Газообмен
| Часть серии о |
| Механика сплошных сред |
|---|

Газообмен — это физический процесс, при котором газы пассивно движутся за счет диффузии по поверхности. Например, эта поверхность может быть границей раздела воздух/вода водоема, поверхностью газового пузыря в жидкости, газопроницаемой мембраной или биологической мембраной , которая образует границу между организмом и его внеклеточной средой.

Газы постоянно потребляются и производятся в ходе клеточных и метаболических реакций у большинства живых существ, поэтому необходима эффективная система газообмена между, в конечном итоге, внутренней частью клетки (клеток) и внешней средой. Маленькие, особенно одноклеточные организмы, такие как бактерии и простейшие , имеют высокое соотношение площади поверхности к объему . У этих существ газообменной мембраной обычно является клеточная мембрана . Некоторые мелкие многоклеточные организмы, такие как плоские черви , также способны осуществлять достаточный газообмен через кожу или кутикулу , окружающую их тело. Однако у большинства более крупных организмов, которые имеют небольшое соотношение площади поверхности к объему, специализированные структуры с извилистой поверхностью, такие как жабры , легочные альвеолы и губчатые мезофиллы, обеспечивают большую площадь, необходимую для эффективного газообмена. Эти извилистые поверхности иногда могут проникать в тело организма. Так обстоит дело с альвеолами, образующими внутреннюю поверхность млекопитающих . легких , губчатый мезофилл, который находится внутри листьев некоторых видов растений , или жабры тех моллюсков , у которых они есть, которые находятся в мантийной полости.
У аэробных организмов газообмен особенно важен для дыхания , при котором происходит поглощение кислорода ( O
2 ) и выделение углекислого газа ( CO
2 ). И наоборот, у оксигенных фотосинтезирующих организмов, таких как большинство наземных растений , поглощение углекислого газа и выделение как кислорода, так и водяного пара являются основными процессами газообмена, происходящими в течение дня. Другие процессы газообмена важны у менее знакомых организмов: например, обмен углекислого газа, метана и водорода происходит через клеточную мембрану метаногенных архей . При фиксации азота диазотрофными ( бактериями и денитрификации бактериями гетеротрофными и такими как Paracoccus denitrificans различными псевдомонадами ) [1] газообразный азот обменивается с окружающей средой, поглощаясь первой и выделяясь в нее второй, в то время как гигантские трубчатые черви полагаются на бактерии для окисления сероводорода, извлеченного из их глубоководной морской среды, [2] использование растворенного в воде кислорода в качестве акцептора электронов.
Диффузия происходит только с градиентом концентрации . Газы будут перетекать от высокой концентрации к низкой концентрации.Высокая концентрация кислорода в альвеолах и низкая концентрация кислорода в капиллярах приводят к перемещению кислорода в капилляры.Высокая концентрация углекислого газа в капиллярах и низкая концентрация углекислого газа в альвеолах приводят к перемещению углекислого газа в альвеолы.
Физические принципы газообмена
[ редактировать ]Диффузия и площадь поверхности
[ редактировать ]Обмен газов происходит в результате диффузии по градиенту концентрации. Молекулы газа перемещаются из области, в которой они имеют высокую концентрацию, в область, в которой они находятся в низкой концентрации. Диффузия — это пассивный процесс , означающий, что для питания транспорта не требуется никакой энергии, и он подчиняется закону Фика : [ нужна ссылка ]
По отношению к типичной биологической системе, где два отсека («внутри» и «снаружи») разделены мембранным барьером и где газу разрешено самопроизвольно диффундировать вниз по градиенту концентрации: [ нужна ссылка ]
- J — поток, количество газа , диффундирующего с единицы площади мембраны в единицу времени. Обратите внимание, что это уже масштабировано для площади мембраны.
- D — коэффициент диффузии , который будет различаться от газа к газу и от мембраны к мембране в зависимости от размера рассматриваемой молекулы газа и природы самой мембраны (особенно ее вязкости , температуры и гидрофобности ).
- φ – концентрация газа.
- x — положение по толщине мембраны.
- d φ /d x , следовательно, представляет собой градиент концентрации через мембрану. Если два отсека хорошо перемешаны по отдельности, то это упрощается до разницы в концентрации газа между внутренним и внешним отсеками, разделенной на толщину мембраны.
- Отрицательный знак указывает на то, что диффузия всегда происходит в направлении, которое со временем разрушит градиент концентрации, т.е. газ движется от высокой концентрации к низкой концентрации, пока в конечном итоге внутренняя и внешняя камеры не достигнут равновесия .

