Лишайник







Лишайник ( / l aɪ k Большинство одинаково / ly - , Великобритания также / ˈ L ɪ Tʃ ən / litch -ən ) является гибридной колонией водорослей живых или цианобактерий, симбиотически среди филаментов множества видов грибов , а также дрожжи, встроенные в кору или «кожу», в взаимных отношениях. [ 1 ] [ 2 ] [ устаревший источник ] [ 3 ] [ устаревший источник ] [ 4 ] [ устаревший источник ] [ 5 ]
Лишайники являются важными актерами в велосипеде питательных веществ и выступают в качестве производителей, которые питаются многие более высокие трофические кормушки, такие как оленя, гастроподы, нематоды, клещи и весенние хвосты. [ 6 ] [ 7 ] [ 8 ] [ 9 ] Лишайники обладают свойствами, отличными от свойств их компонентных организмов. Они бывают разных цветов, размеров и форм и иногда похожи на растения, но не являются растениями . У них могут быть крошечные, безлистные ветви ( фрутикоза ); плоские листообразные структуры ( фолиоз ); выращивать, похожий на кору, плотно прилипающий к поверхности ( субстрат ), как толстый слой краски ( кровь ); [ 10 ] иметь порошкообразную внешний вид ( проказа ); или другие формы роста . [ 11 ]
Макролихен ; -это лишайник, который либо похож на кустарник, либо листовой Все остальные лишайны называются микролихенами . [ 2 ] Здесь «макро» и «микро» не относятся не к размеру, а к форме роста. [ 2 ] Общие названия для лишайников могут содержать слово «Мосс» (например, « оленевой мох », « Исландия Мосс »), а лишайники могут поверхностно выглядеть и расти с мхами , но они не тесно связаны с мхами или каким -либо растением. [4]: 3 Lichens do not have roots that absorb water and nutrients as plants do,[12]: 2 but like plants, they produce their own nutrition by photosynthesis.[13] Когда они растут на растениях, они не живут как паразиты , а вместо этого используют поверхность растения в качестве субстрата.
Lichens occur from sea level to high alpine elevations, in many environmental conditions, and can grow on almost any surface.[13][14] They are abundant growing on bark, leaves, mosses, or other lichens[12] and hanging from branches "living on thin air" (epiphytes) in rainforests and in temperate woodland. They grow on rock, walls, gravestones, roofs, exposed soil surfaces, rubber, bones, and in the soil as part of biological soil crusts. Various lichens have adapted to survive in some of the most extreme environments on Earth: arctic tundra, hot dry deserts, rocky coasts, and toxic slag heaps. They can even live inside solid rock, growing between the grains (endolithic).
There are about 20,000 known species.[15] Some lichens have lost the ability to reproduce sexually, yet continue to speciate.[12][16] They can be seen as being relatively self-contained miniature ecosystems, where the fungi, algae, or cyanobacteria have the potential to engage with other microorganisms in a functioning system that may evolve as an even more complex composite organism.[17][18][19][20] Lichens may be long-lived, with some considered to be among the oldest living things.[4][21] They are among the first living things to grow on fresh rock exposed after an event such as a landslide. The long life-span and slow and regular growth rate of some species can be used to date events (lichenometry). Lichens are a keystone species in many ecosystems and benefit trees and birds.[22]
Etymology and pronunciation
[edit]The English word lichen derives from the Greek λειχήν leichēn ("tree moss, lichen, lichen-like eruption on skin") via Latin lichen.[23][24][25] The Greek noun, which literally means "licker", derives from the verb λείχειν leichein, "to lick".[26][27] In American English, "lichen" is pronounced the same as the verb "liken" (/ˈlaɪkən/). In British English, both this pronunciation and one rhyming with "kitchen" (/ˈlɪtʃən/) are used.[28][29][30]
Anatomy and morphology
[edit]Growth forms
[edit]Lichens grow in a wide range of shapes and forms; this external appearance is known as their morphology. The shape of a lichen is usually determined by the organization of the fungal filaments.[31] The nonreproductive tissues, or vegetative body parts, are called the thallus. Lichens are grouped by thallus type, since the thallus is usually the most visually prominent part of the lichen. Thallus growth forms typically correspond to a few basic internal structure types. Common names for lichens often come from a growth form or color that is typical of a lichen genus.
Common groupings of lichen thallus growth forms are:
- fruticose[32][33][34] – growing like a tuft or multiple-branched leafless mini-shrub, upright or hanging down, 3-dimensional branches with nearly round cross section (terete) or flattened
- foliose[32][33] – growing in 2-dimensional, flat, leaf-like lobes
- crustose[10][32][33] – crust-like, adhering tightly to a surface (substrate) like a thick coat of paint
- squamulose[34] – formed of small leaf-like scales crustose below but free at the tips
- leprose[35] – powdery
- gelatinous – jelly-like
- filamentous – stringy or like matted hair
- byssoid – wispy, like teased wool
- structureless
There are variations in growth types in a single lichen species, grey areas between the growth type descriptions, and overlapping between growth types, so some authors might describe lichens using different growth type descriptions.
When a crustose lichen gets old, the center may start to crack up like old-dried paint, old-broken asphalt paving, or like the polygonal "islands" of cracked-up mud in a dried lakebed. This is called being rimose or areolate, and the "island" pieces separated by the cracks are called areolas.[32] The areolas appear separated, but are (or were)[citation needed] connected by an underlying prothallus or hypothallus.[35] When a crustose lichen grows from a center and appears to radiate out, it is called crustose placodioid. When the edges of the areolas lift up from the substrate, it is called squamulose.[36]: 159 [34]
These growth form groups are not precisely defined. Foliose lichens may sometimes branch and appear to be fruticose. Fruticose lichens may have flattened branching parts and appear leafy. Squamulose lichens may appear where the edges lift up. Gelatinous lichens may appear leafy when dry.[36]: 159
The thallus is not always the part of the lichen that is most visually noticeable. Some lichens can grow inside solid rock between the grains (endolithic lichens), with only the sexual fruiting part visible growing outside the rock.[32] These may be dramatic in color or appearance.[32] Forms of these sexual parts are not in the above growth form categories.[32] The most visually noticeable reproductive parts are often circular, raised, plate-like or disc-like outgrowths, with crinkly edges, and are described in sections below.
Color
[edit]
Lichens come in many colors.[12]: 4 Coloration is usually determined by the photosynthetic component.[31] Special pigments, such as yellow usnic acid, give lichens a variety of colors, including reds, oranges, yellows, and browns, especially in exposed, dry habitats.[37] In the absence of special pigments, lichens are usually bright green to olive gray when wet, gray or grayish-green to brown when dry.[37] This is because moisture causes the surface skin (cortex) to become more transparent, exposing the green photobiont layer.[37] Different colored lichens covering large areas of exposed rock surfaces, or lichens covering or hanging from bark can be a spectacular display when the patches of diverse colors "come to life" or "glow" in brilliant displays following rain.
Different colored lichens may inhabit different adjacent sections of a rock face, depending on the angle of exposure to light.[37] Colonies of lichens may be spectacular in appearance, dominating much of the surface of the visual landscape in forests and natural places, such as the vertical "paint" covering the vast rock faces of Yosemite National Park.[38]
Color is used in identification.[39]: 4 The color of a lichen changes depending on whether the lichen is wet or dry.[39] Color descriptions used for identification are based on the color that shows when the lichen is dry.[39] Dry lichens with a cyanobacterium as the photosynthetic partner tend to be dark grey, brown, or black.[39]
The underside of the leaf-like lobes of foliose lichens is a different color from the top side (dorsiventral), often brown or black, sometimes white. A fruticose lichen may have flattened "branches", appearing similar to a foliose lichen, but the underside of a leaf-like structure on a fruticose lichen is the same color as the top side. The leaf-like lobes of a foliose lichen may branch, giving the appearance of a fruticose lichen, but the underside will be a different color from the top side.[35]
The sheen on some jelly-like gelatinous lichens is created by mucilaginous secretions.[31]
Internal structure
[edit]
(a) The cortex is the outer layer of tightly woven fungus filaments (hyphae)
(b) This photobiont layer has photosynthesizing green algae
(c) Loosely packed hyphae in the medulla
(d) A tightly woven lower cortex
(e) Anchoring hyphae called rhizines where the fungus attaches to the substrate
A lichen consists of a simple photosynthesizing organism, usually a green alga or cyanobacterium, surrounded by filaments of a fungus. Generally, most of a lichen's bulk is made of interwoven fungal filaments,[40] but this is reversed in filamentous and gelatinous lichens.[31] The fungus is called a mycobiont. The photosynthesizing organism is called a photobiont. Algal photobionts are called phycobionts.[41] Cyanobacteria photobionts are called cyanobionts.[41]
The part of a lichen that is not involved in reproduction, the "body" or "vegetative tissue" of a lichen, is called the thallus. The thallus form is very different from any form where the fungus or alga are growing separately. The thallus is made up of filaments of the fungus called hyphae. The filaments grow by branching then rejoining to create a mesh, which is called being "anastomosed". The mesh of fungal filaments may be dense or loose.
Generally, the fungal mesh surrounds the algal or cyanobacterial cells, often enclosing them within complex fungal tissues that are unique to lichen associations. The thallus may or may not have a protective "skin" of densely packed fungal filaments, often containing a second fungal species,[1] which is called a cortex. Fruticose lichens have one cortex layer wrapping around the "branches". Foliose lichens have an upper cortex on the top side of the "leaf", and a separate lower cortex on the bottom side. Crustose and squamulose lichens have only an upper cortex, with the "inside" of the lichen in direct contact with the surface they grow on (the substrate). Even if the edges peel up from the substrate and appear flat and leaf-like, they lack a lower cortex, unlike foliose lichens. Filamentous, byssoid, leprose,[35] gelatinous, and other lichens do not have a cortex; in other words, they are ecorticate.[42]
Fruticose, foliose, crustose, and squamulose lichens generally have up to three different types of tissue, differentiated by having different densities of fungal filaments.[40] The top layer, where the lichen contacts the environment, is called a cortex.[40] The cortex is made of densely tightly woven, packed, and glued together (agglutinated) fungal filaments.[40] The dense packing makes the cortex act like a protective "skin", keeping other organisms out, and reducing the intensity of sunlight on the layers below.[40] The cortex layer can be up to several hundred micrometers (μm) in thickness (less than a millimeter).[43] The cortex may be further topped by an epicortex of secretions, not cells, 0.6–1 μm thick in some lichens.[43] This secretion layer may or may not have pores.[43]
Below the cortex layer is a layer called the photobiontic layer or symbiont layer.[33][40] The symbiont layer has less densely packed fungal filaments, with the photosynthetic partner embedded in them.[40] The less dense packing allows air circulation during photosynthesis, similar to the anatomy of a leaf.[40] Each cell or group of cells of the photobiont is usually individually wrapped by hyphae, and in some cases penetrated by a haustorium.[31] In crustose and foliose lichens, algae in the photobiontic layer are diffuse among the fungal filaments, decreasing in gradation into the layer below. In fruticose lichens, the photobiontic layer is sharply distinct from the layer below.[31]
The layer beneath the symbiont layer is called the medulla. The medulla is less densely packed with fungal filaments than the layers above. In foliose lichens, as in Peltigera,[36]: 159 there is usually another densely packed layer of fungal filaments called the lower cortex.[35][40] Root-like fungal structures called rhizines (usually)[36]: 159 grow from the lower cortex to attach or anchor the lichen to the substrate.[2][35] Fruticose lichens have a single cortex wrapping all the way around the "stems" and "branches".[36] The medulla is the lowest layer, and may form a cottony white inner core for the branchlike thallus, or it may be hollow.[36]: 159 Crustose and squamulose lichens lack a lower cortex, and the medulla is in direct contact with the substrate that the lichen grows on.
In crustose areolate lichens, the edges of the areolas peel up from the substrate and appear leafy. In squamulose lichens the part of the lichen thallus that is not attached to the substrate may also appear leafy. But these leafy parts lack a lower cortex, which distinguishes crustose and squamulose lichens from foliose lichens.[40] Conversely, foliose lichens may appear flattened against the substrate like a crustose lichen, but most of the leaf-like lobes can be lifted up from the substrate because it is separated from it by a tightly packed lower cortex.[35]
Gelatinous,[36]: 159 byssoid, and leprose lichens lack a cortex (are ecorticate), and generally have only undifferentiated tissue, similar to only having a symbiont layer.[citation needed]
In lichens that include both green algal and cyanobacterial symbionts, the cyanobacteria may be held on the upper or lower surface in small pustules called cephalodia.
