Центромер

Centromere вместе во время связывает пару сестринских хроматидов деления клеток. Эта сужаемая область хромосомы соединяет сестринские хроматиды, создавая короткую руку (P) и длинную руку (Q) на хроматидах. Во время митоза прикрепляются волокна шпинделя к центромеру через кинетохор .
Физическая роль центромеры состоит в том, чтобы выступать в качестве места сборки кинетохоров - очень сложной мультипротеиновой структуры, которая отвечает за фактические события сегрегации хромосомы - IE, связывающие микротрубочки и передача сигналов с механическим обеспечением клеточного цикла, когда все хромосомы приняли правильные Прикрепления к шпинделю , так что для деления клеток можно было бы добраться до завершения, и для клетки въезжают анафазу .
В целом, есть два типа центромеров. «Центромеры точек» связываются со специфическими белками , которые распознают определенные ДНК последовательности с высокой эффективностью. Любая часть ДНК с точечной последовательности ДНК Centromere на ней обычно образует центромер, если присутствует у соответствующих видов. Лучше всего охарактеризованные центромеры - это центромеры, которые являются начинающими дрожжами, Saccharomyces cerevisiae . «Региональные центромеры» - это термин, придуманный для описания большинства центромеров, которые обычно образуются в областях предпочтительной последовательности ДНК, но которые также могут образовываться и на других последовательностях ДНК. Сигнал для образования региональной центромеры, по -видимому, является эпигенетическим . Большинство организмов, начиная от делящихся дрожжей Schizosaccharomyces, до людей, имеют региональные центромеры.
Что касается структуры митотической хромосомы, центромеры представляют собой ограниченную область хромосомы (часто называемой основным сужением), где две идентичные сестринские хроматиды наиболее близко находятся в контакте. Когда клетки вступают в митоз, сестринские хроматиды (две копии каждой хромосомной ДНК -молекулы, возникающие в результате репликации ДНК в форме хроматина) связаны вдоль их длины путем действия комплекса когезина . В настоящее время считается, что этот комплекс в основном высвобождается из хромосомных рычагов во время профазы, так что к тому времени, когда хромосомы выстраиваются в очередь на средней плоскости митотического веретена (также известный как метафазная пластина), последнее место, где они связаны друг с другом находится в хроматине в центре и вокруг него.
Позиция
[ редактировать ]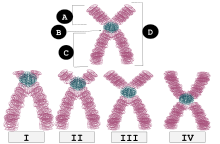
| я | Телоцентрический | Расположение центромеры очень близко к вершине, P Руки едва видимы, если они вообще видимы. |
| II | Акроцентрический | Q Руки по -прежнему намного дольше, чем ружения р, но Руки P длиннее, чем в телеокентрических. |
| Iii | Субметацентричный | Рыб и Q очень близки по длине, но не равны. |
| IV | Метацентрический | Р и Q Руки равны по длине. |
B : Centromere
C : Длинная рука (q Arm)
D : сестринские хроматиды
У людей положения центромеры определяют хромосомный кариотип , в котором каждая хромосома имеет две руки, P (более короче из двух) и Q (чем длиннее). Короткая рука «P», как сообщается, названа в честь французского слова «petit», что означает «маленький». [ 1 ] Положение центромеры относительно какой -либо конкретной линейной хромосомы используется для классификации хромосом как метацентрические, субметацентрические, акроцентрические, телоцентрические или голоцентрические. [ 2 ] [ 3 ]
| Центромерная позиция | Коэффициент длины рук | Знак | Описание |
|---|---|---|---|
| Medial Sensu Stricto | 1.0 – 1.6 | М | Метацентрический |
| Медиальная область | 1.7 | м | Метацентрический |
| Субмедиал | 3.0 | СМ | Субметацентричный |
| Субтерминал | 3.1 – 6.9 | ул | Субтелоцентрический |
| Терминальная область | 7.0 | Т | Акроцентрический |
| Терминальное чувство строгого | ∞ | Т | Телоцентрический |
| Примечания | – | Метацентричный : M + M | Atelocentric : M + M + SM + ST + T |
Метацентрический
[ редактировать ]Метацентричный означает, что центромер расположена на полпути между концами хромосомы, в результате чего руки приблизительно равны по длине. Когда центромеры являются метацентрическими, хромосомы, по-видимому, являются «x-образными».
Субметацентричный
[ редактировать ]Субметацентричный означает, что центромер расположено ниже середины, с одной хромосомы, короче другой, часто приводит к форме L.
Акроцентрический
[ редактировать ]Центромера акроцентрической хромосомы расположена так, что одно из хромосомных рук намного короче другого. «Акро-» в акроцентрике относится к греческому слову «пик». Геном человека имеет шесть акроцентрических хромосом, в том числе пять аутосомных хромосом ( 13 , 14 , 15 , 21 , 22 ) и Y -хромосомы .