Газы должны сначала раствориться в жидкости, чтобы диффундировать через мембрану , поэтому все биологические системы газообмена требуют влажной среды. [3] В общем, чем выше градиент концентрации на газообменной поверхности, тем выше скорость диффузии через нее. И наоборот, чем тоньше газообменная поверхность (при той же разнице концентраций), тем быстрее газы будут диффундировать по ней. [4]
В приведенном выше уравнении J — это поток, выраженный на единицу площади, поэтому увеличение площади не повлияет на его значение. Однако увеличение доступной площади поверхности приведет к увеличению количества газа, который может диффундировать за определенный период времени. [4] Это связано с тем, что количество газа, диффундирующего в единицу времени (d q /d t ), является произведением J и площади газообменной поверхности A :

Одноклеточные организмы, такие как бактерии и амебы, не имеют специализированных поверхностей газообмена, поскольку они могут использовать преимущества большой площади поверхности по сравнению с их объемом. Количество газа, которое организм производит (или требует) в данный момент времени, будет примерно пропорционально объему его цитоплазмы . Объем одноклеточного организма очень мал, поэтому он производит (и требует) относительно небольшое количество газа в данный момент времени. По сравнению с этим небольшим объемом площадь поверхности клеточной мембраны очень велика и достаточна для нужд газообмена без дальнейших модификаций. Однако по мере увеличения размера организма площадь его поверхности и объем не масштабируются одинаково. представляющий собой куб со стороной L. Рассмотрим воображаемый организм , Его объем увеличивается с ростом куба ( L 3 ) его длины, но площадь его внешней поверхности увеличивается только с увеличением площади ( L 2 ) его длины. Это означает, что внешняя поверхность быстро становится неадекватной для быстро растущих потребностей в газообмене большего объема цитоплазмы. Кроме того, толщина поверхности, которую должны пересечь газы (d x в законе Фика), также может быть больше у более крупных организмов: в случае одноклеточного организма типичная клеточная мембрана имеет толщину всего 10 нм; [5] но у более крупных организмов, таких как круглые черви (Nematoda), эквивалентная обменная поверхность - кутикула - значительно толще и составляет 0,5 мкм. [6]
Взаимодействие с кровеносными системами
[ редактировать ]
Поэтому у многоклеточных организмов специализированные органы дыхания, такие как жабры или легкие, часто используются для обеспечения дополнительной площади поверхности для необходимой скорости газообмена с внешней средой. Однако расстояния между газообменником и более глубокими тканями часто слишком велики для диффузии и удовлетворения потребностей этих тканей в газах. Поэтому газообменники часто соединяют с газораспределительными системами кровообращения , которые равномерно транспортируют газы ко всем тканям организма независимо от их расстояния от газообменника. [7]
Некоторые многоклеточные организмы, такие как плоские черви (Platyhelminthes), относительно большие, но очень тонкие, что позволяет их внешней поверхности тела действовать как поверхность газообмена без необходимости в специализированном органе газообмена. Таким образом, у плоских червей отсутствуют жабры и легкие, а также система кровообращения. Другие многоклеточные организмы, такие как губки (Porifera), имеют большую площадь поверхности, поскольку они очень пористые и/или разветвленные. Губкам не требуется система кровообращения или специализированные органы газообмена, поскольку их стратегия питания предполагает одностороннюю прокачку воды через их пористые тела с помощью жгутиковых клеток воротника . Таким образом, каждая клетка тела губки подвергается постоянному потоку свежей насыщенной кислородом воды. Поэтому они могут полагаться на диффузию через клеточные мембраны для осуществления газообмена, необходимого для дыхания. [8]
У организмов, у которых системы кровообращения связаны со специализированными газообменными поверхностями, для взаимодействия между ними используются самые разнообразные системы.
В противоточной системе воздух (или, чаще, вода, содержащая растворенный воздух) втягивается в направлении, противоположном потоку крови в газообменнике. Такая противоточная система поддерживает крутой градиент концентрации по длине газообменной поверхности (см. нижнюю диаграмму на рис. 2). Такая ситуация наблюдается в жабрах рыб и многих других водных существ . [9] Газосодержащая вода окружающей среды тянется через газообменную поверхность в одном направлении, а поток крови в жаберных капиллярах под ней течет в противоположном направлении. [9] [10] [11] Хотя теоретически это обеспечивает почти полный перенос дыхательного газа с одной стороны теплообменника на другую, у рыб в кровь обычно переходит менее 80% кислорода в воде, протекающей через жабры. [9]
Альтернативными вариантами являются системы перекрестного тока, встречающиеся у птиц. [12] [13] и тупиковые системы заполненных воздухом мешков, обнаруженные в легких млекопитающих. [14] [15] В системе с прямоточным потоком кровь и газ (или жидкость, содержащая газ) движутся через газообменник в одном направлении. Это означает, что величина градиента варьируется по длине газообменной поверхности, и обмен в конечном итоге прекратится при достижении равновесия (см. верхнюю диаграмму на рис. 2). [9] Применение прямоточных систем газообмена в природе не известно.
Млекопитающие
[ редактировать ]Газообменник у млекопитающих интернализуется и образует легкие, как и у большинства крупных наземных животных. [ нужна ссылка ] Газообмен происходит в микроскопических тупиковых заполненных воздухом мешочках, называемых альвеолами , где очень тонкая мембрана (называемая гематовоздушным барьером ) отделяет кровь в альвеолярных капиллярах (в стенках альвеол) от альвеолярного воздуха в мешочках. .


