Лишайник







Лишайник ( / ˈ l aɪ k ən / LY -kən , UK также / ˈ l ɪ tʃ ən / LITCH -ən ) представляет собой симбиоз водорослей живущих или цианобактерий, среди нитей нескольких видов грибов , вместе с дрожжами , встроенными в кору или «кожу», во мутуалистических отношениях. [1] [2] [ устаревший источник ] [3] [ устаревший источник ] [4] [ устаревший источник ] [5] Лишайники являются важными участниками круговорота питательных веществ и выступают в качестве продуцентов, которыми питаются многие представители более высоких трофических групп, такие как северные олени, брюхоногие моллюски, нематоды, клещи и коллемболы. [6] [7] [8] [9] Лишайники обладают свойствами, отличными от свойств составляющих их организмов. Они бывают разных цветов, размеров и форм и иногда похожи на растения, но не являются растениями . У них могут быть крошечные безлистные ветви ( кустаристые ); плоские листовидные структуры ( листовидные ); разрастаются в виде корки, плотно прилегая к поверхности ( подложке ), как толстый слой краски ( корка ); [10] иметь порошкообразный вид ( проказа ); или другие формы роста . [11]
Макролишайник ; — это лишайник кустовидной или листовой формы все остальные лишайники называются микролишайниками . [2] Здесь «макро» и «микро» относятся не к размеру, а к форме роста. [2] Общие названия лишайников могут содержать слово «мох» (например, « оленьий мох », « исландский мох »), а лишайники могут внешне выглядеть как мхи и расти вместе с ними , но они не имеют тесного родства со мхами или каким-либо растением. [4] : 3 У лишайников нет корней, поглощающих воду и питательные вещества, как у растений. [12] : 2 но, как и растения, они производят себе питание посредством фотосинтеза . [13] Когда они растут на растениях, они не живут как паразиты , а вместо этого используют поверхность растения в качестве субстрата.
Лишайники встречаются от уровня моря до высоких альпийских возвышенностей, во многих условиях окружающей среды и могут расти практически на любой поверхности. [13] [14] Они в изобилии растут на коре, листьях , мхах и других лишайниках. [12] и свисающие с ветвей, «живущих в воздухе» ( эпифиты ) в тропических лесах и лесах умеренного пояса . Они растут на камнях, стенах, надгробиях , крышах , открытых поверхностях почвы, резине, костях и в почве как часть биологических почвенных корок . Различные лишайники приспособились к выживанию в самых экстремальных условиях на Земле: арктической тундре , жарких сухих пустынях , скалистых побережьях и токсичных шлаковых отвалах. Они могут жить даже внутри твердой породы, разрастаясь между зернами ( эндолит ).
Известно около 20 000 видов. [15] Некоторые лишайники утратили способность к половому размножению, но продолжают видообразоваться . [12] [16] Их можно рассматривать как относительно автономные миниатюрные экосистемы , в которых грибы, водоросли или цианобактерии могут взаимодействовать с другими микроорганизмами в функционирующей системе, которая может развиваться как еще более сложный составной организм . [17] [18] [19] [20] Лишайники могут быть долгожителями , а некоторые из них считаются одними из старейших живых существ. [4] [21] Они являются одними из первых живых существ, которые растут на свежих камнях, обнаженных после такого события, как оползень. Длительная продолжительность жизни, а также медленная и регулярная скорость роста некоторых видов могут быть использованы для датировки событий ( лихенометрия ). Лишайники являются ключевым видом во многих экосистемах и приносят пользу деревьям и птицам . [22]
Этимология и произношение
[ редактировать ]Английское слово «лишайник» происходит от греческого λειχήν leichēn («древесный мох, лишайник, лишайниковая сыпь на коже») через латинский лишайник . [23] [24] [25] Греческое существительное, которое буквально означает «лизать», происходит от глагола λείχειν leichein , «лизать». [26] [27] В американском английском «lichen» произносится так же, как глагол «liken» ( / ˈ l aɪ k ən / ). как это произношение, так и рифма со словом «кухня» ( / ˈ l ɪ tʃ ən / ). В британском английском используются [28] [29] [30]
Анатомия и морфология
[ редактировать ]Формы роста
[ редактировать ]Лишайники растут в самых разных формах и формах; этот внешний вид известен как их морфология . Форма лишайника обычно определяется организацией грибных нитей. [31] Нерепродуктивные ткани или вегетативные части тела называются слоевищем . Лишайники сгруппированы по типу слоевища, поскольку слоевище обычно является наиболее визуально заметной частью лишайника. Формы роста слоевища обычно соответствуют нескольким основным типам внутренней структуры. Общие названия лишайников часто происходят от формы роста или цвета, типичного для рода лишайников .
Обычными группами форм роста слоевища лишайников являются:
- кустистый [32] [33] [34] – растущий в виде пучка или многоразветвленного безлистного мини-кустарника, прямостоячий или свисающий, трехмерные ветви почти круглого сечения ( терете ) или сплюснутые
- листоватый [32] [33] – растущие в двухмерных, плоских, листовидных лепестках
- покрытый коркой [10] [32] [33] – корковая, плотно прилегающая к поверхности ( подложке ), как толстый слой краски
- покрытый мелкими чешуйками [34] – образованы мелкими листовидными чешуйками, покрытыми коркой снизу, но свободными на кончиках
- проказа [35] – пудровый
- студенистый – желеобразный
- нитевидный – волокнистый или похожий на спутанные волосы
- биссоид – тонкий, как взчесанная шерсть
- бесструктурный
Существуют различия в типах роста у одного вида лишайников, серые зоны между описаниями типов роста и совпадение между типами роста, поэтому некоторые авторы могут описывать лишайники, используя разные описания типов роста.
Когда корковый лишайник стареет, центр может начать трескаться, как старая засохшая краска, старое разбитое асфальтовое покрытие или как многоугольные «островки» растрескавшейся грязи на высохшем дне озера. Это называется римозой или ареолой , а «островные» части, разделенные трещинами, называются ареолами. [32] Ареолы кажутся разделенными, но они (или были) [ нужна ссылка ] соединены подлежащим проталломом или гипоталломом . [35] Когда корковый лишайник растет из центра и кажется расходящимся, его называют корковым плакодиоидом. Когда края ареол приподнимаются от субстрата, это называется чешуйчатым . [36] : 159 [34]
Эти группы форм роста точно не определены. Листовидные лишайники иногда могут разветвляться и выглядеть кустистыми. Кустистые лишайники могут иметь уплощенные ветвящиеся части и выглядеть листоватыми. Чешуйчатые лишайники могут появиться там, где приподнимаются края. Желатиновые лишайники в сухом состоянии могут выглядеть листоватыми. [36] : 159
Слоевище не всегда является наиболее заметной визуально частью лишайника. Некоторые лишайники могут расти внутри твердой породы между зернами ( эндолитические лишайники ), при этом за пределами камня растет только видимая часть полового плода. [32] Они могут быть драматичными по цвету или внешнему виду. [32] Формы этих половых частей не относятся к вышеуказанным категориям форм роста. [32] Наиболее визуально заметные репродуктивные части часто представляют собой круглые, приподнятые, пластинчатые или дискообразные выросты с извилистыми краями и описаны в разделах ниже.
Цвет
[ редактировать ]
Лишайники бывают разных цветов. [12] : 4 Окраска обычно определяется фотосинтетическим компонентом. [31] Специальные пигменты, такие как желтая усниновая кислота , придают лишайникам различные цвета, включая красный, оранжевый, желтый и коричневый, особенно в открытых и сухих местах обитания. [37] В отсутствие специальных пигментов лишайники обычно имеют цвет от ярко-зеленого до оливково-серого во влажном состоянии, серого или серовато-зеленого до коричневого в сухом состоянии. [37] Это связано с тем, что влага заставляет поверхность кожи ( кору головного мозга ) становиться более прозрачной, обнажая зеленый слой фотобионта. [37] Разноцветные лишайники, покрывающие большие площади открытых поверхностей камней, или лишайники, покрывающие кору или свисающие с нее, могут представлять собой впечатляющее зрелище, когда пятна разных цветов «оживают» или «светятся» яркими эффектами после дождя.
Лишайники разного цвета могут обитать в разных прилегающих участках скалы, в зависимости от угла воздействия света. [37] Колонии лишайников могут иметь впечатляющий внешний вид, доминируя над большей частью поверхности визуального ландшафта в лесах и природных местах, например, вертикальная «краска», покрывающая обширные скальные поверхности национального парка Йосемити . [38]
Цвет используется для идентификации. [39] : 4 Цвет лишайника меняется в зависимости от того, влажный лишайник или сухой. [39] Описания цветов, используемые для идентификации, основаны на цвете, который проявляется, когда лишайник высыхает. [39] Сухие лишайники с цианобактериями в качестве партнера по фотосинтезу обычно имеют темно-серый, коричневый или черный цвет. [39]
Нижняя сторона листовидных долей листоватых лишайников отличается по цвету от верхней стороны ( дорзивентральная ), чаще бурая или черная, иногда белая. Кустистый лишайник может иметь уплощенные «ветви», похожие на листоватый лишайник, но нижняя сторона листовидной структуры кустистого лишайника имеет тот же цвет, что и верхняя сторона. Листовидные доли листоватого лишайника могут разветвляться, создавая вид кустистого лишайника, но нижняя сторона будет отличаться по цвету от верхней. [35]
Блеск некоторых желеобразных студенистых лишайников создается слизистыми выделениями. [31]
Внутренняя структура
[ редактировать ]
(а) Кора представляет собой внешний слой плотно сплетенных нитей гриба ( гиф ).
(б) В этом слое фотобионтов есть фотосинтезирующие зеленые водоросли.
(в) Рыхло упакованные гифы в мозговом веществе.
(г) Плотно сплетенная нижняя кора
(д) Закрепление гиф, называемых ризинами , где гриб прикрепляется к субстрату.
Лишайник состоит из простого фотосинтезирующего организма, обычно зеленой водоросли или цианобактерии , окруженного нитями гриба. Как правило, большая часть лишайника состоит из переплетенных грибных нитей. [40] но у нитевидных и студенистых лишайников ситуация обратная. [31] Гриб называется микобионт . Фотосинтезирующий организм называется фотобионтом . Водорослевые фотобионты называются фикобионтами . [41] Цианобактерии-фотобионты называются цианобионтами . [41]
Часть лишайника, не участвующая в размножении, «тело» или «вегетативная ткань» лишайника, называется слоевищем . Форма слоевища сильно отличается от любой формы, в которой гриб или водоросль растут отдельно. Таллом состоит из нитей гриба, называемых гифами . Нити растут путем разветвления, а затем снова соединяются, образуя сетку, которая называется « анастомозом ». Сетка грибных нитей может быть плотной или рыхлой.
Обычно грибковая сетка окружает водорослевые или цианобактериальные клетки, часто заключая их в сложные грибковые ткани, уникальные для ассоциаций лишайников. Таллом может иметь или не иметь защитную «кожу» из плотно упакованных грибковых нитей, часто содержащих второй вид грибов, [1] который называется корой. Кустарниковые лишайники имеют один слой коры, охватывающий «ветви». Листовидные лишайники имеют верхнюю кору на верхней стороне «листа» и отдельную нижнюю кору на нижней стороне. Корковые и чешуйчатые лишайники имеют только верхнюю кору, причем «внутренняя часть» лишайника находится в непосредственном контакте с поверхностью, на которой они растут (субстратом ) . Даже если края отслаиваются от субстрата и кажутся плоскими и листовидными, в отличие от листоватых лишайников у них отсутствует нижняя кора. Нитчатая, биссоидная, проказная, [35] студенистые и другие лишайники не имеют коры; другими словами, они экортикаты . [42]
Кустистые, листоватые, корковые и чешуйчатые лишайники обычно имеют до трех различных типов тканей, различающихся разной плотностью грибных нитей. [40] Верхний слой, где лишайник контактирует с окружающей средой, называется корой . [40] Кора состоит из плотно сплетенных, упакованных и склеенных ( агглютинированных ) грибных нитей. [40] Плотная упаковка заставляет кору действовать как защитная «кожа», не допуская проникновения других организмов и снижая интенсивность солнечного света на нижних слоях. [40] Толщина коркового слоя может достигать нескольких сотен микрометров (мкм) (менее миллиметра). [43] Кора может быть дополнительно покрыта эпикортексом, состоящим из секретов, а не клеток, толщиной 0,6–1 мкм у некоторых лишайников . [43] Этот секретный слой может иметь или не иметь поры. [43]
Ниже слоя коры находится слой, называемый фотобионтным слоем или слоем симбионта . [33] [40] Слой симбионта имеет менее плотно упакованные грибковые нити, в которые встроен партнер по фотосинтезу. [40] Менее плотная упаковка обеспечивает циркуляцию воздуха во время фотосинтеза, аналогично анатомии листа. [40] Каждая клетка или группа клеток фотобионта обычно индивидуально обернута гифами, а в некоторых случаях пронизана гаусторием . [31] У накипных и листоватых лишайников водоросли фотобионтного слоя рассеяны среди грибных нитей, постепенно уменьшаясь в нижнем слое. У кустистых лишайников фотобионтный слой резко отличается от нижнего слоя. [31]
Слой под слоем симбионта называется мозговым веществом . Сердцевина менее плотно упакована грибковыми нитями, чем вышележащие слои. У листоватых лишайников, как у Пельтигеры , [36] : 159 обычно существует еще один плотно упакованный слой грибковых нитей, называемый нижней корой. [35] [40] Корнеподобные грибковые структуры, называемые ризинами ( обычно ) [36] : 159 растут из нижней коры, чтобы прикрепить или закрепить лишайник к субстрату. [2] [35] Кустарниковые лишайники имеют единственную кору, охватывающую все «стебли» и «ветви». [36] Медулла является самым нижним слоем и может образовывать ватное белое внутреннее ядро ветвистого слоевища или быть полым. [36] : 159 У накипных и чешуйчатых лишайников нижняя кора отсутствует, а мозговое вещество непосредственно контактирует с субстратом , на котором растет лишайник.
