хроматофор

Хроматофоры — это клетки, производящие цвет, многие из которых представляют собой пигментсодержащие клетки или группы клеток, обнаруженные у широкого круга животных, включая земноводных , рыб , рептилий , ракообразных и головоногих моллюсков . У млекопитающих и птиц , напротив, есть класс клеток, называемых меланоцитами, отвечающими за окраску .
Хроматофоры в значительной степени ответственны за формирование цвета кожи и глаз у экзотермических животных и генерируются в нервном гребне во время эмбрионального развития . Зрелые хроматофоры группируются в подклассы в зависимости от их цвета в белом свете: ксантофоры (желтые), эритрофоры (красные), иридофоры ( отражающие / переливающиеся ), лейкофоры (белые), меланофоры (черные/коричневые) и цианофоры (синие). Хотя большинство хроматофоров содержат пигменты, поглощающие свет определенной длины, цвет лейкофоров и иридофоров определяется их соответствующими свойствами рассеяния и оптической интерференции.
Некоторые виды могут быстро менять цвет с помощью механизмов, которые перемещают пигмент и переориентируют отражающие пластинки внутри хроматофоров. Этот процесс, часто используемый как разновидность камуфляжа , называется физиологическим изменением цвета или метахрозом . [ 1 ] Головоногие моллюски, такие как осьминоги , для достижения этой цели имеют сложные хроматофорные органы, контролируемые мышцами, тогда как позвоночные, такие как хамелеоны, генерируют аналогичный эффект посредством передачи сигналов клетками . Такие сигналы могут быть гормонами или нейротрансмиттерами и могут быть инициированы изменениями настроения, температуры, стресса или видимыми изменениями в местной среде. [ нужна ссылка ] Хроматофоры изучаются учеными для понимания болезней человека и в качестве инструмента для разработки лекарств .
Человеческое открытие
[ редактировать ]Аристотель упомянул способность осьминога менять цвет как для маскировки , так и для подачи сигналов в своей «Истории животных» (около 4 века до н.э.): [ 2 ]
Осьминог... ищет свою добычу, меняя свой цвет так, чтобы сделать его похожим на цвет соседних с ним камней; он делает то же самое, когда встревожен.
Джозуэ Санджованни был первым, кто описал беспозвоночных пигментсодержащие клетки как хромофоро в итальянском научном журнале в 1819 году. [ 3 ]
Чарльз Дарвин описал способность каракатиц менять цвет в книге «Путешествие «Бигля»» (1860 г.): [ 4 ]
Эти животные также избегают обнаружения благодаря необычной, подобной хамелеону, способности менять свой цвет. По-видимому, они меняют свои оттенки в зависимости от характера грунта, по которому они проходят: на глубокой воде их общий оттенок был коричневато-фиолетовым, но когда их помещали на сушу или на мелководье, этот темный оттенок менялся на один из желтовато-зеленый. Цвет, при более внимательном рассмотрении, оказался французско-серым с многочисленными мелкими пятнами ярко-желтого цвета: первые из них различались по интенсивности; последний совершенно исчезал и по очереди появлялся снова. Эти изменения происходили таким образом, что облака, оттенки которых варьировались от гиацинтово-красного до каштаново-коричневого, постоянно проходили над телом. Любая часть, подвергнутая легкому воздействию гальванизма, становилась почти черной: аналогичный эффект, но в меньшей степени, давало расчесывание кожи иглой. Говорят, что эти облака, или покраснения, как их можно назвать, возникают в результате поочередного расширения и сжатия мельчайших пузырьков, содержащих жидкости разного цвета.
Классификация хроматофора
[ редактировать ]
Термин хроматофор Санджованни был принят (вслед за хромофоро ) как название пигментоносных клеток, происходящих из нервного гребня хладнокровных позвоночных и головоногих моллюсков. Само слово происходит от греческих слов chrōma ( χρῶμα ), означающих «цвет», и phoros ( φόρος ), означающих «несущий». Напротив, слово хроматоцит ( kytos ( κύτος ), что означает «клетка»), было принято для обозначения клеток, ответственных за цвет, обнаруженных у птиц и млекопитающих. только один такой тип клеток — меланоциты У этих животных был идентифицирован .
