Лактоилглутатионлиаза
| лактоилглутатионлиаза | |||
|---|---|---|---|
 Ленточная диаграмма человеческой глиоксалазы I с ее каталитическими ионами цинка, показанными в виде двух фиолетовых сфер. Ингибитор S-гексилглутатион показан в виде модели, заполняющей пространство ; зеленые, красные, синие и желтые сферы соответствуют углерода , кислорода , азота и серы атомам соответственно. | |||
| Идентификаторы | |||
| Номер ЕС. | 4.4.1.5 | ||
| Номер CAS. | 9033-12-9 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| ЭксПАСи | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
Фермент лактоилглутатионлиаза (EC 4.4.1.5, также известный как глиоксалаза I ) катализирует изомеризацию , полутиоацеталевых аддуктов, которые образуются в результате спонтанной реакции между глутатионильной группой и альдегидами такими как метилглиоксаль . [1] [2]
- ( R )- S -лактоилглутатион = глутатион + 2-оксопропаналь
Глиоксалаза I получила свое название от катализа первой стадии глиоксалазной системы , критической двухступенчатой системы детоксикации метилглиоксаля . Метилглиоксаль вырабатывается естественным путем как побочный продукт нормальной биохимии, но он очень токсичен из-за химических реакций с белками , нуклеиновыми кислотами и другими клеточными компонентами. Второй этап детоксикации, на котором ( R ) -S -лактоилглутатион расщепляется на глутатион и D-лактат, осуществляется II гидролазой глиоксалаза . Необычно то, что эти реакции, осуществляемые глиоксалазной системой, не окисляют глутатион, который обычно действует как окислительно-восстановительный кофермент . Хотя альдозоредуктаза также может детоксицировать метилглиоксаль, глиоксалазная система более эффективна и, по-видимому, является наиболее важным из этих путей. Глиоксалаза I является привлекательной мишенью для разработки лекарств для лечения инфекций, вызванных некоторыми паразитическими простейшими, и рака . несколько ингибиторов Было идентифицировано глиоксалазы I, таких как S-(N-гидрокси-N-метилкарбамоил)глутатион.
Глиоксалаза I классифицируется как углерод-серная лиаза , хотя, строго говоря, фермент не образует и не разрушает связь углерод-сера. Скорее, фермент сдвигает два атома водорода с одного атома углерода метилглиоксаля на соседний атом углерода. По сути, реакция представляет собой внутримолекулярную окислительно-восстановительную реакцию; один углерод окисляется, а другой восстанавливается. Механизм происходит за счет вычитания, а затем добавления протонов , образуя промежуточный эндиолат, а не за счет переноса гидридов . Что необычно для металлопротеина , этот фермент проявляет активность с несколькими различными металлами. Глиоксалаза I необычна еще и тем, что она стереоспецифична во второй половине своего механизма, но не в первой половине. Структурно фермент представляет собой димер с замененным доменом у многих видов, хотя у дрожжей две субъединицы слились в мономер посредством дупликации гена .
Номенклатура
[ редактировать ]Систематическое название этого класса ферментов: ( R ) -S -лактоилглутатионметилглиоксаль-лиаза (изомеризующаяся; образующая глутатион); другие имена включают:
- метилглиоксалаза,
- альдокетомутаза,
- кетон-альдегидмутаза и
- ( R ) -S- лактоилглутатионметилглиоксаль-лиаза (изомеризация).
В некоторых случаях глутатионильный фрагмент может быть обеспечен трипанотионом , аналогом глутатиона у паразитических простейших, таких как трипаносомы . Человеческий ген этого фермента называется GLO1 .