Короткие акроцентрические P-арма содержат мало генетического материала и могут быть перемещены без значительного вреда, как при сбалансированной транслокации Робертсонского . В дополнение к некоторым генам, кодирующим белок, акроцентрические P-Армы человека также содержат области ядрево-органайзера (NORS), из которых рибосомальная РНК транскрибируется . Тем не менее, доля акроцентрических P-арм в клеточных линиях и тканях от нормальных доноров человека не содержит обнаруживаемых НОР. [ 4 ] Геном домашней лошади включает в себя одну метацентрическую хромосому, которая гомологична двум акроцентрическим хромосомам в конспецифической , но недооцененной лошади Przewalski . Это может отражать либо фиксацию сбалансированной транслокации Робертсона у домашних лошадей, либо, наоборот, фиксацию деления одной метацентрической хромосомы в две акроцентрические хромосомы у лошадей Przewalski. Аналогичная ситуация существует между геномами человека и великой обезьяны, с уменьшением двух акроцентрических хромосом в великих обезьянах до одной метацентрической хромосомы у людей (см. Анеуплоидию и хромосому человека 2 ).
Многие заболевания из результата несбалансированных транслокаций чаще включают акроцентрические хромосомы, чем другие неакроцентрические хромосомы. Акроцентрические хромосомы обычно расположены в ядрах и вокруг него . В результате эти хромосомы имеют тенденцию быть менее плотно упакованными, чем хромосомы на ядерной периферии. Постоянно, хромосомные области, которые менее плотно упакованы, также более подвержены хромосомным транслокациям при раке.
Телоцентрический
[ редактировать ]Телоцентрические хромосомы имеют центромер на одном конце хромосомы и, следовательно, имеют только одну руку на цитологическом (микроскопическом) уровне. Они не присутствуют у людей, но могут образовываться через клеточные хромосомные ошибки. Телоцентрические хромосомы встречаются естественным образом у многих видов, таких как домашняя мышь , в которых все хромосомы, кроме Y, являются телоцентрическими.
Субтелоцентрический
[ редактировать ]Центромеры субтелоцентрических хромосом расположены между серединой и концом хромосом, но находятся ближе к концу хромосом.
Центромерные типы
[ редактировать ]Акентрический
[ редактировать ]Ацентрическая хромосома - это фрагмент хромосомы, в которой отсутствует центромер. Поскольку центромеры являются точкой прикрепления для веретеновых волокон в клеточном делении, фрагменты ацентрических фрагментов не распределены по дочерним клеткам во время деления клеток. В результате дочерней ячейке не хватает ацентрического фрагмента, и могут возникнуть вредные последствия.
Разбивающие хромосомы события также могут генерировать ацентрические хромосомы или фрагменты по ацентрическим.
Дицентрический
[ редактировать ]Дицентрическая хромосома является аномальной хромосомой с двумя центромерами, которые могут быть нестабильными с помощью клеточных делений. Он может образовываться посредством транслокации или слияния двух сегментов хромосом, каждый с центромерой. Некоторые перестройки продуцируют как дицентрические хромосомы, так и фрагменты инспекта, которые не могут прикрепляться к шпинделям при митозе. [ 5 ] Образование дицентрических хромосом было связано с генетическими процессами, такими как Робертсонская транслокация [ 6 ] и парацентрическая инверсия. [ 7 ] Дицентрические хромосомы могут иметь различные судьбы, включая митотическую стабильность. [ 8 ] В некоторых случаях их стабильность происходит от инактивации одного из двух центромеров для создания функционально моноцентрической хромосомы, способной к нормальной передаче дочерних клеток во время деления клеток. [ 9 ]
Например, человеческая хромосома 2 , которая, как полагают, является результатом транслокации Робертсона в какой -то момент эволюции между великими обезьянами и гомо , имеет второй рудиментальный, центромер в середине его длинной руки. [ 10 ]
Моноцентрический
[ редактировать ]Моноцентрическая хромосома - это хромосома, которая имеет только одну центромеру в хромосоме и образует узкое сужение.