Pruinia is a whitish coating on top of an upper surface.[44] An epinecral layer is "a layer of horny dead fungal hyphae with indistinct lumina in or near the cortex above the algal layer".[44]
In August 2016, it was reported that some macrolichens have more than one species of fungus in their tissues.[1]
Physiology
[edit]Symbiotic relation
[edit]Lichens are fungi that have discovered agriculture
— Trevor Goward[45]

A lichen is a composite organism that emerges from algae or cyanobacteria living among the filaments (hyphae) of the fungi in a mutually beneficial symbiotic relationship. The fungi benefit from the carbohydrates produced by the algae or cyanobacteria via photosynthesis. The algae or cyanobacteria benefit by being protected from the environment by the filaments of the fungi, which also gather moisture and nutrients from the environment, and (usually) provide an anchor to it. Although some photosynthetic partners in a lichen can survive outside the lichen, the lichen symbiotic association extends the ecological range of both partners, whereby most descriptions of lichen associations describe them as symbiotic. Both partners gain water and mineral nutrients mainly from the atmosphere, through rain and dust. The fungal partner protects the alga by retaining water, serving as a larger capture area for mineral nutrients and, in some cases, provides minerals obtained from the substrate. If a cyanobacterium is present, as a primary partner or another symbiont in addition to a green alga as in certain tripartite lichens, they can fix atmospheric nitrogen, complementing the activities of the green alga.
In three different lineages the fungal partner has independently lost the mitochondrial gene atp9, which has key functions in mitochondrial energy production. The loss makes the fungi completely dependent on their symbionts.[46]
The algal or cyanobacterial cells are photosynthetic and, as in plants, they reduce atmospheric carbon dioxide into organic carbon sugars to feed both symbionts. Phycobionts (algae) produce sugar alcohols (ribitol, sorbitol, and erythritol), which are absorbed by the mycobiont (fungus).[41] Cyanobionts produce glucose.[41] Lichenized fungal cells can make the photobiont "leak" out the products of photosynthesis, where they can then be absorbed by the fungus.[12]: 5
It appears many, probably the majority, of lichen also live in a symbiotic relationship with an order of basidiomycete yeasts called Cyphobasidiales. The absence of this third partner could explain why growing lichen in the laboratory is difficult. The yeast cells are responsible for the formation of the characteristic cortex of the lichen thallus, and could also be important for its shape.[47]
The lichen combination of alga or cyanobacterium with a fungus has a very different form (morphology), physiology, and biochemistry than the component fungus, alga, or cyanobacterium growing by itself, naturally or in culture. The body (thallus) of most lichens is different from those of either the fungus or alga growing separately. When grown in the laboratory in the absence of its photobiont, a lichen fungus develops as a structureless, undifferentiated mass of fungal filaments (hyphae). If combined with its photobiont under appropriate conditions, its characteristic form associated with the photobiont emerges, in the process called morphogenesis.[4] In a few remarkable cases, a single lichen fungus can develop into two very different lichen forms when associating with either a green algal or a cyanobacterial symbiont. Quite naturally, these alternative forms were at first considered to be different species, until they were found growing in a conjoined manner.[citation needed]
Evidence that lichens are examples of successful symbiosis is the fact that lichens can be found in almost every habitat and geographic area on the planet.[17] Two species in two genera of green algae are found in over 35% of all lichens, but can only rarely be found living on their own outside of a lichen.[48]
In a case where one fungal partner simultaneously had two green algae partners that outperform each other in different climates, this might indicate having more than one photosynthetic partner at the same time might enable the lichen to exist in a wider range of habitats and geographic locations.[17]
At least one form of lichen, the North American beard-like lichens, are constituted of not two but three symbiotic partners: an ascomycetous fungus, a photosynthetic alga, and, unexpectedly, a basidiomycetous yeast.[49]
Phycobionts can have a net output of sugars with only water vapor.[41] The thallus must be saturated with liquid water for cyanobionts to photosynthesize.[41]
Algae produce sugars that are absorbed by the fungus by diffusion into special fungal hyphae called appressoria or haustoria in contact with the wall of the algal cells.[50] The appressoria or haustoria may produce a substance that increases permeability of the algal cell walls, and may penetrate the walls.[50] The algae may contribute up to 80% of their sugar production to the fungus.[50]
Ecology
[edit]Lichen associations may be examples of mutualism or commensalism, but the lichen relationship can be considered parasitic[51] under circumstances where the photosynthetic partner can exist in nature independently of the fungal partner, but not vice versa. Photobiont cells are routinely destroyed in the course of nutrient exchange. The association continues because reproduction of the photobiont cells matches the rate at which they are destroyed.[51] The fungus surrounds the algal cells,[13] often enclosing them within complex fungal tissues unique to lichen associations. In many species the fungus penetrates the algal cell wall,[13] forming penetration pegs (haustoria) similar to those produced by pathogenic fungi that feed on a host.[34][52] Cyanobacteria in laboratory settings can grow faster when they are alone rather than when they are part of a lichen.
Miniature ecosystem and holobiont theory
[edit]Symbiosis in lichens is so well-balanced that lichens have been considered to be relatively self-contained miniature ecosystems in and of themselves.[17][18] It is thought that lichens may be even more complex symbiotic systems that include non-photosynthetic bacterial communities performing other functions as partners in a holobiont.[19][20]
Many lichens are very sensitive to environmental disturbances and can be used to cheaply[13] assess air pollution,[53][54][55] ozone depletion, and metal contamination. Lichens have been used in making dyes, perfumes (oakmoss),[56] and in traditional medicines. A few lichen species are eaten by insects[13] or larger animals, such as reindeer.[57] Lichens are widely used as environmental indicators or bio-indicators. When air is very badly polluted with sulphur dioxide, there may be no lichens present; only some green algae can tolerate those conditions. If the air is clean, then shrubby, hairy and leafy lichens become abundant. A few lichen species can tolerate fairly high levels of pollution, and are commonly found in urban areas, on pavements, walls and tree bark. The most sensitive lichens are shrubby and leafy, while the most tolerant lichens are all crusty in appearance. Since industrialisation, many of the shrubby and leafy lichens such as Ramalina, Usnea and Lobaria species have very limited ranges, often being confined to the areas which have the cleanest air.
Lichenicolous fungi
[edit]Some fungi can only be found living on lichens as obligate parasites. These are referred to as lichenicolous fungi, and are a different species from the fungus living inside the lichen; thus they are not considered to be part of the lichen.[58]
Reaction to water
[edit]Moisture makes the cortex become more transparent.[12]: 4 This way, the algae can conduct photosynthesis when moisture is available, and is protected at other times. When the cortex is more transparent, the algae show more clearly and the lichen looks greener.
Metabolites, metabolite structures and bioactivity
[edit]Lichens can show intense antioxidant activity.[59][60] Secondary metabolites are often deposited as crystals in the apoplast.[61] Secondary metabolites are thought to play a role in preference for some substrates over others.[61]
Growth rate
[edit]Lichens often have a regular but very slow growth rate of less than a millimeter per year.
In crustose lichens, the area along the margin is where the most active growth is taking place.[36]: 159 Most crustose lichens grow only 1–2 mm in diameter per year.
Life span
[edit]Lichens may be long-lived, with some considered to be among the oldest living organisms.[4][21] Lifespan is difficult to measure because what defines the "same" individual lichen is not precise.[62] Lichens grow by vegetatively breaking off a piece, which may or may not be defined as the "same" lichen, and two lichens can merge, then becoming the "same" lichen.[62] One specimen of Rhizocarpon geographicum on East Baffin Island has an estimated age of 9500 years.[63][64] Thalli of Rhizocarpon geographicum and Rhizocarpon eupetraeoides/inarense in the central Brooks Range of northern Alaska have been given a maximum possible age of 10,000–11,500 years.[65][66]
Response to environmental stress
[edit]Unlike simple dehydration in plants and animals, lichens may experience a complete loss of body water in dry periods.[13] Lichens are capable of surviving extremely low levels of water content (poikilohydric).[67]: 5–6 They quickly absorb water when it becomes available again, becoming soft and fleshy.[13]
In tests, lichen survived and showed remarkable results on the adaptation capacity of photosynthetic activity within the simulation time of 34 days under Martian conditions in the Mars Simulation Laboratory (MSL) maintained by the German Aerospace Center (DLR).[68][69]
The European Space Agency has discovered that lichens can survive unprotected in space. In an experiment led by Leopoldo Sancho from the Complutense University of Madrid, two species of lichen—Rhizocarpon geographicum and Rusavskia elegans—were sealed in a capsule and launched on a Russian Soyuz rocket 31 May 2005. Once in orbit, the capsules were opened and the lichens were directly exposed to the vacuum of space with its widely fluctuating temperatures and cosmic radiation. After 15 days, the lichens were brought back to earth and were found to be unchanged in their ability to photosynthesize.[70][71]
Reproduction and dispersal
[edit]Vegetative reproduction
[edit]
Many lichens reproduce asexually, either by a piece breaking off and growing on its own (vegetative reproduction) or through the dispersal of diaspores containing a few algal cells surrounded by fungal cells.[2] Because of the relative lack of differentiation in the thallus, the line between diaspore formation and vegetative reproduction is often blurred. Fruticose lichens can fragment, and new lichens can grow from the fragment (vegetative reproduction). Many lichens break up into fragments when they dry, dispersing themselves by wind action, to resume growth when moisture returns.[72][73] Soredia (singular: "soredium") are small groups of algal cells surrounded by fungal filaments that form in structures called soralia, from which the soredia can be dispersed by wind.[2] Isidia (singular: "isidium") are branched, spiny, elongated, outgrowths from the thallus that break off for mechanical dispersal.[2] Lichen propagules (diaspores) typically contain cells from both partners, although the fungal components of so-called "fringe species" rely instead on algal cells dispersed by the "core species".[74]
Sexual reproduction
[edit]
Structures involved in reproduction often appear as discs, bumps, or squiggly lines on the surface of the thallus.[12]: 4 Though it has been argued that sexual reproduction in photobionts is selected against, there is strong evidence that suggests meiotic activities (sexual reproduction) in Trebouxia.[75][76] Many lichen fungi reproduce sexually like other fungi, producing spores formed by meiosis and fusion of gametes. Following dispersal, such fungal spores must meet with a compatible algal partner before a functional lichen can form.
Some lichen fungi belong to the phylum Basidiomycota (basidiolichens) and produce mushroom-like reproductive structures resembling those of their nonlichenized relatives.
Most lichen fungi belong to Ascomycetes (ascolichens). Among the ascolichens, spores are produced in spore-producing structures called ascomata.[12] The most common types of ascomata are the apothecium (plural: apothecia) and perithecium (plural: perithecia).[12]: 14 Apothecia are usually cups or plate-like discs located on the top surface of the lichen thallus. When apothecia are shaped like squiggly line segments instead of like discs, they are called lirellae.[12]: 14 Perithecia are shaped like flasks that are immersed in the lichen thallus tissue, which has a small hole for the spores to escape the flask, and appear like black dots on the lichen surface.[12]: 14
The three most common spore body types are raised discs called apothecia (singular: apothecium), bottle-like cups with a small hole at the top called perithecia (singular: perithecium), and pycnidia (singular: pycnidium), shaped like perithecia but without asci (an ascus is the structure that contains and releases the sexual spores in fungi of the Ascomycota).[77]
The apothecium has a layer of exposed spore-producing cells called asci (singular: ascus), and is usually a different color from the thallus tissue.[12]: 14 When the apothecium has an outer margin, the margin is called the exciple.[12]: 14 When the exciple has a color similar to colored thallus tissue the apothecium or lichen is called lecanorine, meaning similar to members of the genus Lecanora.[12]: 14 When the exciple is blackened like carbon it is called lecideine meaning similar to members of the genus Lecidea.[12]: 14 When the margin is pale or colorless it is called biatorine.[12]: 14

A "podetium" (plural: podetia) is a lichenized stalk-like structure of the fruiting body rising from the thallus, associated with some fungi that produce a fungal apothecium.[33] Since it is part of the reproductive tissue, podetia are not considered part of the main body (thallus), but may be visually prominent.[33] The podetium may be branched, and sometimes cup-like. They usually bear the fungal pycnidia or apothecia or both.[33] Many lichens have apothecia that are visible to the naked eye.[2]
Most lichens produce abundant sexual structures.[78] Many species appear to disperse only by sexual spores.[78] For example, the crustose lichens Graphis scripta and Ochrolechia parella produce no symbiotic vegetative propagules. Instead, the lichen-forming fungi of these species reproduce sexually by self-fertilization (i.e. they are homothallic). This breeding system may enable successful reproduction in harsh environments.[78]
Mazaedia (singular: mazaedium) are apothecia shaped like a dressmaker's pin in pin lichens, where the fruiting body is a brown or black mass of loose ascospores enclosed by a cup-shaped exciple, which sits on top of a tiny stalk.[12]: 15
Taxonomy and classification
[edit]Lichens are classified by the fungal component. Lichen species are given the same scientific name (binomial name) as the fungus species in the lichen. Lichens are being integrated into the classification schemes for fungi. The alga bears its own scientific name, which bears no relationship to that of the lichen or fungus.[79] There are about 20,000 identified lichen species,[80][81] and taxonomists have estimated that the total number of lichen species (including those yet undiscovered) might be as high as 28,000.[82] Nearly 20% of known fungal species are associated with lichens.[50]
"Lichenized fungus" may refer to the entire lichen, or to just the fungus. This may cause confusion without context. A particular fungus species may form lichens with different algae species, giving rise to what appear to be different lichen species, but which are still classified (as of 2014) as the same lichen species.[83]
Formerly, some lichen taxonomists placed lichens in their own division, the Mycophycophyta, but this practice is no longer accepted because the components belong to separate lineages. Neither the ascolichens nor the basidiolichens form monophyletic lineages in their respective fungal phyla, but they do form several major solely or primarily lichen-forming groups within each phylum.[84] Even more unusual than basidiolichens is the fungus Geosiphon pyriforme, a member of the Glomeromycota that is unique in that it encloses a cyanobacterial symbiont inside its cells. Geosiphon is not usually considered to be a lichen, and its peculiar symbiosis was not recognized for many years. The genus is more closely allied to endomycorrhizal genera. Fungi from Verrucariales also form marine lichens with the brown algae Petroderma maculiforme,[85] and have a symbiotic relationship with seaweed (such as rockweed) and Blidingia minima, where the algae are the dominant components. The fungi is thought to help the rockweeds to resist desiccation when exposed to air.[86][87] In addition, lichens can also use yellow-green algae (Heterococcus) as their symbiotic partner.[88]
Lichens independently emerged from fungi associating with algae and cyanobacteria multiple times throughout history.[89]

Fungi
[edit]The fungal component of a lichen is called the mycobiont. The mycobiont may be an Ascomycete or Basidiomycete.[15] The associated lichens are called either ascolichens or basidiolichens, respectively. Living as a symbiont in a lichen appears to be a successful way for a fungus to derive essential nutrients, since about 20% of all fungal species have acquired this mode of life.[90]
Thalli produced by a given fungal symbiont with its differing partners may be similar,[citation needed] and the secondary metabolites identical,[citation needed] indicating[citation needed] that the fungus has the dominant role in determining the morphology of the lichen. But the same mycobiont with different photobionts may also produce very different growth forms.[83] Lichens are known in which there is one fungus associated with two or even three algal species.