Лишь в 1960-х годах хроматофоры стали достаточно хорошо изучены, чтобы можно было классифицировать их по внешнему виду. Эта система классификации сохраняется и по сей день, хотя биохимия пигментов может быть более полезной для научного понимания того, как функционируют клетки. [ 5 ]
Молекулы, производящие цвет, делятся на два отдельных класса: биохромы и структурные цвета или «схемохромы». [ 6 ] Биохромы включают настоящие пигменты, такие как каротиноиды и птеридины . Эти пигменты избирательно поглощают части спектра видимого света , который составляет белый свет, позволяя при этом другим длинам волн достигать глаза наблюдателя. Структурные цвета создаются в результате различных комбинаций дифракции, отражения или рассеяния света от структур размером около четверти длины волны света. Многие такие структуры интерферируют с некоторыми длинами волн (цветами) света и пропускают другие просто из-за своего масштаба, поэтому они часто создают переливчатость , создавая разные цвета, если смотреть с разных сторон. [ нужна ссылка ]
В то время как все хроматофоры содержат пигменты или отражающие структуры (за исключением случаев мутации , как при альбинизме ), не все содержащие пигмент клетки являются хроматофорами. Гем , например, является биохромом, ответственным за красный цвет крови. Он содержится преимущественно в красных кровяных тельцах (эритроцитах), которые образуются в костном мозге на протяжении всей жизни организма, а не образуются в ходе эмбрионального развития. Поэтому эритроциты не относятся к хроматофорам. [ нужна ссылка ]
Ксантофоры и эритрофоры
[ редактировать ]Хроматофоры, содержащие большое количество желтых птеридиновых пигментов, называются ксантофорами; те, которые содержат преимущественно красные / оранжевые каротиноиды, называются эритрофорами. [ 5 ] Однако иногда в одной клетке обнаруживаются везикулы, содержащие птеридин и каротиноиды, и в этом случае общая окраска зависит от соотношения красного и желтого пигментов. [ 7 ] Поэтому различие между этими типами хроматофоров не всегда четкое.
Большинство хроматофоров могут генерировать птеридины из гуанозинтрифосфата , но ксантофоры, по-видимому, имеют дополнительные биохимические пути, позволяющие им накапливать желтый пигмент. Напротив, каротиноиды метаболизируются и транспортируются в эритрофоры. Впервые это было продемонстрировано при выращивании обычно зеленых лягушек на диете из каротина с ограниченным содержанием сверчков . Отсутствие каротина в рационе лягушек означало, что красно-оранжевый «фильтр» цвета каротиноидов не присутствовал в их эритрофофах. Из-за этого лягушки выглядели синими, а не зелеными. [ 8 ]
Иридофоры и лейкофоры
[ редактировать ]
Иридофоры, иногда также называемые гуанофорами, представляют собой хроматофоры, которые отражают свет с помощью пластинок кристаллических хемохромов, изготовленных из гуанина . [ 9 ] При освещении они создают переливающиеся цвета из-за конструктивной интерференции света. Иридофоры рыб обычно представляют собой сложенные друг на друга пластинки гуанина, разделенные слоями цитоплазмы, образующие микроскопические одномерные зеркала Брэгга . Как ориентация, так и оптическая толщина хемохрома определяют природу наблюдаемого цвета. [ 10 ] Используя биохромы в качестве цветных фильтров, иридофоры создают оптический эффект, известный как рассеяние Тиндаля или Рэлея , создавая ярко- синие или зеленые цвета. [ 11 ]
Родственный тип хроматофора — лейкофор — обнаружен у некоторых рыб, в частности у Tapetum lucidum . Как иридофоры, они используют кристаллические пурины (часто гуанин) для отражения света. В отличие от иридофоров, лейкофоры имеют более организованные кристаллы, уменьшающие дифракцию. При наличии источника белого света они излучают белый блеск. Как и в случае с ксантофорами и эритрофорами, у рыб различие между иридофорами и лейкофорами не всегда очевидно, но, как правило, считается, что иридофоры создают радужные или металлические цвета , тогда как лейкофоры производят отражающие белые оттенки. [ 11 ]
Меланофоры
[ редактировать ]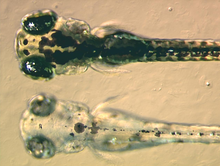