Моноцентрические центромеры являются наиболее распространенной структурой на очень повторяющейся ДНК у растений и животных. [ 11 ]
Голоцентрический
[ редактировать ]В отличие от моноцентрических хромосом, голоцентрические хромосомы не имеют четкого первичного сужения при просмотре при митозе. Вместо этого волокна шпинделя прикрепляются почти всю (греческую: голово) длину хромосомы. В голоцентрических хромосомах центромерные белки, такие как CENPA (CENH3), распространяются по всей хромосоме. [ 12 ] Нематода, Caenorhabditis elegans , является хорошо известным примером организма с голоцентрическими хромосом, [ 13 ] Но этот тип центромеры можно найти у различных видов, растений и животных по всему эукариотам. Holocentromeres фактически состоят из нескольких распределенных центромерных единиц, которые образуют линейную структуру вдоль хромосом во время митоза. [ 14 ] Альтернативные или нетрадиционные стратегии развернуты в мейозе для достижения гомологичной спаривания и сегрегации хромосом, необходимых для производства жизнеспособных гаметов или гаметофитов для сексуального размножения.
Различные типы голоцентромеров существуют у разных видов, а именно с центромерными повторяющимися последовательностями ДНК или без или без него, а также с CENH3 или без него . Holocentricity развивалась по меньшей мере 13 раз независимо в различных зеленых водорослях, простейших, беспозвоночных и различных семействах растений. [ 15 ] В отличие от моноцентрических видов, где ацентрические фрагменты обычно теряются во время деления клеток, разрушение голоцентрических хромосом создает фрагменты с нормальными сайтами прикрепления клетчатки. [ 16 ] Из -за этого организмы с голоцентрическими хромосомами могут быстрее развивать вариацию кариотипа, способные заживлять фрагментированные хромосомы посредством последующего добавления тогоромовых колпачков в местах поломки. [ 17 ]
Полицентрический
[ редактировать ]Полицентрические хромосомы имеют несколько кинетохорных кластеров, то есть центромы. Термин частично перекрывается с «голоцентрическим», но «полицентрический» явно предпочтительнее при обсуждении дефективно сформированных моноцентрических хромосомов. Также существует некоторая фактическая двусмысленность, так как нет четкой линии перехода от кинетохоров, охватывающих всю хромосому к отдельным кластерам. Другими словами, разница между «всей хромосомой - это центром» и «хромосома нет центрома», и варьируется использование. Помимо «полицентричности», используемого больше о дефектах, нет четких предпочтений в других темах, таких как эволюционное происхождение или распределение кинетохора и подробная структура (например, как видно при анализе маркировки или сборки генома ). [ 18 ] [ 19 ] [ 20 ] [ 21 ]
Даже явно отчетливые кластеры белков кинетохор не обязательно вызывают более одного сужения: «метаполицентрические» хромосомы имеют одно удлиненное сужение хромосомы, соединяя более длительный сегмент, который все еще заметно короче, чем хроматиды. [ 22 ] Метаполицентрические хромосомы могут быть этапом появления и подавления Centromere Drive, типа мейотического движения , который нарушает паритет моноцентрическими центромерами, выращивающими дополнительные кинетохорские белки, чтобы получить преимущество во время мейоза. [ 23 ]
Человеческие хромосомы
[ редактировать ]
| Хромосома | Центромер положение ( MBP ) |
Категория | Хромосома Размер (MBP) |
Центромер размер (MBP) |
|---|---|---|---|---|
| 1 | 125.0 | Метацентрический | 247.2 | 7.4 |
| 2 | 93.3 | субметацентричный | 242.8 | 6.3 |
| 3 | 91.0 | Метацентрический | 199.4 | 6.0 |
| 4 | 50.4 | субметацентричный | 191.3 | — |
| 5 | 48.4 | субметацентричный | 180.8 | — |
| 6 | 61.0 | субметацентричный | 170.9 | — |
| 7 | 59.9 | субметацентричный | 158.8 | — |
| 8 | 45.6 | субметацентричный | 146.3 | — |
| 9 | 49.0 | субметацентричный | 140.4 | — |
| 10 | 40.2 | субметацентричный | 135.4 | — |
| 11 | 53.7 | субметацентричный | 134.5 | — |
| 12 | 35.8 | субметацентричный | 132.3 | — |
| 13 | 17.9 | акроцентрический | 114.1 | — |
| 14 | 17.6 | акроцентрический | 106.3 | — |
| 15 | 19.0 | акроцентрический | 100.3 | — |
| 16 | 36.6 | Метацентрический | 88.8 | — |
| 17 | 24.0 | субметацентричный | 78.7 | — |
| 18 | 17.2 | субметацентричный | 76.1 | — |
| 19 | 26.5 | Метацентрический | 63.8 | — |
| 20 | 27.5 | Метацентрический | 62.4 | — |
| 21 | 13.2 | акроцентрический | 46.9 | — |
| 22 | 14.7 | акроцентрический | 49.5 | — |
| Х | 60.6 | субметацентричный | 154.9 | — |
| И | 12.5 | акроцентрический | 57.7 | — |
Основываясь на микрографических характеристиках размера, положении центромеры, а иногда и о наличии хромосомного спутника , хромосомы человека классифицируются в следующих группах: [ 24 ]
| Группа | Хромосомы | Функции |
|---|---|---|