Although each lichen thallus generally appears homogeneous, some evidence seems to suggest that the fungal component may consist of more than one genetic individual of that species.[citation needed]
Two or more fungal species can interact to form the same lichen.[91]
The following table lists the orders and families of fungi that include lichen-forming species.
Photobionts
[edit]
The photosynthetic partner in a lichen is called a photobiont. The photobionts in lichens come from a variety of simple prokaryotic and eukaryotic organisms. In the majority of lichens the photobiont is a green alga (Chlorophyta) or a cyanobacterium. In some lichens both types are present; in such cases, the alga is typically the primary partner, with the cyanobacteria being located in cryptic pockets.[92] Algal photobionts are called phycobionts, while cyanobacterial photobionts are called cyanobionts.[41] About 90% of all known lichens have phycobionts, and about 10% have cyanobionts.[41] Approximately 100 species of photosynthetic partners from 40[41] genera and five distinct classes (prokaryotic: Cyanophyceae; eukaryotic: Trebouxiophyceae, Phaeophyceae, Chlorophyceae) have been found to associate with the lichen-forming fungi.[93]
Common algal photobionts are from the genera Trebouxia, Trentepohlia, Pseudotrebouxia, or Myrmecia. Trebouxia is the most common genus of green algae in lichens, occurring in about 40% of all lichens. "Trebouxioid" means either a photobiont that is in the genus Trebouxia, or resembles a member of that genus, and is therefore presumably a member of the class Trebouxiophyceae.[33] The second most commonly represented green alga genus is Trentepohlia.[34] Overall, about 100 species of eukaryotes are known to occur as photobionts in lichens. All the algae are probably able to exist independently in nature as well as in the lichen.[91]
A "cyanolichen" is a lichen with a cyanobacterium as its main photosynthetic component (photobiont).[94] Most cyanolichen are also ascolichens, but a few basidiolichen like Dictyonema and Acantholichen have cyanobacteria as their partner.[95]
The most commonly occurring cyanobacterium genus is Nostoc.[91] Other[34] common cyanobacterium photobionts are from Scytonema.[15] Many cyanolichens are small and black, and have limestone as the substrate.[citation needed] Another cyanolichen group, the jelly lichens of the genera Collema or Leptogium are gelatinous and live on moist soils. Another group of large and foliose species including Peltigera, Lobaria, and Degelia are grey-blue, especially when dampened or wet. Many of these characterize the Lobarion communities of higher rainfall areas in western Britain, e.g., in the Celtic rain forest. Strains of cyanobacteria found in various cyanolichens are often closely related to one another.[96] They differ from the most closely related free-living strains.[96]
The lichen association is a close symbiosis. It extends the ecological range of both partners but is not always obligatory for their growth and reproduction in natural environments, since many of the algal symbionts can live independently. A prominent example is the alga Trentepohlia, which forms orange-coloured populations on tree trunks and suitable rock faces. Lichen propagules (diaspores) typically contain cells from both partners, although the fungal components of so-called "fringe species" rely instead on algal cells dispersed by the "core species".[74]
The same cyanobiont species can occur in association with different fungal species as lichen partners.[97] The same phycobiont species can occur in association with different fungal species as lichen partners.[41] More than one phycobiont may be present in a single thallus.[41]
A single lichen may contain several algal genotypes.[98][99] These multiple genotypes may better enable response to adaptation to environmental changes, and enable the lichen to inhabit a wider range of environments.[100]
Controversy over classification method and species names
[edit]There are about 20,000 known lichen species.[15] But what is meant by "species" is different from what is meant by biological species in plants, animals, or fungi, where being the same species implies that there is a common ancestral lineage.[15] Because lichens are combinations of members of two or even three different biological kingdoms, these components must have a different ancestral lineage from each other. By convention, lichens are still called "species" anyway, and are classified according to the species of their fungus, not the species of the algae or cyanobacteria. Lichens are given the same scientific name (binomial name) as the fungus in them, which may cause some confusion. The alga bears its own scientific name, which has no relationship to the name of the lichen or fungus.[79]
Depending on context, "lichenized fungus" may refer to the entire lichen, or to the fungus when it is in the lichen, which can be grown in culture in isolation from the algae or cyanobacteria. Some algae and cyanobacteria are found naturally living outside of the lichen. The fungal, algal, or cyanobacterial component of a lichen can be grown by itself in culture. When growing by themselves, the fungus, algae, or cyanobacteria have very different properties than those of the lichen. Lichen properties such as growth form, physiology, and biochemistry, are very different from the combination of the properties of the fungus and the algae or cyanobacteria.
The same fungus growing in combination with different algae or cyanobacteria, can produce lichens that are very different in most properties, meeting non-DNA criteria for being different "species". Historically, these different combinations were classified as different species. When the fungus is identified as being the same using modern DNA methods, these apparently different species get reclassified as the same species under the current (2014) convention for classification by fungal component. This has led to debate about this classification convention. These apparently different "species" have their own independent evolutionary history.[2][83]
There is also debate as to the appropriateness of giving the same binomial name to the fungus, and to the lichen that combines that fungus with an alga or cyanobacterium (synecdoche). This is especially the case when combining the same fungus with different algae or cyanobacteria produces dramatically different lichen organisms, which would be considered different species by any measure other than the DNA of the fungal component. If the whole lichen produced by the same fungus growing in association with different algae or cyanobacteria, were to be classified as different "species", the number of "lichen species" would be greater.
Diversity
[edit]The largest number of lichenized fungi occur in the Ascomycota, with about 40% of species forming such an association.[79] Some of these lichenized fungi occur in orders with nonlichenized fungi that live as saprotrophs or plant parasites (for example, the Leotiales, Dothideales, and Pezizales). Other lichen fungi occur in only five orders in which all members are engaged in this habit (Orders Graphidales, Gyalectales, Peltigerales, Pertusariales, and Teloschistales). Overall, about 98% of lichens have an ascomycetous mycobiont.[101] Next to the Ascomycota, the largest number of lichenized fungi occur in the unassigned fungi imperfecti, a catch-all category for fungi whose sexual form of reproduction has never been observed.[citation needed] Comparatively few basidiomycetes are lichenized, but these include agarics, such as species of Lichenomphalia, clavarioid fungi, such as species of Multiclavula, and corticioid fungi, such as species of Dictyonema.
Identification methods
[edit]Lichen identification uses growth form, microscopy and reactions to chemical tests.
The outcome of the "Pd test" is called "Pd", which is also used as an abbreviation for the chemical used in the test, para-phenylenediamine.[33] If putting a drop on a lichen turns an area bright yellow to orange, this helps identify it as belonging to either the genus Cladonia or Lecanora.[33]
Evolution and paleontology
[edit]The fossil record for lichens is poor.[102] The extreme habitats that lichens dominate, such as tundra, mountains, and deserts, are not ordinarily conducive to producing fossils.[102][103] There are fossilized lichens embedded in amber. The fossilized Anzia is found in pieces of amber in northern Europe and dates back approximately 40 million years.[104] Lichen fragments are also found in fossil leaf beds, such as Lobaria from Trinity County in northern California, US, dating back to the early to middle Miocene.[105]
The oldest fossil lichen in which both symbiotic partners have been recovered is Winfrenatia, an early zygomycetous (Glomeromycotan) lichen symbiosis that may have involved controlled parasitism,[citation needed] is permineralized in the Rhynie Chert of Scotland, dating from early Early Devonian, about 400 million years ago.[106] The slightly older fossil Spongiophyton has also been interpreted as a lichen on morphological[107] and isotopic[108] grounds, although the isotopic basis is decidedly shaky.[109] It has been demonstrated that Silurian-Devonian fossils Nematothallus[110] and Prototaxites[111] were lichenized. Thus lichenized Ascomycota and Basidiomycota were a component of Early Silurian-Devonian terrestrial ecosystems.[112][113] Newer research suggests that lichen evolved after the evolution of land plants.[114]
Передцом экологического состояния как Ascomycota, так и базидиомикоты, вероятно, было сапробизм , а независимые события лишайнизации могли происходить несколько раз. [ 115 ] [ 116 ] В 1995 году Гаргас и его коллеги предположили, что было по меньшей мере пять независимых источников лихенизации; Три в базидиомицетах и по крайней мере два в Аскомицетах. [ 117 ] Lutzoni et al. (2001) предполагают, что лишайна, вероятно, развивалась ранее, и последовала множество независимых потерь. Некоторые нелихнообразующие грибы, возможно, вторично потеряли способность формировать ассоциацию лишайников. В результате лишайная лишайна рассматривалась как очень успешная стратегия питания. [ 118 ] [ 119 ]
Лишайнизированный Glomeromycota может хорошо простираться обратно в докембрий. Лишайноподобные окаменелости, состоящие из коккоидных клеток ( цианобактерии ?) И тонких филаментов (мукоромикотин- гломеромикота ?), Являются перминерализованными в морском фосфорите формирования Душнтуо в южном Китае. Считается, что эти окаменелости составляют от 551 до 635 миллионов лет или эдиакаран . [ 120 ] Эдиакаранские акритархи также имеют много сходства с гломеромикотана . везикулами и спорами [ 121 ] Также утверждалось, что окаменелости эдиакарана, включая Дикинсонию , [ 122 ] были лишайники, [ 123 ] Хотя это утверждение противоречиво. [ 124 ] Эндосимбиотическая гломеромикота, сравнимая с живым геозифоном, может распространяться на протерозой в форме 1500 миллионов лет Городискии [ 125 ] и 2200 миллионов лет дискагма . [ 126 ] Открытие этих окаменелостей предполагает, что грибы развивали симбиотические партнерские отношения с фотоавтотрофами задолго до эволюции сосудистых растений, хотя гипотеза лихарана в основном отвергается из -за неуместного определения лишайников на основе тафономии и субстратной экологии. [ 127 ] Тем не менее, исследование того же ученого в 2019 году, которое отвергла гипотезу ликарана, Нельсен, использовал новые калиброванные по времени филогения, чтобы сделать вывод, что нет никаких признаков лишайника до существования сосудистых растений. [ 128 ]
Lecanoromycetes, один из наиболее распространенных классов грибов, образующих лишайники, отличался от своего предка, который также мог образовывать лишайники, около 258 миллионов лет назад, в течение позднего палеозойского периода. Тем не менее, тесно связанные клад Euritiomycetes, по-видимому, стали лишайными, образующими только 52 миллиона лет назад, в течение раннего кайнозойского периода. [ 129 ]
Экология и взаимодействие с окружающей средой
[ редактировать ]Субстраты и среда обитания
[ редактировать ]
Лишайники растут на широком диапазоне субстратов и среды обитания, в том числе некоторые из самых экстремальных условий на земле. [ 130 ] Они обильно растут на коре, листьях и висят в эпифита ветвях в тропических лесах и в умеренных лесах . Они растут на голой камне, стенах, надгробиях, крышах и открытых поверхностях почвы. Они могут выжить в некоторых из самых экстремальных сред на Земле: арктической тундры , горячие сухие пустыни , скалистые побережья и токсичные кучи шлака . Они могут жить внутри твердой скалы, растущих между зернами и в почве как часть биологической почвенной коры в засушливых местах обитания, таких как пустыни. Некоторые лишайники не растут ни на чем, живя свою жизнь, дуя вокруг окружающей среды. [ 2 ]
При росте на минеральных поверхностях некоторые лишайники медленно разлагают свой субстрат, химически разлагая и физически нарушая минералы, способствуя процессу выветривания , с помощью которого пород постепенно превращаются в почву. Хотя этот вклад в выветривание обычно является доброкачественным, он может вызвать проблемы для искусственных каменных конструкций. Например, существует постоянная проблема роста лишайников на Национальном мемориале Маунт-Рашмор , которая требует применения консерваторов, занимающихся горами для очистки памятника. [ 131 ]
Лишайники - это не паразиты на растениях, на которых они растут, а используют их только в качестве субстрата. Грибы некоторых видов лишайников могут «захватить» водоросли других видов лишайников. [ 13 ] [ 132 ] Лишайники делают свою собственную пищу из их фотосинтетических частей и поглощая минералы из окружающей среды. [ 13 ] Лишайники, растущие на листьях, могут иметь вид паразитов на листьях, но это не так. Некоторые лишайники в Diploschistes паразитируют другие лишайники. Diploschistes Muscorum начинает развитие в тканях вида кладонии хозяина . [ 52 ] : 30 [ 34 ] : 171

В арктической тундре лишайники вместе с мхами и печеночными веществами составляют большую часть наземного покрова , что помогает изолировать землю и может обеспечить корм для пастбищных животных. Примером является « оленевой мох », который является лишайником, а не мхом. [ 13 ]
Есть только два вида известных постоянно погруженных лишайников; Веноза гидротирия встречается в пресной воде, а Verrucaria Serpuloides обнаруживается в морской среде. [ 133 ]
Лишайник, который растет на скале, называется саксиколовым лишайником . [ 33 ] [ 36 ] : 159 Лишайники, которые растут на скале, являются эпилитными , и те, которые погружены в пород, растущие между кристаллами, только их плодовые тела, подверженные воздействию воздуха, называются эндолитическими лишайками . [ 32 ] [ 36 ] : 159 [ 94 ] Лишайник, который растет на коре, называется кортиколевым лишайником . [ 36 ] : 159 Лишайник, который растет на древесине, из которого была раздетала кора, называется лигниколовым лишайником . [ 42 ] Лишайники, которые погружены в ткани растений, называются эндофлоидными лишайниками или эндофлоидальными лишайниками . [ 32 ] [ 36 ] : 159 Лишайники, которые используют листья в качестве субстратов, независимо от того, есть ли лист на дереве или на земле, называются эпифиллыми или фоликологическими . [ 41 ] Террикологический лишайник растет на почве в качестве субстрата. Многие сквамулозные лишайники террикологичны. [ 36 ] : 159 Лишайки для пупок - это лишайки фолиоза, которые прикреплены к субстрату только в одной точке. [ 32 ] Бродячий лишайник вообще не прикреплен к субстрату и живет своей жизнью, взорванной ветром.