Обменная мембрана
[ редактировать ]Мембрана, через которую происходит газообмен в альвеолах (т.е. гематовоздушный барьер), чрезвычайно тонка (толщина у человека в среднем 2,2 мкм). [14] Он состоит из альвеолярных эпителиальных клеток , их базальных мембран и эндотелиальных клеток легочных капилляров (рис. 4). [14] [16] Большая площадь поверхности мембраны возникает в результате складывания мембраны примерно на 300 миллионов альвеол диаметром примерно 75–300 мкм каждая. Это обеспечивает чрезвычайно большую площадь поверхности (около 145 м²). 2 ), в котором может происходить газообмен. [14]
Альвеолярный воздух
[ редактировать ]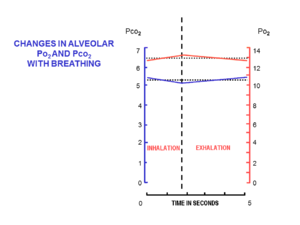

Воздух подается в альвеолы в небольших дозах (так называемый дыхательный объем ), при вдохе ( вдох ) и выдохе ( выдох ) через дыхательные пути , набор относительно узких и умеренно длинных трубок, которые начинаются у носа или рта и заканчиваются. в альвеолах легких, в грудной клетке. Воздух входит и выходит через один и тот же набор трубок, в которых поток движется в одном направлении во время вдоха и в противоположном направлении во время выдоха.
При каждом вдохе в состоянии покоя через нос втекает примерно 500 мл свежего воздуха. Он согревается и увлажняется, проходя через нос и глотку . К моменту достижения трахеи температура вдыхаемого воздуха составляет 37°С и он насыщен водяными парами. По прибытии в альвеолы он разбавляется и тщательно смешивается с примерно 2,5–3,0 л воздуха, оставшимся в альвеолах после последнего выдоха. Этот относительно большой объем воздуха, который полупостоянно присутствует в альвеолах на протяжении всего дыхательного цикла, известен как функциональная остаточная емкость (ФОЕ). [15]
В начале вдоха дыхательные пути наполняются неизмененным альвеолярным воздухом, оставшимся от последнего выдоха. Это объем мертвого пространства , который обычно составляет около 150 мл. [17] Это первый воздух, который снова попадает в альвеолы во время вдоха. Только после того, как воздух мертвого пространства вернулся в альвеолы, остальная часть дыхательного объема (500 мл - 150 мл = 350 мл) поступает в альвеолы. [15] Поступление столь небольшого объема свежего воздуха при каждом вдохе обеспечивает практически полное отсутствие изменений состава ФОЕ в течение дыхательного цикла (рис. 5). [15] Альвеолярное парциальное давление кислорода остается очень близким к 13–14 кПа (100 мм рт. ст.), а парциальное давление углекислого газа минимально варьируется около 5,3 кПа (40 мм рт. ст.) на протяжении всего дыхательного цикла (вдоха и выдоха). [15] Соответствующие парциальные давления кислорода и углекислого газа в окружающем (сухом) воздухе на уровне моря составляют 21 кПа (160 мм рт. ст.) и 0,04 кПа (0,3 мм рт. ст.) соответственно. [15]

