Ку (белок)
| Комплексная семья Ку | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | Гетеродимер Ku70:Ku80Ku70:Ku80Ku Аутоантиген | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Генные карты : [1] ; ОМА : - ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| ремонт рентгена перекрестное дополнение 5 | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Символ | XRCC5 | ||
| Альт. символы | Ку80 | ||
| ген NCBI | 7520 | ||
| HGNC | 12833 | ||
| МОЙ БОГ | 194364 | ||
| ПДБ | 1ДЖЕЙ | ||
| RefSeq | НМ_021141 | ||
| ЮниПрот | P13010 | ||
| Другие данные | |||
| Локус | 2 q35 | ||
| |||
| ремонт рентгена перекрестное дополнение 6 | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | XRCC6 | ||
| Альт. символы | Ку70, Г22П1 | ||
| ген NCBI | 2547 | ||
| HGNC | 4055 | ||
| МОЙ БОГ | 152690 | ||
| ПДБ | 1ДЖЕЙ | ||
| RefSeq | НМ_001469 | ||
| ЮниПрот | P12956 | ||
| Другие данные | |||
| Локус | Хр.22 q11-q13 | ||
| |||
| Ku70/Ku80 N-концевой альфа/бета-домен | |||
|---|---|---|---|
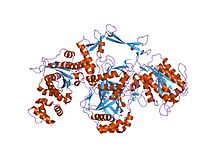 кристаллическая структура ku-гетеродимера | |||
| Идентификаторы | |||
| Символ | Ку_Н | ||
| Пфам | PF03731 | ||
| Пфам Клан | CL0128 | ||
| ИнтерПро | ИПР005161 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 1jey / SCOPe / СУПФАМ | ||
| |||
| Домен бета-ствола Ку70/Ку80 | |||
|---|---|---|---|
 кристаллическая структура гетеродимера ku, связанного с ДНК | |||
| Идентификаторы | |||
| Символ | К | ||
| Пфам | PF02735 | ||
| ИнтерПро | ИПР006164 | ||
| PROSITE | PDOC00252 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 1jey / SCOPe / СУПФАМ | ||
| |||
| Ку70/Ку80 С-концевое плечо | |||
|---|---|---|---|
кристаллическая структура гетеродимера ku, связанного с ДНК | |||
| Идентификаторы | |||
| Символ | Ку_С | ||
| Пфам | PF03730 | ||
| ИнтерПро | ИПР005160 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 1jey / SCOPe / СУПФАМ | ||
| |||
| Терминальный домен Ku C, например | |||
|---|---|---|---|
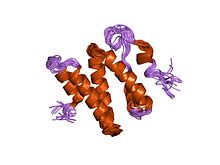 структура 3d-решения c-концевой области ku86 | |||
| Идентификаторы | |||
| Символ | Ku_PK_bind | ||
| Пфам | PF08785 | ||
| ИнтерПро | ИПР014893 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 1q2z / СКОПе / СУПФАМ | ||
| |||
Ku представляет собой димерный белковый комплекс, который связывается с ДНК двухцепочечных разрывов концами и необходим для пути негомологичного соединения концов (NHEJ) репарации ДНК . Ку эволюционно сохраняется от бактерий до человека. Предковый бактериальный Ku представляет собой гомодимер (две копии одного и того же белка, связанные друг с другом). [ 2 ] Эукариотический Ku представляет собой гетеродимер двух полипептидов Ku70 . (XRCC6) и Ku80 (XRCC5), названных так потому, что молекулярная масса человеческих белков Ku составляет около 70 кДа и 80 кДа Две субъединицы Ku образуют структуру в форме корзины, которая прикрепляется к концу ДНК . [ 1 ] После связывания Ku может скользить вниз по цепи ДНК, позволяя большему количеству молекул Ku проникнуть на конец. У высших эукариот Ku образует комплекс с каталитической субъединицей ДНК-зависимой протеинкиназы (DNA-PKcs) с образованием полной ДНК-зависимой протеинкиназы DNA-PK. [ 3 ] Считается, что Ku действует как молекулярный каркас, с которым могут связываться другие белки, участвующие в NHEJ, ориентируя двухцепочечный разрыв для лигирования.