Лишайники и почвы
[ редактировать ]В дополнение к отдельным физическим механизмам, с помощью которых лишайники разбивают сырой камень, исследования показывают, что лишайники химически атакуют камень, вводя недавно хелатированные минералы в экологию. Вещества, источенные лишайниками, известными своей сильной способностью связывать и секвестр металлов, наряду с общим образованием новых минералов, особенно осалатов металлов и признаков субстратов, которые они изменяют, все подчеркивают важные роли лиша химическое выветривание . [ 134 ] Со временем это занятие создает новую плодородную почву с камня.
Лишайники могут быть важны в соревнованиях азота в почвы в некоторых пустынях, когда их съедят, вместе с их подложкой по горным камням, которые затем исправляются, вставляя азот в почвы. [ 135 ] Лишайники помогают связывать и стабилизировать песок почвы в дюнах. [ 2 ] В пустынях и полузасушливых районах лишайники являются частью обширных живых биологических коров почвы , необходимых для поддержания структуры почвы. [ 2 ]
Экологические взаимодействия
[ редактировать ]Лишайники являются пионерскими видами , среди первых живых существ, которые растут на голой скале или районах, обнаруженных в результате стихийного бедствия. [ 2 ] Лишайникам, возможно, придется конкурировать с растениями за доступ к солнечному свету, но из -за их небольшого размера и медленного роста они процветают в местах, где более высокие растения испытывают трудности с ростом. Лишайники часто являются первыми, кто поселился в местах, в которых отсутствует почва, составляя единственную растительность в некоторых экстремальных средах, таких как те, которые встречаются на высоких горных высотах и в высоких широтах. [ 136 ] Некоторые выживают в жестких условиях пустыни, а другие на замороженной почве арктических областей. [ 137 ]
Основным экофизиологическим преимуществом лишайников является то, что они являются пукилогидрическими ( poikilo - переменная, гидрическая - относящаяся к воде), что означает, что, хотя они мало контролируют статус своей гидратации, они могут переносить нерегулярные и длительные периоды тяжелой десаккации . Как и некоторые мхи , печеночные запасы , папоротники и несколько растений воскресения , после высыхания лишайники попадают в метаболическую суспензию или стазис (известный как криптобиоз ), в которых клетки симбионтов лишайников дегидратируются до такой степени, что останавливает наибольшую биохимическую активность. В этом криптобиотическом состоянии лишайники могут пережить более широкие крайности температуры, радиации и засухи в суровых условиях, в которых они часто обитают.

Лишайники не имеют корней, и не нужно использовать непрерывные резервуары воды, как большинство более высоких растений, поэтому они могут расти в местах невозможно для большинства растений, таких как голая порода, стерильная почва или песок, и различные искусственные сооружения, такие как стены, крыши и памятники. Многие лишайники также растут как эпифиты ( Epi - на поверхности, Phyte - Plant) на растениях, особенно на стволах и ветвях деревьев. При выращивании на растениях лишайники не являются паразитами ; Они не потребляют какую -либо часть растения и не отравляют его. Лишайники продуцируют аллелопатические химические вещества, которые ингибируют рост мхов. Некоторые земляные лишайники, такие как члены подрода кладина (лишайки оленей), продуцируют аллелопатические химические вещества, которые выщелачивают в почву и ингибируют прорастание семян, ели и других растений. [ 138 ] Стабильность (то есть долговечность) их субстрата является основным фактором среды обитания лишайников. Большинство лишайников растут на стабильных поверхностях скал или коре старых деревьев, но многие другие растут на почве и песке. В этих последних случаях лишайники часто являются важной частью стабилизации почвы; Действительно, в некоторых пустынных экосистемах сосудистые (высшие) семена растений не могут быть установлены, кроме как в местах, где коры лишайников стабилизируют песок и помогают сохранить воду.
Лишайники могут быть съедены некоторыми животными, такими как оленя , живущие в арктических регионах. Личинки . ряда видов Lepidoptera питаются исключительно на лишайниках К ним относятся обыкновенный лаков и мраморная красота . У них очень мало белка и с высоким содержанием углеводов, что делает их непригодными для некоторых животных. Северная летающая белка использует его для гнездования, еды и зимней воды.
Влияние загрязнения воздуха
[ редактировать ]
Если лишайники всегда подвергаются воздействию загрязнителей воздуха, без каких -либо лиственных частей, они не могут избежать накопления загрязняющих веществ. Также отсутствуют устья и кутикулу , лишайники могут поглощать аэрозоли и газы на всю поверхность таллоуса, с которой они могут легко диффундировать на слой фотобионта. [ 139 ] Поскольку лишайники не обладают корнями, их основным источником большинства элементов является воздух, и, следовательно, элементные уровни в лишайниках часто отражают накопленный состав окружающего воздуха. Процессы, с помощью которых происходит атмосферное осаждение, включают туман и росу , газообразное поглощение и сухое осаждение. [ 140 ] Следовательно, экологические исследования с лишайниками подчеркивают их осуществимость как эффективные биомониторы качества атмосферы. [ 139 ]
Не все лишайники одинаково чувствительны к загрязняющим веществам воздуха , поэтому разные виды лишайников демонстрируют различные уровни чувствительности к конкретным атмосферным загрязнителям. [ 141 ] Чувствительность лишайника к загрязнению воздуха напрямую связана с энергетическими потребностями микобионта, так что чем сильнее зависимость микобионта от фотобионта, тем чувствительным является лишайник от загрязнения воздуха. [ 142 ] После воздействия загрязнения воздуха фотобионт может использовать метаболическую энергию для восстановления своих клеточных структур, которые в противном случае использовались бы для поддержания ее фотосинтетической активности, что оставляет меньше метаболической энергии, доступной для микобионта. Изменение баланса между фотобионтом и микобионтом может привести к разрушению симбиотической ассоциации. Следовательно, снижение лишайника может быть вызвано не только накоплением токсичных веществ, но и из -за измененных питательных принадлежностей, которые предпочитают один симбионт над другим. [ 139 ]
Это взаимодействие между лишайниками и загрязнением воздуха использовалось в качестве средства мониторинга качества воздуха с 1859 года, причем более систематические методы, разработанные Уильямом Ниландером в 1866 году. [ 2 ]
Использование человека
[ редактировать ]
Еда
[ редактировать ]Лишайники едят множество различных культур по всему миру. Хотя некоторые лишайники едят только во времена голода , другие являются основной пищей или даже деликатесом . При употреблении лишайников часто встречаются два препятствия: полисахариды лишайника , как правило, не усваиваемы для людей, а лишайники обычно содержат легкие токсичные вторичные соединения , которые следует удалить перед едой. Очень немногие лишайники являются ядовитыми, но те, которые с высоким содержанием вульпиновой кислоты или USNIC, токсичны. [ 143 ] Большинство ядовитых лишайников желтые. [ Цитация необходима ]
В прошлом Исландия Мосс ( Cetraria Islandica ) была важным источником еды для людей в Северной Европе и была приготовлена в виде хлеба, каши, пудинга, супа или салата. Bryoria Fremontii (Edible Horsehair Lichen) была важной едой в некоторых частях Северной Америки, где обычно он был пробиты . Северные народы в Северной Америке и Сибири традиционно съедают частично перевариваемых лишайников оленей ( Cladina spp.) После того, как они удаляют его из рубца Карибу или оленя, которые были убиты. Rock Tripe ( Umbilicaria spp. И Lasalia Spp.) - это лишайник, который часто использовался в качестве аварийной еды в Северной Америке, а один вид, Esculanta usbilicaria ( iwatake на японском Полем
Лишайнометрия
[ редактировать ]

Лихенометрия - это метод, используемый для определения возраста обнаженных горных поверхностей в зависимости от размера лишайника Талли. Введен Бесчел в 1950 -х годах, [ 144 ] Техника нашла много приложений. Он используется в археологии , палеонтологии и геоморфологии . Он использует предполагаемую регулярную, но медленную скорость роста лишайников, чтобы определить возраст открытой породы . [ 38 ] : 9 [ 145 ] Измерение диаметра (или другого размера измерения) самого большого лишайника вида на поверхности породы указывает на продолжительность времени, так как поверхность породы была впервые обнажена. Лишайник может быть сохранен на старых скальных лицах для [ Цитация необходима ] 10000 лет, обеспечивая максимальную возрастную ограничение метода, хотя он наиболее точен (в пределах 10% ошибки) при применении к поверхностям, которые были выставлены менее 1000 лет. [ 146 ] Лихенометрия особенно полезна для поверхностей датировки менее 500 лет, так как в течение этого периода методы радиоуглеродного датирования менее точны. [ 147 ] Лишайники, наиболее часто используемые для лихенометрии, являются линии рода -ризокарпона (например, виды Rhizocarpon Geographicum , Map Lichen) и Xanthoria .
Биодеградация
[ редактировать ]Было показано, что лишайники разлагают полиэфирные смолы , как видно в археологических участках в римском городе Баело Клаудия в Испании. [ 148 ] Лишайники могут накапливать несколько загрязнителей окружающей среды, таких как свинец, медь и радионуклиды . [ 149 ] Некоторые виды лишайника, такие как Parmelia Sulcata (называемые забитыми щитом лишайника, среди других названий) и лобария Pulmonaria (Lung Lichen), и многие из Cladonia родов , как было показано, продуцируют сериновые протеазы, способные деградации патогенных форм Прион белок (PRP), который может быть полезен при лечении загрязненных окружающих резервуаров. [ 150 ] [ 151 ] [ 152 ]
Красители
[ редактировать ]Многие лишайники производят вторичные соединения, в том числе пигменты , которые уменьшают вредное количество солнечного света и мощные токсины, которые сдерживают травоядные животные или убивают бактерии. Эти соединения очень полезны для идентификации лишайников и имеют экономическое значение, как красители, такие как кудбир или примитивные антибиотики .
Индикатор pH (который может указывать на кислые или основные вещества), называемый лакмусом, представляет собой краситель, извлеченный из Roccella Tinctoria лишайника («Сорняки Дайера») [ 153 ] кипящим. Это дает свое название известному лакмусовому тесту .
