
- Регейро, М; Цепи, АМ; Гайден, Т; Андерхилл, Пенсильвания; Эррера, Р.Дж. (2006). «Иран: трехконтинентальная связь для миграции, вызванной Y-хромосомой». Хум Херед 61 (3): 132–143. дои : 10.1159/000093774 . ПМИД 16770078 . S2CID 7017701 .
- Рико Ф., Кейзер-Тракки С., Буржуа I, Крубези Э., Людес Б. (2004). «Генетический анализ скифо-сибирского скелета и его значение для древних миграций в Центральной Азии». Биология человека . 76 (1): 109–25. дои : 10.1353/hub.2004.0025 . ПМИД 15222683 . S2CID 35948291 .
- Россер Ж.Х., Зержал Т., Хёрлес М.Е., Адохаан М., Алавантик Д., Аморим А., Амос В., Арментерос М. и др. (2000). «Разнообразие Y-хромосомы в Европе является клинальным и зависит в первую очередь от географии, а не от языка» . Американский журнал генетики человека . 67 (6): 1526–1543. дои : 10.1086/316890 . ПМЦ 1287948 . ПМИД 11078479 .
- Сааг, Лехти; Варул, Лийви; Шейб, Кристиана Лин; Стендерап, Йеспер; Аллентофт, Мортен Э.; Сааг, Лаури; Пагани, Лука; Рейдла, Маере; и др. (24 июля 2017 г.). «Экстенсивное земледелие в Эстонии началось благодаря миграции из степи по признаку пола» . Современная биология . 27 (14). Cell Press : 2185–2193. Бибкод : 2017CBio...27E2185S . дои : 10.1016/j.cub.2017.06.022 . ПМИД 28712569 .
- Saag, Lehti; Vasilyev, Sergey V.; Varul, Liivi; Kosorukova, Natalia V.; Gerasimov, Dmitri V.; Oshibkina, Svetlana V.; Griffith, Samuel J.; Solnik, Anu; et al. (July 3, 2020). "Genetic ancestry changes in Stone to Bronze Age transition in the East European plain". bioRxiv 10.1101/2020.07.02.184507 .
- Саху, С; Сингх, А; Химабинду, Г; Банерджи, Дж; Ситалакшими, Т; Гайквад, С; Триведи, Р; Эндикотт, П; и др. (2006). «Предыстория индийских Y-хромосом: оценка сценариев демического распространения» . Труды Национальной академии наук . 103 (4): 843–848. Бибкод : 2006PNAS..103..843S . дои : 10.1073/pnas.0507714103 . ПМК 1347984 . ПМИД 16415161 .
- Скоццари Р., Кручиани Ф., Панграцио А., Сантоламазца П., Вона Дж., Морал П., Латини В., Варези Л. и др. (2001). «Вариации Y-хромосомы человека в районе Западного Средиземноморья: последствия для заселения региона» (PDF) . Иммунология человека . 62 (9): 871–84. CiteSeerX 10.1.1.408.4857 . дои : 10.1016/S0198-8859(01) 00286-5 ПМИД 11543889 . Архивировано из оригинала (PDF) 17 декабря . Получено 15 . марта
- Семенов Александр С.; Булат, Владимир В. (2016), «Древняя палео-ДНК Северо-Восточной Европы домедного века: установление следов миграции гаплогруппы Y-ДНК R1a1», Европейский журнал молекулярной биотехнологии , 11 (1): 40–54 , doi : 10.13187/ejmb.2016.11.40 , S2CID 172131289
- Семино, О; Пассарино, Дж; Оефнер, П.Дж.; Лин, А.А.; Арбузова С; Бекман, Ле; Де Бенедиктис, Дж; Франкалаччи, П; и др. (2000). «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: хромосомная перспектива AY» (PDF) . Наука . 290 (5494): 1155–1159. Бибкод : 2000Sci...290.1155S . дои : 10.1126/science.290.5494.1155 . ПМИД 11073453 . Архивировано из оригинала (PDF) 25 ноября 2003 г.
- Сенгупта, С; Животовский, Л.А.; Кинг, Р; Мехди, SQ; Эдмондс, Калифорния; Чоу, CE; Лин, А.А.; Митра, М; и др. (2006). «Полярность и временной характер распределения Y-хромосомы с высоким разрешением в Индии определяют как местное, так и экзогенное распространение и выявляют незначительное генетическое влияние скотоводов Центральной Азии» . Американский журнал генетики человека . 78 (2): 202–21. дои : 10.1086/499411 . ПМЦ 1380230 . ПМИД 16400607 .