| Группа а | Хромосома 1–3 | Большой, метацентричный и субметоцентричный |
| Группа б | Хромосома 4–5 | Большой, субметацентричный |
| Группа c | Хромосома 6–12, х | Средний, субметацентричный |
| Группа d | Хромосома 13–15 | Средний, акроцентричный, со спутником |
| Группа e | Хромосома 16–18 | Маленький, метацентричный и субметоцентричный |
| Группа f | Хромосома 19–20 | Очень маленький, метацентричный |
| Группа G. | Хромосома 21–22, y | Очень маленький, акроцентричный, со спутником |
Последовательность
[ редактировать ]Есть два типа центромеров. [ 25 ] У региональных центромеров последовательности ДНК способствуют, но не определяют функцию. Региональные центромеры содержат большое количество ДНК и часто упаковываются в гетерохроматин . У большинства эукариот последовательность ДНК Centromere состоит из больших массивов повторяющейся ДНК (EG -спутниковой ДНК ), где последовательность внутри отдельных повторяющихся элементов аналогична, но не идентична. У людей первичная центромерная повторная единица называется α-сателлитом (или альфоидом), хотя в этой области обнаружено ряд других типов последовательностей. [ 26 ] Спутники Centromere предполагают, что они развиваются путем процесса, называемого многоуровневым расширением. Они быстро развиваются между видами, и анализ у диких мышей показывает, что количество спутниковых копий и неоднородности связаны с происхождением популяции и подвидом. [ 27 ] Кроме того, спутниковые последовательности могут зависеть от инбридинга. [ 27 ]
Центромеры точек меньше и компактны. Последовательности ДНК необходимы и достаточны для указания идентичности и функции центромер в организмах с точечными центромерами. У подавляющих дрожжей центромерная область относительно небольшая (около 125 п.н. ДНК) и содержит две высококонсервативные последовательности ДНК, которые служат сайтами связывания для незаменимых белков кинетохора . [ 26 ]
Наследование
[ редактировать ]Поскольку центромерная последовательность ДНК не является ключевой детерминантой центромерной идентичности у метазоаций , считается, что эпигенетическое наследование играет важную роль в определении центромеры. [ 28 ] Дочерние хромосомы собирают центромеры в том же месте, что и родительская хромосома, независимая от последовательности. Было предложено, чтобы вариант гистона H3 CENP-A (белок Centromere A) является эпигенетической меткой центромеры. [ 29 ] Возникает вопрос о том, должен ли все еще быть какой -то оригинальный способ, которым центромер указан, даже если он впоследствии распространяется эпигенетически. Если центромера эпигенетически унаследована от одного поколения к другому, проблема перенесена на начало первых метазоанов.
С другой стороны, благодаря сравнению центромеров в Х -хромосомах, в этих регионах наблюдались эпигенетические и структурные изменения. Кроме того, недавняя сборка генома человека обнаружила возможный механизм того, как развиваются перицентромерные и центромерные структуры, посредством слоистой модели расширения для последовательностей αSAT. Эта модель предполагает, что различные повторения последовательности αSAT периодически возникают и расширяются в активном векторе, вытесняя старые последовательности и становясь местом сборки кинетохора. ΑSAT может происходить из одного и того же или из разных векторов. Поскольку этот процесс повторяется с течением времени, слои, которые окружают активную центромеру, сокращаются и ухудшаются. Этот процесс поднимает вопросы о взаимосвязи между этим динамическим эволюционным процессом и положением центромеры. [ 30 ]
Структура
[ редактировать ]Центромерная ДНК обычно находится в состоянии гетерохроматина , что необходимо для рекрутирования комплекса когезина , который опосредует сплоченность сестринских хроматидов после репликации ДНК, а также координации разделения сестринских хроматидов во время анафазы. В этом хроматине нормальный гистон H3 заменяется центром-специфическим вариантом CENP-A у людей. [ 31 ] Присутствие CENP-A считается важным для сборки кинетохора на центромере. Было показано, что CENP-C локализуется почти исключительно в этих областях CENP-A, связанного с хроматином. В клетках человека гистоны обнаруживаются наиболее обогащены H4K20ME 3 и H3K9ME3 [ 32 ] которые известны гетерохроматические модификации. У Drosophila острова ретроэлементов являются основными компонентами центромеров. [ 33 ]
У дрожжей Schizosaccharomyces pombe (и, вероятно, у других эукариот) образование центромерного гетерохроматина связано с РНКи . [ 34 ] У нематод, таких как Caenorhabditis Elegans , некоторые растения и заказы насекомых Lepidoptera и Hemiptera, хромосомы являются «голоцентрическими», что указывает на то, что нет первичного места прикрепления микротрубочек или первичного сужения и «диффузное» Kinetochore сборка вдоль всего Длина хромосомы.