Меланофоры содержат эумеланин , тип меланина , который кажется черным или темно- коричневым из-за его светопоглощающих свойств. Он упакован в пузырьки, называемые меланосомами, и распределяется по всей клетке. Эумеланин образуется из тирозина в результате серии катализируемых химических реакций. Это сложное химическое вещество, содержащее звенья дигидроксииндола и дигидроксииндол-2- карбоновой кислоты с некоторыми пиррольными кольцами. [ 12 ] Ключевым ферментом синтеза меланина является тирозиназа . Когда этот белок дефектен, меланин не вырабатывается, что приводит к определенным типам альбинизма. У некоторых видов земноводных наряду с эумеланином имеются и другие пигменты. Например, в меланофорах филломедузиновых лягушек был идентифицирован новый пигмент глубокого (винного) красного цвета . [ 13 ] Некоторые виды ящериц-анолов, такие как Anolis grahami , используют меланоциты в ответ на определенные сигналы и гормональные изменения и способны приобретать цвета от ярко-синего, коричневого и черного. Впоследствии он был идентифицирован как птерорродин птеридина , димер , который накапливается вокруг ядра эумеланина, и он также присутствует у различных видов древесных лягушек из Австралии и Папуа-Новой Гвинеи . Хотя вполне вероятно, что другие менее изученные виды имеют сложные пигменты меланофоров, тем не менее верно, что большинство меланофоров, изученных на сегодняшний день, действительно содержат исключительно эумеланин. [ 14 ]
У людей есть только один класс пигментных клеток, эквивалент меланофоров у млекопитающих, отвечающий за цвет кожи, волос и глаз. По этой причине, а также потому, что большое количество и контрастный цвет клеток обычно облегчают их визуализацию, меланофоры на сегодняшний день являются наиболее широко изученными хроматофорами. Однако существуют различия между биологией меланофоров и меланоцитов . Помимо эумеланина, меланоциты могут генерировать желтый/красный пигмент, называемый феомеланин . [ нужна ссылка ]

Цианофоры
[ редактировать ]Почти весь яркий синий цвет у животных и растений создается структурной окраской, а не пигментами. Однако некоторые виды Synchiropus splendidus обладают в клетках пузырьками цианового биохрома неизвестной химической структуры, называемыми цианофорами. [ 11 ] Хотя они кажутся необычными в своем ограниченном таксономическом диапазоне, цианофоры (а также другие необычные типы хроматофоров) могут встречаться и у других рыб и амфибий. Например, ярко окрашенные хроматофоры с неопределенными пигментами обнаружены как у ядовитых древолазов, так и у стеклянных лягушек . [ 15 ] и атипичные дихроматические хроматофоры, названные эритроиридофоры , были описаны у Pseudochromis diadema . [ 16 ]
Транслокация пигмента
[ редактировать ]
Многие виды способны перемещать пигмент внутри своих хроматофоров, что приводит к явному изменению цвета тела. Этот процесс, известный как физиологическое изменение цвета , наиболее широко изучен у меланофоров, поскольку меланин является самым темным и наиболее видимым пигментом. У большинства видов с относительно тонкой дермой дермальные меланофоры имеют тенденцию быть плоскими и покрывать большую площадь поверхности. Однако у животных с толстым кожным слоем, таких как взрослые рептилии, дермальные меланофоры часто образуют трехмерные единицы с другими хроматофорами. Эти дермальные хроматофорные единицы (DCU) состоят из самого верхнего слоя ксантофоров или эритрофоров, затем слоя иридофора и, наконец, корзинчатого меланофорного слоя с отростками, покрывающими иридофоры. [ 17 ]
Оба типа меланофоров играют важную роль в физиологическом изменении цвета. Плоские дермальные меланофоры часто накладываются на другие хроматофоры, поэтому, когда пигмент распределяется по клетке, кожа кажется темной. Когда пигмент агрегируется по направлению к центру клетки, пигменты других хроматофоров подвергаются воздействию света, и кожа приобретает их оттенок. Аналогичным образом, после агрегации меланина в DCU кожа кажется зеленой из-за ксантофорной (желтой) фильтрации рассеянного света от слоя иридофора. При дисперсии меланина свет больше не рассеивается, и кожа кажется темной. Поскольку другие биохроматические хроматофоры также способны к транслокации пигментов, животные с несколькими типами хроматофоров могут генерировать впечатляющий набор цветов кожи, эффективно используя эффект разделения. [ 18 ] [ 19 ]