Традиционные красители шотландского нагорья для Харриса Твида [ 2 ] и другие традиционные ткани были изготовлены из лишайников, в том числе апельсиновая ксантория Parietina («Общий апельсиновый лишай») и серую насыщенную Пармелия Saxatilis, обыкновенные на скалах и известный как «Crowtle».
Есть сообщения, датируемые почти 2000 лет лишайника, используемые для изготовления фиолетовых и красных красителей. [ 154 ] Большое историческое и коммерческое значение - это лишайники, принадлежащие к семейству Roccellaceae , обычно называемым Orchella Weed или Orchil. Орцеин и другие красители лишайника в основном были заменены синтетическими версиями .
Традиционная медицина и исследования
[ редактировать ]Исторически, в традиционной медицине Европы лобария пульмонария была собрана в больших количествах как «легкоеворт» из-за его легкого, похожая на легкие (« о подписях », предполагая, что травы могут лечить части тела, которые они физически похожи). Учение Leucophlebia («рюшинный веснушливый шкура») использовался в качестве предполагаемого лекарства от молочника , из -за сходства его цефалодий с появлением заболевания. [ 34 ]
Лишайники продуцируют метаболиты , исследуемые для их потенциальной терапевтической или диагностической ценности. [ 155 ] Некоторые метаболиты, продуцируемые лишайниками, структурно и функционально сходны с антибиотиками широкого спектра, в то время как немногие связаны в соответствии с антисептическими сходствами. [ 156 ] Усновая кислота является наиболее часто изучаемым метаболитом, продуцируемым лишайниками. [ 156 ] Он также находится под исследованием в качестве бактерицидного агента против Escherichia coli и Staphylococcus aureus . [ 157 ]
Эстетическая привлекательность
[ редактировать ]
Колонии лишайников могут быть впечатляющими по внешнему виду, доминируя на поверхности визуального ландшафта как часть эстетической привлекательности для посетителей национального парка Йосемити , национального парка Секвойи и залива пожаров . [ 38 ] : 2 Оранжевые и желтые лишайны добавляют в атмосферу пустынных деревьев, тундры и скалистых побережье. Сложные сетки лишайников, висящих из ветвей деревьев, добавляют таинственный аспект в леса. Фрутикозные лишайники используются в модельных железных дорогах [ 158 ] и другие моделирующие хобби как материал для изготовления миниатюрных деревьев и кустарников.
В литературе
[ редактировать ]В ранней мидрашической литературе еврейское слово « Вайлафет » в Рут 3: 8 объясняется как ссылка на Рут, развязывающуюся вокруг Боаза, как Лишай. [ 159 ] Арабский врач 10-го века Аль-Тамими упоминает, что лишайники растворяются в уксусе и розовой воде, используемых в его день для лечения кожных заболеваний и сыпей. [ 160 ]
Сюжет Джона Уиндхэма научно-фантастического романа , с которыми сталкивается с лишайкой, вращается вокруг антивозрастного химического вещества, извлеченного из лишайника.
История
[ редактировать ]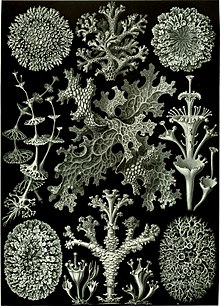
Хотя лишайники были признаны организмами в течение довольно долгого времени, только в 1867 году, когда швейцарский ботаник Саймон Швенденер предложил свою двойную теорию лишайников, лишайники представляют собой комбинацию грибов с водорослями или цианобактериями, посредством которых истинная природа лишайна ассоциации лишайна начал появляться. [ 161 ] Гипотеза Швенденера, которая в то время не имела экспериментальных доказательств, возникла из -за его обширного анализа анатомии и развития у лишайников, водорослей и грибов с использованием светового микроскопа . Многие из ведущих лихенологов в то время, такие как Джеймс Кромби и Ниланддер , отвергли гипотезу Швенденгера, потому что консенсус заключался в том, что все живые организмы были автономными. [ 161 ]
Другие выдающиеся биологи, такие как Генрих Антон де Бари , Альберт Бернхард Фрэнк , Беатрикс Поттер , Мелхиор Треуб и Герман Хелригель , не так быстро отвергли идеи Швенденгера и концепция вскоре распространились в другие области изучения, такие как микробные, растения, животное, животное, и концепция распространились в другие области изучения, такие как микробные, растения, животное, и человеческие патогены. [ 161 ] [ 162 ] [ 163 ] Когда были окончательно идентифицированы сложные отношения между патогенными микроорганизмами и их хозяевами, гипотеза Швенденера начала набирать популярность. Дальнейшее экспериментальное доказательство двойной природы лишайников было получено, когда Евген Томас опубликовал свои результаты в 1939 году в первом успешном эксперименте по повторному синтезу. [ 161 ]
В 2010 -х годах был обнаружен новый аспект партнерства с грибами. Toby Spribille и коллеги обнаружили, что многие типы лишайников, которые давно считали пары Ascomycete - Algae, на самом деле были Ascomycete - Basidiomycete - Algae Trios. Третий симбиотический партнер во многих лишайниках - базидиомицетовые дрожжи. [ 1 ] [ 164 ]
Смотрите также
[ редактировать ]- Лихенология
- Лишайники и азот езды на велосипеде
- Микофикобиоз - симбиоз, в котором гриб живет в макроскопическом таллосе пресной воды и морских водорослей; Технически не лишайник, а подобное явление, где грибы и водоросли находятся в симбиозе
Примечания
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый Сприбиль, Тоби; Туовинен, Вера; Resl, Филипп; Вандерпул, Дэн; Волински, Хеймо; Айме, М. Кэтрин; Шнайдер, Кевин; Stabentheiner, Edith; TOOME-HELLER, MERJE (21 июля 2016 г.). «Дрожжи базидиомицета в коре макролихенов аскомицета» . Наука . 353 (6298): 488–92. Bibcode : 2016sci ... 353..488s . doi : 10.1126/science.aaf8287 . ISSN 0036-8075 . PMC 5793994 . PMID 27445309 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а Лепп, Хейно (7 марта 2011 г.). "Что такое лишайник?" Полем Австралийский национальный ботанический сад. Архивировано из оригинала 2 июля 2014 года . Получено 10 октября 2014 года .
- ^ « Введение в лишайники - альянс между королевствами ». Калифорнийский музей палеонтологии. Архивировано 22 августа 2014 года на машине Wayback .
- ^ Jump up to: а беременный в дюймовый и Бродо, Ирвин М. и Дулаан Шарнофф, Сильвия (2001) Лишайники Северной Америки ISBN 978-0300082494 .
- ^ «Дрожжи появляются как скрытый третий партнер в лишайном симбиозе» . Scienceday . 21 июля 2016 года . Получено 31 марта 2024 года .
- ^ Кумпула, Жуко; Lefrère, Stéphanie C.; Nieminen, Mauri (1 января 2004 г.). «Использование пастбищ из лесных лишай на оленях зимой с легкими снежными условиями» . Арктика . 57 (3). doi : 10.14430/arctic504 . ISSN 1923-1245 .
- ^ Фрёберг, Ларс; Баур, Анетт; Баур, Бруно (январь 1993 г.). «Дифференциальное повреждение травоядных животных кальцикологическим лишатам от улиток» . Лихенолог . 25 (1): 83. DOI : 10.1017/S002428299300009X . ISSN 0024-2829 .
- ^ Лейнаас, Ганс Петтер; Fjellberg, Arne (июнь 1985 г.). «Структура среды обитания и стратегии жизненной истории двух частично симпатрических и тесно связанных, лишайных кормляющих видов Коллемболан» . Оикос . 44 (3): 448. Bibcode : 1985oikos..44..448L . doi : 10.2307/3565786 . ISSN 0030-1299 . JSTOR 3565786 .
- ^ Герсон, США (октябрь 1973 г.). «Ассоциации лишайников-артропода» . Лихенолог . 5 (5–6): 434–443. doi : 10.1017/s0024282973000484 . ISSN 0024-2829 . S2CID 85020765 .
- ^ Jump up to: а беременный Галлоуэй, DJ (13 мая 1999 г.). "Лишайский глоссарий" . Австралийский национальный ботанический сад. Архивировано из оригинала 6 декабря 2014 года.
- ^ Маргулис, Линн ; Баррено, Ева (2003). «Глядя на лишайники» . Биоссака . 53 (8): 776. doi : 10.1641/0006-3568 (2003) 053 [0776: lal] 2.0.co; 2 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. Шарнофф, Стивен (2014 ) . ISBN 978-0-300-19500-2
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k Шпеер, Брайан Р; Бен Вагонер (май 1997 г.). «Лишайники: история жизни и экология» . Калифорнийский музей палеонтологии. Архивировано из оригинала 2 мая 2015 года . Получено 28 апреля 2015 года .
- ^ Сиссон, Лив; Вигус, Паула (2023). Грибы Aotearoa: полезное полевое гид . Окленд, Новая Зеландия: книги пингвинов. п. 66. ISBN 978-1-76104-787-9 Полем OCLC 1372569849 .
- ^ Jump up to: а беременный в дюймовый и «Лишайники: систематика, Музей палеонтологии Калифорнийского университета» . Архивировано с оригинала 24 февраля 2015 года . Получено 10 октября 2014 года .
- ^ Lendemer, JC (2011). «Таксономическое пересмотр североамериканских видов Lepraria SL, которые производят диварикатическую кислоту, с заметками о видах вида рода L. Incana ». Микология . 103 (6): 1216–1229. doi : 10.3852/11-032 . PMID 21642343 . S2CID 34346229 .
- ^ Jump up to: а беременный в дюймовый Casano, LM; Поля, эм; García-Breijo, FJ; REIG-ACT, J; Gasulla, F; Из Хойо, а; Гера, а; Баррено и (2011). «Два водоросли Требуксии с различными физиологическими характеристиками постоянно присутствуют в Лишайне Талли из Рамалины Фаринацеи . Соавтор против конкуренции?». Микробиология окружающей среды (представленная рукопись). 13 (3): 806–818. Bibcode : 2011envi..13..806c . Doi : 10.1111/j.1462-2920.2010.02386.x . HDL : 10251/60269 . PMID 21134099 .
- ^ Jump up to: а беременный Honegger, R. (1991) Эволюция грибов: симбиоз и морфогенез, симбиоз как источник эволюционных инноваций , Margulis, L. и Fester, R. (Eds). Кембридж, Массачусетс, США: MIT Press, с. 319–340.
- ^ Jump up to: а беременный Grube, M; Кардинал, м; De Castro, JV Jr.; Мюллер, ч; Берг, Г. (2009). «Видоспецифичные структурные и функциональные разнообразие бактериальных сообществ у лишайных симбиозов» . Журнал ISME . 3 (9): 1105–1115. Bibcode : 2009ismej ... 3.1105G . doi : 10.1038/ismej.2009.63 . PMID 19554038 .
- ^ Jump up to: а беременный Э., Эррера-Камос М., Газулла Ф., Дж. ( 2008 Баррено ) . Совместное собрание ABLS.
- ^ Jump up to: а беременный Моррис Дж., Пурвис В. (2007). Лишайники (жизнь) . Лондон: Музей естественной истории. п. 19. ISBN 978-0-565-09153-8 .
- ^ «Лишайник - мелочи, которые имеют значение (Служба национальных парков США)» . www.nps.gov . Получено 17 ноября 2023 года .
- ^ Харпер, Дуглас. «Лишайник» . Онлайн этимологический словарь .
- ^ лишайник . Чарлтон Т. Льюис и Чарльз Шорт. Латинский словарь по проекту Персей .
- ^ λειχήν . Лидделл, Генри Джордж ; Скотт, Роберт ; Грек -английский лексикон в проекте Персея .
- ^ отсутствует в Лидделле и Скотте .
- ^ Beekes, Robert SP (2010). "SV λειχήν, λείχω". Этимологический словарь греческого . Лейден Индоевропейский этимологический словарь. Тол. 1. С помощью ЛЮДИАНА ВАН БИК. Лейден, Бостон: Брилл. С. 846–47. ISBN 9789004174184 .
- ^ «Лишайник» . Кембриджский словарь . Получено 8 сентября 2022 года .
- ^ Вордсворт, Дот (17 ноября 2012 г.). «Лишайник» . Spectator.co.uk. Архивировано из оригинала 23 декабря 2014 года . Получено 8 сентября 2022 года .
- ^ Оксфордский словарь английского языка цитирует только произношение "СИКОВАЯ": «Лишайник» . Оксфордский английский словарь (онлайн изд.). Издательство Оксфордского университета . Получено 10 января 2018 года . в учреждении или (Требуется членство участвующее учреждение .)