Центромерные аберрации
[ редактировать ]В редких случаях неоцентромеры могут образовываться на новых участках на хромосоме в результате перемещения центромеры. Это явление наиболее хорошо известно в клинических исследованиях человека, и в настоящее время более 90 известных неоцентромеров человека, выявленные на 20 различных хромосомах. [ 35 ] [ 36 ] Образование неоцентромеры должно быть связано с инактивацией предыдущей центромеры, поскольку хромосомы с двумя функциональными центромерами ( дицентрическая хромосома ) приведут к разрушению хромосомы во время митоза. В некоторых необычных случаях неоцентромеры человека спонтанно образуются на фрагментированных хромосомах. Некоторые из этих новых позиций были первоначально эухроматическими и не имели альфа -спутниковой ДНК в целом. У неоцентромеров не хватает повторяющейся структуры, наблюдаемой в нормальных центромерах, которые предполагают, что образование центромеры в основном контролируется эпигенетически . [ 37 ] [ 38 ] Со временем неоцентромер может накапливать повторяющиеся элементы и превратиться в то, что известно как эволюционная новая центромера. В хромосомах приматов есть несколько известных примеров, где положение центромеры отличается от центральной центромеры той же хромосомы и, как считается, является эволюционным новым центромером. [ 37 ] Предполагается, что перемещение центромер и формирование эволюционных новых центромеров является механизмом видообразования . [ 39 ]
Белки Centromere также являются аутоантигенной мишенью для некоторых антителеарных антител , таких как антицентромерные антитела .
Дисфункция и болезнь
[ редактировать ]Было известно, что недостаток центромеры способствует неправильной сегрегации хромосом, что тесно связано с раком и выкидышами. Примечательно, что сверхэкспрессия многих генов центромер была связана с злокачественными фенотипами рака. Сверхэкспрессия этих генов центромер может увеличить нестабильность геномной раковой заболевания. Повышенная геномная нестабильность, с одной стороны, относится к злокачественным фенотипам; С другой стороны, это делает опухолевые клетки более уязвимыми к специфической адъювантной терапии, такой как определенная химиотерапия и лучевая терапия. [ 40 ] Нестабильность повторяющейся ДНК центромер была недавно показана при раке и старении. [ 41 ]
Восстановление центромерной ДНК
[ редактировать ]Когда разрывы ДНК происходят в центромерах в фазе G1 клеточного цикла , клетки способны рекрутировать гомологичную рекомбинационную рефлекционную машину в поврежденный участок, даже в отсутствие сестринского хроматида . [ 42 ] По -видимому, гомологичный рекомбинационный восстановление может происходить при центромерных разрывах по всему клеточному циклу, чтобы предотвратить активацию неточных путей восстановления мутагенной ДНК и сохранить центромерную целостность. [ 42 ]
Этимология и произношение
[ редактировать ]Слово Centromere ( / ˈ S ɛ n t r ə ˌ m ɪər / [ 43 ] [ 44 ] Использует сочетание форм центро- и мерера , давая «центральную часть», описывая местоположение центромеры в центре хромосомы.
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ «P + Q = Решено, будучи истинной историей о том, как хромосома получила свое название» . 2011-05-03.
- ^ "Какие различные типы хромосом существуют?" , Уроки генетики Николай , YouTube, 2013-10-12, архивированы из оригинала 2021-12-11 , извлечен 2017-05-28
- ^ Jump up to: а беременный Леван А., Фредга К, Сандберг А.А. (декабрь 1964 г.). «Номенклатура для центромерного положения на хромосомах». Hereditas . 52 (2): 201–220. doi : 10.1111/j.1601-5223.1964.tb01953.x .
- ^ Van Sluis M, Van Vuuren C, Mangan H, McStay B (май 2020). «NOR на акроцентрическую хромосому человека P-арма активны по умолчанию и могут ассоциироваться с нуклеоли независимо от рДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 117 (19): 10368–10377. Bibcode : 2020pnas..11710368V . doi : 10.1073/pnas.2001812117 . PMC 7229746 . PMID 32332163 .
- ^ Nussbaum R, McInnes R, Willard H, Hamosh A (2007). Thompson & Thompson Genetics in Medicine . Филадельфия (Пенсильвания): Сондерс. п. 72. ISBN 978-1-4160-3080-5 .