Этот альвеолярный воздух, составляющий ФОК, полностью окружает кровь в альвеолярных капиллярах (рис. 6). Газообмен у млекопитающих происходит между альвеолярным воздухом (который существенно отличается от свежего воздуха) и кровью в альвеолярных капиллярах. Газы по обе стороны газообменной мембраны уравновешиваются простой диффузией. Это гарантирует, что парциальное давление кислорода и углекислого газа в крови, покидающей альвеолярные капилляры и в конечном итоге циркулирующей по всему телу, будет таким же, как и в FRC. [15]
Заметное различие между составом альвеолярного воздуха и составом окружающего воздуха может сохраняться за счет того, что функциональная остаточная емкость содержится в тупиковых мешочках, соединенных с наружным воздухом длинными узкими трубками (дыхательные пути: нос , глотка , гортань , трахея , бронхи и их ветви и подветви вплоть до бронхиол ). Эта анатомия, а также тот факт, что легкие не опорожняются, а повторно наполняются при каждом вдохе, обеспечивают млекопитающим «переносимую атмосферу», состав которой значительно отличается от современного окружающего воздуха . [18]
За составом воздуха в ФРК тщательно следят путем измерения парциального давления кислорода и углекислого газа в артериальной крови. Если давление газа отклоняется от нормального, возникают рефлексы, которые изменяют частоту и глубину дыхания таким образом, что нормальное состояние восстанавливается в течение нескольких секунд или минут. [15]
Легочное кровообращение
[ редактировать ]Вся кровь, возвращающаяся из тканей тела в правую часть сердца, течет через альвеолярные капилляры , а затем снова перекачивается по организму. При прохождении через легкие кровь вступает в тесный контакт с альвеолярным воздухом, отделенным от него очень тонкой диффузионной мембраной, толщина которой в среднем составляет всего около 2 мкм. [14] Таким образом, давление газов в крови быстро уравновешивается с давлением в альвеолах , гарантируя, что артериальная кровь, циркулирующая во всех тканях по всему телу, будет иметь напряжение кислорода 13–14 кПа (100 мм рт. ст.), а напряжение углекислого газа — 13–14 кПа (100 мм рт. ст.). 5,3 кПа (40 мм рт. ст.). Эти артериальные парциальные давления кислорода и углекислого газа контролируются гомеостатически . Повышение артериального давления и, в меньшей степени, падение артериального , рефлекторно вызовет более глубокое и учащенное дыхание, пока напряжение газов крови не вернется к норме. Обратное происходит, когда напряжение углекислого газа падает или, опять же, в меньшей степени, повышается напряжение кислорода: частота и глубина дыхания уменьшаются до тех пор, пока не восстановится нормальное состояние газов крови.