- Шарма, С; Рай, Э; Шарма, П; Йена, М; Сингх, С; Дарвиши, К; Бхат, АК; Бханвер, AJ; и др. (2009). «Индийское происхождение отцовской гаплогруппы R1a1(*) обосновывает автохтонное происхождение брахманов и кастовой системы» . Журнал генетики человека . 54 (1): 47–55. дои : 10.1038/jhg.2008.2 . ПМИД 19158816 .
- Шильц, Феликс (2006). Анализ молекулярно-генетического родства доисторического коллектива скелетов пещеры Лихтенштейн [ Молекулярно-генетический анализ родства доисторического коллектива скелетов пещеры Лихтенштейн ] (PDF) (Диссертация) (на немецком языке). Геттинген: Факультет математики и естественных наук Университета Георга Августа. Архивировано (PDF) из оригинала 8 июня 2011 г. Проверено 9 ноября 2009 г.
- Сильва, Марина; и др. (2017). «Генетическая хронология Индийского субконтинента указывает на сильное расселение по признаку пола» . Эволюционная биология BMC . 17 (1): 88. Бибкод : 2017BMCEE..17...88S . дои : 10.1186/s12862-017-0936-9 . ПМК 5364613 . ПМИД 28335724 .
- Тамбетс К., Роотси С., Кивисилд Т., Хелп Х., Серк П., Лугвяли Э.Л., Толк Х.В., Рейдла М. и др. (2004). «Западные и восточные корни саамов - история генетических «выбросов», рассказанная митохондриальной ДНК и Y-хромосомами» . Американский журнал генетики человека . 74 (4): 661–682. дои : 10.1086/383203 . ПМК 1181943 . ПМИД 15024688 .
- Тангарадж, Кумарасами; Найду, Б. Пратап; Кривелларо, Федерика; Таманг, Ракеш; Упадхьяй, Шашанк; Шарма, Варун Кумар; Редди, Алла Г.; Валимбе, СР; и др. (2010). Кордо, Ричард (ред.). «Влияние естественных барьеров на формирование генетической структуры популяций Махараштры» . ПЛОС ОДИН . 5 (12): e15283. Бибкод : 2010PLoSO...515283T . дои : 10.1371/journal.pone.0015283 . ПМК 3004917 . ПМИД 21187967 .
- Андерхилл, Пенсильвания; Майрес, Нью-Мексико; Рутси, С; Метспалу, М; Животовский, Л.А.; Кинг, Р.Дж.; Лин, А.А.; Чоу, CE; и др. (4 ноября 2009 г.). «Разделение постледникового происхождения европейских и азиатских Y-хромосом в пределах гаплогруппы R1a» . Европейский журнал генетики человека . 18 (4) (опубликовано в апреле 2010 г.): 479–84. дои : 10.1038/ejhg.2009.194 . ПМЦ 2987245 . ПМИД 19888303 .
- Андерхилл, Питер А.; и др. (26 марта 2014 г.). «Филогенетическая и географическая структура гаплогруппы R1a Y-хромосомы» . Европейский журнал генетики человека . 23 (1) (опубликовано в январе 2015 г.): 124–131. дои : 10.1038/ejhg.2014.50 . ПМК 4266736 . ПМИД 24667786 . «ПДФ» (PDF) . Архивировано из оригинала (PDF) 16 августа 2016 года . Проверено 12 июня 2016 г.
- Ван, Вэй; Мудрая, Шерил; Барик, Том; Блэк, Майкл Л.; Биттлс, Алан Х. (2003). «Происхождение и генетическая структура трех совместно проживающих в Китае мусульманских популяций: Салар, Боань и Дунсян». Генетика человека . 113 (3): 244–52. дои : 10.1007/s00439-003-0948-y . ПМИД 12759817 . S2CID 11138499 .
- Уэллс, Р.С. (2001), «Евразийский центр: континентальный взгляд на разнообразие Y-хромосомы», Труды Национальной академии наук США , 98 (18): 10244–10249, Бибкод : 2001PNAS...9810244W , doi : 10.1073/pnas.171305098 , PMC 56946 , PMID 11526236
- Ян, Ши; Ван, Чуан-Чао; Чжэн, Хун-Сян; Ван, Вэй; Цинь, Чжэнь-Дун; Вэй, Лан-Хай; Ван, Йи; Пан, Сюэ-Донг; и др. (29 августа 2014 г.). «Y-хромосомы 40% китайцев происходят от трех неолитических супердедушек» . ПЛОС ОДИН . 9 (8). е105691. arXiv : 1310.3897 . Бибкод : 2014PLoSO...9j5691Y . дои : 10.1371/journal.pone.0105691 . ПМК 4149484 . ПМИД 25170956 .