У накипных ареоловых лишайников края ареол отслаиваются от субстрата и кажутся облиственными. У чешуйчатых лишайников облиственной может оказаться и часть слоевища лишайника, не прикрепленная к субстрату. Но в этих облиственных частях отсутствует нижняя кора, отличающая накипные и чешуйчатые лишайники от листоватых лишайников. [40] И наоборот, листовидные лишайники могут казаться приплюснутыми к субстрату, как корковый лишайник, но большая часть листовидных долей может быть приподнята над субстратом, поскольку они отделены от него плотно упакованной нижней корой. [35]
Желатиновый, [36] : 159 Биссоидные и проказные лишайники лишены коры (являются экортикатами ) и обычно имеют только недифференцированную ткань, как будто имеют только слой симбионта. [ нужна ссылка ]
В лишайниках, которые включают как зеленые водоросли, так и цианобактериальные симбионты, цианобактерии могут удерживаться на верхней или нижней поверхности в небольших пустулах, называемых цефалодиями .
Пруиния представляет собой беловатый налет на верхней поверхности. [44] Эпинекральный слой — это «слой роговых мертвых гиф грибов с нечетким просветом внутри или рядом с корой над слоем водорослей». [44]
В августе 2016 года сообщалось, что некоторые макролишайники содержат в своих тканях более одного вида грибов. [1]
Физиология
[ редактировать ]Симбиотические отношения
[ редактировать ]Лишайники — это грибы, которые открыли сельское хозяйство.
— Тревор Говард [45]

Лишайник — сложный организм, возникающий из водорослей или цианобактерий, живущих среди нитей ( гиф ) грибов во взаимовыгодных симбиотических отношениях. Грибы извлекают выгоду из углеводов, вырабатываемых водорослями или цианобактериями посредством фотосинтеза . Водоросли или цианобактерии получают выгоду от защиты от окружающей среды нитями грибов, которые также собирают влагу и питательные вещества из окружающей среды и (обычно) служат для нее якорем. Хотя некоторые фотосинтетические партнеры лишайника могут выжить вне лишайника, симбиотическая ассоциация лишайника расширяет экологический диапазон обоих партнеров, при этом в большинстве описаний ассоциаций лишайников они описываются как симбиотические. Оба партнера получают воду и минеральные питательные вещества в основном из атмосферы, через дождь и пыль. Грибковый партнер защищает водоросль, удерживая воду, служа большей площадью захвата минеральных питательных веществ и, в некоторых случаях, обеспечивает минералы, полученные из субстрата . Если цианобактерии присутствуют в качестве основного партнера или другого симбионта в дополнение к зеленым водорослям, как в некоторых трехчастных лишайниках, они могут фиксировать атмосферный азот , дополняя деятельность зеленых водорослей.
В трех различных линиях грибной партнер независимо потерял митохондриальный ген atp9, который выполняет ключевые функции в производстве митохондриальной энергии. Потеря делает грибы полностью зависимыми от своих симбионтов. [46]
Клетки водорослей или цианобактерий являются фотосинтезирующими и, как и растения, восстанавливают из атмосферы углекислый газ в органические углеродные сахара, чтобы питать обоих симбионтов. Фикобионты (водоросли) производят сахарные спирты ( рибит , сорбит и эритрит ), которые поглощаются микобионтом (грибом). [41] Цианобионты производят глюкозу . [41] Лихенизированные клетки гриба могут заставить фотобионт «вытекать» продукты фотосинтеза, где они затем могут быть поглощены грибом. [12] : 5
Похоже, что многие, а возможно и большинство, лишайников также живут в симбиотических отношениях с отрядом базидиомицетов, называемых Cyphobasidiales . Отсутствие этого третьего партнера может объяснить, почему выращивание лишайников в лаборатории затруднено. Дрожжевые клетки ответственны за формирование характерной коры слоевища лишайника, а также могут иметь значение для его формы. [47]
Сочетание лишайника водоросли или цианобактерии с грибом имеет совершенно другую форму (морфологию), физиологию и биохимию, чем составляющий его гриб, водоросль или цианобактерия, растущий сам по себе, в природе или в культуре. Тело ( таллом ) большинства лишайников отличается от тела гриба или водоросли, растущих отдельно. При выращивании в лаборатории в отсутствие своего фотобионта гриб-лишайник развивается как бесструктурная, недифференцированная масса грибных нитей ( гиф ). При сочетании с фотобионтом в соответствующих условиях возникает его характерная форма, связанная с фотобионтом, в процессе, называемом морфогенезом . [4] В некоторых примечательных случаях один лишайниковый гриб может развиться в две совершенно разные формы лишайника при взаимодействии либо с зелеными водорослями, либо с цианобактериальным симбионтом. Вполне естественно, что эти альтернативные формы сначала считались разными видами, пока не было обнаружено, что они растут слитно. [ нужна ссылка ]
Доказательством того, что лишайники являются примером успешного симбиоза, является тот факт, что лишайники можно найти почти в каждой среде обитания и географическом районе на планете. [17] Два вида двух родов зеленых водорослей встречаются более чем в 35% всех лишайников, но лишь изредка их можно встретить живущими отдельно вне лишайника. [48]
В случае, когда у одного грибного партнера одновременно было два партнера из зеленых водорослей, которые превосходят друг друга в разных климатических условиях, это может указывать на то, что наличие более чем одного партнера по фотосинтезу одновременно может позволить лишайнику существовать в более широком диапазоне сред обитания и географических мест. [17]
По крайней мере, одна форма лишайника, североамериканские бородообразные лишайники, состоит не из двух, а из трех симбиотических партнеров: аскомицетного гриба, фотосинтезирующей водоросли и, что неожиданно, базидиомицетовых дрожжей. [49]
Фикобионты могут производить чистый сахар только с водяным паром. [41] Для того чтобы цианобионты могли фотосинтезировать, слоевище должно быть насыщено жидкой водой. [41]
Водоросли производят сахара, которые поглощаются грибом путем диффузии в специальные грибные гифы, называемые аппрессориями или гаусториями, при контакте со стенками клеток водоросли. [50] Апрессории или гаустории могут производить вещество, которое увеличивает проницаемость стенок клеток водорослей и может проникать через стенки. [50] Водоросли могут обеспечивать грибу до 80% производства сахара. [50]
Экология
[ редактировать ]Ассоциации лишайников могут быть примерами мутуализма или комменсализма , но отношения с лишайниками можно считать паразитическими. [51] при обстоятельствах, когда партнер по фотосинтезу может существовать в природе независимо от партнера-гриба, но не наоборот. Клетки фотобионтов обычно разрушаются в ходе обмена питательных веществ . Ассоциация продолжается, поскольку размножение клеток фотобионтов соответствует скорости их разрушения. [51] Гриб . окружает клетки водорослей [13] часто заключая их в сложные грибковые ткани, уникальные для ассоциаций лишайников. У многих видов гриб проникает в клеточную стенку водорослей. [13] образуя проникающие колышки ( гаустории ), подобные тем, которые производят патогенные грибы , питающиеся хозяином. [34] [52] Цианобактерии в лабораторных условиях могут расти быстрее, когда они одни, а не когда они являются частью лишайника.
Миниатюрная экосистема и теория холобионтов
[ редактировать ]Симбиоз лишайников настолько хорошо сбалансирован, что лишайники сами по себе считаются относительно автономными миниатюрными экосистемами. [17] [18] Считается, что лишайники могут быть еще более сложными симбиотическими системами, включающими нефотосинтезирующие бактериальные сообщества, выполняющие другие функции в качестве партнеров холобионта . [19] [20]
Многие лишайники очень чувствительны к воздействиям окружающей среды и могут быть дешево использованы для [13] оценить загрязнение воздуха , [53] [54] [55] разрушение озона и загрязнение металлами. Лишайники использовались при изготовлении красок , духов ( дубовый мох ), [56] и в традиционной медицине . Некоторые виды лишайников поедаются насекомыми. [13] или более крупных животных, таких как северный олень. [57] Лишайники широко используются в качестве индикаторов окружающей среды или биоиндикаторов. При очень сильном загрязнении воздуха сернистым газом лишайников может не быть; только некоторые зеленые водоросли могут переносить такие условия. Если воздух чистый, то в изобилии появляются кустарниковые, волосатые и листоватые лишайники. Некоторые виды лишайников могут переносить довольно высокие уровни загрязнения и обычно встречаются в городских районах, на тротуарах, стенах и коре деревьев. Наиболее чувствительные лишайники — кустарниковые и листовые, тогда как наиболее устойчивые лишайники имеют корковый вид. С момента индустриализации многие кустарниковые и листовые лишайники, такие как виды Ramalina , Usnea и Lobaria , имеют очень ограниченный ареал, часто ограничиваясь районами с самым чистым воздухом.
Лихеноподобные грибы
[ редактировать ]Некоторые грибы обитают на лишайниках только как облигатные паразиты . Их называют лихеноподобными грибами , и они представляют собой разные виды грибов, живущих внутри лишайника; таким образом, они не считаются частью лишайника. [58]
Реакция на воду
[ редактировать ]Влага делает кору более прозрачной. [12] : 4 Таким образом, водоросли могут осуществлять фотосинтез при наличии влаги и защищены в другое время. Когда кора более прозрачна, водоросли видны более четко, а лишайник выглядит зеленее.
Метаболиты, структуры метаболитов и биологическая активность
[ редактировать ]Лишайники могут проявлять интенсивную антиоксидантную активность. [59] [60] Вторичные метаболиты часто откладываются в виде кристаллов в апопласте . [61] Считается, что вторичные метаболиты играют роль в предпочтении одних субстратов другим. [61]
Темпы роста
[ редактировать ]Лишайники часто имеют регулярный, но очень медленный темп роста – менее миллиметра в год.
У накипных лишайников наиболее активный рост происходит в зоне вдоль края. [36] : 159 Большинство корковых лишайников вырастают всего на 1–2 мм в диаметре в год.
Продолжительность жизни
[ редактировать ]Лишайники могут быть долгожителями , а некоторые из них считаются одними из старейших живых организмов. [4] [21] Продолжительность жизни трудно измерить, поскольку неточно определить «один и тот же» отдельный лишайник. [62] Лишайники растут путем вегетативного отрыва части, которая может определяться, а может и не определяться как «один и тот же» лишайник, а два лишайника могут сливаться, становясь затем «одним и тем же» лишайником. [62] Возраст одного экземпляра Rhizocarpon geographicum на Восточном Баффиновом острове оценивается в 9500 лет. [63] [64] Талломам и Rhizocarpon geographicum Rhizocarpon eupetraeoides / inarense в центральном хребте Брукс на севере Аляски присвоен максимально возможный возраст 10 000–11 500 лет. [65] [66]
Реакция на экологический стресс
[ редактировать ]В отличие от простого обезвоживания у растений и животных, лишайники могут испытывать полную потерю воды в организме в засушливые периоды. [13] Лишайники способны переносить чрезвычайно низкие уровни содержания воды ( пойкилогидридные ). [67] : 5–6 Они быстро впитывают воду, когда она снова становится доступной, становясь мягкими и мясистыми. [13]
В ходе испытаний лишайник выжил и показал замечательные результаты по адаптационной способности фотосинтетической активности в течение моделирования 34 дней в марсианских условиях в Лаборатории моделирования Марса (MSL), поддерживаемой Немецким аэрокосмическим центром (DLR). [68] [69]
Европейское космическое агентство обнаружило, что лишайники могут выжить в космосе без защиты. В ходе эксперимента, проведенного Леопольдо Санчо из Мадридского университета Комплутенсе, два вида лишайников — Rhizocarpon geographicum и Rusavskia elegans — были запечатаны в капсулу и запущены на российской ракете «Союз» 31 мая 2005 года. На орбите капсулы были открыты и лишайники подвергались прямому воздействию космического вакуума с его широко колеблющимися температурами и космическим излучением. Через 15 дней лишайники были возвращены на землю, и было обнаружено, что их способность к фотосинтезу не изменилась. [70] [71]
Размножение и распространение
[ редактировать ]Вегетативное размножение
[ редактировать ]
Многие лишайники размножаются бесполым путем: либо путем отрыва части и роста самостоятельно ( вегетативное размножение ), либо путем распространения диаспор , содержащих несколько клеток водорослей, окруженных грибковыми клетками. [2] Из-за относительной недифференцированности слоевища грань между образованием диаспор и вегетативным размножением часто размыта. Кустистые лишайники могут фрагментироваться, и из фрагмента могут вырасти новые лишайники ( вегетативное размножение ). Многие лишайники при высыхании распадаются на фрагменты, разносясь под действием ветра, чтобы возобновить рост при возвращении влаги. [72] [73] Соредии (единственное число: «соредий») представляют собой небольшие группы клеток водорослей, окруженные грибковыми нитями, которые образуют структуры, называемые соралиями, из которых соредии могут разноситься ветром. [2] Изидии (единственное число: «исидий») представляют собой разветвленные, колючие, удлиненные выросты слоевища, которые отрываются при механическом распространении. [2] Размножения лишайников ( диаспоры ) обычно содержат клетки обоих партнеров, хотя грибковые компоненты так называемых «маргинальных видов» вместо этого полагаются на клетки водорослей, рассеянные «основными видами». [74]
Половое размножение
[ редактировать ]
Структуры, участвующие в размножении, часто выглядят как диски, выпуклости или волнистые линии на поверхности слоевища. [12] : 4 Хотя утверждалось, что половое размножение у фотобионтов отвергается, существуют убедительные доказательства, свидетельствующие о мейотической активности (половом размножении) у Trebouxia . [75] [76] Многие лишайниковые грибы, как и другие грибы, размножаются половым путем, образуя споры, образующиеся в результате мейоза и слияния гамет. После распространения такие грибные споры должны встретиться с совместимым партнером-водорослем, прежде чем сможет сформироваться функциональный лишайник.
Некоторые лишайниковые грибы относятся к типу Basidiomycota ( базидиолишайники ) и образуют грибовидные репродуктивные структуры, напоминающие структуры своих нелихенизированных родственников.