Ген
[ редактировать ]| ГЛО1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | GLO1 , GLOD1, GLYI, HEL-S-74, глиоксалаза I | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 138750 ; МГИ : 95742 ; Гомологен : 4880 ; Генные карты : GLO1 ; ОМА : GLO1 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Лактоилглутатионлиаза у человека кодируется GLO1 геном . [7] [8] [9]
Структура
[ редактировать ]несколько структур Расшифровано глиоксалазы I. Были опубликованы четыре структуры человеческой формы с PDB кодами доступа 1BH5 , 1FRO , 1QIN и 1QIP . пять структур формы Escherichia coli Опубликованы с кодами доступа 1FA5 , 1FA6 , 1FA7 , 1FA8 и 1F9Z . одна структура специфичной для трипанотиона версии Leishmania major Наконец, была решена - 2C21 . Во всех этих случаях четвертичная структура биологической единицы представляет собой димер с замененным доменом, в котором активный центр и 8-нитевого бета-листа вторичная структура образуются из обеих субъединиц. Однако у дрожжей , таких как Saccharomyces cerevisiae , две субъединицы слились в один мономер двойного размера посредством дупликации генов . Каждая половина структурного димера представляет собой сэндвич из 3-4 альфа-спиралей по обе стороны 8-нитевого антипараллельного бета-листа; интерфейс димера в основном состоит из встречи лицом к лицу двух бета-листов. [1]
Третичная и четвертичная структуры глиоксалазы I аналогичны структурам некоторых других типов белков. Например, глиоксалаза I напоминает несколько белков, которые позволяют бактериям противостоять антибиотикам, таким как фосфомицин , блеомицин и митомицин . Аналогичным образом, несвязанные ферменты метилмалонил-КоА эпимераза , 3-деметилубихинон-9, 3-О-метилтрансфераза и многочисленные диоксигеназы, такие как бифенил-2,3-диол-1,2-диоксигеназа , катехол-2,3-диоксигеназа , 3,4-дигидроксифенилацетат 2,3-диоксигеназа и 4-гидроксифенилпируватдиоксигеназа по структуре напоминают глиоксалазу I. [1] Наконец, многие белки с неизвестной или неопределенной функцией также напоминают глиоксалазу I, например At5g48480 из растения Arabidopsis thaliana .
Активный сайт состоит из четырех основных регионов.
Функция
[ редактировать ]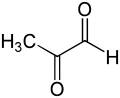
Основная физиологическая функция глиоксалазы I — детоксикация метилглиоксаля , реактивного 2-оксоальдегида , который является цитостатическим при низких концентрациях. [10] и цитотоксичны в миллимолярных концентрациях. [11] Метилглиоксаль — побочный продукт нормальной биохимии, канцероген и мутаген. [12] и может химически повредить некоторые компоненты клетки, такие как белки и нуклеиновые кислоты. [11] [13] Метилглиоксаль образуется спонтанно из дигидроксиацетонфосфата ферментативно с помощью триозофосфатизомеразы и метилглиоксальсинтазы, а также при катаболизме треонина . [14]
Чтобы свести к минимуму количество токсичного метилглиоксаля и других реакционноспособных 2-оксоальдегидов, была разработана система глиоксалазы . Метилглиоксаль спонтанно реагирует с восстановленным глутатионом (или его эквивалентом трипанотионом ), [15] ) с образованием гемитиоацеталя. Глиоксалазная система превращает такие соединения в D- лактат и восстанавливает глутатион. [14] При этом превращении два карбонильных атома углерода 2-оксоальдегида окисляются и восстанавливаются соответственно, причем альдегид окисляется до карбоновой кислоты, а ацетальная группа восстанавливается до спирта. Система глиоксалазы развилась очень рано в истории жизни и встречается почти повсеместно во всех формах жизни.
Глиоаксалазная система состоит из двух ферментов: глиоксалазы I и глиоксалазы II . Первый фермент, описанный здесь, перестраивает гемитиоацеталь, образующийся естественным путем при атаке глутатиона на метилглиоксаль, в продукт. Глиоксалаза II гидролизует продукт с образованием глутатиона и образованием D- лактата . Таким образом, глутатион необычно действует как кофермент и требуется только в каталитических (т. е. очень малых) количествах; обычно вместо этого глутатион действует как окислительно-восстановительная пара в окислительно-восстановительных реакциях.
Также предполагается, что система глиоксалазы играет роль в регуляции роста клеток. [16] и в сборке микротрубочек . [17]
Характеристики
[ редактировать ]Глиоксалаза I требует для катализа связанных ионов металлов. [18] Человеческий фермент [19] и его аналоги в дрожжах ( Saccharomyces cerevisiae ) [20] и Pseudomonas putida [21] используйте двухвалентный цинк , Zn 2+ . Напротив, прокариотические версии часто используют ион никеля . Глиоксалаза, которую я обнаружил у эукариотических трипаносомных паразитов, таких как Leishmania major и Trypanosoma cruzi, также может использовать никель для своей активности. [15] возможно, это отражает приобретение гена GLO1 путем горизонтального переноса генов . [22]
Свойством глиоксалазы I является отсутствие специфичности к каталитическому иону металла. Большинство ферментов связывают один конкретный тип металла, и их каталитическая активность зависит от связывания этого металла. Например, оксидоредуктазы часто используют ион определенного металла, такого как железо , марганец или медь , и не смогут функционировать, если их предпочтительный ион металла будет заменен, из-за различий в окислительно-восстановительном потенциале ; железа таким образом, супероксиддисмутаза не может функционировать, если ее каталитическое железо заменено марганцем, и наоборот. Напротив, хотя человеческая глиоксалаза I предпочитает использовать двухвалентный цинк, она способна функционировать со многими другими двухвалентными металлами, включая магний , марганец , кобальт , никель и даже кальций .; [23] однако фермент неактивен по отношению к катиону железа. [24] Точно так же, хотя прокариотическая глиоксалаза I предпочитает никель, она способна функционировать с кобальтом, марганцем и кадмием ; однако фермент инертен по отношению к связанному цинку из-за изменения геометрии координации с октаэдрической на тригонально-бипирамидальную . [15] Структурные и вычислительные исследования показали, что металл связывает два карбонильных атома кислорода метилглиоксаля в двух его координационных сайтах, стабилизируя промежуточный эндиолат-анион.