Белки Ku70 и Ku80 состоят из трех структурных доменов . домен N-концевой представляет собой альфа /бета-домен. Этот домен вносит лишь небольшой вклад в интерфейс димеров. Домен состоит из шестинитевого бета-листа складки Россмана . [ 4 ] Центральный домен Ku70 и Ku80 представляет собой ДНК -связывающий домен бета-бочонка . Ку устанавливает лишь несколько контактов с сахаро-фосфатным остовом и ни одного контакта с основаниями ДНК , но он стерически прилегает к контурам больших и малых бороздок, образуя кольцо, которое окружает дуплексную ДНК, удерживая два полных оборота молекулы ДНК. Образуя мостик между разорванными концами ДНК, Ku структурно поддерживает и выравнивает концы ДНК, защищает их от деградации и предотвращает беспорядочное связывание с целой ДНК. Ku эффективно выравнивает ДНК, в то же время обеспечивая доступ полимераз , нуклеаз и лигаз к разорванным концам ДНК, чтобы способствовать соединению концов. [ 5 ] С -концевое плечо представляет собой альфа-спиральную область, которая охватывает центральный домен бета-цилиндра противоположной субъединицы . [ 1 ] В некоторых случаях на С-конце присутствует четвертый домен, который связывается с каталитической субъединицей ДНК-зависимой протеинкиназы . [ 6 ]
Обе субъединицы Ku были экспериментально выведены из строя у мышей . У этих мышей наблюдается хромосомная нестабильность , что указывает на то, что NHEJ важен для поддержания генома. [ 7 ] [ 8 ]
У многих организмов Ku выполняет дополнительные функции в теломерах . помимо своей роли в репарации ДНК [ 9 ]
Обилие Ku80, по-видимому, связано с долголетием вида. [ 10 ]
Старение
[ редактировать ]Мутантные мыши с дефектом Ku70 или Ku80 или двойные мутантные мыши с дефицитом Ku70 и Ku80 демонстрируют раннее старение. [ 11 ] Средняя продолжительность жизни трех мутантных линий мышей была схожа друг с другом и составляла около 37 недель по сравнению со 108 неделями для контроля дикого типа. Было исследовано шесть специфических признаков старения, и было обнаружено, что три мутантные мыши проявляют те же признаки старения, что и контрольные мыши, но в гораздо более раннем возрасте. Заболеваемость раком у мутантных мышей не увеличивалась. Эти результаты позволяют предположить, что функция Ku важна для обеспечения долголетия и что путь репарации ДНК NHEJ (опосредованный Ku) играет ключевую роль в восстановлении двухцепочечных разрывов ДНК, которые в противном случае могли бы вызвать раннее старение. [ 12 ] (Также см. теорию старения, связанную с повреждением ДНК .)
Растения
[ редактировать ]Ku70 и Ku80 также были экспериментально охарактеризованы на растениях, где они, по-видимому, играют ту же роль, что и у других эукариот. Было показано, что в рисе подавление любого белка способствует гомологичной рекомбинации (HR). [ 13 ] Этот эффект был использован для повышения эффективности нацеливания на гены (GT) у Arabidopsis thaliana . В исследовании частота ГТ на основе HR с использованием нуклеазы цинковых пальцев (ZFN) была увеличена до шестнадцати раз у ku70 . мутантов [ 14 ] Этот результат имеет многообещающее значение для редактирования генома эукариот, поскольку механизмы репарации DSB высоко консервативны. Существенная разница состоит в том, что у растений Ku также участвует в поддержании альтернативной морфологии теломер , характеризующейся тупыми концами или короткими (≤ 3-нт) 3'-выступами. [ 15 ] Эта функция не зависит от роли Ku в репарации DSB, поскольку было показано, что устранение способности комплекса Ku перемещаться вдоль ДНК сохраняет теломеры с тупыми концами, одновременно препятствуя репарации ДНК. [ 16 ]
Бактерии и археи
[ редактировать ]Бактерии обычно имеют только один ген Ku (если он у них вообще есть). Необычно, что у Mesorhizobium loti их два: mlr9624 и mlr9623 . [ 17 ]
У архей обычно также есть только один ген Ku (для ~ 4% видов, у которых он вообще есть). Эволюционная история размыта обширным горизонтальным переносом генов с бактериями. [ 18 ]
Бактериальные и архейные Ku-белки отличаются от своих эукариотических аналогов тем, что имеют только центральный домен бета-бочонка.