Контроль и механизм быстрой транслокации пигментов хорошо изучены у ряда различных видов, в частности у амфибий и костистых рыб. [ 11 ] [ 20 ] Было продемонстрировано, что этот процесс может находиться под гормональным или нейронным контролем, или под тем и другим, и для многих видов костистых рыб известно, что хроматофоры могут напрямую реагировать на стимулы окружающей среды, такие как видимый свет, УФ-излучение, температура, pH, химические вещества и т. д. [ 21 ] Нейрохимические вещества, которые, как известно, перемещают пигмент, включают норадреналин через его рецептор на поверхности меланофоров. [ 22 ] Основными гормонами, участвующими в регуляции транслокации, по-видимому, являются меланокортины , мелатонин и меланин-концентрирующий гормон (MCH), которые производятся главным образом в гипофизе, шишковидной железе и гипоталамусе соответственно. Эти гормоны также могут вырабатываться паракринным образом клетками кожи. Было показано, что на поверхности меланофора гормоны активируют специфические рецепторы, связанные с G-белком, которые, в свою очередь, передают сигнал в клетку. Меланокортины приводят к дисперсии пигмента, а мелатонин и MCH – к агрегации. [ 23 ]
У рыб идентифицированы многочисленные рецепторы меланокортина, MCH и мелатонина. [ 24 ] и лягушки, [ 25 ] включая гомолог MC1R , [ 26 ] рецептор меланокортина, который, как известно, регулирует цвет кожи и волос у людей. [ 27 ] Было продемонстрировано, что MC1R необходим рыбкам данио для диспергирования меланина. [ 28 ] внутри клетки циклический аденозинмонофосфат Было показано, что (цАМФ) является важным вторичным посредником транслокации пигмента. По механизму, который еще не до конца понятен, цАМФ влияет на другие белки, такие как протеинкиназа А, приводя в движение молекулярные моторы, переносящие пигментсодержащие пузырьки как по микротрубочкам , так и по микрофиламентам . [ 29 ] [ 30 ] [ 31 ]
Фоновая адаптация
[ редактировать ]
Большинство рыб, рептилий и земноводных претерпевают ограниченное физиологическое изменение цвета в ответ на изменение окружающей среды. Этот тип камуфляжа, известный как фоновая адаптация , чаще всего проявляется как небольшое затемнение или осветление тона кожи, приблизительно имитирующее оттенок непосредственной среды. Было продемонстрировано, что процесс фоновой адаптации зависит от зрения (похоже, что животное должно иметь возможность видеть окружающую среду, чтобы адаптироваться к ней). [ 32 ] и что транслокация меланина в меланофорах является основным фактором изменения цвета. [ 23 ] Некоторые животные, такие как хамелеоны и анолисы , имеют высокоразвитую фоновую адаптационную реакцию, способную очень быстро генерировать множество различных цветов. [ 33 ] Они адаптировали способность менять цвет в зависимости от температуры, настроения, уровня стресса и социальных сигналов, а не просто имитировать окружающую среду.
Разработка
[ редактировать ]
Во время эмбрионального развития позвоночных хроматофоры являются одним из многих типов клеток, образующихся в нервном гребне , парной полоске клеток, возникающей на краях нервной трубки . Эти клетки обладают способностью мигрировать на большие расстояния, что позволяет хроматофорам заселять многие органы тела, включая кожу, глаза, уши и мозг. Было обнаружено, что меланофоры и иридофоры рыб содержат регуляторные белки гладких мышц [кальпонин] и кальдесмон . [ 34 ] Покидая нервный гребень волнообразно, хроматофоры идут либо дорсолатеральным путем через дерму, попадая в эктодерму через небольшие отверстия в базальной пластинке , либо вентромедиальным путем между сомитами и нервной трубкой. Исключение составляют меланофоры пигментного эпителия сетчатки глаза. Они не происходят из нервного гребня. Вместо этого выпячивание нервной трубки образует глазной бокал , который, в свою очередь, образует сетчатку . [ нужна ссылка ]
Когда и как мультипотентные клетки-предшественники хроматофоров (называемые хроматобластами ) развиваются в свои дочерние подтипы, является областью текущих исследований. Известно, например, что у эмбрионов рыбок данио уже через 3 дня после оплодотворения присутствуют все классы клеток, обнаруженные у взрослых рыб — меланофоры, ксантофоры и иридофоры. Исследования с использованием мутантных рыб показали, что факторы транскрипции , такие как kit , sox10 и mitf , важны для контроля дифференцировки хроматофоров. [ 35 ] Если эти белки дефектны, хроматофоры могут частично или полностью отсутствовать, что приводит к лейцистическому расстройству.