Еще одним необычным свойством глиоксалазы I является ее непостоянная стереоспецифичность. Первый этап механизма его реакции (отрыв протона от C 1 и последующее протонирование O 2 ) не является стереоспецифичным и работает одинаково хорошо независимо от исходной хиральности по C 1 в гемитиоацетальном субстрате. Образующийся промежуточный эндиолат является ахиральным, но вторая стадия механизма реакции (отрыв протона от O 1 и последующее протонирование C 2 ) определенно стереоспецифична, образуя только ( S ) форму D-лактоилглутатиона. Считается, что это происходит из-за того, что два глутамата противоположно связываются с ионом металла; любой из них способен выполнить первый шаг, но только один способен выполнить второй шаг. Причина этой асимметрии еще до конца не определена.
Механизм реакции
[ редактировать ]Молекула метилглиоксаля состоит из двух карбонильных групп, окруженных атомом водорода и метильной группой. В обсуждении ниже эти два карбонильных атома углерода будут обозначаться как C1 и C2 соответственно. Как в гемитиоацеталевом субстрате, так и в продукте (R)-S-лактоилглутатиона глутатионовый фрагмент связан с карбонильной группой C1.
Основной механизм действия глиоксалазы I заключается в следующем. Субстрат гемитиоацеталь образуется, когда молекула глутатиона — вероятно, в его реакционноспособной тиолатной форме — атакует карбонил C1 метилглиоксаля или родственного соединения, делая этот углерод четырехвалентным. Эта реакция происходит в клетке спонтанно, без участия фермента. Затем этот гемитиоацеталь связывается ферментом, который перемещает водород с C1 на C2. Карбонил C2 восстанавливается до формы четырехвалентного спирта путем присоединения двух протонов, тогда как карбонил C1 восстанавливается за счет потери водорода, сохраняя при этом связь с фрагментом глутатиона.
Компьютерное исследование в сочетании с имеющимися экспериментальными данными предполагает следующий механизм атомного разрешения для глиоксалазы I. [25] В активном центре каталитический металл принимает октаэдрическую координационную геометрию и в отсутствие субстрата связывает две воды, два противоположных глутамата , гистидин и еще одну боковую цепь, обычно еще один гистидин или глутаматы . Когда субстрат попадает в активный центр, две воды теряются, и два карбонильных кислорода субстрата связываются непосредственно с ионом металла. Два противоположных глутамата добавляют и вычитают протоны из C1 и C2 и соответствующие им кислороды O1 и O2. Первая половина реакции переносит протон от C1 к O2, тогда как вторая половина переносит протон от O1 к C2. Первая реакция может осуществляться любым из противоположных глутаматов, в зависимости от исходной хиральности C1 в гемитиоацетальном субстрате; однако вторая половина стереоспецифична и осуществляется только одним из противоположных глутаматов.
Примечательно, что первый теоретически подтвержденный механизм действия R -субстрата глиоксалазы. недавно опубликован [26]
Каталитический механизм глиоксалазы был изучен с помощью теории функционала плотности, молекулярно-динамического моделирования и гибридных методов КМ/ММ. Причиной особой специфичности фермента (он принимает оба энантиомера своего хирального субстрата, но превращает их в один и тот же энантиомер продукта) является более высокая основность и гибкость одного из глутаматов активного центра (Glu172). [27] [28] [29]
Перенос протона и гидрида
[ редактировать ]Первоначально считалось, что глиоксалаза I действует путем переноса гидрида , который представляет собой протон, окруженный двумя электронами (H – ). [30] Считалось, что в этом он напоминает классический механизм реакции Канниццаро , при котором атака гидроксилата на альдегид превращает его в анион четырехвалентного спирта; этот анион отдает свои атомы водорода второму альдегиду, образуя карбоновую кислоту и спирт. (Фактически, два идентичных альдегида восстанавливают и окисляют друг друга, оставляя чистую степень окисления одинаковой.)
В глиоксалазе I такой механизм переноса гидрида будет работать следующим образом. Атака глутатиона оставит заряженный O – и альдегидный водород, связанный с C 1 . Если карбонильный кислород C 2 может отсоединить водород от послушной кислой боковой цепи фермента, образуя спирт, то водород C 1 может одновременно со своими электронами переместиться на C 2 (гидридный перенос). В то же время дополнительный электрон кислорода C 1 может реформировать двойную связь карбонила, давая конечный продукт.