- ^ Thompson & Thompson Genetics in Medicine (7 -е изд.). п. 62
- ^ Хартвелл Л., Худ Л., Голдберг М., Рейнольдс А., Ли С. (2011). Генетика от генов до геномов (4 -е изд.). Нью-Йорк: МакГроу-Хилл. ISBN 9780073525266 .
- ^ Линч С.А., Эшкрофт К.А., Зволински С., Кларк С., Берн Дж (март 1995 г.). «Синдром Кабуки, подобные монозиготным мальчикам-близнецам с псевдодикентрической хромосомой 13» . Журнал медицинской генетики . 32 (3): 227–230. doi : 10.1136/jmg.32.3.227 . PMC 1050324 . PMID 7783176 .
- ^ Стимпсон, Кейтлин М.; Matheny, Justyne E.; Салливан, Бет А. (2012). «Дицентрические хромосомы: уникальные модели для изучения функции центромеры и инактивации» . Исследование хромосом . 20 (5): 595–605. doi : 10.1007/s10577-012-9302-3 . PMC 3557915 .
- ^ Аварелло; и др. (1992). «Свидетельство о наследственном альфоидном домене на длинной руке хромосомы человека 2». Человеческая генетика . 89 (2): 247–9. doi : 10.1007/bf00217134 . PMID 1587535 . S2CID 1441285 .
- ^ Barra V, Fachinetti D (октябрь 2018 г.). «Темная сторона центромеров: типы, причины и последствия структурных аномалий, подразумевающих центромерную ДНК» . Природная связь . 9 (1): 4340. Bibcode : 2018natco ... 9.4340b . doi : 10.1038/s41467-018-06545-y . PMC 6194107 . PMID 30337534 .
- ^ Neumann P, Navrátilová A, Schroeder-Reiter E, Koblížková A, Steinbauer V, Chochcholová E, et al. (2012). «Растяжение правил: моноцентрические хромосомы с несколькими доменами центромер» . PLOS Genetics . 8 (6): E1002777. Doi : 10.1371/journal.pgen.1002777 . PMC 3380829 . PMID 22737088 .
- ^ Дернбург AF (июнь 2001 г.). «Здесь, там, и везде: Кинетохор функционирует на голоцентрических хромосомах» . Журнал клеточной биологии . 153 (6): F33 - F38. doi : 10.1083/jcb.153.6.f33 . PMC 2192025 . PMID 11402076 .
- ^ Marques A, Ribeiro T, Neumann P, Macas J, Novák P, Schubert V, et al. (Ноябрь 2015). «Holocentromeres в Rhynchospora связаны с геномными центромерами, специфичными для повторных массивов, перемежающихся среди эухроматина» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (44): 13633–13638. Bibcode : 2015pnas..11213633M . doi : 10.1073/pnas.15122555112 . PMC 4640781 . PMID 26489653 .
- ^ Мелтерс DP, Paliulis LV, Korf If, Chan SW (июль 2012 г.). «Холоцентрические хромосомы: конвергентная эволюция, мейотическая адаптация и геномный анализ» . Исследование хромосом . 20 (5): 579–593. doi : 10.1007/s10577-012-9292-1 . PMID 22766638 . S2CID 3351527 .
- ^ Hughes-Schrader S ; RIS H (август 1941 г.). «Прикрепление диффузного шпинделя кокцитов, подтвержденное митотическим поведением индуцированных фрагментов хромосом» . Журнал экспериментальной зоологии . 87 (3): 429–456. doi : 10.1002/jez.1400870306 . ISSN 0022-104X .
- ^ Jankowska M, Fuchs J, Klocke E, Fojtová M, Polanská P, Fajkus J, et al. (Декабрь 2015). «Голокинетические центромеры и эффективное заживление теломер позволяют быстро эволюцию кариотипа». Хромосома . 124 (4): 519–528. doi : 10.1007/s00412-015-0524-y . PMID 26062516 . S2CID 2530401 .
- ^ Божественный MB (апрель 1954 г.). «Диффузная» центромер или полицентрические хромосомы в спирогире ». Анналы ботаники . 18 (2): 143–144. doi : 10.1093/oxfordjournals.aob.a083387 .
- ^ Куо, yt; Камара, как; Шуберт, V (2023-06-13). «Холоцентромеры могут состоять только из нескольких спутниковых массивов размером с мегабазу» . Природная связь . 14 doi : 10.1038/s41467-023-38922-7 . PMC 10264360 .
- ^ Senaratne, Aruni P.; Кортес-Сильва, Нурия; Дринненберг, Инес А. (июль 2022 г.). «Эволюция голоцентрических хромосом: драйверы, разнообразие и сдерживание» . Семинары в биологии клеток и развития . 127 : 90–99. doi : 10.1016/j.semcdb.2022.01.003 . Получено 2024-09-11 .