Практическое применение
[ редактировать ]Хроматофоры иногда используются в прикладных исследованиях. Например, личинки рыбок данио используются для изучения того, как хроматофоры организуются и взаимодействуют, чтобы точно генерировать регулярный узор из горизонтальных полос, который наблюдается у взрослых рыб. [ 36 ] Это рассматривается как полезная модельная система для понимания закономерностей в области эволюционной биологии развития . Биология хроматофоров также использовалась для моделирования состояния или болезней человека, включая меланому и альбинизм. Недавно было показано, что ген, ответственный за меланофор-специфичный штамм золотых рыбок данио, Slc24a5 , имеет человеческий эквивалент, который сильно коррелирует с цветом кожи . [ 37 ]
Хроматофоры также используются в качестве биомаркера слепоты у хладнокровных видов, поскольку животные с определенными дефектами зрения не могут адаптироваться к световой среде. [ 32 ] Считается, что человеческие гомологи рецепторов, которые опосредуют транслокацию пигментов в меланофорах, участвуют в таких процессах, как подавление аппетита и загар , что делает их привлекательными мишенями для лекарств . [ 26 ] Поэтому фармацевтические компании разработали биологический анализ для быстрой идентификации потенциальных биологически активных соединений с использованием меланофоров африканской когтистой лягушки . [ 38 ] Другие ученые разработали методы использования меланофоров в качестве биосенсоров . [ 39 ] и для быстрого выявления заболеваний (на основе открытия того, что коклюшный токсин блокирует агрегацию пигментов в меланофорах рыб). [ 40 ] Были предложены потенциальные военные применения изменения цвета, опосредованного хроматофором, главным образом в качестве типа активного камуфляжа , может , который, как и у каракатиц делать объекты почти невидимыми. [ 41 ] [ 42 ]
Хроматофоры головоногих моллюсков
[ редактировать ]
Колеоидные головоногие моллюски (включая осьминогов, кальмаров и каракатиц ) имеют сложные многоклеточные органы, которые они используют для быстрого изменения цвета, создавая большое разнообразие ярких цветов и узоров. Каждая хроматофорная единица состоит из одной хроматофорной клетки и множества мышечных, нервных, глиальных и оболочечных клеток. [ 43 ] Внутри хроматофорной клетки пигментные гранулы заключены в эластичный мешочек, называемый цитоэластическим мешочком. Чтобы изменить цвет, животное искажает форму или размер саккулюса за счет мышечного сокращения, изменяя его прозрачность , отражательную способность или непрозрачность . Это отличается от механизма, используемого у рыб, амфибий и рептилий, тем, что форма саккулюса изменяется, а не перемещается пигментные пузырьки внутри клетки. Однако аналогичный эффект достигается.
Осьминоги и большинство каракатиц [ 44 ] могут управлять хроматофорами в сложных, волнообразных хроматических проявлениях, что приводит к множеству быстро меняющихся цветовых схем. Считается, что нервы, управляющие хроматофорами, расположены в мозге по схеме, изоморфной хроматофорам, каждый из которых ими управляет. Это означает, что паттерн изменения цвета функционально соответствует паттерну активации нейронов . Это может объяснить, почему, когда нейроны активируются в итеративном сигнальном каскаде, можно наблюдать волны изменения цвета. [ 45 ] Подобно хамелеонам, головоногие моллюски используют физиологическое изменение цвета для социального взаимодействия . Они также являются одними из самых искусных в маскировке, имея способность с удивительной точностью соответствовать как распределению цвета, так и текстуре окружающей среды.
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ Скотт М. Бобак и Линн М. Сифферман (2010). «Изменение цвета и изменение цвета островных и материковых удавов ( Удав )». Журнал герпетологии . 44 (4): 506–515. дои : 10.1670/09-026.1 . S2CID 53634890 .
- ^ Аристотель. История животных . IX, 622а: 2-10. Около 400 г. до н.э. Цитируется у Лусианы Боррелли, Франчески Герарди , Грациано Фиорито. Каталог рисунков тела головоногих . Издательство Firenze University Press, 2006. Аннотация. Архивировано 6 февраля 2018 г. в книгах Wayback Machine Google.
- ^ Санджованни, Дж. (1819). «Описание конкретной экспансивно-дермоидной хромофорной системы органов и явлений, которые она вызывает, обнаруженных у головоногих моллюсков». Г. Энциклопедика Неаполя . 9 :1–13.
- ^ Дарвин, Чарльз (1860). «Глава 1. Повадки морского слизняка и каракатицы». Журнал исследований естественной истории и геологии стран, посещенных во время кругосветного путешествия корабля HMS «Бигль» под командованием капитана Фитца Роя, RN . Джон Мюррей, Лондон. п. 7.
- ^ Перейти обратно: а б Баньяра, Дж.Т. (1966). Цитология и цитофизиология немеланофорных пигментных клеток . Международный обзор цитологии. Том. 20. стр. 173–205. дои : 10.1016/S0074-7696(08)60801-3 . ISBN 978-0-12-364320-9 . ПМИД 5337298 .
- ^ Фокс, ДЛ. Биохромы животных и структурные цвета: физические, химические, распределительные и физиологические особенности цветных тел в животном мире. Калифорнийский университет Press, Беркли, 1976. ISBN 0-520-02347-1
- ^ Мацумото, Дж (1965). «Исследование тонкой структуры и цитохимических свойств эритрофоров меченосца Xiphophorus helleri с особым упором на их пигментные гранулы (птериносомы)» . J Клеточная Биол . 27 (3): 493–504. дои : 10.1083/jcb.27.3.493 . ПМК 2106771 . ПМИД 5885426 .