Альтернативный (и в конечном итоге правильный) механизм с использованием протона (H + ) передача была выдвинута в 1970-х годах. [31] В этом механизме основная боковая цепь фермента отрывает протон альдегида от C 1 ; в то же время к кислороду C 2 присоединяется протон a , образуя эндиол . Эн означает , образовалась двойная связь что между C 2 и C 1 из электронов, оставшихся в результате отрыва альдегидного протона; Диол относится к тому факту , что из первых двух карбонильных групп образовались два спирта. В этом механизме промежуточное соединение образует продукт путем добавления еще одного протона к C 2 .
Ожидалось, что протоны растворителя будут способствовать образованию продукта из эндиола, интермедиата механизма переноса протона, и когда такой вклад не наблюдался в тритированной воде, 3 H 1 O, механизм переноса гидрида был предпочтительным. Однако альтернативная гипотеза о том, что активный центр фермента был глубоко погребен вдали от воды, не могла быть исключена и в конечном итоге оказалась верной. Первые признаки появились, когда постоянно повышающиеся температуры показали постоянно увеличивающееся включение трития, что согласуется с переносом протона и неожиданно происходит с переносом гидрида. Убедительными доказательствами могут служить исследования эффекта изотопа водорода и дейтерия на субстраты, фторированные по метильной группе и дейтерированные по альдегиду. Фторид является хорошей уходящей группой; механизм переноса гидрида предсказывает меньшее удаление ионов фтора из дейтерированного образца, тогда как механизм переноса протона предсказывает большее . Эксперименты с тремя типами глиоксалазы I (дрожжевая, крысиная и мышиная формы) во всех случаях подтвердили механизм переноса протона. [32] Этот механизм наконец был обнаружен в кристаллических структурах глиоксалазы I.
Клиническое значение
[ редактировать ]Поведение
[ редактировать ]Экспрессия Glo1 коррелирует с различиями в тревожном поведении у мышей [33] [34] а также поведение в тесте подвешивания за хвост , чувствительном к антидепрессантам ; [35] однако направление этих эффектов не всегда было последовательным, что вызывает скептицизм. [36] Различия в экспрессии Glo1 у мышей, по-видимому, вызваны вариантом числа копий , который распространен среди инбредных линий мышей. [37] Было высказано предположение, что поведенческие эффекты обусловлены активностью его основного субстрата метилглиоксаля на ГАМК А. рецепторах Glo1 [38] Было показано, что низкомолекулярный ингибитор глиоксалазы I обладает анксиолитическими свойствами, что указывает на еще одно возможное показание для применения ингибиторов глиоксалазы I. [38]
Как мишень для наркотиков
[ редактировать ]Глиоксалаза I является мишенью для разработки фармацевтических препаратов против бактерий, простейших (особенно Trypanosoma cruzi и Leishmania ) и рака человека. [39] Было разработано множество ингибиторов, большинство из которых имеют глутатионовый фрагмент. К семейству ингибиторов, наиболее прочно связывающихся с человеческим ферментом, относятся производные S- ( N -арил- N -гидроксикарбамоил)глутатиона, особенно производное п -бромфенила, константа диссоциации которого составляет 14 нМ. [40] Ближайшим аналогом переходного состояния считают S- ( N -гидрокси- N - p -йодфенилкарбамоил)глутатион; кристаллическая структура этого соединения, связанного с человеческим ферментом, была решена с разрешением 2 Å (код доступа PDB 1QIN ). [41]
Эксперименты показывают, что метилглиоксаль преимущественно токсичен для пролиферирующих клеток, например, раковых. [42]
Недавние исследования показывают, что экспрессия GLO1 повышается в различных злокачественных опухолях человека, включая метастатическую меланому. [43] [44]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Торнэлли П.Дж. (декабрь 2003 г.). «Глиоксалаза I - структура, функция и решающая роль в ферментативной защите от гликирования». Труды Биохимического общества . 31 (Часть 6): 1343–1348. дои : 10.1042/BST0311343 . ПМИД 14641060 .
- ^ Фаррера Д.О., Галлиган Дж.Дж. (октябрь 2022 г.). «Семейство генов глиоксалазы человека в здоровье и болезнях» . Химические исследования в токсикологии . 35 (10): 1766–1776. doi : 10.1021/acs.chemrestox.2c00182 . ПМЦ 10013676 . ПМИД 36048613 . S2CID 251978989 .
- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000124767 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: Ensembl, выпуск 89: ENSMUSG00000024026 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Ранганатан С., Уолш Э.С., Годвин А.К., Тью К.Д. (март 1993 г.). «Клонирование и характеристика глиоксалазы-I толстой кишки человека» . Журнал биологической химии . 268 (8): 5661–7. дои : 10.1016/S0021-9258(18)53370-6 . ПМИД 8449929 .
- ^ Ким Н.С., Умезава Ю., Омура С., Като С. (май 1993 г.). «Человеческая глиоксалаза I. Клонирование кДНК, экспрессия и сходство последовательностей с глиоксалазой I из Pseudomonas putida» . Журнал биологической химии . 268 (15): 11217–21. дои : 10.1016/S0021-9258(18)82113-5 . ПМИД 7684374 .
- ^ «Ген Энтрез: GLO1 глиоксалаза I» .
- ^ Кэмерон А.Д., Олин Б., Риддерстрем М., Маннервик Б., Джонс Т.А. (июнь 1997 г.). «Кристаллическая структура человеческой глиоксалазы I — свидетельства дупликации генов и трехмерной замены доменов» . Журнал ЭМБО . 16 (12): 3386–95. дои : 10.1093/emboj/16.12.3386 . ПМК 1169964 . ПМИД 9218781 .
- ^ Перейти обратно: а б Иноуэ Ю, Кимура А (1995). «Неизвестное название главы». В Р. К. Пуле (ред.). Достижения микробной физиологии (том 37 изд.). Лондон: Академическая пресса. стр. 177–227.
- ^ Нагао М., Фудзита Ю., Вакабаяши К., Нукая Х., Косуге Т., Сугимура Т. (август 1986 г.). «Мутагены в кофе и других напитках» . Перспективы гигиены окружающей среды . 67 : 89–91. дои : 10.1289/ehp.866789 . JSTOR 3430321 . ПМЦ 1474413 . ПМИД 3757962 .
- ^ Фергюсон Г.П., Тотемейер С., Маклин М.Дж., Бут И.Р. (октябрь 1998 г.). «Продуцирование метилглиоксаля бактериями: самоубийство или выживание?». Архив микробиологии . 170 (4): 209–18. Бибкод : 1998ArMic.170..209F . дои : 10.1007/s002030050635 . ПМИД 9732434 . S2CID 21289561 .
Оя Т., Хаттори Н., Мизуно Й., Мията С., Маэда С., Осава Т. и др. (июнь 1999 г.). «Метилглиоксаль-модификация белка. Химическая и иммунохимическая характеристика аддуктов метилглиоксаля-аргинина» . Журнал биологической химии . 274 (26): 18492–502. дои : 10.1074/jbc.274.26.18492 . ПМИД 10373458 .
Торнэлли П.Дж. (1998). «Глутатион-зависимая детоксикация α-оксоальдегидов системой глиоксалазы: участие в механизмах заболевания и антипролиферативная активность ингибиторов глиоксалазы I». хим. Биол. Взаимодействуйте . 112–112: 137–151. Бибкод : 1998CBI...111..137T . дои : 10.1016/s0009-2797(97)00157-9 . ПМИД 9679550 . - ^ Перейти обратно: а б Торнелли П.Дж. (1996). «Фармакология метилглиоксаля: образование, модификация белков и нуклеиновых кислот и ферментативная детоксикация - роль в патогенезе и антипролиферативной химиотерапии». Генерал Фармак . 27 (4): 565–573. дои : 10.1016/0306-3623(95)02054-3 . ПМИД 8853285 .
- ^ Перейти обратно: а б с Ариза А., Викерс Т.Дж., Грейг Н., Армор К.А., Диксон М.Дж., Эгглстон И.М. и др. (февраль 2006 г.). «Специфика трипанотион-зависимой глиоксалазы I Leishmania major: структура и биохимическое сравнение с человеческим ферментом» . Молекулярная микробиология . 59 (4): 1239–48. дои : 10.1111/j.1365-2958.2006.05022.x . ПМИД 16430697 . S2CID 10113958 .
- ^ Сент-Дьердьи А (июль 1965 г.). «Деление клеток и рак». Наука . 149 (3679): 34–7. Бибкод : 1965Sci...149...34S . дои : 10.1126/science.149.3679.34 . ПМИД 14300523 .
- ^ Гиллеспи Э. (январь 1979 г.). «Влияние S-лактоилглутатиона и ингибиторов глиоксалазы I на высвобождение гистамина из лейкоцитов человека». Природа . 277 (5692): 135–7. Бибкод : 1979Natur.277..135G . дои : 10.1038/277135a0 . ПМИД 83539 . S2CID 2153821 .
- ^ Вандер Ягт Д.Л. (1989). «Неизвестное название главы». В Д. Дельфин, Р. Поулсон, О. Аврамович (ред.). Коферменты и кофакторы VIII: Глутатион, часть А. Нью-Йорк: Джон Уайли и сыновья.