Имя
[ редактировать ]Название «Ку» происходит от фамилии японского пациента, у которого он был обнаружен. [ 19 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с ВВП : 1JEY ; Уокер-младший, Корпина Р.А., Голдберг Дж. (август 2001 г.). «Структура Ku-гетеродимера, связанного с ДНК, и ее значение для восстановления двухцепочечного разрыва». Природа . 412 (6847): 607–14. Бибкод : 2001Natur.412..607W . дои : 10.1038/35088000 . ПМИД 11493912 . S2CID 4371575 .
- ^ Доэрти А.Дж., Джексон С.П., Веллер Г.Р. (июль 2001 г.). «Идентификация бактериальных гомологов белков репарации ДНК Ku» . ФЭБС Летт . 500 (3): 186–8. дои : 10.1016/S0014-5793(01)02589-3 . ПМИД 11445083 . S2CID 43588474 .
- ^ Картер Т., Ванкурова И., Сан И., Лу В., ДеЛеон С. (декабрь 1990 г.). «ДНК-активируемая протеинкиназа из ядер клеток HeLa» . Мол. Клетка. Биол . 10 (12): 6460–71. дои : 10.1128/MCB.10.12.6460 . ПМК 362923 . ПМИД 2247066 .
- ^ Сугихара Т., Вадхва Р., Каул С.К., Мицуи Ю. (апрель 1999 г.). «Новый металлотионеиноподобный белок, специфичный для семенников, тесмин, является ранним маркером дифференцировки мужских зародышевых клеток». Геномика . 57 (1): 130–6. дои : 10.1006/geno.1999.5756 . ПМИД 10191092 .
- ^ Аравинд Л., Кунин Е.В. (август 2001 г.). «Прокариотические гомологи эукариотического белка Ku, связывающего концы ДНК, новые домены в белке Ku и предсказание прокариотической системы восстановления двухцепочечных разрывов» . Геном Рез . 11 (8): 1365–74. дои : 10.1101/гр.181001 . ПМК 311082 . ПМИД 11483577 .
- ^ Харрис Р., Эспозито Д., Санкар А., Маман Дж.Д., Хинкс Дж.А., Перл Л.Х., Дрисколл ПК (январь 2004 г.). «Структура 3D-решения С-концевой области Ku86 (Ku86CTR)». Дж. Мол. Биол . 335 (2): 573–82. дои : 10.1016/j.jmb.2003.10.047 . ПМИД 14672664 .
- ^ Дифилиппантонио М.Дж., Чжу Дж., Чен Х.Т., Меффре Э., Нуссенцвейг М.К., Макс Э.Э., Рид Т., Нуссенцвейг А. (март 2000 г.). «Белок репарации ДНК Ku80 подавляет хромосомные аберрации и злокачественную трансформацию» . Природа 404 (6777): 510–4. Бибкод : 2000Nature.404..510D . дои : 10.1038/35006670 . ПМЦ 4721590 . ПМИД 10761921 .
- ^ Фергюсон Д.О., Секигути Дж.М., Чанг С., Фрэнк К.М., Гао Ю., ДеПиньо Р.А., Альт Ф.В. (июнь 2000 г.). «Номологичный путь репарации ДНК с соединением концов необходим для стабильности генома и подавления транслокаций» . Учеб. Натл. акад. наук. США . 97 (12): 6630–3. Бибкод : 2000PNAS...97.6630F . дои : 10.1073/pnas.110152897 . ЧВК 18682 . ПМИД 10823907 .
- ^ Бултон С.Дж., Джексон С.П. (март 1998 г.). «Компоненты Ku-зависимого негомологичного пути соединения концов участвуют в поддержании длины теломер и молчании теломер» . ЭМБО Дж . 17 (6): 1819–28. дои : 10.1093/emboj/17.6.1819 . ПМК 1170529 . ПМИД 9501103 .