Поскольку кровь, поступающая в альвеолярные капилляры, имеет в среднем 6 кПа (45 мм рт. ст.), а давление в альвеолярном воздухе составляет 13 кПа (100 мм рт. ст.), произойдет чистая диффузия кислорода в капиллярную кровь, изменяя состав 3 литров альвеолярного воздуха. немного. Аналогично, поскольку кровь, поступающая в альвеолярные капилляры, имеет также около 6 кПа (45 мм рт. ст.), тогда как альвеолярный воздух составляет 5,3 кПа (40 мм рт. ст.), происходит чистое перемещение углекислого газа из капилляров в альвеолы. Изменения, вызванные этими чистыми потоками отдельных газов в функциональную остаточную емкость и из нее, требуют замены около 15% альвеолярного воздуха окружающим воздухом каждые 5 секунд или около того. Это очень строго контролируется непрерывным мониторингом напряжений газов артериальной крови (которые точно отражают парциальные давления дыхательных газов в альвеолярном воздухе) с помощью тел аорты , сонных артерий , а также газов крови и датчика pH на передней поверхности. продолговатого мозга в головном мозге. В легких также имеются датчики кислорода и углекислого газа, но они в первую очередь определяют диаметры бронхиол и легочных капилляров , а следовательно, отвечают за направление потока воздуха и крови в разные отделы легких.
Только в результате точного поддержания состава 3-х литров альвеолярного воздуха при каждом вдохе часть углекислого газа выбрасывается в атмосферу, а часть кислорода поглощается из наружного воздуха. потеряется больше углекислого газа, чем обычно Если за короткий период гипервентиляции , дыхание будет замедлено или остановлено до тех пор, пока не произойдет альвеолярная недостаточность. вернулось к 5,3 кПа (40 мм рт. ст.). Поэтому, строго говоря, неверно, что основной функцией дыхательной системы является избавление организма от «отходов» углекислого газа. Фактически общая концентрация углекислого газа в артериальной крови составляет около 26 мМ (или 58 мл на 100 мл). [19] по сравнению с концентрацией кислорода в насыщенной артериальной крови около 9 мМ (или 20 мл на 100 мл крови). [15] Эта большая концентрация углекислого газа играет ключевую роль в определении и поддержании pH внеклеточной жидкости . внеклеточной жидкости организма Углекислый газ, который выдыхается при каждом вдохе, вероятно, правильнее было бы рассматривать как побочный продукт углекислого газа и гомеостата рН.
Если эти гомеостаты нарушены, респираторный ацидоз или респираторный алкалоз возникает . В долгосрочной перспективе это может быть компенсировано почечной корректировкой уровня H. + и ОХС 3 − концентрации в плазме; но поскольку это требует времени, синдром гипервентиляции может возникнуть, например, когда возбуждение или тревога заставляют человека дышать быстро и глубоко. [20] тем самым выбрасывая слишком много CO 2 из крови в наружный воздух, вызывая ряд неприятных симптомов, которые возникают в результате чрезмерно высокого pH внеклеточных жидкостей. [21]
Кислород очень плохо растворяется в воде и поэтому переносится кровью в слабом соединении с гемоглобином . Кислород удерживается в гемоглобине четырьмя двухвалентными гемовыми железосодержащими группами на молекулу гемоглобина. Когда все группы гема несут по одной молекуле O 2 каждая, кровь считается «насыщенной» кислородом, и дальнейшее увеличение парциального давления кислорода не приведет к значительному увеличению концентрации кислорода в крови. Большая часть углекислого газа в крови переносится в виде HCO3 . − ионы в плазме. Однако превращение растворенного CO 2 в HCO 3 − (за счет добавления воды) слишком медленна для скорости, с которой кровь циркулирует по тканям с одной стороны и альвеолярным капиллярам с другой. Таким образом, реакция катализируется карбоангидразой , ферментом внутри эритроцитов . [22] Реакция может идти в любом направлении в зависимости от преобладающего парциального давления углекислого газа. Небольшое количество углекислого газа переносится на белковой части молекул гемоглобина в виде карбаминовых групп. Общая концентрация углекислого газа (в виде ионов бикарбоната, растворенного СО 2 и карбаминовых групп) в артериальной крови (т.е. после ее уравновешивания с альвеолярным воздухом) составляет около 26 мМ (или 58 мл/100 мл), [19] по сравнению с концентрацией кислорода в насыщенной артериальной крови около 9 мМ (или 20 мл/100 мл крови). [15]
Другие позвоночные
[ редактировать ]Рыба
[ редактировать ]
Содержание растворенного кислорода в пресной воде составляет примерно 8–10 миллилитров на литр, тогда как в воздухе — 210 миллилитров на литр. [23] Вода в 800 раз плотнее воздуха [24] и в 100 раз более вязкая. [23] Следовательно, скорость диффузии кислорода в воздухе в 10 000 раз выше, чем в воде. [23] Таким образом, использование мешкообразных легких для удаления кислорода из воды было бы недостаточно эффективным для поддержания жизни. [23] Вместо использования легких газообмен происходит через поверхность сильно васкуляризированных жабр . Жабры — специализированные органы, содержащие нити , которые в дальнейшем делятся на пластинки . Ламели содержат капилляры , которые обеспечивают большую площадь поверхности и короткие диффузионные расстояния, поскольку их стенки чрезвычайно тонкие. [25] Жаберные тычинки находятся внутри обменной системы, чтобы фильтровать пищу и поддерживать чистоту жабр.
В жабрах используется система противотока , которая увеличивает эффективность поглощения кислорода (и потери отходящих газов). [9] [10] [11] Кислородная вода всасывается через рот и проходит через жабры в одном направлении, тогда как кровь течет через пластинки в противоположном направлении. Этот противоток поддерживает крутые градиенты концентрации по всей длине каждого капилляра (см. схему в разделе «Взаимодействие с системами кровообращения» выше). Кислород способен непрерывно диффундировать вниз по градиенту в кровь, а углекислый газ — вниз по градиенту в воду. [10] Деоксигенированная вода в конечном итоге выйдет через жаберную крышку (жаберную крышку). Хотя противоточные системы обмена теоретически допускают почти полную передачу дыхательного газа с одной стороны теплообменника на другую, у рыб в кровь обычно переносится менее 80% кислорода в воде, протекающей через жабры. [9]
Земноводные
[ редактировать ]У амфибий есть три основных органа, участвующих в газообмене: легкие, кожа и жабры, которые могут использоваться по отдельности или в различных комбинациях. Относительная важность этих структур различается в зависимости от возраста, окружающей среды и вида амфибии. Кожа земноводных и их личинок сильно васкуляризирована, что приводит к относительно эффективному газообмену, когда кожа влажная. Личинки амфибий, например предметаморфозная головастика стадия лягушек , также имеют наружные жабры . Жабры всасываются в тело во время метаморфоза , после чего легкие берут на себя управление. Легкие обычно проще, чем у других наземных позвоночных , с небольшим количеством внутренних перегородок и более крупными альвеолами; однако жабы, которые проводят больше времени на суше, имеют большую альвеолярную поверхность и более развитые легкие. Чтобы увеличить скорость газообмена за счет диффузии, амфибии поддерживают градиент концентрации на поверхности дыхательных путей, используя процесс, называемый буккальной накачкой. . [26] Нижнее дно рта перемещается «качающимся» образом, что можно наблюдать невооруженным глазом.
Рептилии
[ редактировать ]Все рептилии дышат легкими. У чешуеногих ( ящериц и змей ) вентиляция осуществляется за счет осевой мускулатуры , но эта мускулатура также используется во время движения, поэтому некоторые чешуйчатые полагаются на буккальную накачку для поддержания эффективности газообмена. [27]
Из-за жесткости черепахового и черепахового панциря существенное расширение и сжатие грудной клетки затруднено. Черепахи и черепахи зависят от мышечных слоев, прикрепленных к их панцирям, которые окружают их легкие, заполняя и опорожняя их. [28] Некоторые водные черепахи также могут закачивать воду в хорошо васкуляризированный рот или клоаку для обеспечения газообмена. [29] [30]
Крокодилы имеют структуру, похожую на диафрагму млекопитающих — диафрагму, — но эта мышца помогает создавать однонаправленный поток воздуха через легкие, а не приливный поток: это больше похоже на поток воздуха, наблюдаемый у птиц, чем на тот, который наблюдается у млекопитающих. [31] Во время вдоха диафрагмальная мышца оттягивает печень назад, раздувая легкие в образовавшееся пространство. [32] [33] Воздух поступает в легкие из бронхов при вдохе, но при выдохе воздух выходит из легких в бронх другим путем: такое одностороннее движение газа достигается аэродинамическими клапанами в дыхательных путях. [34] [35]
Птицы
[ редактировать ]