- ^ Аронссон AC, Мармстол Э, Маннервик Б (апрель 1978 г.). «Глиоксалаза I, металлофермент цинка млекопитающих и дрожжей». Связь с биохимическими и биофизическими исследованиями . 81 (4): 1235–40. дои : 10.1016/0006-291X(78)91268-8 . ПМИД 352355 .
- ^ Риддерстрем М., Маннервик Б. (март 1996 г.). «Оптимизированная гетерологичная экспрессия человеческого фермента цинка глиоксалазы I» . Биохимический журнал . 314 (Часть 2): 463–7. дои : 10.1042/bj3140463 . ПМК 1217073 . ПМИД 8670058 .
- ^ Сен-Жан А.П., Филлипс К.Р., Крейтон DJ, Stone MJ (июль 1998 г.). «Активные мономерные и димерные формы глиоксалазы I Pseudomonas putida: доказательства трехмерной замены доменов». Биохимия . 37 (29): 10345–53. дои : 10.1021/bi980868q . ПМИД 9671502 .
- ^ Грейг Н., Уилли С., Викерс Т.Дж., Фэрламб А.Х. (декабрь 2006 г.). «Трипанотион-зависимая глиоксалаза I в Trypanosoma cruzi» . Биохимический журнал . 400 (2): 217–23. дои : 10.1042/BJ20060882 . ПМЦ 1652828 . ПМИД 16958620 .
- ^ Селлин С., Эрикссон Л.Е., Аронссон А.С., Маннервик Б. (февраль 1983 г.). «Октаэдрическая координация металлов в активном центре глиоксалазы I, о чем свидетельствуют свойства Co (II)-глиоксалазы I» . Журнал биологической химии . 258 (4): 2091–3. дои : 10.1016/S0021-9258(18)32886-2 . ПМИД 6296126 .
Селлин С, Маннервик Б (1984). «Константы диссоциации металлов для глиоксалазы I, восстановленной Zn 2+ , Ко 2+ , Мн 2+ и мг 2+ 10.1016/S0021-9258 ( Журнал биологической химии . 259 (18): 11426–11429. doi : 18)90878-1 . PMID 6470005 . - ^ Уотила Л., Койвусало М. (апрель 1975 г.). «Очистка и свойства глиоксалазы I из печени овцы» . Европейский журнал биохимии . 52 (3): 493–503. дои : 10.1111/j.1432-1033.1975.tb04019.x . ПМИД 19241 .
- ^ Химо Ф, Siegbahn PE (октябрь 2001 г.). «Каталитический механизм глиоксалазы I: теоретическое исследование». Журнал Американского химического общества . 123 (42): 10280–9. дои : 10.1021/ja010715h . ПМИД 11603978 .
- ^ Джафари С., Райд У., Фуда А.Е., Алави Ф.С., Донг Г., Ирани М. (февраль 2020 г.). «Квантовая механика/молекулярная механика. Исследование механизма реакции глиоксалазы I» . Неорганическая химия . 59 (4): 2594–2603. doi : 10.1021/acs.inorgchem.9b03621 . ПМИД 32011880 . S2CID 211022551 .
- ^ Джафари С., Райд У., Ирани М. (сентябрь 2016 г.). «Каталитический механизм человеческой глиоксалазы, который я изучил с помощью квантово-механических кластерных расчетов» . Журнал молекулярного катализа B: Enzymatic . 131 : 18–30. дои : 10.1016/j.molcatb.2016.05.010 . S2CID 54031819 .
- ^ Джафари С., Каземи Н., Райд У., Ирани М. (май 2018 г.). «Более высокая гибкость Glu-172 объясняет необычную стереоспецифичность глиоксалазы I». Неорганическая химия . 57 (9): 4944–4958. doi : 10.1021/acs.inorgchem.7b03215 . ПМИД 29634252 .
- ^ Джафари С., Райд У., Ирани М. (01.01.2019). «КМ/ММ-исследование стереоспецифического протонного обмена глутатиогидроксиацетона глиоксалазой I» . Результаты по химии . 1 : 100011. doi : 10.1016/j.rechem.2019.100011 .
- ^ Роуз И.А. (июль 1957 г.). «Механизм действия глиоксалазы I». Биохимика и биофизика Acta . 25 (1): 214–5. дои : 10.1016/0006-3002(57)90453-5 . ПМИД 13445752 .
Францен V (1956). «Механизм действия глиоксалазы I». Химические отчеты/Recueil . 89 (4): 1020–1023. дои : 10.1002/cber.19560890427 .