- ^ Jump up to: а беременный в дюймовый и фон «Лишайники и Бриофиты, Мичиганский государственный университет, 10-25-99» . Архивировано из оригинала 5 октября 2011 года . Получено 10 октября 2014 года .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж Словарь лишайников, лишайника Северной Америки Информация, Сильвия и Стивен Шарнофф, [1] Архивировано 20 января 2015 года на машине Wayback
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k «Глоссарий лишайника Алана Сильверсайда (PZ), Алан Сильверсейд» . Архивировано из оригинала 31 октября 2014 года . Получено 10 октября 2014 года .
- ^ Jump up to: а беременный в дюймовый и фон глин час Dobson, FS (2011). Лишайны, иллюстрированное руководство по британским и ирландским видам . Slough, Великобритания: Richmond Publishing Co. ISBN 9780855463151 .
- ^ Jump up to: а беременный в дюймовый и фон глин «Фолиозные лишайники, типы лишайников, Аллан Сильверсейд» . Архивировано с оригинала 19 октября 2014 года . Получено 10 октября 2014 года .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м Mosses Lichens & Ferns из Северо -Западной Северной Америки , Дейл Х. Витт, Джанет Э. Марш, Робин Б. Бови, издательская компания Lone Pine, ISBN 0-295-966666-1
- ^ Jump up to: а беременный в дюймовый «Лишайники, корпорация Saguaro-Juniper» . Архивировано из оригинала 10 мая 2015 года . Получено 10 октября 2014 года .
- ^ Jump up to: а беременный в McCune, B.; Grenon, J.; Martin, E.; Mutch, LS; Мартин, EP (март 2007 г.). «Лишайники в отношении вопросов управления в национальных парках Сьерра -Невада». Североамериканские грибы . 2 : 1–39. doi : 10.2509/pnwf.2007.002.003 (неактивный 16 апреля 2024 г.).
{{cite journal}}: CS1 Maint: doi неактивен с апреля 2024 года ( ссылка ) - ^ Jump up to: а беременный в дюймовый Мичиган Лишайны, Джули Джонс Медлин, Б. Джайн издательство, 1996, ISBN 0877370397 , 9780877370390, [2] Архивировано 24 ноября 2016 года на машине Wayback
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж Лишайны: подробнее о морфологии, Музей палеонтологии Калифорнийского университета, [3] Архивировано 28 февраля 2015 года на машине Wayback
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л «Лишайные фотобионты, Университет штата Небраска Омаха» (PDF) . Архивировано из оригинала (PDF) 6 октября 2014 года.
- ^ Jump up to: а беременный «Глоссарий лишайника Алана Сильверсайда (GO), Алан Сильверсейд» . Архивировано с оригинала 2 ноября 2014 года . Получено 10 октября 2014 года .
- ^ Jump up to: а беременный в Büdel, B.; Scheidegger, C. (1996). «Морфология таллуса и анатомия». Лишайская биология . С. 37–64. doi : 10.1017/cbo9780511790478.005 . ISBN 9780511790478 .
- ^ Jump up to: а беременный Хеймарссон, Старри; Heidmarsson, Starri (1996). «Пруна как таксономический характер в роде лишайника Dermatocarpon ». Бриолог . 99 (3): 315–320. doi : 10.2307/32444302 . JSTOR 3244302 .
- ^ Шарнофф, Сильвия и Шарнофф, Стивен. «Биология лишайника и окружающая среда», архивировав 17 октября 2015 года на машине Wayback . sharnoffphotos.com
- ^ Pogoda, CS; Хранители, кг; Lendemer, JC; Кейн, Северная Каролина; Трипп, EA (2018). «Снижение сложности митохондриальных геномов в лишайных грибах проливает свет на архитектуру генома облигатных симбиозов-онлайн-библиотека Wiley». Молекулярная экология . 27 (5): 1155–1169. doi : 10.1111/mec.14519 . PMID 29417658 . S2CID 4238109 .
- ^ Сприбиль, Тоби; Туовинен, Вера; Resl, Филипп; Вандерпул, Дэн; Волински, Хеймо; Айме, М. Кэтрин; Шнайдер, Кевин; Stabentheiner, Edith; Toome-Heller, Merje; Тор, Гёран; Mayrhofer, Helmut; Йоханнессон, Ханна; Маккатчон, Джон П. (29 июля 2016 г.). «Дрожжи базидиомицета в коре макролихенов аскомицета» . Наука . 353 (6298): 488–492. Bibcode : 2016sci ... 353..488s . doi : 10.1126/science.aaf8287 . PMC 5793994 . PMID 27445309 .
- ^ Skaloud, P; Peksa, O (2010). «Эволюционные выводы, основанные на ее последовательностях рДНК и актина, показывают обширное разнообразие общей лишайники, содержащей алга, астерохлорис (Trebouxiophyceae, Chlorophyta)». Молекулярная филогенетика и эволюция . 54 (1): 36–46. doi : 10.1016/j.ympev.2009.09.035 . PMID 19853051 .
- ^ Сприбиль, Тоби; Туовинен, Вера; Resl, Филипп; Вандерпул, Дэн; Волински, Хеймо; Айме, М. Кэтрин; Шнайдер, Кевин; Stabentheiner, Edith; Toome-Heller, Merje; Тор, Гёран; Mayrhofer, Helmut (29 июля 2016 года). «Дрожжи базидиомицета в коре макролихенов аскомицета» . Наука . 353 (6298): 488–492. Bibcode : 2016sci ... 353..488s . doi : 10.1126/science.aaf8287 . ISSN 0036-8075 . PMC 5793994 . PMID 27445309 .
- ^ Jump up to: а беременный в дюймовый Рамель, Гордон. "Что такое лишайник?" Полем Веб. Архивировано с оригинала 19 января 2015 года . Получено 20 января 2015 года .
- ^ Jump up to: а беременный Ахмаджян В. (1993). Симбиоз лишайника . Нью -Йорк: Джон Уайли и сыновья. ISBN 978-0-471-57885-7 .
- ^ Jump up to: а беременный Honeger, R. (1988). "Микобонты" В Нэш III, Th (ed.). Лишайская биология Кембридж: ISBN 978-0-521-45368-4 .
- ^ Ferry, BW, Baddeley, MS & Hawksworth, DL (редакторы) (1973) Загрязнение воздуха и лишайники . Athlone Press, Лондон.
- ^ Rose CI, Hawksworth DL (1981). «Реколонизация лишайника в лондонском чистого воздуха». Природа . 289 (5795): 289–292. Bibcode : 1981natur.289..289r . doi : 10.1038/289289a0 . S2CID 4320709 .
- ^ Hawksworth, DL и Rose, F. (1976) Лишайники как мониторы загрязнения . Эдвард Арнольд, серия Института биологии, № 66. ISBN 07131255551
- ^ "Oak Moss Absolute Oil, Evernia prunastri, парфюмерная фиксация" . Архивировано из оригинала 25 декабря 2014 года . Получено 19 сентября 2014 года .
- ^ Skogland, Terje (1984). «Организация диких оленей в Niche-Niche». Экография . 7 (4): 345. Bibcode : 1984ecogr ... 7..345s . doi : 10.1111/j.1600-0587.1984.tb01138.x .
- ^ Lawrey, James D.; Дидерих, Пол (2003). «Лихениколевые грибы: взаимодействия, эволюция и биоразнообразие» (PDF) . Бриолог . 106 : 80. doi : 10.1639/0007-2745 (2003) 106 [0080: lfieab] 2.0.co; 2 . S2CID 85790408 . Архивировано (PDF) из оригинала 3 января 2011 года . Получено 2 мая 2011 года .
- ^ Hagiwara K, Wright PR, et al. (Март 2015 г.). «Сравнительный анализ антиоксидантных свойств исландских и гавайских лишайников». Экологическая микробиология . 18 (8): 2319–2325. doi : 10.1111/1462-2920.12850 . PMID 25808912 . S2CID 13768322 .
- ^ Odabasoglu F, Aslan A, Cakir A, et al. (Март 2005 г.). «Антиоксидантная активность, снижение мощности и общее содержание фенольных видов у некоторых видов лишайников». Fitoterapia . 76 (2): 216–219. doi : 10.1016/j.fitote.2004.05.012 . PMID 15752633 .
- ^ Jump up to: а беременный Хаук, Маркус; Юргенс, Саша-Рен; Leuschner, Christoph (2010). «Норстиктивная кислота: корреляции между ее физико-химическими характеристиками и экологическими предпочтениями лишайников, производящих этот депсидон». Экологическая и экспериментальная ботаника . 68 (3): 309. Bibcode : 2010enveb..68..309h . doi : 10.1016/j.envexpbot.2010.01.003 .
- ^ Jump up to: а беременный «Веб, рост и развитие земной жизни в лишайках» . Earthlife.net. Архивировано из оригинала 28 мая 2015 года . Получено 12 октября 2014 года .
- ^ Rosenwinkel, Swenja; Korup, Oliver; Landgraf, Angela; Dzhumabaeva, Atyrgul (2015). "Limits to lichenometry" . Quaternary Science Reviews . 129 : 229–238. Bibcode : 2015QSRv..129..229R . doi : 10.1016/j.quascirev.2015.10.031 .
- ^ Миллер, GH; Эндрюс, JT (апрель 1972 г.). «Кватернарная история северного полуострова Камберленд, Остров Ист -Баффин, NWT, Канада, часть VI: предварительная кривая роста лишайников для ризокарпона Geographicum» . Бюллетень GSA . 83 (4): 1133–1138. doi : 10.1130/0016-7606 (1972) 83 [1133: Qhoncp] 2.0.co; 2 .
- ^ Haworth, Leah A.; Calkin, Parker E.; Эллис, Джеймс М. (1986). «Прямое измерение роста лишайников в диапазоне Центрального Брукса, Аляски, США, и его применение к лишайнометрическим датированию, арктическим и альпийским исследованиям» . Арктика и альпийские исследования . 18 (3): 289–296. doi : 10.2307/1550886 . JSTOR 1550886 .
- ^ Бенедикт, Джеймс Б. (январь 2009 г.). «Обзор лихенометрических знакомств и его применения к археологии» . Американская древность . 74 (1): 143–172. doi : 10.1017/s0002731600047545 . S2CID 83108496 .
- ^ Нэш III, Томас Х. (2008). "Введение" В Нэш III, Th (ed.). Биология лишайника (2 -е изд.). Кембридж: стр. 1–8 doi : 10.1017/ cbo9705050478.002 ISBN 978-0-521-69216-8 .
- ^ Болдуин, Эмили (26 апреля 2012 г.). «Лишайник переживает суровую среду Марса» . Skymania News. Архивировано из оригинала 28 мая 2012 года . Получено 27 апреля 2012 года .
- ^ Шелдрак, Мерлин (2020). Запутанная жизнь: как грибы делают наши миры, передумают наше мнение и формируют наше будущее . Бодли голова. п. 94. ISBN 978-1847925206 .
- ^ «ESA - человеческий космический полет и разведка - лишайник выживает в космосе» . Архивировано из оригинала 26 февраля 2010 года . Получено 16 февраля 2010 года .
- ^ Санчо, LG; De la Torre, R.; Horneck, G.; Ascaso, C.; De los reos, a.; Pintado, A.; Wierzchos, J.; Шустер М. (2007). «Лишайники выживают в космосе: результаты эксперимента лишайника 2005 года». Астробиология . 7 (3): 443–454. Bibcode : 2007asbio ... 7..443s . Doi : 10.1089/ast2006.0046 . PMID 17630840 . S2CID 4121180 .
- ^ Eichorn, Susan E., Evert, Ray F. и Raven, Peter H. (2005). Биология растений . Нью -Йорк: WH Freeman and Company. п. 1 ISBN 0716710072 .
- ^ Кук, Ребекка; McFarland, Kenneth (1995). Генеральная ботаника 111 Лабораторное руководство . Ноксвилл, Теннесси: Университет Теннесси. п. 104
- ^ Jump up to: а беременный Я Рай; Б. Бергман; Улла Расмуссен (31 июля 2002 г.). Цианобактер в симбиозе Спрингер. П. 59. ISBN 978-1-4020-0777-4 Полем Архивировано из оригинала 31 декабря 2013 года . Получено 2 июня 2013 года .
- ^ Закон, Р.; Льюис, DH (ноябрь 1983 г.). «Биотическая среда и поддержание показаний по полу-абсолютному положению от взаимных симбиозов» . Биологический журнал Линневого общества . 20 (3): 249–276. doi : 10.1111/j.1095-8312.1983.tb01876.x . ISSN 0024-4066 .
- ^ Шкалуд, Павел; Стейн, Яна; Редкий, Тереза; Ванчарова, Люси; Пекса, Ондхей (4 мая 2015 г.). «Сборка сложной головоломки биоразнообразия водорослей: разграничение видов в Genusasterochloris (Trebouxiophyceae, хлорофит)» . Журнал Phycology . 51 (3): 507–527. Bibcode : 2015jpcgy..51..507s . Doi : 10.1111/jpy.12295 . ISSN 0022-3646 . PMID 26986666 . S2CID 25190572 .
- ^ Рамель, Гордон. «Репродуктивные структуры лишайников» . Архивировано из оригинала 28 февраля 2014 года . Получено 22 августа 2014 года .