У птиц есть легкие, но нет диафрагмы . они полагаются главным образом на воздушные мешки Для вентиляции . Эти воздушные мешки не играют непосредственной роли в газообмене, но помогают перемещать воздух в одном направлении через поверхности газообмена в легких. При вдохе свежий воздух забирается из трахеи вниз в задние воздухоносные мешки и в парабронхи, ведущие из задних воздухоносных мешков в легкие. Воздух, поступающий в легкие, соединяется с воздухом, уже находящимся в легких, и вытягивается через газообменник в передние воздушные мешки. Во время выдоха задние воздушные мешки нагнетают воздух в те же парабронхи легких, текущий в том же направлении, что и во время вдоха, обеспечивая непрерывный газообмен независимо от дыхательного цикла. Воздух, выходящий из легких при выдохе, присоединяется к воздуху, выбрасываемому из передних воздухоносных мешков (оба состоят из «отработанного воздуха», прошедшего через газообменник), поступающему в трахею для выдыхания (рис. 10). [13] Избирательная бронхоконстрикция в различных точках бронхиальных ветвей обеспечивает то, что воздух не приливы и отливы через бронхи при вдохе и выдохе, как это происходит у млекопитающих, а следует описанным выше путям.
Однонаправленный поток воздуха через парабронхи обменивается дыхательными газами с перекрестным током крови (рис. 9). [12] [13] Парциальное давление O 2 ( ) в парабронхиолах снижается по их длине по мере диффундирования О 2 в кровь. Капилляры, выходящие из обменника вблизи входа воздушного потока, поглощают больше О 2 , чем капилляры, выходящие из около выходного конца парабронхов. Когда содержимое всех капилляров смешается, происходит окончательное смешанной легочной венозной крови выше, чем у выдыхаемого воздуха, но ниже, чем у вдыхаемого. [12] [13]
Растения
[ редактировать ]В газообмене у растений преобладают углекислый газ, кислород и водяной пар . СО
2 является единственным источником углерода для автотрофного роста посредством фотосинтеза , и когда растение активно фотосинтезирует на свету, оно поглощает углекислый газ и теряет водяной пар и кислород. Ночью растения дышат , и газообмен частично меняется на обратный: водяной пар все еще теряется (но в меньшей степени), но теперь поглощается кислород и выделяется углекислый газ. [36]