- ^ Баньяра Дж.Т. Сравнительная анатомия и физиология пигментных клеток в тканях немлекопитающих. В: Пигментная система: физиология и патофизиология , Oxford University Press , 1998. ISBN 0-19-509861-7
- ^ Тейлор, Джей Ди. (1969). «Влияние интермедина на ультраструктуру иридофоров амфибий». Ген Комп Эндокринол . 12 (3): 405–16. дои : 10.1016/0016-6480(69)90157-9 . ПМИД 5769930 .
- ^ Моррисон, РЛ. (1995). «Метод просвечивающей электронной микроскопии (ТЕМ) для определения структурных цветов, отражаемых иридофорами ящерицы». Ресурс пигментных клеток . 8 (1): 28–36. дои : 10.1111/j.1600-0749.1995.tb00771.x . ПМИД 7792252 .
- ^ Перейти обратно: а б с д Фуджи, Р. (2000). «Регуляция подвижной активности хроматофоров рыб». Ресурс пигментных клеток . 13 (5): 300–19. дои : 10.1034/j.1600-0749.2000.130502.x . ПМИД 11041206 .
- ^ Ито, С; Вакамацу, К. (2003). «Количественный анализ эумеланина и феомеланина у людей, мышей и других животных: сравнительный обзор». Ресурс пигментных клеток . 16 (5): 523–31. дои : 10.1034/j.1600-0749.2003.00072.x . ПМИД 12950732 .
- ^ Баньяра, Джей Ти; Тейлор, доктор медицинских наук; Прота, Г (1973). «Изменения цвета, необычные меланосомы и новый пигмент листовых лягушек». Наука . 182 (4116): 1034–5. Бибкод : 1973Sci...182.1034B . дои : 10.1126/science.182.4116.1034 . ПМИД 4748673 . S2CID 33900017 .
- ^ Баньяра, Дж. Т. (2003). «Загадки птерородина, красного меланосомного пигмента древесных лягушек». Исследование пигментных клеток . 16 (5): 510–516. дои : 10.1034/j.1600-0749.2003.00075.x . ПМИД 12950730 .
- ^ Швальм, Пенсильвания; Старретт, PH; МакДиармид, RW (1977). «Инфракрасное отражение у неотропических лягушек, сидящих на листьях». Наука . 196 (4295): 1225–7. Бибкод : 1977Sci...196.1225S . дои : 10.1126/science.860137 . ПМИД 860137 . S2CID 45014819 .
- ^ Года М., Охата М., Икома Х., Фудзиёси Ю., Сугимото М., Фуджи Р. (2011). «Покровная красновато-фиолетовая окраска обусловлена новыми дихроматическими хроматофорами у костистых рыб Pseudochromis diadema». Пигментно-клеточная меланома Res . 24 (4): 614–7. дои : 10.1111/j.1755-148X.2011.00861.x . ПМИД 21501419 . S2CID 3299089 .
- ^ Баньяра, Джей Ти; Тейлор, доктор медицинских наук; Хэдли, Мэн (1968). «Кожный хроматофорный аппарат» . J Клеточная Биол . 38 (1): 67–79. дои : 10.1083/jcb.38.1.67 . ПМК 2107474 . ПМИД 5691979 .
- ^ Палаццо, RE; Линч, Ти Джей; Ло, С.Дж.; Тейлор, доктор медицинских наук; Чен, Т.Т. (1989). «Перестройки птериносом и цитоскелета, сопровождающие дисперсию пигментов в ксантофорах золотых рыбок». Цитоскелет клеточного мотора . 13 (1): 9–20. дои : 10.1002/см.970130103 . ПМИД 2543509 .
- ^ Поррас, МГ; Де Луф, А; Брейер, М; Аречига, Х (2003). «Прокамбарус Кларкии». Пептиды . 24 (10): 1581–9. doi : 10.1016/j.peptides.2003.08.016 . ПМИД 14706537 . S2CID 2593619 .
- ^ Дьякон, SW; Серпинская А.С.; Воган, PS; Лопес Фанаррага, М; Вернос, я; Воган, Коннектикут; Гельфанд, В.И. (2003). «Динактин необходим для двунаправленного транспорта органелл» . Журнал клеточной биологии . 160 (3): 297–301. дои : 10.1083/jcb.200210066 . ПМК 2172679 . ПМИД 12551954 .
- ^ Мейер-Рохов, В.Б. (2001). «Хроматофоры рыб как сенсоры раздражителей окружающей среды». В Капур Б.Г. и Хара Т.Дж. (ред.). Сенсорная биология челюстных рыб . Издательство Science Publishers Энфилд (Нью-Хэмпшир), США. стр. 317–334.