- ^ Jump up to: а беременный в Муртаг Г.Дж., Дайер П.С., Криттенден П.Д. (апрель 2000 г.). «Секс и единственный лишайник». Природа . 404 (6778): 564. Bibcode : 2000natur.404..564M . doi : 10.1038/35007142 . PMID 10766229 . S2CID 4425228 .
- ^ Jump up to: а беременный в Kirk PM, Cannon PF, Minter DW, Stalpers JA (2008). Словарь грибов (10 -е изд.). Уоллингфорд: Каби. С. 378–381. ISBN 978-0-85199-826-8 .
- ^ Люкинг, Роберт; Ходкинсон, Брендан П.; Ливитт, Стивен Д. (2017). «Классификация лишайных грибов 2016 года в Ascomycota и Basidiomycota, поднимающей тысячу родов». Бриолог . 119 (4): 361–416. doi : 10.1639/0007-2745-119.4.361 . JSTOR 44250015 . S2CID 90258634 .
- ^ Люкинг, Роберт; Ходкинсон, Брендан П.; Ливитт, Стивен Д. (2017). «Исправления и поправки к классификации лишайных грибов 2016 года в Ascomycota и Basidiomycota». Бриолог . 120 (1): 58–69. doi : 10.1639/0007-2745-120.1.058 . S2CID 90363578 .
- ^ Люкинг, Роберт; Rivas-Plata, E.; Чавес, JL; Umaña, L.; Sipman, HJM (2009). «Сколько там тропических лишайников… правда?». Bibliotheca Lichenologica . 100 : 399–418.
- ^ Jump up to: а беременный в «Форма и структура - Sticta и Dendriscocaulon » . Австралийский национальный ботанический сад. Архивировано из оригинала 28 апреля 2014 года . Получено 18 сентября 2014 года .
- ^ Lutzoni, F.; Кауфф, Ф.; Кокс, CJ; McLaughlin, D.; Celio, G.; Dentinger, B.; Padamsee, M.; Хиббетт, Д.; и др. (2004). «Сборка грибкового дерева жизни: прогресс, классификация и эволюция субклеточных черт» . Американский журнал ботаники . 91 (10): 1446–1480. doi : 10.3732/ajb.91.10.1446 . PMID 21652303 . S2CID 9432006 .
- ^ Сандерс, WB; Мо, RL; Ascaso, C. (2004). «Стоиндальный морской лишайник, образованный грибом пиреномицета Verrucaria tavaresiae (Ascomycotina) и коричневой водоросли Petga Petroderma Maculiforme (Phaeophyceae): организация Thallus и взаимодействие Symbiont - NCBI». Американский журнал ботаники . 91 (4): 511–22. doi : 10.3732/ajb.91.4.511 . HDL : 10261/31799 . PMID 21653406 .
- ^ «Взаимные музея между грибами и водорослями - музей Нью -Брансуика» . Архивировано из оригинала 18 сентября 2018 года . Получено 4 октября 2018 года .
- ^ Миллер, Кэти Энн; Перес-Форга, Серхио (май 2018 г.). «Вызов концепции лишайника: Turgidosculum ulvae - Cambridge» . Лихенолог . 50 (3): 341–356. doi : 10.1017/s0024282918000117 . S2CID 90117544 . Архивировано с оригинала 7 октября 2018 года . Получено 7 октября 2018 года .
- ^ Rybalka, N.; Волк, М.; Андерсен, Ра; Фридл, Т. (2013). «Конгруэнтность хлоропласта - эволюционная биология BMC - Biomed Central» . BMC Эволюционная биология . 13 : 39. DOI : 10.1186/1471-2148-13-39 . PMC 3598724 . PMID 23402662 .
- ^ Лутцони, Франсуа; Пейджел, Марк; Риб, Валери (21 июня 2001 г.). «Основные грибковые линии получены из лишайных симбиотических предков» . Природа . 411 (6840): 937–940. Bibcode : 2001natur.411..937L . doi : 10.1038/35082053 . PMID 11418855 . S2CID 4414913 .
- ^ Hawksworth, DL (1988). «Разнообразие грибко-альгальских симбиозов, их эволюционное значение и природа лишайников». Ботанический журнал Линневого общества . 96 : 3–20. doi : 10.1111/j.1095-8339.1988.tb00623.x . S2CID 49717944 .
- ^ Jump up to: а беременный в Rikkinen J. (1995). «Что за красивыми цветами? Исследование фотобиологии лишайников». Bryobrothera . 4 (3): 375–376. doi : 10.2307/32444316 . JSTOR 3244316 .
- ^ Лукинг, Роберт (25 февраля 2015 г.). «Один гриб - два лишайника» . Полевой музей естественной истории . Получено 25 февраля 2022 года .
- ^ Friedl, T.; Бюдель Б. (1996). «Photobionts». В Нэш III, Th (ed.). Лишайская биология . Кембридж: издательство Кембриджского университета. С. 9–26. doi : 10.1017/cbo9780511790478.003 . ISBN 978-0-521-45368-4 .
- ^ Jump up to: а беременный «Глоссарий лишайника Алана Сильверсайда (AF), Алан Сильверсейд» . Архивировано из оригинала 31 октября 2014 года . Получено 10 октября 2014 года .
- ^ Халленбек, Патрик С. (18 апреля 2017 г.). Современные темы в фототрофных прокариотах: экологические и прикладные аспекты . Спрингер. ISBN 9783319462615 Полем Архивировано с оригинала 4 октября 2018 года . Получено 4 октября 2018 года .
- ^ Jump up to: а беременный Rikkinen, J. (2002). «Гильдии лишайника разделяют связанные цианобактериальные симбионты». Наука . 297 (5580): 357. doi : 10.1126/science.1072961 . PMID 12130774 . S2CID 35731669 .
- ^ О'Брайен, H.; Miadlikowska, J.; Lutzoni, F. (2005). «Оценка специализации хозяина в симбиотических цианобактериях, связанных с четырьмя близкородственными видами гриба лишайника Peltigera » . Европейский журнал Phycology . 40 (4): 363–378. Bibcode : 2005ejphy..40..363o . doi : 10.1080/09670260500342647 . S2CID 4094256 .
- ^ Guzow-Krzeminska, B (2006). «Photobiont? Extibility у Thelichen Protoparmeliopsis muralis, как выявит его анализ рДНК». Лихенолог . 38 (5): 469–476. doi : 10.1017/s002428290600005068 . S2CID 85895322 .
- ^ Ohmura, Y.; Kawachi, M.; Касаи, Ф.; Ватанабе М. (2006). «Генетические комбинации симбионтов в вегетативно воспроизводимом лишайнике, Parmotrema tinctorum , на основе его последовательностей рДНК» (2006) ». Бриолог . 109 : 43–59. DOI : 10.1639/0007-2745 (2006) 109 [0043: GCOSIA] 2.0. ; . 2 .co
- ^ Пирси-Нормор (2006). «Лишайник, образующий лишайник Asco-Mycete Evernia Mesomorpha, ассоциируется с множествомгенгенотипами Trebouxia Jamesii » . Новый фитолог . 169 (2): 331–344. doi : 10.1111/j.1469-8137.2005.01576.x . PMID 16411936 .
- ^ Лутцони, Франсуа; Пейджел, Марк; Reeb, Valérie (2001). «Основные грибковые линии получены из лишайных симбиотических предков» . Природа . 411 (6840): 937–940. Bibcode : 2001natur.411..937L . doi : 10.1038/35082053 . PMID 11418855 . S2CID 4414913 .
- ^ Jump up to: а беременный «Лишайники: ископаемые записи» Архивировали 25 января 2010 года на машине Wayback , Музей палеонтологии Калифорнийского университета.
- ^ Speer BR, Wagoner B. Калифорнийский музей палеонтологии. Архивировано из оригинала 25 января 2010 года . Получено 16 февраля 2010 года .
- ^ Пунар -младший, иди. (1992). Жизнь в янтаре . Издательство Стэнфордского университета.
- ^ Петерсон Эб. (2000). «Опубликованный ископаемый лишайник (Lobariaceae)». Лихенолог . 32 (3): 298–300. Bibcode : 2000thlic..32..298p . doi : 10.1006/lich.1999.0257 . S2CID 83629248 .
- ^ Тейлор, Теннесси; Хасс, Х.; Реми, W.; Kerp, H. (1995). «Самый старый ископаемый лишайник» . Природа . 378 (6554): 244. Bibcode : 1995nater.378..244t . doi : 10.1038/378244A0 . S2CID 4353572 .
- ^ Тейлор WA, Free CB, Helgemo R, Ochoada J (2004). «SEM -анализ Spongiophyton, интерпретируемый как ископаемый лишайник». Международный журнал растительных наук . 165 (5): 875–881. doi : 10.1086/422129 . S2CID 85382155 .
- ^ Ярен, ах; Портер, с.; Kuglitsch, JJ (2003). «Лишайский метаболизм выявлен в ранних девонских наземных организмах». Геология . 31 (2): 99–102. Bibcode : 2003geo .... 31 ... 99J . doi : 10.1130/0091-7613 (2003) 031 <0099: lmiied> 2,0.co; 2 . ISSN 0091-7613 . S2CID 49241170 .
- ^ Флетчер, BJ; Пиво, диджей ; Chaloner, WG (2004). «Стабильные изотопы углерода и метаболизм наземного девонского организма Spongiophyton ». Геобиология . 2 (2): 107–119. Bibcode : 2004gbio .... 2..107f . doi : 10.1111/j.1472-4677.2004.00026.x . S2CID 85079041 .
- ^ Эдвардс D; AX L (2012). «Доказательства грибкового сродства к Nematasketum, близкому союзнику прототакситов» . Ботанический журнал Линневого общества . 168 : 1–18. doi : 10.1111/j.1095-8339.2011.01195.x .
- ^ Retallack GJ; Landing, E. (2014). «Сродство и архитектура девонских стволов прототакситов Loganii ». Микология . 106 (6): 1143–1156. doi : 10.3852/13-390 . PMID 24990121 . S2CID 20013625 .
- ^ Каратигин IV; Snigirevskaya ns; Викулин С.В. (2009). «Самая древняя наземная лишайника Winfrenatia reticulata : новая находка и новая интерпретация». Палеонтологический журнал . 43 (1): 107–114. Bibcode : 2009palj ... 43..107K . doi : 10.1134/s0031030109010110 . S2CID 85262818 .
- ^ Karatygin IV, Snigirevskaya NS, Vikulin SV (2007). «Два типа симбиоза с участием грибов из ранних девонских экосистем» . XV Конгресс европейских микологов, Сен -Петербург, Россия, 16–21 сентября 2007 года . 1 (1): 226. Архивировано из оригинала 24 апреля 2013 года . Получено 18 февраля 2011 года .
- ^ «Лишайники намного моложе, чем думали ученые - вероятно, развивались миллионы лет после растений» . 15 ноября 2019 года. Архивировано с оригинала 18 декабря 2019 года . Получено 18 ноября 2019 года .
- ^ Хансен, Карен; Перри, Брайан А.; Dranginis, Andrew W.; Пфистер, Дональд Х. (май 2013). «Филогения очень разнообразного семейства Кубка-Фунгуса Pyronemataceae (Pezizomycetes, Ascomycota) проясняет отношения и эволюцию отбранных черт жизни» . Молекулярная филогенетика и эволюция . 67 (2): 311–335. Bibcode : 2013molpe..67..311h . doi : 10.1016/j.ympev.2013.01.014 . ISSN 1055-7903 . PMID 23403226 .
- ^ Schoch Cl; Спел GH; Лопес-Гральдес F; Townsend JP; Miadlikowska J; Hofstetter V; Robbertse B; Matheny PB; и др. (2009). «Ascomycota Tree of Life: филогения по типу, проясняет происхождение и эволюцию фундаментальных репродуктивных и экологических признаков» . Система Биол . 58 (2): 224–239. doi : 10.1093/sysbio/syp020 . PMID 20525580 .
- ^ Гаргас, а; Depriest, Pt; Grube, M; Tehler, A (1995). «Многочисленные происхождения лишайников симбиозов в грибах, предложенных филогениями RDNA SSU». Наука . 268 (5216): 1492–1495. Bibcode : 1995sci ... 268.1492g . doi : 10.1126/science.7770775 . PMID 7770775 . S2CID 563601 .
- ^ Honegger R. (1998). «Симбиоз лишайника - что такого впечатляющего в этом?» (PDF) . Лихенолог . 30 (3): 193–212. doi : 10.1017/s002428299200015x . Архивировано (PDF) из оригинала 26 апреля 2019 года . Получено 30 января 2019 года .
- ^ Wedin M, Döring H, Gilenstam G (2004). «Сапротрофия и лишайнизация как варианты одного и того же вида Furgl на различных субстратах: экологическая пластичность и грибковой образ жизни в комплексе Contrema - Conotrema » . Новый фитолог . 16 (3): 459–465. doi : 10.1111/j.1469-8137.2004.01198.x .