Францен V (1957). «Связь между строением и каталитической активностью тиоламинов в катализе внутримолекулярной реакции Канниццаро». Химические отчеты/Recueil . 90 (4): 623–633. дои : 10.1002/cber.19570900427 . - ^ Холл С.С., Довейко А.М., Джордан Ф. (ноябрь 1976 г.). «Исследование фермента глиоксалазы I. 2. Доказательства ядерного магнитного резонанса механизма переноса эндиола-протона». Журнал Американского химического общества . 98 (23): 7460–1. дои : 10.1021/ja00439a077 . ПМИД 977876 .
Холл С.С., Довейко А.М., Джордан Ф. (1978). «Исследования фермента глиоксалазы I. 4. Общая катализируемая основаниями эндиоловая перегруппировка протон-переноса метил-глиоксальглутатионилгемитиола и фенилглиоксальглутатионилгемитиола ацеталя в S-лактоилглутатион и S-манделоилглутатион с последующим гидролизом - модель ферментной системы глиоксалазы». Журнал Американского химического общества . 100 (18): 5934–5939. дои : 10.1021/ja00486a054 . - ^ Чари Р.В., Козарич Дж.В. (октябрь 1981 г.). «Влияние изотопа дейтерия на разделение продукта фторметилглиоксаля глиоксалазой I. Доказательство механизма переноса протона» . Журнал биологической химии . 256 (19): 9785–8. дои : 10.1016/S0021-9258(19)68690-4 . ПМИД 7024272 .
Козарич Дж.В., Чари Р.В., Ву Дж.К., Лоуренс Т.Л. (1981). «Фторметилглиоксаль - Синтез и глиоксалаза I катализировали разделение продуктов через предполагаемое промежуточное соединение эндиола». Журнал Американского химического общества . 103 (15): 4593–4595. дои : 10.1021/ja00405a057 . - ^ Оватта И., Теннант Р.С., Хелтон Р., Марр Р.А., Сингер О., Редвин Дж.М. и др. (декабрь 2005 г.). «Глиоксалаза 1 и глутатионредуктаза 1 регулируют тревогу у мышей». Природа . 438 (7068): 662–6. Бибкод : 2005Natur.438..662H . дои : 10.1038/nature04250 . ПМИД 16244648 . S2CID 4425579 .
- ^ Кремер С.А., Кесслер М.С., Милфей Д., Бирг И.Н., Банк М., Чибере Л. и др. (апрель 2005 г.). «Идентификация глиоксалазы-I как белкового маркера в мышиной модели крайних проявлений тревожности» . Журнал неврологии . 25 (17): 4375–84. doi : 10.1523/JNEUROSCI.0115-05.2005 . ПМК 6725100 . ПМИД 15858064 .
- ^ Бентон К.С., Миллер Б.Х., Скверер С., Сузуки О., Шульц Л.Е., Кэмерон М.Д. и др. (май 2012 г.). «Оценка генетических маркеров и нейробиохимических аналитов для реакции на флуоксетин с использованием панели инбредных линий мышей» . Психофармакология . 221 (2): 297–315. дои : 10.1007/s00213-011-2574-z . ПМЦ 3337404 . ПМИД 22113448 .
- ^ Торнэлли П.Дж. (май 2006 г.). «Беспокойство по поводу роли глиоксалазы 1 в поведении, связанном с высокой тревожностью». Тенденции молекулярной медицины . 12 (5): 195–9. doi : 10.1016/j.molmed.2006.03.004 . ПМИД 16616641 .
- ^ Уильямс Р., Лим Дж.Э., Харр Б., Винг С, Уолтерс Р., Дистлер М.Г. и др. (2009). «Распространенный и нестабильный вариант числа копий связан с различиями в экспрессии Glo1 и тревожным поведением» . ПЛОС ОДИН . 4 (3): е4649. Бибкод : 2009PLoSO...4.4649W . дои : 10.1371/journal.pone.0004649 . ПМК 2650792 . ПМИД 19266052 .
- ^ Перейти обратно: а б Дистлер М.Г., Плант Л.Д., Соколофф Г., Хоук А.Дж., Анеас И., Вуеншелл Г.Е. и др. (июнь 2012 г.). «Глиоксалаза 1 усиливает тревогу за счет снижения уровня агониста ГАМК-рецептора метилглиоксаля» . Журнал клинических исследований . 122 (6): 2306–15. дои : 10.1172/JCI61319 . ПМК 3366407 . ПМИД 22585572 .
- ^ Торнэлли П.Дж. (1993). «Система глиоксалазы в здоровье и болезни». Молекулярные аспекты медицины . 14 (4): 287–371. дои : 10.1016/0098-2997(93)90002-У . ПМИД 8277832 . S2CID 45479697 .