Газообмен у растений происходит преимущественно через листья. Газообмен между листом и атмосферой происходит одновременно двумя путями: 1) через эпидермальные клетки и кутикулярный воск (обычно называемый « кутикулой »), которые всегда присутствуют на каждой поверхности листа, и 2) через устьица , которые обычно контролируют большую часть обмен. [37] Газы поступают в фотосинтетическую ткань листа путем растворения на влажной поверхности палисадных и губчатых клеток мезофилла . Клетки губчатого мезофилла упакованы рыхло, что позволяет увеличить площадь поверхности и, следовательно, увеличить скорость газообмена. Поглощение углекислого газа обязательно приводит к некоторой потере водяного пара. [38] поскольку обе молекулы входят и выходят через одни и те же устьица, поэтому растения сталкиваются с дилеммой газообмена: получить достаточное количество CO.
2, не теряя слишком много воды. Таким образом, потеря воды из других частей листа сводится к минимуму благодаря восковой кутикуле на эпидермисе листа . Размер стомы регулируется открытием и закрытием двух замыкающих клеток : набухание этих клеток определяет состояние устьичного отверстия, а оно само регулируется водным стрессом. Растения, демонстрирующие толстянковый кислотный обмен, являются засухоустойчивыми ксерофитами и почти весь газообмен осуществляют ночью, так как только ночью эти растения открывают устьица. Открывая устьица только ночью, потери водяного пара, связанные с поглощением углекислого газа, сводятся к минимуму. Однако за это приходится платить медленным ростом: растению приходится запасать углекислый газ в форме яблочной кислоты для использования в течение дня, и оно не может хранить неограниченное количество. [39]