- Консорциум Y-хромосомы «YCC» (2002). «Система номенклатуры дерева бинарных гаплогрупп Y-хромосомы человека» . Геномные исследования . 12 (2): 339–348. дои : 10.1101/гр.217602 . ПМК 155271 . ПМИД 11827954 .
- Зержал, Татьяна; Уэллс, Р. Спенсер; Юлдашева, Надира; Рузибакиев Руслан; Тайлер-Смит, Крис (2002). «Генетический ландшафт, измененный недавними событиями: взгляд на Y-хромосому в Центральной Азии» . Американский журнал генетики человека . 71 (3): 466–82. дои : 10.1086/342096 . ПМК 419996 . ПМИД 12145751 .
- Чжун Х, Ши Х, Ци XB, Дуань Ю, Тан П.П., Цзинь Л, СУ Б, Ма РЗ (январь 2011 г.). «Расширенное исследование Y-хромосомы позволяет предположить, что современные люди мигрировали в Восточную Азию северным путем после ледникового периода» . Молекулярная биология и эволюция . 28 (1): 717–27. дои : 10.1093/molbev/msq247 . ПМИД 20837606 .
- Чжоу, Жуйся; Ан, Лиже; Ван, Сюньлин; Шао, Вэй; Линь, Гунхуа; Ю, Вейпин; Йи, Лин; Сюй, Шицзянь; и др. (2007). «Проверка гипотезы о древнеримском солдатском происхождении народа Лицянь на северо-западе Китая: взгляд на Y-хромосому» . Журнал генетики человека . 52 (7): 584–91. дои : 10.1007/s10038-007-0155-0 . ПМИД 17579807 .
Дальнейшее чтение
[ редактировать ]- Гимбутас (1970). Индоевропейцы и индоевропейцы . Филадельфия, Пенсильвания: Univ. из Пенсильвании Пресс. стр. 155–195.
- Патриция Маласпина; Андрей Иванович Козлов; Фульвио Кручиани; Пьеро Сантоламазца; Неджат Акар; Димитр Ковачев; Керимова Марина Георгиевна; Юрий Парик; Ричард Виллемс; Розана Скоццари; Андреа Новеллетто (2003). «Анализ вариаций Y-хромосомы в современных популяциях на европейско-азиатской границе» (PDF) . У К. Бойля; К. Ренфрю; М. Левин (ред.). Древние взаимодействия: Восток и Запад в Евразии . Монографии Института Макдональда. Кембридж: Издательство Кембриджского университета. стр. 309–313. Архивировано из оригинала (PDF) 24 июня 2023 года . Проверено 10 сентября 2009 г.
- Парпола, Аско (2015). Корни индуизма. Ранние арийцы и цивилизация Инда . Издательство Оксфордского университета.
- Шарма, С. (2007). «1344/T: Автохтонное происхождение и племенная связь индийских браминов: оценка с помощью молекулярно-генетических маркеров». 57-е ежегодное собрание Американского общества генетики человека, 23–27 октября 2007 г.; Сан-Диего, Калифорния (PDF) (Аннотация). п. 273. Архивировано из оригинала (PDF) 26 июня 2008 г.
- Траутманн, Томас (2005). Арийские дебаты . Издательство Оксфордского университета.
- Варзари, Александр (2006). Популяционная история Днестровско-Карпат: данные по вставке Alu и полиморфизму Y-хромосомы (PDF) (Диссертация). Мюнхен: Университет Людвига-Максимилиана. Архивировано (PDF) из оригинала 28 октября 2008 г. Проверено 30 октября 2008 г.
- Уэллс, Спенсер (2002). Путешествие человека: генетическая одиссея . Издательство Принстонского университета. ISBN 978-0-691-11532-0 .
журнальные статьи
|
|---|
Внешние ссылки
[ редактировать ]- ДНК-дерево
- Проект гаплогруппы FTDNA R1a Y-хромосомы
- Проект R1a1a1 и субкладов Y-ДНК - Фоновое генеалогическое древо ДНК R1a1a1
- ТМРЦА
- Различный
- Датский региональный проект ДНК Демеса: Гаплогруппа Y-ДНК R1a. Архивировано 20 августа 2008 г. в Wayback Machine.
- Блог Eurogenes, Полтовский выброс
- Авотайну Онлайн, Отпечаток Y-ДНК Шполера Зейды, цадика, коснувшегося мира