- ^ Мурти Н.С., Бакерис Т., Каварана М.Дж., Гамильтон Д.С., Лан Ю., Крейтон DJ (июль 1994 г.). «Производные S-(N-арил-N-гидроксикарбамоил)глутатиона являются сильносвязывающими ингибиторами глиоксалазы I и медленными субстратами глиоксалазы II». Журнал медицинской химии . 37 (14): 2161–6. дои : 10.1021/jm00040a007 . ПМИД 8035422 .
- ^ Кэмерон А.Д., Риддерстрем М., Олин Б., Каварана М.Дж., Крейтон DJ, Маннервик Б. (октябрь 1999 г.). «Механизм реакции глиоксалазы I исследован с помощью рентгеноструктурного анализа человеческого фермента в комплексе с аналогом переходного состояния». Биохимия . 38 (41): 13480–90. дои : 10.1021/bi990696c . ПМИД 10521255 .
- ^ Эгюд Л.Г., Сент-Дьёрдьи А. (июнь 1968 г.). «Ранковое действие метилглиоксаля» . Наука . 160 (3832): 1140. Бибкод : 1968Sci...160.1140E . дои : 10.1126/science.160.3832.1140 . ПМИД 5647441 .
Аюб FM, Аллен Р.Э., Торналли П.Дж. (май 1993 г.). «Ингибирование пролиферации клеток лейкемии человека 60 метилглиоксалем in vitro». Исследования лейкемии . 17 (5): 397–401. дои : 10.1016/0145-2126(93)90094-2 . ПМИД 8501967 . - ^ Баир В.Б., Кабельо СМ, Учида К., Баузе А.С., Wondrak GT (апрель 2010 г.). «Сверхэкспрессия GLO1 при злокачественной меланоме человека» . Исследования меланомы . 20 (2): 85–96. дои : 10.1097/CMR.0b013e3283364903 . ПМЦ 2891514 . ПМИД 20093988 .
- ^ Сантариус Т., Бигнелл Г.Р., Гринман К.Д., Видаа С., Чен Л., Махони К.Л. и др. (август 2010 г.). «GLO1-новый амплифицированный ген рака человека» . Гены, хромосомы и рак . 49 (8): 711–25. дои : 10.1002/gcc.20784 . ПМЦ 3398139 . ПМИД 20544845 .
Дальнейшее чтение
[ редактировать ]- Эквалл К., Маннервик Б. (февраль 1973 г.). «Стереохимическая конфигурация лактоильной группы S-лактоилглутатионина, образующейся под действием глиоксалазы I из эритроцитов свиньи и дрожжей». Biochimica et Biophysical Acta (BBA) – Общие предметы . 297 (2): 297–9. дои : 10.1016/0304-4165(73)90076-7 . ПМИД 4574550 .
- Рэкер Э (июнь 1951 г.). «Механизм действия глиоксалазы» . Журнал биологической химии . 190 (2): 685–96. дои : 10.1016/S0021-9258(18)56017-8 . ПМИД 14841219 .
- Аллен Р.Э., Ло Т.В., Торнэлли П.Дж. (апрель 1993 г.). «Упрощенный метод очистки глиоксалазы эритроцитов человека. I. Характеристики, иммуноблоттинг и исследования ингибиторов». Журнал белковой химии . 12 (2): 111–9. дои : 10.1007/BF01026032 . ПМИД 8489699 . S2CID 31587421 .
- Ларсен К., Аронссон А.С., Мармстол Э., Маннервик Б. (1985). «Иммунологическое сравнение глиоксалазы I дрожжей и млекопитающих и количественное определение фермента в тканях человека радиоиммуноанализом». Сравнительная биохимия и физиология. Б. Сравнительная биохимия . 82 (4): 625–38. дои : 10.1016/0305-0491(85)90499-7 . ПМИД 3937656 .
- Вандер Ягт Д.Л., Дауб Э., Крон Дж.А., Хан Л.П. (август 1975 г.). «Влияние pH и тиолов на кинетику дрожжевой глиоксалазы I. Оценка механизма случайного пути». Биохимия . 14 (16): 3669–75. дои : 10.1021/bi00687a024 . ПМИД 240387 .
- Филлипс С.А., Торналли П.Дж. (февраль 1993 г.). «Образование метилглиоксаля из триозофосфатов. Исследование с помощью специфического анализа на метилглиоксаль» . Европейский журнал биохимии . 212 (1): 101–5. дои : 10.1111/j.1432-1033.1993.tb17638.x . ПМИД 8444148 .
Внешние ссылки
[ редактировать ]- Обзор всей структурной информации, доступной в PDB для UniProt : Q04760 (человеческая лактоилглутатионлиаза) на PDBe-KB .
- Обзор всей структурной информации, доступной в PDB для UniProt : Q9CPU0 (мышиная лактоилглутатионлиаза) на PDBe-KB .
галерея PDB |
|---|