- ^ Ma B, Wang H, Liu J, Chen L, Xia X, Wei W, Yang Z, Yuan J, Luo Y, He N. (2023-05-31). «Без разрывы геном шелковицы выясняет архитектуру и эволюцию полицентрических хромосом» . Исследование садоводства . 10 (7). doi : 10.1093/hr/uhad111 . PMC 10541557 . Получено 2024-09-11 .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Исии, Мидори; Акийоши, Бунго (февраль 2022 г.). «Пластичность в организации Centromere и композиции кинетохора: уроки разнообразия» . Современное мнение в клеточной биологии . 74 (клеточное ядро 2022): 47–54. doi : 10.1016/j.ceb.2021.12.007 . Получено 2024-09-11 .
- ^ Зедек, Франтисек; Бур, Петр (декабрь 2016 г.). «Отсутствие положительного отбора на CENH3 в Лузуле предполагает, что голокинетические хромосомы могут подавлять Centromere Drive» . Анналы ботаники . 118 : 1347–1352. doi : 10.1093/aob/mcw186 . PMC 5155603 . Получено 2024-09-11 .
- ^ Erwinsyah, R., Riandi, & Nurjhani, M. (2017). «Актуальность активности анализа хромосом человека против концепции мутации в курсе генетики. Серия конференций IOP» . Материалогическое наук и инженерия . doi : 10.1088/1757-899x/180/1/012285 . S2CID 90739754 .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Плута А.Ф., Маккей А.М., Эйнштейн А.М., Голдберг И.Г., Эрншоу В.К. (декабрь 1995 г.). «Центромер: центр хромосомной активности». Наука . 270 (5242): 1591–1594. Bibcode : 1995sci ... 270.1591p . doi : 10.1126/science.270.5242.1591 . PMID 7502067 . S2CID 44632550 .
- ^ Jump up to: а беременный Мехта Г.Д., Агарвал М.П., Гош С.К. (август 2010 г.). «Centromere Identity: проблема, с которой нужно столкнуться». Молекулярная генетика и геномика . 284 (2): 75–94. doi : 10.1007/s00438-010-0553-4 . PMID 20585957 . S2CID 24881938 .
- ^ Jump up to: а беременный Arora Up, Charlebois C, Lawal RA, Dumont BL (апрель 2021 г.). «Разнообразие населения и подвида на спутниках Centromere мыши» . BMC Genomics . 22 (1): 279. DOI : 10.1186/S12864-021-07591-5 . PMC 8052823 . PMID 33865332 .
- ^ Далал Y (февраль 2009 г.). «Эпигенетическая спецификация центромеров». Биохимия и клеточная биология . 87 (1): 273–282. doi : 10.1139/O08-135 . PMID 19234541 .
- ^ Бернад Р., Санчес П., Лосада А (ноябрь 2009 г.). «Эпигенетическая спецификация центромеров CENP-A». Экспериментальные исследования клеток . 315 (19): 3233–3241. doi : 10.1016/j.yexcr.2009.07.023 . PMID 19660450 .
- ^ Altmose, Николас; Logsdon, Glennis A.; Bzikadze, Andrey V.; Сидхвани, Прагая; Лэнгли, Саша А.; Caldas, Gina v.; Хойт, Саванна Дж.; Уральский, Лев; Yabov, Fedor D.; Shew, Colin J.; Саурия, Майкл Эг; Борчерс, Мэтью; Гершман, Ариэль; Михинко, Алла; Шепелев, Валерий А. (апрель 2022 г.). «Полные геномные и эпигенетические карты центтромеров человека » Наука 376 (6588): EABL4 Doi : 10.1126/ science.abl4 ISSN 0036-8 PMC 9233505 PMID 35357911 .
- ^ Chueh AC, Wong LH, Wong N, Choo KH (январь 2005 г.). «Распределение переменного и иерархического размера в L1-рефрупционных кластерах CENP-A в функциональной неоцентромере человека» . Молекулярная генетика человека . 14 (1): 85–93. doi : 10.1093/hmg/ddi008 . PMID 15537667 .
- ^ Rosenfeld JA, Wang Z, Schones DE, Zhao K, Desalle R, Zhang MQ (март 2009 г.). «Определение обогащенных модификаций гистонов в негенических частях генома человека» . BMC Genomics . 10 : 143. DOI : 10.1186/1471-2164-10-143 . PMC 2667539 . PMID 19335899 .
- ^ Чанг Ч., Чаван А., Палладино Дж., Вэй Х, Мартинс Н.М., Сантинелло Б. и др. (Май 2019). «Острова ретроэлементов являются основными компонентами центромеров дрозофилы» . PLOS Биология . 17 (5): E3000241. doi : 10.1371/journal.pbio.3000241 . PMC 6516634 . PMID 31086362 .