- ^ Аспенгрен, С; Скельд, Х.Н.; Кирога, Дж; Мортенссон, Л; Валлин, М. (2003). «Норадреналин- и мелатонин-опосредованная регуляция агрегации пигментов в меланофорах рыб». Ресурс пигментных клеток . 16 (1): 59–64. дои : 10.1034/j.1600-0749.2003.00003.x . ПМИД 12519126 .
- ^ Перейти обратно: а б Логан, Д.В.; Берн, Сан-Франциско; Джексон, Ай-Джей (2006). «Регуляция пигментации меланофоров рыбок данио». Исследование пигментных клеток . 19 (3): 206–213. дои : 10.1111/j.1600-0749.2006.00307.x . ПМИД 16704454 .
- ^ Логан, Д.В.; Брайсон-Ричардсон, Р.Дж.; Тейлор, MS; Карри, П; Джексон, Эй Джей (2003). «Характеристика последовательностей рецепторов меланокортина костистых рыб». Энн, Нью-Йоркская академия наук . 994 (1): 319–30. Бибкод : 2003NYASA.994..319L . дои : 10.1111/j.1749-6632.2003.tb03196.x . ПМИД 12851332 . S2CID 37853710 .
- ^ Сагден, Д; Дэвидсон, К; Хаф, К.А.; Тех, МТ (2004). «Мелатонин, рецепторы мелатонина и меланофоры: трогательная история» . Ресурс пигментных клеток . 17 (5): 454–60. дои : 10.1111/j.1600-0749.2004.00185.x . ПМИД 15357831 .
- ^ Перейти обратно: а б Логан, Д.В.; Брайсон-Ричардсон, Р.Дж.; Паган, Кентукки; Тейлор, MS; Карри, PD; Джексон, Эй Джей (2003). «Структура и эволюция рецепторов меланокортина и MCH у рыб и млекопитающих». Геномика . 81 (2): 184–91. дои : 10.1016/S0888-7543(02)00037-X . ПМИД 12620396 .
- ^ Вальверде, П; Хили, Э; Джексон, я; Рис, Дж.Л.; Тоди, Эй Джей (1995). «Варианты гена рецептора меланоцитстимулирующего гормона связаны с рыжими волосами и светлой кожей у людей». Нат Жене . 11 (3): 328–30. дои : 10.1038/ng1195-328 . ПМИД 7581459 . S2CID 7980311 .
- ^ Ричардсон, Дж; Лундегаард, PR; Рейнольдс, Нидерланды; Дорин, младший; Портеус, диджей; Джексон, Ай-Джей; Паттон, Э.Э. (2008). «Регуляция пути mc1r дисперсии меланосом рыбок данио». Рыбка данио . 5 (4): 289–95. дои : 10.1089/zeb.2008.0541 . ПМИД 19133827 .
- ^ Снайдер, Дж; Лин, Ф; Захеди, Н; Родионов В; Ю, СС; Гросс, СП (2004). «Внутриклеточный транспорт на основе актина: как далеко вы продвинетесь, зависит от того, как часто вы переключаетесь» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (36): 13204–9. Бибкод : 2004PNAS..10113204S . дои : 10.1073/pnas.0403092101 . ПМК 516548 . ПМИД 15331778 .
- ^ Родионов В.И.; Надежда, Эй Джей; Свиткина, ТМ; Борисы, Г.Г. (1998). «Функциональная координация подвижности микротрубочек и актина в меланофорах» . Современная биология . 8 (3): 165–8. Бибкод : 1998CBio....8..165R . дои : 10.1016/S0960-9822(98)70064-8 . ПМИД 9443917 .
- ^ Кашина А.С.; Семенова, ИВ; Иванов, П.А.; Потехина Е.С.; Заляпин И.; Родионов, В.И. (2004). «Протеинкиназа А, регулирующая внутриклеточный транспорт, образует комплексы с молекулярными моторами на органеллах» . Современная биология . 14 (20): 1877–81. Бибкод : 2004CBio...14.1877K . дои : 10.1016/j.cub.2004.10.003 . ПМИД 15498498 .
- ^ Перейти обратно: а б Нойхаусс, Южная Каролина. (2003). «Поведенческие генетические подходы к развитию и функционированию зрительной системы рыбок данио» (PDF) . Дж. Нейробиол . 54 (1): 148–60. дои : 10.1002/neu.10165 . ПМИД 12486702 . [ мертвая ссылка ]
- ^ «Порошки-хамелеоны» . 13 июня 2023 г. Проверено 8 июня 2024 г.