Большинство лишайниковых грибов относятся к аскомицетам ( асколишенам ). У асколишайников споры образуются в спорообразующих структурах, называемых аскоматами . [12] Наиболее распространенными типами аскомат являются апотеций (множественное число: апотеции) и перитеций (множественное число: перитеции). [12] : 14 Апотеции обычно представляют собой чашечки или пластинчатые диски, расположенные на верхней поверхности слоевища лишайника. Когда апотеции имеют форму сегментов волнистой линии, а не дисков, их называют лиреллами . [12] : 14 Перитеции имеют форму колб, погруженных в ткань слоевища лишайника, имеющую небольшое отверстие для выхода спор из колбы, и выглядят как черные точки на поверхности лишайника. [12] : 14
Три наиболее распространенных типа споровых тел — это приподнятые диски, называемые апотециями (единственное число: апотеций), бутылкообразные чашечки с небольшим отверстием наверху, называемые перитециями (единственное число: перитеций), и пикниды (единственное число: пикнидий), имеющие форму перитеция, но без asci ( аска — это структура, которая содержит и высвобождает половые споры грибов Ascomycota ) . [77]
Апотеций имеет слой открытых спорообразующих клеток, называемый асками (единственное число: аски), и обычно его цвет отличается от ткани слоевища. [12] : 14 Когда апотеций имеет внешний край, этот край называется эксциплиной . [12] : 14 Когда эксциплина имеет цвет, похожий на окрашенную ткань слоевища, апотеций или лишайник называется леканорином , что означает сходство с представителями рода Lecanora . [12] : 14 Когда эксциплина чернеет, как углерод, ее называют лецидином, что означает сходство с представителями рода Lecidea . [12] : 14 Когда край бледный или бесцветный, его называют биаторином . [12] : 14

« Подеций » (множественное число: podetia ) — лихенизированная стебельчатая структура плодового тела, поднимающаяся из слоевища, связанная с некоторыми грибами, образующими грибной апотеций . [33] Поскольку подеции являются частью репродуктивной ткани, они не считаются частью основного тела (таллома), но могут быть визуально заметными. [33] Подеций может быть разветвленным, иногда чашевидным. Обычно они несут грибные пикниды или апотеции , или и то, и другое. [33] Многие лишайники имеют апотеции , видимые невооруженным глазом. [2]
Большинство лишайников образуют обильные половые структуры. [78] Многие виды, по-видимому, распространяются только половыми спорами. [78] Например, корковые лишайники Graphis scripta и Ochrolechia parella не образуют симбиотических вегетативных размножений. Вместо этого лишайниковообразующие грибы этих видов размножаются половым путем путем самооплодотворения (т. е. они гомоталличны ). Эта система разведения может обеспечить успешное размножение в суровых условиях. [78]
Мазаедия (единственное число: mazaedium) представляет собой апотеции в форме портновской булавки у булавовидных лишайников , где плодовое тело представляет собой коричневую или черную массу рыхлых аскоспор, заключенных в чашеобразную эксциплию, расположенную на вершине крошечного стебля. [12] : 15
Таксономия и классификация
[ редактировать ]Лишайники классифицируются по грибковому компоненту. Видам лишайников присвоено то же научное название ( биномиальное название ), что и видам грибов в лишайнике. Лишайники интегрируются в схемы классификации грибов. Водоросль имеет собственное научное название, не имеющее никакого отношения к лишайнику или грибу. [79] Известно около 20 000 видов лишайников. [80] [81] а систематики подсчитали, что общее количество видов лишайников (включая еще не открытые) может достигать 28 000. [82] Около 20% известных видов грибов связаны с лишайниками. [50]
« Лихенизированный гриб » может относиться ко всему лишайнику или только к грибу. Без контекста это может вызвать путаницу. Определенный вид грибов может образовывать лишайники с разными видами водорослей, в результате чего возникают, казалось бы, разные виды лишайников, но которые по-прежнему классифицируются (по состоянию на 2014 год) как один и тот же вид лишайников. [83]
Раньше некоторые систематики лишайников помещали лишайники в отдельный отдел Mycophycophyta , но эта практика больше не принимается, поскольку компоненты принадлежат к разным линиям . Ни асколишайники, ни базидиолишайники не образуют монофилетических линий в своих соответствующих типах грибов, но они образуют несколько основных групп, образующих исключительно или преимущественно лишайники, внутри каждого типа. [84] Еще более необычным, чем базидиолишайники, является гриб Geosiphonpyriforme , представитель Glomeromycota , который уникален тем, что содержит внутри своих клеток цианобактериальный симбионт. Геосифон обычно не считают лишайником, и его своеобразный симбиоз долгие годы не признавался. Род более близок к эндомикоризным родам. Грибы Verrucariales вместе с бурыми водорослями Petroderma maculiforme образуют морские лишайники . [85] и имеют симбиотические отношения с морскими водорослями (такими как водоросли ) и Blidingia minima , где водоросли являются доминирующими компонентами. Считается, что грибы помогают водорослям противостоять высыханию на воздухе. [86] [87] Кроме того, лишайники также могут использовать желто-зеленые водоросли ( Heterococcus ) в качестве своего симбиотического партнера. [88]
Лишайники на протяжении всей истории независимо возникали из грибов, связанных с водорослями и цианобактериями. [89]

Грибы
[ редактировать ]Грибковый компонент лишайника называется микобионтом . Микобионт может быть аскомицетом или базидиомицетом . [15] Сопутствующие лишайники называются либо асколишайниками , либо базидиолишайниками соответственно. Жизнь в качестве симбионта в лишайнике, по-видимому, является для гриба успешным способом получения необходимых питательных веществ, поскольку около 20% всех видов грибов приобрели этот образ жизни. [90]
Талломы, продуцируемые данным грибным симбионтом с разными партнерами, могут быть сходными. [ нужна ссылка ] и вторичные метаболиты идентичны, [ нужна ссылка ] указывая [ нужна ссылка ] что грибу принадлежит доминирующая роль в определении морфологии лишайника. Но один и тот же микобионт с разными фотобионтами может давать и совершенно разные формы роста. [83] Известны лишайники, у которых один гриб связан с двумя и даже тремя видами водорослей.
Хотя каждый слоевище лишайника обычно выглядит однородным, некоторые данные позволяют предположить, что грибной компонент может состоять из более чем одной генетической особи этого вида. [ нужна ссылка ]
Два или более вида грибов могут взаимодействовать, образуя один и тот же лишайник. [91]
В следующей таблице перечислены отряды и семейства грибов, в которые входят виды, образующие лишайники.
Фотобионты
[ редактировать ]
партнер Фотосинтетический лишайника называется фотобионтом . Фотобионты лишайников происходят от множества простых прокариотических и эукариотических организмов. У большинства лишайников фотобионт — зеленая водоросль ( Chlorophyta ) или цианобактерия . В некоторых лишайниках присутствуют оба типа; в таких случаях основным партнером обычно являются водоросли, а цианобактерии располагаются в загадочных карманах. [92] Водорослевые фотобионты называются фикобионтами , а цианобактериальные фотобионты — цианобионтами . [41] Около 90% всех известных лишайников имеют фикобионты, а около 10% — цианобионты. [41] Около 100 видов партнеров по фотосинтезу из 40 [41] роды и пять отдельных классов (прокариотические: Cyanophyceae ; эукариотические: Trebouxiophyceae , Phaeophyceae , Chlorophyceae ) связаны с грибами, образующими лишайники. Было обнаружено, что [93]
Обычные водорослевые фотобионты из родов Trebouxia , Trentepohlia , Pseudotrebouxia или Myrmecia . Trebouxia — наиболее распространенный род зеленых водорослей лишайников, встречающийся примерно в 40% всех лишайников. «Требуксиоид» означает либо фотобионт, принадлежащий к роду Trebouxia , либо напоминающий представителя этого рода и, следовательно, предположительно принадлежащий к классу Trebouxiophyceae . [33] Второй по распространенности род зеленых водорослей — Trentepohlia . [34] Всего известно около 100 видов эукариот, встречающихся в виде фотобионтов в лишайниках. Все водоросли, вероятно, способны существовать самостоятельно как в природе, так и в лишайнике. [91]
« Цианолишайник » — это лишайник, основным фотосинтетическим компонентом которого является цианобактерия (фотобионт). [94] Большинство цианолишайников также являются асколишайниками, но некоторые базидиолишайники, такие как Dictyonema и Acantholichen, имеют в качестве партнера цианобактерии. [95]
Наиболее часто встречающийся род цианобактерий — Nostoc . [91] Другой [34] распространенные фотобионты цианобактерий происходят из Scytonema . [15] Многие цианолишайники маленькие и черные, а является известняк . субстратом [ нужна ссылка ] Другая группа цианолишайников — желейные лишайники родов Collema или Leptogium — студенистые и обитают на влажных почвах. Другая группа крупных и листоватых видов, включая Peltigera , Lobaria и Degelia , имеет серо-голубой цвет, особенно в сыром или мокром состоянии. Многие из них характеризуют сообщества Лобариона в районах с большим количеством осадков на западе Британии, например, в кельтских тропических лесах . Штаммы цианобактерий, обнаруженные в различных цианолишайниках, часто тесно связаны друг с другом. [96] Они отличаются от наиболее близкородственных свободноживущих штаммов. [96]
Ассоциация лишайников представляет собой тесный симбиоз. Оно расширяет экологический ареал обоих партнеров, но не всегда является обязательным для их роста и размножения в природных средах, поскольку многие водорослевые симбионты могут жить независимо. Ярким примером является водоросль Trentepohlia , которая образует популяции оранжевого цвета на стволах деревьев и подходящих скалах. Размножения лишайников ( диаспоры ) обычно содержат клетки обоих партнеров, хотя грибковые компоненты так называемых «маргинальных видов» вместо этого полагаются на клетки водорослей, рассеянные «основными видами». [74]
Один и тот же вид цианобионтов может встречаться в ассоциации с разными видами грибов в качестве партнеров лишайников. [97] Один и тот же вид фикобионтов может встречаться в сочетании с разными видами грибов в качестве партнеров лишайников. [41] В одном слоевище может присутствовать более одного фикобионта. [41]
В одном лишайнике может присутствовать несколько генотипов водорослей . [98] [99] Эти множественные генотипы могут лучше реагировать на адаптацию к изменениям окружающей среды и позволить лишайнику обитать в более широком диапазоне сред. [100]
Споры по поводу метода классификации и названий видов.
[ редактировать ]Известно около 20 000 видов лишайников . [15] Но то, что подразумевается под «видами», отличается от того, что подразумевается под биологическими видами у растений, животных или грибов, где принадлежность к одному и тому же виду подразумевает наличие общей предковой линии . [15] Поскольку лишайники представляют собой комбинации представителей двух или даже трех разных биологических царств , эти компоненты должны иметь различную наследственную линию друг от друга. По соглашению лишайники все равно называются «видами» и классифицируются по видам грибов, а не по видам водорослей или цианобактерий. Лишайникам дано то же научное название ( биномиальное название ), что и грибу в них, что может вызвать некоторую путаницу. Водоросль носит собственное научное название, не имеющее никакого отношения к названию лишайника или гриба. [79]
В зависимости от контекста «лихенизированный гриб» может относиться ко всему лишайнику или к грибу, когда он находится в лишайнике, который можно выращивать в культуре изолированно от водорослей или цианобактерий. Некоторые водоросли и цианобактерии встречаются в природе вне лишайника. Грибковый, водорослевый или цианобактериальный компонент лишайника можно выращивать отдельно в культуре. Растущие сами по себе грибы, водоросли или цианобактерии обладают совершенно иными свойствами, чем лишайники. Свойства лишайника, такие как форма роста, физиология и биохимия, сильно отличаются от сочетания свойств гриба и водорослей или цианобактерий.
Один и тот же гриб, растущий в сочетании с разными водорослями или цианобактериями, может давать лишайники, которые сильно различаются по большинству свойств и соответствуют критериям, не связанным с ДНК, для обозначения разных «видов». Исторически эти разные комбинации классифицировались как разные виды. Когда с помощью современных методов ДНК идентифицируется один и тот же гриб, эти явно разные виды переклассифицируются как один и тот же вид в соответствии с действующим (2014 г.) соглашением о классификации по грибковому компоненту. Это привело к дискуссиям по поводу этого соглашения о классификации. Эти, казалось бы, разные «виды» имеют свою независимую эволюционную историю. [2] [83]
Также ведутся споры относительно целесообразности присвоения одного и того же биномиального названия грибу и лишайнику, сочетающему этот гриб с водорослью или цианобактерией ( синекдоха ). Это особенно актуально, когда сочетание одного и того же гриба с разными водорослями или цианобактериями приводит к совершенно разным лишайниковым организмам, которые можно было бы считать разными видами по любым критериям, кроме ДНК грибкового компонента. Если бы весь лишайник, произведенный одним и тем же грибом, растущим в сочетании с разными водорослями или цианобактериями, был классифицирован как разные «виды», количество «видов лишайников» было бы больше.
Разнообразие
[ редактировать ]Наибольшее количество лихенизированных грибов встречается у Ascomycota , около 40% видов образуют такую ассоциацию. [79] Некоторые из этих лихенизированных грибов встречаются в отрядах с нелихенизированными грибами, живущими как сапротрофы или паразиты растений (например, Leotiales , Dothideales , Pezizales ). Другие лишайниковые грибы встречаются только в пяти отрядах , все представители которых имеют эту привычку (отряды Graphidales , Gyalectales , Peltigerales , Pertusariales и Teloschistales ). В целом около 98% лишайников имеют аскомицетный микобионт. [101] После Ascomycota наибольшее количество лихенизированных грибов встречается в неопределенных грибах несовершенных , всеобъемлющей категории грибов, у которых половая форма размножения никогда не наблюдалась. [ нужна ссылка ] Сравнительно немногие базидиомицеты лихенизированы, но к ним относятся опята , например виды Lichenomphalia , клавариоидные грибы , например виды Multiclavula , и кортицоидные грибы , например виды Dictyonema .
Методы идентификации
[ редактировать ]Для идентификации лишайников используются формы роста, микроскопия и реакции на химические тесты.