- ^ Лоренцини А., Джонсон Ф.Б., Оливер А., Трезини М., Смит Дж.С., Хдейб М., Селл С., Кристофало В.Дж., Стамато Т.Д. (ноябрь – декабрь 2009 г.). «Значительная корреляция продолжительности жизни видов с распознаванием разрыва двойной цепи ДНК, но не с длиной теломер» . Разработчик старения мехов . 130 (11–12): 784–92. дои : 10.1016/j.mad.2009.10.004 . ПМК 2799038 . ПМИД 19896964 .
- ^ Ли Х, Фогель Х, Холкомб В.Б., Гу Ю, Хэсти П. (2007). «Удаление Ku70, Ku80 или обоих вызывает раннее старение без существенного увеличения количества случаев рака» . Мол. Клетка. Биол . 27 (23): 8205–14. дои : 10.1128/MCB.00785-07 . ПМК 2169178 . ПМИД 17875923 .
- ^ Бернштейн Х, Пейн СМ, Бернштейн С, Гаревал Х, Дворжак К (2008). «Рак и старение как последствия невосстановленного повреждения ДНК». В: Новое исследование повреждений ДНК (редакторы: Хонока Кимура и Аой Судзуки) Nova Science Publishers, Нью-Йорк, глава 1, стр. 1–47. открытый доступ, но только для чтения https://www.novapublishers.com/catalog/product_info.php?products_id=43247. Архивировано 25 октября 2014 г. на Wayback Machine. ISBN 978-1604565812
- ^ Нисидзава-Ёкои А., Нонака С., Сайка Х., Квон Й.И., Осакабе К., Токи С. (декабрь 2012 г.). «Подавление Ku70/80 или Lig4 приводит к снижению стабильной трансформации и усилению гомологичной рекомбинации в рисе» . Новый фитолог . 196 (4): 1048–59. дои : 10.1111/j.1469-8137.2012.04350.x . ПМЦ 3532656 . ПМИД 23050791 .
- ^ Ци Ю, Чжан Ю, Чжан Ф, Баллер Дж. А., Клеланд С. С., Рю Ю, Старкер К. Г., Войтас Д. Ф. (март 2013 г.). «Увеличение частоты сайт-специфического мутагенеза и нацеливания на гены у Arabidopsis путем манипулирования путями репарации ДНК» . Геномные исследования . 23 (3): 547–54. дои : 10.1101/гр.145557.112 . ПМЦ 3589543 . ПМИД 23282329 .
- ^ Казда А., Зеллингер Б., Рёсслер М., Дербовен Е., Кусенда Б., Риха К. (август 2012 г.). «Защита концов хромосомы с помощью теломеров с тупыми концами» . Гены и развитие . 26 (15): 1703–13. дои : 10.1101/gad.194944.112 . ПМЦ 3418588 . ПМИД 22810623 .
- ^ Валухова С., Фулнечек Дж., Прокоп З., Столт-Бергнер П., Янушкова Е., Хофр С., Риха К. (июнь 2017 г.). «Защита теломер Arabidopsis с тупыми концами опосредована физической ассоциацией с Ku-гетеродимером» . Растительная клетка . 29 (6): 1533–1545. дои : 10.1105/tpc.17.00064 . ПМК 5502450 . ПМИД 28584163 .
- ^ Питчер Р.С., Бриссетт, Северная Каролина, Доэрти Эй.Дж. (2007). «Номологическое соединение концов у бактерий: микробная перспектива». Ежегодный обзор микробиологии . 61 (1). Годовые обзоры : 259–82. дои : 10.1146/annurev.micro.61.080706.093354 . ПМИД 17506672 .
- ^ Шарда М., Бадринараянан А., Сешасаи А.С. (декабрь 2020 г.). «Эволюционный и сравнительный анализ бактериальной репарации негомологичных концевых соединений» . Геномная биология и эволюция . 12 (12): 2450–2466. дои : 10.1093/gbe/evaa223 . ПМЦ 7719229 . ПМИД 33078828 .
- ^ Дайнан В.С., Ю С. (апрель 1998 г.). «Взаимодействие белка Ku и каталитической субъединицы ДНК-зависимой протеинкиназы с нуклеиновыми кислотами» . Исследования нуклеиновых кислот . 26 (7): 1551–9. дои : 10.1093/нар/26.7.1551 . ПМК 147477 . ПМИД 9512523 .