- ^ Юань Х, Сяо С., Тейлор Т.Н. (2005). «Симбиоз, похожий на лишайника 600 миллионов лет назад». Наука . 308 (5724): 1017–1020. Bibcode : 2005sci ... 308.1017y . doi : 10.1126/science.1111347 . PMID 15890881 . S2CID 27083645 .
- ^ Retallack GJ (2015). «Акритарх свидетельство позднего докембрийского адаптивного излучения грибов» (PDF) . Botanica Pacifica . 4 (2): 19–33. doi : 10.17581/bp.2015.04203 . Архивировано (PDF) из оригинала 22 декабря 2016 года . Получено 22 декабря 2016 года .
- ^ Retallack GJ. (2007). Знаменитая эдиакарран окаменелости " журнал Алхрирование: австралийский 31 (3): 215–2 Bibcode : 2007 Alch ... 215 ... 215r doi : 10.1080/03151510701484705 . S2CID 17181699 .
- ^ Retallack GJ. (1994). «Были ли лишасины эдиакарана?». Палеобиология . 20 (4): 523–544. Bibcode : 1994pbio ... 20..523r . doi : 10.1017/s0094837300012975 . JSTOR 2401233 . S2CID 129180481 .
- ^ Switek B (2012). «Спорный утверждение ставит жизнь на землю на 65 миллионов лет раньше» . Природа . doi : 10.1038/nature.2012.12017 . S2CID 130305901 . Архивировано из оригинала 1 января 2013 года . Получено 2 января 2013 года .
- ^ Retallack, GJ; Данн, Кл; Saxby, J. (2015). «Проблемная мезопротерозойскую ископаемость Городиския из национального парка Глейшер, штат Монтана, США». Докембрийское исследование . 226 : 125–142. Bibcode : 2013prer..226..125r . doi : 10.1016/j.precamres.2012.12.005 .
- ^ Retallack, GJ; Krull, Es; Thackray, GD; Паркинсон Д. (2013). «Проблемные урн-в форме окаменелости от палеопротерозойского (2,2 га) палеозола в Южной Африке». Докембрийское исследование . 235 : 71–87. Bibcode : 2013prer..235 ... 71r . doi : 10.1016/j.precamres.2013.05.015 .
- ^ Люкинг, Роберт; Нельсен, Мэтью П. (2018), «Эдиакаранцы, протолихены и лишайные пенициллиум» , преобразующая палеоботания , Elsevier, с. 551–590, doi : 10.1016/b978-0-12-813012-4.00023-1 , ISBN /B978-0-12-813012- 978-0-12-813012-4 , Получено 14 ноября 2020 года
- ^ Нельсен, Мэтью П.; Люкинг, Роберт; Бойс, С. Кевин; Lumbsch, H. Thorsten; Ри, Ричард Х. (январь 2020 г.). «Нет поддержки появления лишайников до эволюции сосудистых растений» . Геобиология . 18 (1): 3–13. Бибкод : 2020GBIO ... 18 .... 3n . doi : 10.1111/gbi.12369 . ISSN 1472-4677 . PMID 31729136 . S2CID 208034624 .
- ^ Песня, Hyunjeong; Ким, Ки-Тэ; Парк, Сук-Янг; Ли, Гир-Вон; Чой, Jaeyoung; Чон, Чонбум; Cheong, Kyeongchae; Чой, Гобонг; Хур, Чже-Сеун; Ли, Юн-Хван (24 июня 2022 года). «Сравнительный геномный анализ грибов, образующих лишайника, выявляет новое понимание грибкового образа жизни» . Научные отчеты . 12 (1): 10724. BIBCODE : 2022NATSR..1210724S . doi : 10.1038/s41598-022-14340-5 . ISSN 2045-2322 . PMC 9232553 . PMID 35750715 .
- ^ Кляйн, Джоанна (19 ноября 2019 г.). «В гонке, чтобы жить на суше, лишайники не обыграли растения» . New York Times .
- ^ «Для Маунт -Рашмор, просроченное мытье лица» . The Washington Post . Ассошиэйтед Пресс. 11 июля 2005 г. Получено 20 августа 2022 года .
- ^ «Загрязнение, заводный мир» . Австралийский национальный ботанический сад. Архивировано из оригинала 17 февраля 2014 года . Получено 10 октября 2014 года .
- ^ Ахмаджян, В. (1 марта 1995 г.). «Лишайники важнее, чем вы думаете» . Биоссака . 45 (3): 1. DOI : 10.1093/Bioscience/45.3.124 . Получено 14 сентября 2021 года .
- ^ Чен, Цзе; Блюм, Ганс-Петер; Бейер, Лотар (2000). «Выветривание камней, вызванных колонизацией лишайников - обзор» (PDF) . Катена . 39 (2): 121. Bibcode : 2000caten..39..121c . doi : 10.1016/s0341-8162 (99) 00085-5 . Архивировано (PDF) из оригинала 2 апреля 2015 года . Получено 21 марта 2015 года .
- ^ Джонс, Клайв Г.; Шачак, Моше (1990). «Оборудование пустынной почвы с помощью горных улитков». Природа . 346 (6287): 839. Bibcode : 1990natur.346..839J . doi : 10.1038/346839a0 . S2CID 4311333 .
- ^ Уокер, Т.Р. (2007). «Лишайники бореальных лесов Лабрадора, Канада: контрольный список». Эвансия . 24 (3): 85–90. doi : 10.1639/0747-9859-24.3.85 . S2CID 129100097 .
- ^ Oksanen, I. (2006). «Экологические и биотехнологические аспекты лишайников». Прикладная микробиология и биотехнология . 73 (4): 723–734. doi : 10.1007/s00253-006-0611-3 . PMID 17082931 . S2CID 12172616 .
- ^ Lawrey, James D. (1994). «Аллелопатия лишайника: обзор». В Inderjit; KMM Дакшини; Фрэнк А. Эйнхеллиг (ред.). Аллелопатия. Организмы, процессы и приложения . Симпозиум ACS Series. Тол. 582. Американское химическое общество. С. 26–38. doi : 10.1021/bk-1995-0582.ch002 . ISBN 978-0-8412-3061-3 .
- ^ Jump up to: а беременный в Нэш III, Томас Х. (2008). «Чувствительность лишайников к загрязнению воздуха». В Нэш III, Th (ed.). Биология лишайника (2 -е изд.). Кембридж: издательство Кембриджского университета. С. 299–314. doi : 10.1017/cbo9780511790478.016 . ISBN 978-0-521-69216-8 .
- ^ Knops, JMH; Нэш, TH III; Boucher, VL; Schlesinger, WH (1991). «Минеральный велосипед и эпифитные лишайники: последствия на уровне экосистемы». Лихенолог . 23 (3): 309–321. doi : 10.1017/s0024282991000452 . S2CID 86133860 .
- ^ Хоган, С. Майкл (2010). «Абиотический фактор» . Энциклопедия Земли . Вашингтон, округ Колумбия: Национальный совет по науке и окружающей среде. Архивировано из оригинала 8 июня 2013 года . Получено 27 октября 2013 года .
- ^ Beltman IH, De Kok LJ, Kuiper PJC, Van Hasselt PR (1980). «Состав жирной кислоты и содержание хлорофилла в эпифитных лишанках и возможное отношение к их чувствительности к загрязнению воздуха». Оикос . 35 (3): 321–326. Bibcode : 1980oikos..35..321b . doi : 10.2307/3544647 . JSTOR 3544647 .
- ^ Эммерих Р., Гиз I, Ланге О.Л., Прокш П. (1993). «Токсичность и анти -жизнь соединений лишайников против полифагозного травоядного насекомых Сподоптера Littoralis ». Фитохимия . 33 (6): 1389–1394. Bibcode : 1993pchem..33.1389e . doi : 10.1016/0031-9422 (93) 85097-b .
- ^ Чартер RE (1950). «Флекс как возрастная мачта недавних моренс». Журнал ледника и ледниковой геологии . 1 : 152–161.
- ^ Curry, RR (1969) «Голоценовая климатическая и ледниковая история Центральной Сьерра -Невада, Калифорния», стр. 1–47, Специальная статья Геологического общества Америки, 123, Sa Schumm и WC Bradley, eds.
- ^ Sowers, JM, Noller, JS, и Lettis, WR (Eds.) (1997). Знакомства и землетрясения: обзор четвертичной геохронологии и ее применение к палеосеизму . Комиссия по ядерному регулированию США, Nureg/CR 5562.
- ^ Innes, JL (1985). «Лишайнометрия». Прогресс в физической географии . 9 (2): 187. Bibcode : 1985prpg .... 9..187i . doi : 10.1177/030913338500900202 . S2CID 220949784 .
- ^ Каппителли, Франческа; Сорлини, Клаудия (2008). «Микроорганизмы атакуют синтетические полимеры в предметах, представляющих наше культурное наследие» . Прикладная и экологическая микробиология . 74 (3): 564–569. Bibcode : 2008apenm..74..564c . doi : 10.1128/aem.01768-07 . PMC 2227722 . PMID 18065627 .
- ^ Гадд, Джеффри Майкл (март 2010 г.). «Металлы, минералы и микробы: геомикробиология и биоремедиация» . Микробиология . 156 (Pt 3): 609–643. doi : 10.1099/mic.0.037143-0 . PMID 20019082 .
- ^ Джонсон, Кристофер Дж; Беннет, Джеймс П; Биро, Стивен М; Дуке-Веласкес, Хуан Камило; Родригес, Синтия М.; Бессен, Ричард А; Рок, Тони Е (май 2011 г.). Барц, Джейсон С (ред.). «Разложение прионного белка, ассоциированного с заболеванием, сериновой протеазой из лишайников» . Plos один . 6 (5): E19836. BIBCODE : 2011PLOSO ... 619836J . doi : 10.1371/journal.pone.0019836 . PMC 3092769 . PMID 21589935 .
- ^ Родригес, Синтия М; Беннет, Джеймс П; Джонсон, Кристофер Дж (1 января 2012 г.). "Лишайны: неожиданные антиприонные агенты?" Полем Прион 6 (1): 11–16. doi : 10.4161/pri.6.1.17414 . PMC 3338958 . PMID 22453171 .
- ^ Беннет, Джеймс П; Родриугес, Синтия М; Джонсон, Кристофер (июль 2012 г.). «Прионная деградация белка лишайками рода Cladonia» . Лихенолог . 44 (4): 523–531. doi : 10.1017/s0024282912000102 . S2CID 85819801 .
- ^ «Современный травяной | лакмус» . Botanical.com . Получено 24 октября 2021 года .
- ^ Кассельман, Карен Ли; Дин, Дженни (1999). Дикий цвет: [Полное руководство по изготовлению и использованию натуральных красителей] . Нью-Йорк: Watson-Guptill Publications. ISBN 978-0-8230-5727-6 .
- ^ Мюллер, К. (2001). «Фармацевтически соответствующие метаболиты из лишайников». Прикладная микробиология и биотехнология . 56 (1–2): 9–10. doi : 10.1007/s002530100684 . PMID 11499952 . S2CID 11958438 .
- ^ Jump up to: а беременный Morton, E.; Winters, J. and Smith, L. (2010). «Анализ антисептических и антибиотических свойств разноходно обработанных мхов и лишайников», архивировав 20 августа 2017 года на машине Wayback . Биологическая станция Мичиганского университета
- ^ Бустинза, Ф. (1952). «Антибактериальные вещества из лишайников». Экономическая ботаника . 6 (4): 402–406. doi : 10.1007/bf02984888 . S2CID 39335883 .
- ^ "ThemodelrailReader.com" . Архивировано с оригинала 15 октября 2014 года . Получено 10 октября 2014 года .
- ^ Таким образом, объясняется раввином Енохом Зунелом Беном Джозефом , в своем комментарии Этц Йосеф («Дерево Иосифа»), на Sefer Midrash Rabbah , vol. 2, Нью -Йорк 1987, SV Ruth Rabba 6: 3
- ^ Зохар Амар и Ярон Серри, Земля Израиля и Сирии, как описано Аль-Тамими , Рамат-Ган 2004, с. 56, 108–109 ISBN 965-226-252-8 (иврит)
- ^ Jump up to: а беременный в дюймовый Honegger R. (2000). «Саймон Швендер (1829–1919) и двойная гипотеза в лишайниках». Бриолог . 103 (2): 307–313. Doi : 10.1639/0007-2745 (2000) 103 [0307: ssatdh] 2.0.co ; ISSN 0007-2745 . JSTOR 3244159 . S2CID 84580224 .
- ^ Treub, Мелхиор (1873) Расследования о природе лишайника. Диссертация Университета Лейдена.
- ^ Уэйкфорд, Том (2002). Связывание жизни: от Hornworts до гиппопов, как скромный микроб способствовал эволюции . Джон Уайли и сыновья, включенные. ISBN 978-0-470-24400-5 Полем OCLC 922973303 .
- ^ Юн, Эд (21 июля 2016 года), «Как парень из парка трейлера Монтаны отменил 150 лет биологии» , Атлантика , архивировав с оригинала 23 июля 2017 года , извлеченные 23 июля 2017 года .