Результат «теста на палладий» называется «палладий», что также используется как аббревиатура химического вещества, использованного в тесте, пара-фенилендиамина . [33] Если капля на лишайник окрасит участок от ярко-желтого до оранжевого, это поможет идентифицировать его как принадлежность к роду Cladonia или Lecanora . [33]
Эволюция и палеонтология
[ редактировать ]Летопись окаменелостей лишайников скудна. [102] Экстремальные среды обитания, в которых доминируют лишайники, такие как тундра, горы и пустыни, обычно не способствуют образованию окаменелостей. [102] [103] В янтаре заключены окаменелые лишайники. Окаменевшая Анзия найдена в кусках янтаря в Северной Европе и датируется примерно 40 миллионами лет назад. [104] Фрагменты лишайников также находят в пластах ископаемых листьев, таких как Lobaria из округа Тринити в северной Калифорнии, США, относящихся к раннему и среднему миоцену . [105]
Самый старый ископаемый лишайник, в котором были обнаружены оба симбиотических партнера, - это Winfrenatia , ранний симбиоз сзигомицетов ( Glomeromycotan ), который, возможно, включал контролируемый паразитизм. [ нужна ссылка ] перминерализован в Рини-Черте в Шотландии, датируемом ранним девоном , около 400 миллионов лет назад. [106] Немного более древнее ископаемое Spongiophyton также было интерпретировано как лишайник по морфологическим признакам. [107] и изотопный [108] основания, хотя изотопная основа явно шаткая. [109] Показано, что силурийско - девонские окаменелости Nematothallus [110] и прототакситы [111] были лихенизированы. Таким образом, лихенизированные Ascomycota и Basidiomycota были компонентом раннесилурийско - девонских наземных экосистем. [112] [113] Новые исследования показывают, что лишайник развился после эволюции наземных растений. [114]
Исходное экологическое состояние как Ascomycota , так и Basidiomycota, вероятно, было сапробизмом , и независимые события лихенизации могли происходить несколько раз. [115] [116] В 1995 году Гаргас и его коллеги предположили, что существует как минимум пять независимых источников лихенизации; три у базидиомицетов и не менее двух у аскомицетов. [117] Лутзони и др. (2001) предполагают, что лихенизация, вероятно, развилась раньше и сопровождалась множественными независимыми потерями. Некоторые грибы, не образующие лишайников, возможно, вторично утратили способность образовывать лишайниковые ассоциации. В результате лихенизация стала рассматриваться как весьма успешная стратегия питания. [118] [119]
Лихенизированная Glomeromycota могла распространиться еще в докембрий. Лишайниковые окаменелости, состоящие из кокковидных клеток ( цианобактерий ?) и тонких нитей (мукоромикотинан Glomeromycota ?), перминерализованы в морских фосфоритах формации Доушантуо на юге Китая. Считается, что возраст этих окаменелостей составляет от 551 до 635 миллионов лет или эдиакарского периода . [120] Эдиакарские акритархи также имеют много общего с пузырьками и спорами Glomeromycotan . [121] Также утверждалось, что эдиакарские окаменелости , включая дикинсонию , [122] были лишайники, [123] хотя это утверждение спорно. [124] Эндосимбиотическая Glomeromycota, сравнимая с современным Geosiphon, может возникнуть еще в протерозое в виде Horodyskia возрастом 1500 миллионов лет. [125] возрастом 2200 миллионов лет и Дискагма . [126] Обнаружение этих окаменелостей позволяет предположить, что грибы развили симбиотические партнерства с фотоавтотрофами задолго до эволюции сосудистых растений, хотя гипотеза эдиакарского лишайника в значительной степени отвергается из-за неподходящего определения лишайников, основанного на тафономии и экологии субстрата. [127] Однако в исследовании 2019 года, проведенном тем же ученым, который отверг гипотезу эдиакарского лишайника, Нельсеном, использовались новые филогении, калиброванные по времени, чтобы прийти к выводу, что не существует никаких доказательств существования лишайников до существования сосудистых растений. [128]
Lecanoromycetes, один из наиболее распространенных классов грибов, образующих лишайники, отделился от своего предка, который, возможно, также образовывал лишайники, около 258 миллионов лет назад, в конце палеозоя. Однако близкородственная клада Euritiomycetes, по-видимому, начала образовывать лишайники только 52 миллиона лет назад, в ранний кайнозойский период. [129]
Экология и взаимодействие с окружающей средой
[ редактировать ]Субстраты и среда обитания
[ редактировать ]
Лишайники растут на самых разных субстратах и средах обитания, включая некоторые из самых экстремальных условий на Земле. [130] Они в изобилии растут на коре, листьях и свисают с ветвей эпифитов в тропических лесах и лесах умеренного пояса . Они растут на голых камнях, стенах, надгробиях, крышах и открытых поверхностях почвы. Они могут выжить в самых экстремальных условиях на Земле: арктической тундре , жарких сухих пустынях , скалистых побережьях и токсичных отвалах шлака . Они могут жить внутри твердых пород, растущих между зернами, а также в почве как часть биологической почвенной корки в засушливых местах обитания, таких как пустыни. Некоторые лишайники ни на чем не растут, доживая свою жизнь, разносясь по окружающей среде. [2]
Растущие на минеральных поверхностях некоторые лишайники медленно разлагают свой субстрат, химически разлагая и физически разрушая минералы, способствуя процессу выветривания , в результате которого горные породы постепенно превращаются в почву. Хотя этот вклад в выветривание обычно незначителен, он может вызвать проблемы для конструкций из искусственного камня. Например, на Национальном мемориале на горе Рашмор постоянно наблюдается проблема роста лишайников , что требует привлечения реставраторов-альпинистов для очистки памятника. [131]
Лишайники не являются паразитами растений, на которых растут, а лишь используют их в качестве субстрата. Грибы некоторых видов лишайников могут «поглотить» водоросли других видов лишайников. [13] [132] Лишайники производят себе пищу из своих фотосинтезирующих частей и поглощая минералы из окружающей среды. [13] Лишайники, растущие на листьях, могут выглядеть как паразиты на листьях, но это не так. Некоторые лишайники Diploschistes паразитируют на других лишайниках. Diploschistes muscorum начинает свое развитие в ткани хозяина вида Cladonia . [52] : 30 [34] : 171

В арктической тундре лишайники вместе со мхами и печеночниками составляют большую часть напочвенного покрова , который помогает изолировать почву и может служить кормом для пасущихся животных. Примером может служить « оленьий мох », который представляет собой лишайник, а не мох. [13]
Известны только два вида постоянно погруженных лишайников; Hydrothyria venosa встречается в пресной воде, а Verrucaria serpuloides встречается в морской среде. [133]
Корковый лишайник, растущий на камнях, называется камнеломным . [33] [36] : 159 Корковые лишайники, растущие на камнях, относятся к эпилитным , а те, которые растут погруженными внутрь скалы, растущими между кристаллами, оставляя на воздухе только свои плодовые тела, называются эндолитными лишайниками . [32] [36] : 159 [94] Накипистый лишайник, растущий на коре, называется корковым лишайником . [36] : 159 Лишайник, растущий на древесине, с которой снята кора, называется древовидным лишайником . [42] Лишайники, которые растут погруженными в ткани растений, называются эндофлоидными лишайниками или эндофлоидными лишайниками . [32] [36] : 159 Лишайники, использующие листья в качестве субстрата, независимо от того, находятся ли они на дереве или на земле, называются эпифиллистными или листовидными . [41] террикольный лишайник В качестве субстрата на почве растет . Многие чешуйчатые лишайники террицветны. [36] : 159 Пупковидные лишайники — листовидные лишайники, прикрепляющиеся к субстрату только в одной точке. [32] Бродячий лишайник вообще не прикрепляется к субстрату и живет своей жизнью, раздуваясь ветром.
Лишайники и почвы
[ редактировать ]В дополнение к различным физическим механизмам, с помощью которых лишайники разрушают необработанный камень, исследования показывают, что лишайники химически воздействуют на камень, внося в окружающую среду новые хелатные минералы. Вещества, выделяемые лишайниками, известные своей сильной способностью связывать и изолировать металлы, а также обычное образование новых минералов, особенно оксалатов металлов , а также свойства субстратов, которые они изменяют, все это подчеркивает важную роль, которую лишайники играют в процессе химическое выветривание . [134] Со временем эта деятельность создает из камня новую плодородную почву.
Лишайники могут играть важную роль в обеспечении азота почв в некоторых пустынях, поскольку их поедают вместе с каменным субстратом улитки, которые затем испражняются, отдавая азот в почву. [135] Лишайники помогают связывать и стабилизировать почвенный песок в дюнах. [2] В пустынях и полузасушливых районах лишайники являются частью обширных живых биологических почвенных корок , необходимых для поддержания структуры почвы. [2]
Экологические взаимодействия
[ редактировать ]Лишайники — виды-первопроходцы , одни из первых живых существ, которые растут на голых камнях или на участках, лишенных жизни в результате стихийного бедствия. [2] Лишайникам, возможно, придется конкурировать с растениями за доступ к солнечному свету, но из-за их небольшого размера и медленного роста они процветают в местах, где высшим растениям трудно расти. Лишайники часто первыми поселяются в местах, где нет почвы, составляя единственную растительность в некоторых экстремальных условиях, например, на высоких горах и в высоких широтах. [136] Некоторые выживают в суровых условиях пустынь, другие – на мерзлой почве Арктики. [137]
Основным экофизиологическим преимуществом лишайников является то, что они являются пойкилогидными ( пойкило — изменчивый, гидрический — относящийся к воде), что означает, что, хотя они мало контролируют состояние своей гидратации, они могут переносить нерегулярные и длительные периоды сильного высыхания . Подобно некоторым мхам , печеночникам , папоротникам и некоторым воскресным растениям , после высыхания лишайники впадают в метаболическую суспензию или стазис (известный как криптобиоз ), при котором клетки лишайников-симбионтов обезвоживаются до такой степени, что прекращается большая часть биохимической активности. В этом криптобиотическом состоянии лишайники могут пережить более экстремальные температуры, радиацию и засуху в суровых условиях, в которых они часто обитают.

У лишайников нет корней, и им не нужно использовать непрерывные резервуары с водой, как большинству высших растений, поэтому они могут расти в местах, недоступных для большинства растений, таких как голые камни, стерильная почва или песок, а также различные искусственные конструкции, такие как стены, крыши. и памятники. Многие лишайники растут также как эпифиты ( эпи — на поверхности, фит — растение) на растениях, особенно на стволах и ветвях деревьев. Растущие на растениях лишайники не являются паразитами ; они не потребляют какие-либо части растения и не отравляют его. Лишайники производят аллелопатические химические вещества, подавляющие рост мхов. Некоторые наземные лишайники, например представители подрода Cladina (лишайники северного оленя), производят аллелопатические химические вещества, которые выщелачиваются в почву и подавляют прорастание семян, ели и других растений. [138] Стабильность (то есть долговечность) их субстрата является основным фактором обитания лишайников. Большинство лишайников растут на устойчивых поверхностях камней или коре старых деревьев, но многие другие растут на почве и песке. В этих последних случаях лишайники часто являются важной частью стабилизации почвы; действительно, в некоторых пустынных экосистемах семена сосудистых (высших) растений не могут прижиться, за исключением тех мест, где корки лишайников стабилизируют песок и помогают удерживать воду.
Лишайники могут поедаться некоторыми животными, например северными оленями , обитающими в арктических регионах. Личинки . ряда видов чешуекрылых питаются исключительно лишайниками К ним относятся обыкновенный лакей и мраморная красавица . В них очень мало белка и много углеводов, что делает их непригодными для некоторых животных. Северная летяга использует его для гнездования, еды и зимней воды.
Последствия загрязнения воздуха
[ редактировать ]
Если лишайники постоянно подвергаются воздействию загрязнителей воздуха, без каких-либо лиственных частей, они не могут избежать накопления загрязнителей. Также не имея устьиц и кутикулы , лишайники могут поглощать аэрозоли и газы всей поверхностью слоевища, откуда они могут легко диффундировать в слой фотобионта. [139] Поскольку у лишайников нет корней, основным источником большинства элементов для них является воздух, и поэтому уровни элементов в лишайниках часто отражают накопленный состав окружающего воздуха. Процессы, посредством которых происходят атмосферные осаждения, включают туман и росу , газообразное поглощение и сухое осаждение. [140] Следовательно, экологические исследования лишайников подчеркивают их возможность использования в качестве эффективных биомониторов качества атмосферы. [139]
Не все лишайники одинаково чувствительны к загрязнителям воздуха , поэтому разные виды лишайников демонстрируют разные уровни чувствительности к конкретным загрязнителям атмосферы. [141] Чувствительность лишайника к загрязнению воздуха напрямую связана с энергетическими потребностями микобионта, так что чем сильнее зависимость микобионта от фотобионта, тем чувствительнее лишайник к загрязнению воздуха. [142] При воздействии загрязнения воздуха фотобионт может использовать метаболическую энергию для восстановления своих клеточных структур, которая в противном случае использовалась бы для поддержания его фотосинтетической активности, в результате чего для микобионта остается меньше метаболической энергии. Изменение баланса фотобионта и микобионта может привести к разрыву симбиотической ассоциации. Следовательно, упадок лишайников может быть результатом не только накопления токсичных веществ, но и изменения запасов питательных веществ, в результате которых один симбионт отдает предпочтение другому. [139]
Это взаимодействие между лишайниками и загрязнением воздуха использовалось как средство мониторинга качества воздуха с 1859 года, а более систематические методы были разработаны Уильямом Нюландером в 1866 году. [2]
Использование человеком
[ редактировать ]
Еда
[ редактировать ]Лишайники едят представители разных культур по всему миру. Хотя некоторые лишайники едят только во время голода , другие являются основным продуктом питания или даже деликатесом . При употреблении в пищу лишайников часто встречаются два препятствия: полисахариды лишайника обычно не перевариваются человеком, а лишайники обычно содержат слаботоксичные вторичные соединения , которые следует удалить перед едой. Очень немногие лишайники ядовиты, но токсичными являются лишайники с высоким содержанием вульпиновой или усниновой кислоты . [143] Большинство ядовитых лишайников имеют желтый цвет. [ нужна ссылка ]
В прошлом исландский мох ( Cetraria Islandica ) был важным источником пищи для людей в Северной Европе, его готовили в виде хлеба, каши, пудинга, супа или салата. Bryoria fremontii (съедобный лишайник из конского волоса) был важным продуктом питания в некоторых частях Северной Америки, где его обычно готовили в яме . Северные народы Северной Америки и Сибири традиционно едят частично переваренный олений лишайник ( Cladina spp.) после того, как извлекли его из рубца убитого северного оленя или карибу. Каменный рубец ( Umbilicaria spp. и Lasalia spp.) — это лишайник, который часто используется в качестве экстренной пищи в Северной Америке, а один вид, Umbilicaria esculenta ( по-японски iwatake ), используется в различных традиционных корейских и японских блюдах. .