- ^ Мейер-Рохов, В.Б.; Ройуэла, М (2002). «Кальпонин, кальдесмон и хроматофоры: связь гладких мышц». Микроск. Рез. Тех . 58 (6): 504–513. дои : 10.1002/jemt.10169 . ПМИД 12242708 . S2CID 30194336 .
- ^ Келш, Р.Н.; Шмид, Б; Эйзен, Дж. С. (2000). «Генетический анализ развития меланофоров у эмбрионов рыбок данио» . Дев Биол . 225 (2): 277–93. дои : 10.1006/dbio.2000.9840 . ПМИД 10985850 .
- ^ Келш, Р.Н. (2004). «Генетика и эволюция пигментных пятен у рыб» . Исследование пигментных клеток . 17 (4): 326–36. дои : 10.1111/j.1600-0749.2004.00174.x . ПМИД 15250934 . S2CID 15112204 .
- ^ Ламасон, РЛ; Мохидин, Массачусетс; Мест, младший; Вонг, AC; Нортон, Х.Л.; Арос, MC; Юринец, MJ; Мао, X; и др. (2005). «SLC24A5, предполагаемый катионообменник, влияет на пигментацию у рыбок данио и людей». Наука . 310 (5755): 1782–6. Бибкод : 2005Sci...310.1782L . дои : 10.1126/science.1116238 . ПМИД 16357253 . S2CID 2245002 .
- ^ Джаявикрем, СК; Саулс, Х; Болио, Н.; Руан, Дж; Мойер, М; Беркхарт, В; Маррон, Б; Римеле, Т; Шаффер, Дж (1999). «Использование клеточного анализа в формате газона для быстрого скрининга библиотеки пептидов на основе 442 368 шариков». J Фармаколические токсиколовые методы . 42 (4): 189–97. дои : 10.1016/S1056-8719(00)00083-6 . ПМИД 11033434 .
- ^ Андерссон, ТП; Филиппини, Д; Суска, А; Йоханссон, ТЛ; Свенссон, СП; Лундстрём, я (2005). «Меланофоры лягушки, культивированные на флуоресцентных микрогранулах: биосенсорство на основе биомимики». Биосенс Биоэлектрон . 21 (1): 111–20. дои : 10.1016/j.bios.2004.08.043 . ПМИД 15967358 .
- ^ Карлссон, Дж. О.; Андерссон, Р.Г.; Аскелёф, П; Элвинг, Х; Гранстрем, М; Грундстрем, Н.; Лундстрём, я; Оман, Л. (1991). «Реакция агрегации меланофоров изолированной рыбьей чешуи: очень быстрая и чувствительная диагностика коклюша» . FEMS Microbiol Lett . 66 (2): 169–75. дои : 10.1111/j.1574-6968.1991.tb04860.x . ПМИД 1936946 .
- ^ Хансфорд, Дэйв (6 августа 2008 г.). «Каракатицы меняют цвет и форму, чтобы ускользнуть от хищников» . Национальные географические новости . Веллингтон, Новая Зеландия. Архивировано из оригинала 10 августа 2008 года.
[...] вместо этого каракатицы полагались на невидимость, талант, который может найти применение в человеческих технологиях. Норман сказал, что военные проявили интерес к камуфляжу в виде каракатиц с целью однажды включить подобные механизмы в солдатскую униформу.
- ^ Ли И. Нанотрубки для обработки шумных сигналов . Кандидатская диссертация . 2005 г.; Университет Южной Калифорнии .
- ^ Клони, РА; Флори, Э. (1968). «Ультраструктура хроматофорных органов головоногих». Журнал клеточных исследований и микроскопической анатомии . 89 (2): 250–80. дои : 10.1007/BF00347297 . ПМИД 5700268 . S2CID 26566732 .
- ^ Лаан, А; Гутник, Т; Куба, МЮ; Лоран, Дж. (2014). «Поведенческий анализ бегущих волн каракатиц и его значение для нейронного контроля» . Современная биология . 24 (15): 1737–1742. Бибкод : 2014CBio...24.1737L . дои : 10.1016/j.cub.2014.06.027 . ПМИД 25042589 .
- ^ Демский, Л.С. (1992). «Хроматофорные системы костистых и головоногих моллюсков: многоуровневый анализ конвергентных систем». Мозг, поведение и эволюция . 40 (2–3): 141–56. дои : 10.1159/000113909 . ПМИД 1422807 .
Внешние ссылки
[ редактировать ]- «Палитра природы - как животные производят цвет» (PDF) . (1,20 МБ)
- Видеозапись адаптации фона осьминога
- Видеозапись формирования рисунка хроматофора кальмара
- Веб-проект «Древо жизни»: хроматофоры головоногих моллюсков
Зрение у животных |
|---|

головоногих моллюсков Анатомия |
|---|