- ^ Volpe TA, Kidner C, Hall IM, Teng G, Grewal SI, Martienssen RA (сентябрь 2002 г.). «Регуляция гетерохроматического молчания и метилирования лизина-9 гистона H3 с помощью РНКи» . Наука . 297 (5588): 1833–1837. Bibcode : 2002sci ... 297.1833v . doi : 10.1126/science.1074973 . PMID 12193640 . S2CID 2613813 .
- ^ Marshall OJ, Chueh AC, Wong LH, Choo KH (февраль 2008 г.). «Нецентромеры: новое понимание структуры центромеры, развития болезней и эволюции кариотипа» . Американский журнал человеческой генетики . 82 (2): 261–282. doi : 10.1016/j.ajhg.2007.11.009 . PMC 2427194 . PMID 18252209 .
- ^ Warburton PE (2004). «Хромосомная динамика образования неоцентромеры человека». Исследование хромосом . 12 (6): 617–626. doi : 10.1023/b: chro.0000036585.44138.4b . PMID 15289667 . S2CID 29472338 .
- ^ Jump up to: а беременный Rocchi M, Archidiacono N, Schempp W, Capozzi O, Stanyon R (январь 2012 г.). «Центромерное положение у млекопитающих» . Наследственность . 108 (1): 59–67. doi : 10.1038/hdy.2011.101 . PMC 3238114 . PMID 22045381 .
- ^ Tolomeo D, Capzzi O, Stanyon RR, Archidiacono N, D'Adbbo P, Catacchio CR, et al. (Февраль 2017 г.). «Эпигенетическое происхождение эволюционных центров романов » Научные отчеты 7 (1): 41980. Bibcode : 2017natsr ... 741980t Doi : 10.1038/ srep41980 PMC 5290474 28155877PMID
- ^ Браун Д.Д., О'Нил Р.Дж. (сентябрь 2010 г.). «Хромосомы, конфликт и эпигенетика: повторное представление хромосом». Ежегодный обзор геномики и генетики человека . 11 (1): 291–316. doi : 10.1146/annurev-Genom-082509-141554 . PMID 20438362 .
- ^ Zhang W, Mao JH, Zhu W, Jain AK, Liu K, Brown JB, Karpen GH (август 2016 г.). «Misexpression« Центромер и кинетохорская гена предсказывает выживаемость пациента с раком и реакцию на лучевую терапию и химиотерапию » . Природная связь . 7 : 12619. BIBCODE : 2016Natco ... 712619Z . doi : 10.1038/ncomms12619 . PMC 5013662 . PMID 27577169 .
- ^ Giunta S, Funabiki H (февраль 2017 г.). «Целостность повторений ДНК Centromere человека защищена CENP-A, CENP-C и CENP-T» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (8): 1928–1933. Bibcode : 2017pnas..114.1928g . doi : 10.1073/pnas.1615133114 . PMC 5338446 . PMID 28167779 .
- ^ Jump up to: а беременный Yilmaz D, Furst A, Meaburn K, Lezaja A, Wen Y, Altmeyer M, Reina-San-Martin B, Soutoglou E (декабрь 2021 г.). «Активация гомологичной рекомбинации в G1 сохраняет центромерную целостность». Природа . 600 (7890): 748–753. Bibcode : 2021natur.600..748y . doi : 10.1038/s41586-021-04200-z . PMID 34853474 . S2CID 244800481 .
- ^ "Centromere" . Merriam-Webster.com Словарь . Мерриам-Уэбстер.
- ^ "Centromere" . Dictionary.com unabridged (онлайн). н.д.
Дальнейшее чтение
[ редактировать ]- Мехта Г.Д., Агарвал М.П., Гош С.К. (август 2010 г.). «Centromere Identity: проблема, с которой нужно столкнуться». Молекулярная генетика и геномика . 284 (2): 75–94. doi : 10.1007/s00438-010-0553-4 . PMID 20585957 . S2CID 24881938 .
- Lodish H, Berk A, Kaiser CA, Kaiser C, Krieger M, Scott MP, Bretscher A, Ploegh H, Matsudaira (2008). Молекулярная клеточная биология (6 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-7601-7 .
- Нагки, Ченг З., Оуян С., Талберт П.Б., Ким М., Джонс К.М. и др. (Февраль 2004 г.). «Секвенирование центра рисового центра раскрывает активные гены » Природа генетика 36 (2): 138–1 Doi : 10.1038/ ng1 PMID 14716315
Внешние ссылки
[ редактировать ]- «Центромер риса, предположительно тихой генетический домен, сюрпризы» . ScienceDaily (пресс -релиз). 13 января 2004 г.