Лихенометрия
[ редактировать ]

Лихенометрия — это метод, используемый для определения возраста обнаженных поверхностей горных пород на основе размера слоевищ лишайников. Представленный Бешелем в 1950-х годах, [144] этот метод нашел множество применений. используется в археологии , палеонтологии и геоморфологии . Он использует предполагаемую регулярную, но медленную скорость роста лишайников для определения возраста обнаженной породы . [38] : 9 [145] Измерение диаметра (или другое измерение размера) самого крупного лишайника данного вида на поверхности камня указывает на продолжительность времени с момента первого обнажения поверхности камня. Лишайник может сохраняться на старых скалах до [ нужна ссылка ] 10 000 лет, что обеспечивает максимальный возраст метода, хотя он наиболее точен (с погрешностью 10%) при применении к поверхностям, подвергавшимся воздействию менее 1000 лет. [146] Лихенометрия особенно полезна для датирования поверхностей возрастом менее 500 лет, поскольку методы радиоуглеродного датирования в этот период менее точны. [147] Для лихенометрии чаще всего используются лишайники родов Rhizocarpon (например, виды Rhizocarpon geographicum , картографический лишайник) и Xanthoria .
Биодеградация
[ редактировать ]Было доказано, что лишайники разлагают полиэфирные смолы , что можно увидеть на археологических раскопках в римском городе Баэло-Клаудия в Испании. [148] Лишайники могут накапливать несколько загрязнителей окружающей среды, таких как свинец, медь и радионуклиды . [149] некоторые виды лишайников, такие как Parmelia sulcata (называемый, среди прочего, щитовидным лишайником) и Lobaria pulmonaria (легочный лишайник), а также многие представители Cladonia рода Было показано, что , продуцируют сериновые протеазы , способные расщеплять патогенные формы лишайников. прионный белок (PrP), который может быть полезен при очистке загрязненных водоемов окружающей среды. [150] [151] [152]
Красители
[ редактировать ]Многие лишайники производят вторичные соединения, в том числе пигменты , которые уменьшают вредное количество солнечного света, и мощные токсины, отпугивающие травоядных или убивающие бактерии. Эти соединения очень полезны для идентификации лишайников и имели экономическое значение в качестве красителей, таких как жвачка, или примитивных антибиотиков .
Индикатор pH (который может указывать на кислые или основные вещества), называемый лакмусом, представляет собой краситель, извлекаемый из лишайника Roccellatinctoria («трава красильщика»). [153] путем кипячения. Оно дало название известной лакмусовой бумажке .
Традиционные красители Шотландского Хайленда для твида Харриса. [2] и другие традиционные ткани были сделаны из лишайников, в том числе оранжевого Xanthoria parietina («обыкновенный оранжевый лишайник») и серой листовой Parmelia saxatilis, распространенной на камнях и известной в просторечии как «кроттл».
Есть сообщения, датируемые почти 2000 лет назад, о том, что лишайники использовались для изготовления фиолетовых и красных красок. [154] Большое историческое и коммерческое значение имеют лишайники, принадлежащие к семейству Roccellaceae , обычно называемые орхеллами или орхиллами. Орсеин и другие красители лишайника в значительной степени были заменены синтетическими аналогами .
Традиционная медицина и исследования
[ редактировать ]Исторически сложилось так, что в традиционной медицине Европы Lobaria pulmonaria собиралась в больших количествах как «медуница» из-за ее внешнего вида, напоминающего легкие (« доктрина сигнатур », предполагающая, что травы могут лечить части тела, которые они внешне напоминают). Аналогичным образом, пельтигера leucophlebia («взъерошенная веснушчатая шкура») использовалась как предполагаемое лекарство от молочницы из-за сходства ее цефалодий с внешним видом заболевания. [34]
Лишайники производят метаболиты, которые исследуются на предмет их потенциальной терапевтической или диагностической ценности. [155] Некоторые метаболиты, продуцируемые лишайниками, структурно и функционально подобны антибиотикам широкого спектра действия, тогда как лишь немногие из них имеют антисептическое сходство. [156] Усниновая кислота является наиболее часто изучаемым метаболитом, вырабатываемым лишайниками. [156] Он также исследуется как бактерицидное средство против Escherichia coli и Staphylococcus aureus . [157]
Эстетическая привлекательность
[ редактировать ]
Колонии лишайников могут иметь впечатляющий внешний вид, доминируя над поверхностью визуального ландшафта, что является частью эстетической привлекательности для посетителей национального парка Йосемити , национального парка Секвойя и залива Огней . [38] : 2 Оранжевые и желтые лишайники создают атмосферу пустынных деревьев, тундры и скалистых берегов моря. Замысловатая паутина лишайников, свисающая с ветвей деревьев, придает лесу таинственный вид. Кустарниковые лишайники используются в моделях железных дорог. [158] и другие хобби по лепке как материал для изготовления миниатюрных деревьев и кустарников.
В литературе
[ редактировать ]В ранней мидрашской литературе еврейское слово « вайлафет » в Руфь 3:8 объясняется как относящееся к Руфи, обвивающей себя вокруг Вооза, как лишайник. [159] Арабский врач X века Аль-Тамими упоминает, что лишайники, растворенные в уксусе и розовой воде, использовались в его время для лечения кожных заболеваний и сыпи. [160]
Сюжет Джона Уиндема научно-фантастического романа «Проблема с лишайником» вращается вокруг химического вещества, замедляющего старение, полученного из лишайника.
История
[ редактировать ]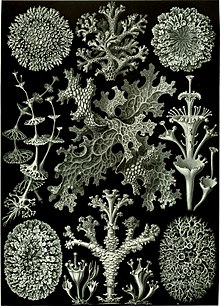
Хотя лишайники уже довольно давно были признаны организмами, только в 1867 году, когда швейцарский ботаник Саймон Швенденер предложил свою двойную теорию лишайников, согласно которой лишайники представляют собой комбинацию грибов с водорослями или цианобактериями, что раскрывает истинную природу ассоциации лишайников. начал проявляться. [161] Гипотеза Швенденера, которая в то время не имела экспериментальных подтверждений, возникла в результате его обширного анализа анатомии и развития лишайников, водорослей и грибов с использованием светового микроскопа . Многие из ведущих лихенологов того времени, такие как Джеймс Кромби и Нюландер , отвергли гипотезу Швенденера, поскольку все живые организмы были автономны. [161]
Другие выдающиеся биологи, такие как Генрих Антон де Бари , Альберт Бернхард Франк , Беатрикс Поттер , Мельхиор Треуб и Герман Хеллригель , не так быстро отвергли идеи Швенденера, и концепция вскоре распространилась на другие области исследования, такие как микробиология, растения, животные. и патогены человека. [161] [162] [163] Когда наконец были выявлены сложные взаимоотношения между патогенными микроорганизмами и их хозяевами, гипотеза Швенденера начала набирать популярность. Дальнейшее экспериментальное доказательство двойственной природы лишайников было получено, когда Ойген Томас опубликовал в 1939 году свои результаты первого успешного эксперимента по повторному синтезу. [161]
В 2010-х годах была открыта новая грань партнерства грибов и водорослей. Тоби Сприбилль и его коллеги обнаружили, что многие виды лишайников, которые долгое время считались парами аскомицет -водоросли, на самом деле представляли собой трио аскомицет- базидиомицет -водоросли. Третьим симбиотическим партнером многих лишайников являются базидиомицетовые дрожжи. [1] [164]
См. также
[ редактировать ]- Лихенология
- Лишайники и круговорот азота
- Микофикобиоз – симбиоз, при котором гриб обитает в макроскопическом слоевище пресноводных и морских водорослей; технически это не лишайник, а похожее явление, в котором грибы и водоросли находятся в симбиозе
Примечания
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д Сприбилле, Тоби; Туовинен, Веера; Ресл, Филипп; Вандерпул, Дэн; Волински, Хеймо; Эм, мсье Кэтрин; Шнайдер, Кевин; Штабентайнер, Эдит; Тооме-Хеллер, Мерье (21 июля 2016 г.). «Базидиомицетовые дрожжи в коре аскомицетов макролишайников» . Наука . 353 (6298): 488–92. Бибкод : 2016Sci...353..488S . doi : 10.1126/science.aaf8287 . ISSN 0036-8075 . ПМЦ 5793994 . ПМИД 27445309 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот Лепп, Хейно (7 марта 2011 г.). «Что такое лишайник?» . Австралийский национальный ботанический сад. Архивировано из оригинала 2 июля 2014 года . Проверено 10 октября 2014 г.
- ^ « Введение в лишайники - Союз между королевствами ». Музей палеонтологии Калифорнийского университета. Архивировано 22 августа 2014 года в Wayback Machine .
- ^ Jump up to: а б с д и Бродо, Ирвин М. и Дюран Шарнофф, Сильвия (2001) Лишайники Северной Америки . ISBN 978-0300082494 .
- ^ «Дрожжи становятся скрытым третьим партнером в лишайниковом симбиозе» . ScienceDaily . 21 июля 2016 года . Проверено 31 марта 2024 г.
- ^ Кумпула, Йоуко; Лефрер, Стефани К.; Ниеминен, Маури (1 января 2004 г.). «Использование лесных лишайниковых пастбищ северными оленями зимой в условиях легкого заснежения» . Арктика . 57 (3). дои : 10.14430/arctic504 . ISSN 1923-1245 .
- ^ Фрёберг, Ларс; Баур, Анетт; Баур, Бруно (январь 1993 г.). «Дифференциальное повреждение известковых лишайников улитками у травоядных» . Лихенолог . 25 (1): 83. дои : 10.1017/s002428299300009x . ISSN 0024-2829 .
- ^ Лейнаас, Ганс Петтер; Фьельберг, Арне (июнь 1985 г.). «Структура среды обитания и стратегии жизненного цикла двух частично симпатрических и тесно связанных видов коллембол, питающихся лишайниками» . Ойкос . 44 (3): 448. Бибкод : 1985Ойкос..44..448Л . дои : 10.2307/3565786 . ISSN 0030-1299 . JSTOR 3565786 .
- ^ Герсон, У. (октябрь 1973 г.). «Лишайниково-членистоногие ассоциации» . Лихенолог . 5 (5–6): 434–443. дои : 10.1017/S0024282973000484 . ISSN 0024-2829 . S2CID 85020765 .
- ^ Jump up to: а б Галлоуэй, ди-джей (13 мая 1999 г.). «Лишайниковый словарь» . Австралийский национальный ботанический сад. Архивировано из оригинала 6 декабря 2014 года.
- ^ Маргулис, Линн ; Баррено, Ева (2003). «Глядя на лишайники» . Бионаука . 53 (8): 776. doi : 10.1641/0006-3568(2003)053[0776:LAL]2.0.CO;2 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д Шарнофф, Стивен (2014) Полевой справочник по калифорнийским лишайникам , издательство Йельского университета. ISBN 978-0-300-19500-2
- ^ Jump up to: а б с д и ж г час я дж к Спир, Брайан Р.; Бен Вагонер (май 1997 г.). «Лишайники: история жизни и экология» . Музей палеонтологии Калифорнийского университета. Архивировано из оригинала 2 мая 2015 года . Проверено 28 апреля 2015 г.
- ^ Сиссон, Лив; Вигус, Паула (2023). Грибы Аотеароа: путеводитель для любопытного собирателя . Окленд, Новая Зеландия: Penguin Books. п. 66. ИСБН 978-1-76104-787-9 . OCLC 1372569849 .
- ^ Jump up to: а б с д и «Лишайники: систематика, Палеонтологический музей Калифорнийского университета» . Архивировано из оригинала 24 февраля 2015 года . Проверено 10 октября 2014 г.
- ^ Лендемер, Дж. К. (2011). «Таксономический пересмотр североамериканских видов Lepraria sl, продуцирующих диварикатовую кислоту, с примечаниями о типовых видах рода L. incana ». Микология . 103 (6): 1216–1229. дои : 10.3852/11-032 . ПМИД 21642343 . S2CID 34346229 .
- ^ Jump up to: а б с д Казано, Л.М.; Дель Кампо, EM; Гарсиа-Брейхо, Ф.Дж.; Рейг-Арминьяна, Дж; Гасулла, Ф; Дель Хойо, А; Гера, А; Баррено, Э (2011). « Trebouxia постоянно присутствуют две водоросли В слоевищах лишайника Ramalina Farinacea с разными физиологическими характеристиками . Сосуществование или конкуренция?». Экологическая микробиология (Представлена рукопись). 13 (3): 806–818. Бибкод : 2011EnvMi..13..806C . дои : 10.1111/j.1462-2920.2010.02386.x . hdl : 10251/60269 . ПМИД 21134099 .
- ^ Jump up to: а б Онеггер Р. (1991) Эволюция грибов: симбиоз и морфогенез, Симбиоз как источник эволюционных инноваций , Маргулис Л. и Фестер Р. (ред.). Кембридж, Массачусетс, США: MIT Press, стр. 319–340.
- ^ Jump up to: а б Грубе, М; Кардинале, М; Де Кастро, СП-младший; Мюллер, Х; Берг, Г (2009). «Видообразное структурное и функциональное разнообразие бактериальных сообществ лишайниковых симбиозов» . Журнал ISME . 3 (9): 1105–1115. Бибкод : 2009ISMEJ...3.1105G . дои : 10.1038/ismej.2009.63 . ПМИД 19554038 .
- ^ Jump up to: а б Баррено Э., Эррера-Кампос М., Гарсиа-Брейхо Ф., Гасулла Ф. и Рейг-Арминьяна Дж. (2008) «Нефотосинтезирующие бактерии, связанные с корковыми структурами слоевищ Рамалины и Усни из Мексики» . Асиломар, Пасифик-Гроув, Калифорния, США: Тезисы докладов IAL 6-совместного заседания ABLS.
- ^ Jump up to: а б Моррис Дж., Первис В. (2007). Лишайники (Жизнь) . Лондон: Музей естественной истории. п. 19. ISBN 978-0-565-09153-8 .
- ^ «Лишайник — мелочи, которые имеют значение (Служба национальных парков США)» . www.nps.gov . Проверено 17 ноября 2023 г.
- ^ Харпер, Дуглас. «лишайник» . Интернет-словарь этимологии .
- ^ лишайник . Чарльтон Т. Льюис и Чарльз Шорт. Латинский словарь по проекту «Персей» .
- ^ λειχήν . Лидделл, Генри Джордж ; Скотт, Роберт ; Греко-английский лексикон в проекте «Персей» .
- ^ читайте у Лидделла и Скотта .
- ^ Бикс, Роберт С.П. (2010). «св λειχήν, λείχω». Этимологический словарь греческого языка . Серия Лейденских индоевропейских этимологических словарей. Том. 1. При содействии Люсьена ван Бека. Лейден, Бостон: Брилл. стр. 846–47. ISBN 9789004174184 .
- ^ «Лишайник» . Кембриджский словарь . Проверено 8 сентября 2022 г.
- ^ Вордсворт, Дот (17 ноября 2012 г.). «Лишайник» . spectator.co.uk. Архивировано из оригинала 23 декабря 2014 года . Проверено 8 сентября 2022 г.
- ^ Оксфордский словарь английского языка приводит только произношение «liken»: «лишайник» . Оксфордский словарь английского языка (онлайн-изд.). Издательство Оксфордского университета . Проверено 10 января 2018 г. (Требуется подписка или членство участвующей организации .)
- ^ Jump up to: а б с д и ж «Лишайники и мохообразные, Университет штата Мичиган, 25.10.99» . Архивировано из оригинала 5 октября 2011 года . Проверено 10 октября 2014 г.
- ^ Jump up to: а б с д и ж г час я дж Словарь лишайников, Информация о лишайниках Северной Америки, Сильвия и Стивен Шарнофф, [1] Архивировано 20 января 2015 г. в Wayback Machine.
- ^ Jump up to: а б с д и ж г час я дж к «Глоссарий Алана Сильверсайда по лишайникам (pz), Алан Сильверсайд» . Архивировано из оригинала 31 октября 2014 года . Проверено 10 октября 2014 г.
- ^ Jump up to: а б с д и ж г час Добсон, Ф.С. (2011). Лишайники, иллюстрированный справочник по британским и ирландским видам . Слау, Великобритания: ISBN Richmond Publishing Co. 9780855463151 .
- ^ Jump up to: а б с д и ж г «Листистые лишайники, типы слоевища лишайников, Аллан Сильверсайд» . Архивировано из оригинала 19 октября 2014 года . Проверено 10 октября 2014 г.
- ^ Jump up to: а б с д и ж г час я дж к л м Мхи, лишайники и папоротники северо-запада Северной Америки , Дейл Х. Витт, Джанет Э. Марш, Робин Б. Бови, издательство Lone Pine Publishing Company, ISBN 0-295-96666-1
- ^ Jump up to: а б с д «Лишайники, корпорация Сагуаро-Можжевельник» . Архивировано из оригинала 10 мая 2015 года . Проверено 10 октября 2014 г.
- ^ Jump up to: а б с МакКьюн, Б.; Гренон, Дж.; Мартин, Э.; Матч, Л.С.; Мартин, EP (март 2007 г.). «Лишайники в связи с проблемами управления в национальных парках Сьерра-Невады». Североамериканские грибы . 2 :1–39. doi : 10.2509/pnwf.2007.002.003 (неактивен 16 апреля 2024 г.).
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на апрель 2024 г. ( ссылка ) - ^ Jump up to: а б с д Мичиганские лишайники, Джули Джонс Медлин, издательство B. Jain Publishers, 1996, ISBN 0877370397 , 9780877370390, [2] Архивировано 24 ноября 2016 г. в Wayback Machine.
- ^ Jump up to: а б с д и ж г час я дж Лишайники: больше о морфологии, Музей палеонтологии Калифорнийского университета, [3] Архивировано 28 февраля 2015 г. в Wayback Machine.
- ^ Jump up to: а б с д и ж г час я дж к л «Лишайниковые фотобионты, Университет Небраски, Омаха» (PDF) . Архивировано из оригинала (PDF) 6 октября 2014 года.
- ^ Jump up to: а б «Глоссарий Алана Сильверсайда по лишайнику (вперед), Алан Сильверсайд» . Архивировано из оригинала 2 ноября 2014 года . Проверено 10 октября 2014 г.
- ^ Jump up to: а б с Бюдель, Б.; Шайдеггер, К. (1996). «Морфология и анатомия слоевища». Биология лишайников . стр. 37–64. дои : 10.1017/CBO9780511790478.005 . ISBN 9780511790478 .
- ^ Jump up to: а б Хейдмарссон, Старри; Хейдмарссон, Старри (1996). «Пруина как таксономический признак лишайника рода Dermatocarpon ». Бриолог . 99 (3): 315–320. дои : 10.2307/3244302 . JSTOR 3244302 .
- ^ Шарнофф, Сильвия и Шарнофф, Стивен. «Биология лишайников и окружающая среда». Архивировано 17 октября 2015 года в Wayback Machine . sharnoffphotos.com
- ^ Погода, CS; Хранители, КГ; Лендемер, Дж.К.; Кейн, Северная Каролина; Трипп, Э.А. (2018). «Снижение сложности митохондриальных геномов лишайниковообразующих грибов проливает свет на архитектуру генома облигатных симбиозов - Интернет-библиотека Wiley». Молекулярная экология . 27 (5): 1155–1169. дои : 10.1111/mec.14519 . ПМИД 29417658 . S2CID 4238109 .
- ^ Сприбилле, Тоби; Туовинен, Веера; Ресл, Филипп; Вандерпул, Дэн; Волински, Хеймо; Эм, мсье Кэтрин; Шнайдер, Кевин; Штабентайнер, Эдит; Туме-Хеллер, Мерье; Тор, Горан; Майрхофер, Гельмут; Йоханнессон, Ханна; Маккатчеон, Джон П. (29 июля 2016 г.). «Базидиомицетовые дрожжи в коре аскомицетов макролишайников» . Наука . 353 (6298): 488–492. Бибкод : 2016Sci...353..488S . doi : 10.1126/science.aaf8287 . ПМЦ 5793994 . ПМИД 27445309 .
- ^ Скалуд, П; Пекса, О (2010). «Эволюционные выводы, основанные на последовательностях ITS рДНК и актина, показывают обширное разнообразие обыкновенной лишайниковой водоросли Asterochromis (Trebouxiophyceae, Chlorophyta)». Молекулярная филогенетика и эволюция . 54 (1): 36–46. дои : 10.1016/j.ympev.2009.09.035 . ПМИД 19853051 .
- ^ Сприбилле, Тоби; Туовинен, Веера; Ресл, Филипп; Вандерпул, Дэн; Волински, Хеймо; Эм, мсье Кэтрин; Шнайдер, Кевин; Штабентайнер, Эдит; Туме-Хеллер, Мерье; Тор, Горан; Майрхофер, Хельмут (29 июля 2016 г.). «Базидиомицетовые дрожжи в коре аскомицетов макролишайников» . Наука . 353 (6298): 488–492. Бибкод : 2016Sci...353..488S . doi : 10.1126/science.aaf8287 . ISSN 0036-8075 . ПМЦ 5793994 . ПМИД 27445309 .
- ^ Jump up to: а б с д Рамель, Гордон. «Что такое лишайник?» . Сеть земной жизни. Архивировано из оригинала 19 января 2015 года . Проверено 20 января 2015 г.
- ^ Jump up to: а б Ахмаджян В. (1993). Лишайниковый симбиоз . Нью-Йорк: Джон Уайли и сыновья. ISBN 978-0-471-57885-7 .
- ^ Jump up to: а б Онеггер, Р. (1988). «Микобионты». В Нэше III, TH (ред.). Биология лишайников . Кембридж: Издательство Кембриджского университета (опубликовано в 1996 г.). ISBN 978-0-521-45368-4 .
- ^ Ферри, Б.В., Баддели, М.С. и Хоксворт, Д.Л. (редакторы) (1973) Загрязнение воздуха и лишайники . Атлон Пресс, Лондон.
- ^ Роуз С.И., Хоксворт Д.Л. (1981). «Реколонизация лишайников в более чистом воздухе Лондона». Природа . 289 (5795): 289–292. Бибкод : 1981Natur.289..289R . дои : 10.1038/289289a0 . S2CID 4320709 .
- ^ Хоксворт, Д.Л. и Роуз, Ф. (1976) Лишайники как мониторы загрязнения . Эдвард Арнольд, Серия Института биологии, № 66. ISBN 0713125551
- ^ «Абсолютное масло дубового мха, Evernia prunastri, фиксатор парфюма» . Архивировано из оригинала 25 декабря 2014 года . Проверено 19 сентября 2014 г.
- ^ Скогланд, Терье (1984). «Организация кормовой ниши диких оленей». Экография . 7 (4): 345. Бибкод : 1984Экогр...7..345С . дои : 10.1111/j.1600-0587.1984.tb01138.x .
- ^ Лоури, Джеймс Д.; Дидерих, Пол (2003). «Лихениколезные грибы: взаимодействие, эволюция и биоразнообразие» (PDF) . Бриолог . 106 :80. doi : 10.1639/0007-2745(2003)106[0080:LFIEAB]2.0.CO;2 . S2CID 85790408 . Архивировано (PDF) из оригинала 3 января 2011 года . Проверено 2 мая 2011 г.
- ^ Хагивара К., Райт П.Р. и др. (март 2015 г.). «Сравнительный анализ антиоксидантных свойств исландских и гавайских лишайников». Экологическая микробиология . 18 (8): 2319–2325. дои : 10.1111/1462-2920.12850 . ПМИД 25808912 . S2CID 13768322 .
- ^ Одабасоглу Ф., Аслан А., Чакир А. и др. (март 2005 г.). «Антиоксидантная активность, снижение мощности и общего содержания фенолов некоторых видов лишайников». Фитотерапия . 76 (2): 216–219. дои : 10.1016/j.fitote.2004.05.012 . ПМИД 15752633 .
- ^ Jump up to: а б Хаук, Маркус; Юргенс, Саша-Рене; Лейшнер, Кристоф (2010). «Норститиновая кислота: корреляция между ее физико-химическими характеристиками и экологическими предпочтениями лишайников, продуцирующих этот депсидон». Экологическая и экспериментальная ботаника . 68 (3): 309. Бибкод : 2010EnvEB..68..309H . дои : 10.1016/j.envexpbot.2010.01.003 .
- ^ Jump up to: а б «Сеть земной жизни, рост и развитие лишайников» . Earthlife.net. Архивировано из оригинала 28 мая 2015 года . Проверено 12 октября 2014 г.
- ^ Rosenwinkel, Swenja; Korup, Oliver; Landgraf, Angela; Dzhumabaeva, Atyrgul (2015). "Limits to lichenometry" . Quaternary Science Reviews . 129 : 229–238. Bibcode : 2015QSRv..129..229R . doi : 10.1016/j.quascirev.2015.10.031 .
- ^ Миллер, Г.Х.; Эндрюс, JT (апрель 1972 г.). «Четвертичная история северного полуострова Камберленд, Восточный Баффинов остров, СЗТ, Канада. Часть VI: Предварительная кривая роста лишайников для Rhizocarpon geographicum» . Бюллетень ГСА . 83 (4): 1133–1138. doi : 10.1130/0016-7606(1972)83[1133:QHONCP]2.0.CO;2 .
- ^ Хаворт, Лия А.; Калкин, Паркер Э.; Эллис, Джеймс М. (1986). «Прямое измерение роста лишайников в хребте Центральный Брукс, Аляска, США, и его применение для лихенометрического датирования, арктических и альпийских исследований» . Арктические и альпийские исследования . 18 (3): 289–296. дои : 10.2307/1550886 . JSTOR 1550886 .
- ^ Бенедикт, Джеймс Б. (январь 2009 г.). «Обзор лихенометрического датирования и его применения в археологии» . Американская древность . 74 (1): 143–172. дои : 10.1017/S0002731600047545 . S2CID 83108496 .
- ^ Нэш III, Томас Х. (2008). "Введение". В Нэше III, TH (ред.). Биология лишайников (2-е изд.). Кембридж: Издательство Кембриджского университета. стр. 1–8. дои : 10.1017/CBO9780511790478.002 . ISBN 978-0-521-69216-8 .
- ^ Болдуин, Эмили (26 апреля 2012 г.). «Лишайник выживает в суровых условиях Марса» . Новости Скаймании. Архивировано из оригинала 28 мая 2012 года . Проверено 27 апреля 2012 г.
- ^ Шелдрейк, Мерлин (2020). Запутанная жизнь: как грибы создают наши миры, меняют наше сознание и формируют наше будущее . Бодли Хед. п. 94. ИСБН 978-1847925206 .
- ^ «ЕКА – Пилотируемые космические полеты и исследования – Лишайник выживает в космосе» . Архивировано из оригинала 26 февраля 2010 года . Проверено 16 февраля 2010 г.
- ^ Санчо, LG; Де Ла Торре, Р.; Хорнек, Г.; Аскасо, К.; Де Лос Риос, А.; Пинтадо, А.; Вержос, Дж.; Шустер, М. (2007). «Лишайники выживают в космосе: результаты эксперимента LICHENS 2005 года». Астробиология . 7 (3): 443–454. Бибкод : 2007AsBio...7..443S . дои : 10.1089/ast.2006.0046 . ПМИД 17630840 . S2CID 4121180 .
- ^ Эйхорн, Сьюзан Э., Эверт, Рэй Ф. и Рэйвен, Питер Х. (2005). Биология растений . Нью-Йорк: WH Freeman and Company. п. 1. ISBN 0716710072 .
- ^ Кук, Ребекка; Макфарланд, Кеннет (1995). Общая ботаника 111 Лабораторное пособие . Ноксвилл, Теннесси: Университет Теннесси. п. 104.
- ^ Jump up to: а б А.Н. Рай; Б. Бергман; Улла Расмуссен (31 июля 2002 г.). Цианобактерии в симбиозе . Спрингер. п. 59. ИСБН 978-1-4020-0777-4 . Архивировано из оригинала 31 декабря 2013 года . Проверено 2 июня 2013 г.
- ^ Закон, Р.; Льюис, Д.Х. (ноябрь 1983 г.). «Биотическая среда и поддержание пола — некоторые свидетельства мутуалистического симбиоза» . Биологический журнал Линнеевского общества . 20 (3): 249–276. дои : 10.1111/j.1095-8312.1983.tb01876.x . ISSN 0024-4066 .
- ^ Скалоуд, Павел; Стейнова, Яна; Редко, Тереза; Ванчурова, Люси; Пекса, Ондржей (4 мая 2015 г.). «Собираем сложную головоломку биоразнообразия водорослей: разграничение видов внутри рода Asterochromis (Trebouxiophyceae, Chlorophyta)» . Журнал психологии . 51 (3): 507–527. Бибкод : 2015JPcgy..51..507S . дои : 10.1111/jpy.12295 . ISSN 0022-3646 . ПМИД 26986666 . S2CID 25190572 .
- ^ Рамель, Гордон. «Репродуктивные структуры лишайников» . Архивировано из оригинала 28 февраля 2014 года . Проверено 22 августа 2014 г.
- ^ Jump up to: а б с Мурта Дж.Дж., Дайер П.С., Криттенден П.Д. (апрель 2000 г.). «Секс и одиночный лишайник». Природа . 404 (6778): 564. Бибкод : 2000Natur.404..564M . дои : 10.1038/35007142 . ПМИД 10766229 . S2CID 4425228 .
- ^ Jump up to: а б с Кирк П.М., Кэннон П.Ф., Минтер Д.В., Сталперс Дж.А. (2008). Словарь грибов (10-е изд.). Уоллингфорд: CABI. стр. 378–381. ISBN 978-0-85199-826-8 .
- ^ Люкинг, Роберт; Ходкинсон, Брендан П.; Ливитт, Стивен Д. (2017). «Классификация лихенизированных грибов Ascomycota и Basidiomycota 2016 года – около тысячи родов». Бриолог . 119 (4): 361–416. дои : 10.1639/0007-2745-119.4.361 . JSTOR 44250015 . S2CID 90258634 .
- ^ Люкинг, Роберт; Ходкинсон, Брендан П.; Ливитт, Стивен Д. (2017). «Исправления и дополнения к классификации лихенизированных грибов Ascomycota и Basidiomycota 2016 г.». Бриолог . 120 (1): 58–69. дои : 10.1639/0007-2745-120.1.058 . S2CID 90363578 .
- ^ Люкинг, Роберт; Ривас-Плата, Э.; Чавес, Дж.Л.; Умана, Л.; Сипман, HJM (2009). «Сколько там тропических лишайников… правда?». Лихенологическая библиотека . 100 : 399–418.
- ^ Jump up to: а б с «Форма и структура – Стикта и Дендрискокаулон » . Австралийский национальный ботанический сад. Архивировано из оригинала 28 апреля 2014 года . Проверено 18 сентября 2014 г.
- ^ Лутзони, Ф.; Кауфф, Ф.; Кокс, CJ; Маклафлин, Д.; Селио, Г.; Дентингер, Б.; Падамси, М.; Хиббетт, Д.; и др. (2004). «Сборка грибного древа жизни: прогресс, классификация и эволюция субклеточных признаков» . Американский журнал ботаники . 91 (10): 1446–1480. дои : 10.3732/ajb.91.10.1446 . ПМИД 21652303 . S2CID 9432006 .
- ^ Сандерс, Всемирный банк; Мо, РЛ; Аскасо, К. (2004). «Приливной морской лишайник, образованный пиреномицетным грибом Verrucaria tavaresiae (Ascomycotina) и бурой водорослью Petroderma maculiforme (Phaeophyceae): организация слоевища и взаимодействие симбионтов - NCBI». Американский журнал ботаники . 91 (4): 511–22. дои : 10.3732/ajb.91.4.511 . hdl : 10261/31799 . ПМИД 21653406 .
- ^ «Взаимосвязь между грибами и водорослями - Музей Нью-Брансуика» . Архивировано из оригинала 18 сентября 2018 года . Проверено 4 октября 2018 г.
- ^ Миллер, Кэти Энн; Перес-Ортега, Серхио (май 2018 г.). «Оспаривание концепции лишайника: Turgidosculum ulvae - Кембридж» . Лихенолог . 50 (3): 341–356. дои : 10.1017/S0024282918000117 . S2CID 90117544 . Архивировано из оригинала 7 октября 2018 года . Проверено 7 октября 2018 г.
- ^ Рыбалка, Н.; Вольф, М.; Андерсен, РА; Фридл, Т. (2013). «Конгруэнтность хлоропластов - Эволюционная биология BMC - BioMed Central» . Эволюционная биология BMC . 13:39 . дои : 10.1186/1471-2148-13-39 . ПМЦ 3598724 . ПМИД 23402662 .
- ^ Лутзони, Франсуа; Пейджел, Марк; Риб, Валери (21 июня 2001 г.). «Основные линии грибов произошли от симбиотических предков лишайников» . Природа . 411 (6840): 937–940. Бибкод : 2001Natur.411..937L . дои : 10.1038/35082053 . ПМИД 11418855 . S2CID 4414913 .
- ^ Хоксворт, Д.Л. (1988). «Многообразие грибно-водорослевых симбиозов, их эволюционное значение и природа лишайников». Ботанический журнал Линнеевского общества . 96 : 3–20. дои : 10.1111/j.1095-8339.1988.tb00623.x . S2CID 49717944 .
- ^ Jump up to: а б с Риккинен Дж. (1995). «Что стоит за красивыми цветами? Исследование фотобиологии лишайников». Бриобротера . 4 (3): 375–376. дои : 10.2307/3244316 . JSTOR 3244316 .
- ^ Люкинг, Роберт (25 февраля 2015 г.). «Один гриб – два лишайника» . Полевой музей естественной истории . Проверено 25 февраля 2022 г.
- ^ Фридл, Т.; Бюдель, Б. (1996). «Фотобионты». В Нэше III, TH (ред.). Биология лишайников . Кембридж: Издательство Кембриджского университета. стр. 9–26. дои : 10.1017/CBO9780511790478.003 . ISBN 978-0-521-45368-4 .
- ^ Jump up to: а б «Глоссарий Алана Сильверсайда по лишайникам (af), Алан Сильверсайд» . Архивировано из оригинала 31 октября 2014 года . Проверено 10 октября 2014 г.
- ^ Халленбек, Патрик К. (18 апреля 2017 г.). Современные проблемы фототрофных прокариот: экологические и прикладные аспекты . Спрингер. ISBN 9783319462615 . Архивировано из оригинала 4 октября 2018 года . Проверено 4 октября 2018 г.
- ^ Jump up to: а б Риккинен, Дж. (2002). «Гильдии лишайников разделяют родственные цианобактериальные симбионты». Наука . 297 (5580): 357. doi : 10.1126/science.1072961 . ПМИД 12130774 . S2CID 35731669 .
- ^ О'Брайен, Х.; Мядликовска, Ю.; Лутзони, Ф. (2005). «Оценка специализации хозяина на симбиотических цианобактериях, связанных с четырьмя близкородственными видами лишайникового гриба Peltigera » . Европейский журнал психологии . 40 (4): 363–378. Бибкод : 2005EJPhy..40..363O . дои : 10.1080/09670260500342647 . S2CID 4094256 .
- ^ Гузов-Кржеминска, Б (2006). «Гибкость фотобионтов в лишайнике Protoparmeliopsis muralis, выявленная с помощью анализа ITS рДНК». Лихенолог . 38 (5): 469–476. дои : 10.1017/s0024282906005068 . S2CID 85895322 .
- ^ Омура, Ю.; Кавачи, М.; Касаи, Ф.; Ватанабэ, М. (2006). Генетические комбинации симбионтов в вегетативно размножающемся лишайнике основе ITS рДНК» (2006 на . ) » последовательностей « Parmotrematinctorum .co; 2 .S2CID 83701566 .
- ^ Пирси-Нормор (2006). «Лишенообразующий аскомицет Evernia mesomorpha ассоциируется с множественными генотипами Trebouxia jamesii » . Новый фитолог . 169 (2): 331–344. дои : 10.1111/j.1469-8137.2005.01576.x . ПМИД 16411936 .
- ^ Лутзони, Франсуа; Пейджел, Марк; Риб, Валери (2001). «Основные линии грибов произошли от симбиотических предков лишайников» . Природа . 411 (6840): 937–940. Бибкод : 2001Natur.411..937L . дои : 10.1038/35082053 . ПМИД 11418855 . S2CID 4414913 .
- ^ Jump up to: а б «Лишайники: летопись окаменелостей». Архивировано 25 января 2010 года в Wayback Machine , Музей палеонтологии Калифорнийского университета.
- ^ Шпеер Б.Р., Ваггонер Б. «Ископаемая летопись лишайников» . Музей палеонтологии Калифорнийского университета. Архивировано из оригинала 25 января 2010 года . Проверено 16 февраля 2010 г.
- ^ Пойнар-младший, ГО. (1992). Жизнь в Янтаре . Издательство Стэнфордского университета.
- ^ Петерсон ЭБ. (2000). «Незамеченный ископаемый лишайник (Lobariaceae)». Лихенолог . 32 (3): 298–300. Бибкод : 2000ThLic..32..298P . дои : 10.1006/lich.1999.0257 . S2CID 83629248 .
- ^ Тейлор, Теннесси; Хасс, Х.; Реми, В.; Керп, Х. (1995). «Старейший ископаемый лишайник» . Природа . 378 (6554): 244. Бибкод : 1995Natur.378..244T . дои : 10.1038/378244a0 . S2CID 4353572 .
- ^ Тейлор В.А., Фри CB, Хельгемо Р., Очоада Дж. (2004). «СЭМ-анализ спонгиофитона, интерпретируемого как ископаемый лишайник». Международный журнал наук о растениях . 165 (5): 875–881. дои : 10.1086/422129 . S2CID 85382155 .
- ^ Джарен, А.Х.; Портер, С.; Куглич, Джей Джей (2003). «Метаболизм лишайников выявлен у наземных организмов раннего девона». Геология . 31 (2): 99–102. Бибкод : 2003Geo....31...99J . doi : 10.1130/0091-7613(2003)031<0099:LMIIED>2.0.CO;2 . ISSN 0091-7613 . S2CID 49241170 .
- ^ Флетчер, Би Джей; Берлинг, диджей ; Чалонер, WG (2004). «Стабильные изотопы углерода и метаболизм наземного девонского организма Spongiophyton ». Геобиология . 2 (2): 107–119. Бибкод : 2004Gbio....2..107F . дои : 10.1111/j.1472-4677.2004.00026.x . S2CID 85079041 .
- ^ Эдвардс Д; Топор Л (2012). «Доказательства родства грибов с Nematasketum, близким союзником прототакситов» . Ботанический журнал Линнеевского общества . 168 : 1–18. дои : 10.1111/j.1095-8339.2011.01195.x .
- ^ Реталлак Г.Дж.; Лендинг, Э. (2014). «Сродство и архитектура девонских стволов Prototaxites loganii ». Микология . 106 (6): 1143–1156. дои : 10.3852/13-390 . ПМИД 24990121 . S2CID 20013625 .
- ^ Каратыгин И.В.; Снигиревская Н.С.; Викулин СВ. (2009). «Самый древний наземный лишайник Winfrenatia reticulata : новая находка и новая интерпретация». Палеонтологический журнал . 43 (1): 107–114. Бибкод : 2009PalJ...43..107K . дои : 10.1134/S0031030109010110 . S2CID 85262818 .
- ^ Каратыгин И.В., Снигиревская Н.С., Викулин С.В. (2007). «Два типа симбиоза с участием грибов раннедевонских экосистем» . XV Конгресс европейских микологов, Санкт-Петербург, Россия, 16–21 сентября 2007 г. 1 (1): 226. Архивировано из оригинала 24 апреля 2013 года . Проверено 18 февраля 2011 г.
- ^ «Лишайники намного моложе, чем думали ученые, и, вероятно, эволюционировали на миллионы лет позже растений» . 15 ноября 2019 года. Архивировано из оригинала 18 декабря 2019 года . Проверено 18 ноября 2019 г.
- ^ Хансен, Карен; Перри, Брайан А.; Дрангинис, Эндрю В.; Пфистер, Дональд Х. (май 2013 г.). «Филогения весьма разнообразного семейства чашечных грибов Pyronemataceae (Pezizomycetes, Ascomycota) проясняет взаимоотношения и эволюцию избранных черт жизненного цикла» . Молекулярная филогенетика и эволюция . 67 (2): 311–335. Бибкод : 2013MolPE..67..311H . дои : 10.1016/j.ympev.2013.01.014 . ISSN 1055-7903 . ПМИД 23403226 .
- ^ Шох КЛ; Сун Г.Х.; Лопес-Хиральдес Ф; Таунсенд Дж. П.; Мядликовска Дж; Хофстеттер В; Робберце Б; Мэтени П.Б.; и др. (2009). «Древо жизни Ascomycota: филогения всего типа проясняет происхождение и эволюцию фундаментальных репродуктивных и экологических признаков» . Сист. Биол . 58 (2): 224–239. дои : 10.1093/sysbio/syp020 . ПМИД 20525580 .
- ^ Гаргас, А; Деприст, ПТ; Грубе, М; Тейлер, А (1995). «Множественное происхождение симбиозов лишайников у грибов, предполагаемое филогенией рДНК СГУ». Наука . 268 (5216): 1492–1495. Бибкод : 1995Sci...268.1492G . дои : 10.1126/science.7770775 . ПМИД 7770775 . S2CID 563601 .
- ^ Онеггер Р. (1998). «Лишайниковый симбиоз – что в нем такого впечатляющего?» (PDF) . Лихенолог . 30 (3): 193–212. дои : 10.1017/s002428299200015x . Архивировано (PDF) из оригинала 26 апреля 2019 года . Проверено 30 января 2019 г.
- ^ Ведин М., Деринг Х., Гиленстам Г. (2004). «Сапротрофия и лихенизация как варианты одного и того же вида грибов на разных субстратах: экологическая пластичность и образ жизни грибов в комплексе Strictis - Conotrema » . Новый фитолог . 16 (3): 459–465. дои : 10.1111/j.1469-8137.2004.01198.x .
- ^ Юань Х, Сяо С, Тейлор Т.Н. (2005). «Лишайноподобный симбиоз 600 миллионов лет назад». Наука . 308 (5724): 1017–1020. Бибкод : 2005Sci...308.1017Y . дои : 10.1126/science.1111347 . ПМИД 15890881 . S2CID 27083645 .
- ^ Реталлак Г.Дж. (2015). «Свидетельства Акритарха о поздней докембрийской адаптивной радиации грибов» (PDF) . Ботаника Пасифика . 4 (2): 19–33. дои : 10.17581/bp.2015.04203 . Архивировано (PDF) из оригинала 22 декабря 2016 года . Проверено 22 декабря 2016 г.
- ^ Реталлак Г.Дж. (2007). «Рост, распад и уплотнение захоронений дикинсонии, культового эдиакарского ископаемого» . Алчеринга: Австралазийский журнал палеонтологии . 31 (3): 215–240. Бибкод : 2007Алч...31..215Р . дои : 10.1080/03115510701484705 . S2CID 17181699 .
- ^ Реталлак Г.Дж. (1994). «Были ли эдиакарские ископаемые лишайники?». Палеобиология . 20 (4): 523–544. Бибкод : 1994Pbio...20..523R . дои : 10.1017/s0094837300012975 . JSTOR 2401233 . S2CID 129180481 .
- ^ Свитек Б (2012). «Спорное утверждение указывает на то, что жизнь на суше появилась на 65 миллионов лет раньше» . Природа . дои : 10.1038/nature.2012.12017 . S2CID 130305901 . Архивировано из оригинала 1 января 2013 года . Проверено 2 января 2013 г.
- ^ Реталлак, Дж.Дж.; Данн, КЛ; Саксби, Дж. (2015). «Проблематическая мезопротерозойская окаменелость Horodyskia из национального парка Глейшер, Монтана, США». Докембрийские исследования . 226 : 125–142. Бибкод : 2013PreR..226..125R . doi : 10.1016/j.precamres.2012.12.005 .
- ^ Реталлак, Дж.Дж.; Крулл, ЕС; Текрей, Джорджия; Паркинсон, Д. (2013). «Проблемные окаменелости в форме урны из палеопротерозойской (2,2 млрд лет) палеопочвы в Южной Африке». Докембрийские исследования . 235 : 71–87. Бибкод : 2013PreR..235...71R . doi : 10.1016/j.precamres.2013.05.015 .
- ^ Люкинг, Роберт; Нельсен, Мэтью П. (2018), «Эдиакарцы, протолишайники и пенициллы, полученные из лишайников» , Трансформационная палеоботаника , Elsevier, стр. 551–590, doi : 10.1016/b978-0-12-813012-4.00023-1 , ISBN 978-0-12-813012-4 , получено 14 ноября 2020 г.
- ^ Нельсен, Мэтью П.; Люкинг, Роберт; Бойс, К. Кевин; Лумбш, Х. Торстен; Ри, Ричард Х. (январь 2020 г.). «Нет подтверждения появлению лишайников до эволюции сосудистых растений» . Геобиология . 18 (1): 3–13. Бибкод : 2020Gbio...18....3N . дои : 10.1111/gbi.12369 . ISSN 1472-4677 . ПМИД 31729136 . S2CID 208034624 .
- ^ Сон, Хёнджон; Ким, Ки-Тэ; Пак, Сук-Янг; Ли, Гир-Вон; Чхве, Джеён; Чон, Чонбом; Чонг, Кёнче; Чой, Гобонг; Хур, Джэ-Сын; Ли, Ён Хван (24 июня 2022 г.). «Сравнительный геномный анализ лишайниковообразующих грибов открывает новое понимание образа жизни грибов» . Научные отчеты . 12 (1): 10724. Бибкод : 2022NatSR..1210724S . дои : 10.1038/s41598-022-14340-5 . ISSN 2045-2322 . ПМЦ 9232553 . ПМИД 35750715 .
- ^ Кляйн, ДжоАнна (19 ноября 2019 г.). «В гонке за выживание на суше лишайники не победили растения» . Нью-Йорк Таймс .
- ^ «Для горы Рашмор: запоздалое умывание лица» . Вашингтон Пост . Ассошиэйтед Пресс. 11 июля 2005 г. Проверено 20 августа 2022 г.
- ^ «Загрязнение, подземный мир растений» . Австралийский национальный ботанический сад. Архивировано из оригинала 17 февраля 2014 года . Проверено 10 октября 2014 г.
- ^ Ахмаджян, В. (1 марта 1995 г.). «Лишайники важнее, чем вы думаете» . Бионаука . 45 (3): 1. doi : 10.1093/bioscience/45.3.124 . Проверено 14 сентября 2021 г.
- ^ Чен, Цзе; Блюм, Ганс-Петер; Бейер, Лотар (2000). «Выветривание горных пород, вызванное колонизацией лишайников – обзор» (PDF) . КАТЕНА . 39 (2): 121. Бибкод : 2000Caten..39..121C . дои : 10.1016/S0341-8162(99)00085-5 . Архивировано (PDF) из оригинала 2 апреля 2015 г. Проверено 21 марта 2015 г.
- ^ Джонс, Клайв Г.; Шачак, Моше (1990). «Удобрение пустынной почвы камнеядными улитками». Природа . 346 (6287): 839. Бибкод : 1990Natur.346..839J . дои : 10.1038/346839a0 . S2CID 4311333 .
- ^ Уокер, Т.Р. (2007). «Лишайники бореальных лесов Лабрадора, Канада: Контрольный список». Эвансия . 24 (3): 85–90. дои : 10.1639/0747-9859-24.3.85 . S2CID 129100097 .
- ^ Оксанен, И. (2006). «Экологические и биотехнологические аспекты лишайников». Прикладная микробиология и биотехнология . 73 (4): 723–734. дои : 10.1007/s00253-006-0611-3 . ПМИД 17082931 . S2CID 12172616 .
- ^ Лоури, Джеймс Д. (1994). «Аллелопатия лишайников: обзор». В Индержите; КММ Дакшини; Фрэнк А. Эйнхеллиг (ред.). Аллелопатия. Организмы, процессы и приложения . Серия симпозиумов ACS. Том. 582. Американское химическое общество. стр. 26–38. дои : 10.1021/bk-1995-0582.ch002 . ISBN 978-0-8412-3061-3 .
- ^ Jump up to: а б с Нэш III, Томас Х. (2008). «Чувствительность лишайников к загрязнению воздуха». В Нэше III, TH (ред.). Биология лишайников (2-е изд.). Кембридж: Издательство Кембриджского университета. стр. 299–314. дои : 10.1017/CBO9780511790478.016 . ISBN 978-0-521-69216-8 .
- ^ Кнопс, JMH; Нэш, TH III; Буше, В.Л.; Шлезингер, WH (1991). «Минеральный круговорот и эпифитные лишайники: последствия на уровне экосистемы». Лихенолог . 23 (3): 309–321. дои : 10.1017/S0024282991000452 . S2CID 86133860 .
- ^ Хоган, К. Майкл (2010). «Абиотический фактор» . Энциклопедия Земли . Вашингтон, округ Колумбия: Национальный совет по науке и окружающей среде. Архивировано из оригинала 8 июня 2013 года . Проверено 27 октября 2013 г.
- ^ Белтман И.Х., де Кок Л.Дж., Койпер П.Дж.К., ван Хасселт П.Р. (1980). «Жирнокислотный состав и содержание хлорофилла в эпифитных лишайниках и возможная связь с их чувствительностью к загрязнению воздуха». Ойкос . 35 (3): 321–326. Бибкод : 1980Оикос..35..321Б . дои : 10.2307/3544647 . JSTOR 3544647 .
- ^ Эммерих Р., Гиз И., Ланге О.Л., Прокш П. (1993). «Токсичность и антифидантная активность соединений лишайника в отношении многоядных травоядных насекомых Spodopteralittoralis ». Фитохимия . 33 (6): 1389–1394. Бибкод : 1993PChem..33.1389E . дои : 10.1016/0031-9422(93)85097-Б .
- ^ Бешель Р.Э. (1950). «Флектен как возрастной ориентир для современных морен». Журнал ледниковой науки и ледниковой геологии . 1 : 152-161.
- ^ Карри, Р.Р. (1969) «Голоценовая климатическая и ледниковая история центральной Сьерра-Невады, Калифорния», стр. 1–47, Специальный доклад Геологического общества Америки, 123, С.А. Шумм и У.К. Брэдли, ред.
- ^ Соуерс, Дж. М., Ноллер, Дж. С., и Леттис, В. Р. (ред.) (1997) Датирование и землетрясения: обзор четвертичной геохронологии и ее применение к палеосейсмологии . Комиссия по ядерному регулированию США, NUREG/CR 5562.
- ^ Иннес, Дж.Л. (1985). «Лихенометрия». Успехи физической географии . 9 (2): 187. Бибкод : 1985ПрПГ....9..187И . дои : 10.1177/0309133338500900202 . S2CID 220949784 .
- ^ Каппителли, Франческа; Сорлини, Клаудия (2008). «Микроорганизмы атакуют синтетические полимеры в предметах, представляющих наше культурное наследие» . Прикладная и экологическая микробиология . 74 (3): 564–569. Бибкод : 2008ApEnM..74..564C . дои : 10.1128/АЕМ.01768-07 . ПМК 2227722 . ПМИД 18065627 .
- ^ Гэдд, Джеффри Майкл (март 2010 г.). «Металлы, минералы и микробы: геомикробиология и биоремедиация» . Микробиология . 156 (Часть 3): 609–643. дои : 10.1099/mic.0.037143-0 . ПМИД 20019082 .
- ^ Джонсон, Кристофер Дж; Беннет, Джеймс П.; Биро, Стивен М; Дуке-Веласкес, Хуан Камило; Родригес, Синтия М.; Бессен, Ричард А; Рок, Тони Э (май 2011 г.). Барц, Джейсон С. (ред.). «Деградация прионного белка, связанного с заболеванием, сериновой протеазой лишайников» . ПЛОС ОДИН . 6 (5): e19836. Бибкод : 2011PLoSO...619836J . дои : 10.1371/journal.pone.0019836 . ПМЦ 3092769 . ПМИД 21589935 .
- ^ Родригес, Синтия М; Беннет, Джеймс П.; Джонсон, Кристофер Дж (1 января 2012 г.). «Лишайники: неожиданные антиприонные агенты?» . Прион . 6 (1): 11–16. дои : 10.4161/при.6.1.17414 . ПМЦ 3338958 . ПМИД 22453171 .
- ^ Беннет, Джеймс П.; Родригес, Синтия М; Джонсон, Кристофер (июль 2012 г.). «Деградация прионного белка лишайниками рода Cladonia» . Лихенолог . 44 (4): 523–531. дои : 10.1017/S0024282912000102 . S2CID 85819801 .
- ^ «Современный травяной препарат | Лакмусовая бумажка» . botanical.com . Проверено 24 октября 2021 г.
- ^ Кассельман, Карен Ли; Дин, Дженни (1999). Дикий цвет: [полное руководство по изготовлению и использованию натуральных красителей] . Нью-Йорк: Публикации Уотсона-Гуптилла. ISBN 978-0-8230-5727-6 .
- ^ Мюллер, К. (2001). «Фармацевтически значимые метаболиты лишайников». Прикладная микробиология и биотехнология . 56 (1–2): 9–10. дои : 10.1007/s002530100684 . ПМИД 11499952 . S2CID 11958438 .
- ^ Jump up to: а б Мортон, Э.; Уинтерс Дж. и Смит Л. (2010). «Анализ антисептических и антибиотических свойств мхов и лишайников, обработанных различными способами». Архивировано 20 августа 2017 г. на Wayback Machine . Биологическая станция Мичиганского университета
- ^ Бустинца, Ф. (1952). «Антибактериальные вещества из лишайников». Экономическая ботаника . 6 (4): 402–406. дои : 10.1007/bf02984888 . S2CID 39335883 .
- ^ «Themodelrailroader.com» . Архивировано из оригинала 15 октября 2014 года . Проверено 10 октября 2014 г.
- ↑ Так объяснил раввин Енох Зундель бен Иосиф в своем комментарии Эц Йосеф («Древо Иосифа») к Сефер Мидраш Рабба , том. 2, Нью-Йорк 1987, св. Рут Рабба 6:3
- ^ Зохар Амар и Ярон Серри, Земля Израиля и Сирия в описании Аль-Тамими , Рамат-Ган, 2004 г., стр. 56, 108–109. ISBN 965-226-252-8 (иврит)
- ^ Jump up to: а б с д Онеггер Р. (2000). «Саймон Швендер (1829–1919) и двойная гипотеза лишайников». Бриолог . 103 (2): 307–313. doi : 10.1639/0007-2745(2000)103[0307:SSATDH]2.0.CO;2 . ISSN 0007-2745 . JSTOR 3244159 . S2CID 84580224 .
- ^ Треуб, Мельхиор (1873) Исследования природы лишайников. Диссертация Лейденского университета.
- ^ Уэйкфорд, Том (2002). Жизненные связи: от роголистников до гиппопотамов, как скромный микроб управлял эволюцией . Джон Уайли и сыновья, Инкорпорейтед. ISBN 978-0-470-24400-5 . OCLC 922973303 .
- ^ Йонг, Эд (21 июля 2016 г.), «Как парень из трейлерного парка Монтаны перевернул 150 лет биологии» , The Atlantic , заархивировано из оригинала 23 июля 2017 г. , получено 23 июля 2017 г.