Измерения газообмена являются важными инструментами в науке о растениях: обычно это включает в себя помещение растения (или части растения) в камеру и измерение изменений концентрации углекислого газа и водяного пара с помощью инфракрасного газоанализатора . Если условия окружающей среды ( влажность , CO
2 концентрация, свет и температура ) полностью контролируются, измерения CO
2 поглощение и выделение воды раскрывают важную информацию о CO
2 скорости ассимиляции и транспирации . Межклеточный CO
2 концентрация раскрывает важную информацию о фотосинтетическом состоянии растений. [40] [41] В определенных обстоятельствах можно использовать более простые методы: можно использовать индикатор гидрокарбоната. для контроля потребления CO
2 в растворе, содержащем один лист растения при разной интенсивности света, [42] а выработку кислорода рдестом элодеей можно измерить, просто собрав газ в погруженную пробирку, содержащую небольшой кусочек растения.
Беспозвоночные
[ редактировать ]Механизм газообмена у беспозвоночных зависит от их размера, стратегии питания и среды обитания (водной или наземной).

Губки (Porifera) — сидячие существа, то есть они не могут двигаться самостоятельно и обычно остаются прикрепленными к субстрату . Они получают питательные вещества за счет потока воды через клетки и обмениваются газами путем простой диффузии через клеточные мембраны. Поры, называемые отверстиями, втягивают воду в губку, и вода впоследствии циркулирует по губке клетками, называемыми хоаноцитами , которые имеют волосоподобные структуры , которые перемещают воду через губку. [43]

К книдариям относятся кораллы , морские анемоны , медузы и гидры . Эти животные всегда встречаются в водной среде, от пресной до соленой. У них нет выделенных органов дыхания ; вместо этого каждая клетка их тела может поглощать кислород из окружающей воды и выделять в нее отработанные газы. Одним из ключевых недостатков этой особенности является то, что книдарии могут умереть в условиях застоя в воде воды, поскольку они истощают запасы кислорода . [44] Кораллы часто образуют симбиоз с другими организмами, особенно с фотосинтезирующими динофлагеллятами . В этом симбиозе коралл обеспечивает убежище, а другой организм обеспечивает коралл питательными веществами, включая кислород. [ нужна ссылка ]

( Круглые черви Nematoda), плоские черви (Platyhelminthes) и многие другие мелкие беспозвоночные животные, обитающие в водных или других влажных средах обитания, не имеют выделенной поверхности газообмена или системы кровообращения. Вместо этого они полагаются на диффузию CO .
2 и О
2 прямо через кутикулу. [45] [46] Кутикула — это полупроницаемый внешний слой их тела. [ нужна ссылка ]
Другие водные беспозвоночные, такие как большинство моллюсков (Mollusca) и более крупные ракообразные (Crustacea), такие как омары , имеют жабры, аналогичные рыбьим, которые действуют аналогичным образом.

В отличие от групп беспозвоночных, упомянутых до сих пор, насекомые обычно ведут наземный образ жизни и обменивают газы через влажную поверхность в прямом контакте с атмосферой, а не в контакте с окружающей водой. насекомых Экзоскелет непроницаем для газов, включая водяной пар, поэтому у них более специализированная система газообмена, требующая прямой транспортировки газов к тканям через сложную сеть трубок. Эта дыхательная система отделена от системы кровообращения. Газы входят и выходят из тела через отверстия, называемые дыхалцами , расположенными латерально вдоль грудной клетки и живота . Подобно растениям, насекомые способны контролировать открытие и закрытие дыхалец, но вместо того, чтобы полагаться на тургорное давление , они полагаются на сокращения мышц . [47] Эти сокращения приводят к тому, что брюшко насекомого втягивается и выдыхается. Дыхальца соединены с трубками, называемыми трахеями , которые неоднократно разветвляются и разветвляются в тело насекомого. Эти ветви заканчиваются специализированными клетками трахеолы , которые обеспечивают тонкую влажную поверхность для эффективного газообмена непосредственно с клетками. [48]
Другая основная группа наземных членистоногих , паукообразные ( пауки , скорпионы , клещи и их родственники), обычно осуществляют газообмен с помощью книжных легких . [49]
Краткое описание основных систем газообмена
[ редактировать ]| Площадь поверхности | Диффузионное расстояние | Поддержание градиента концентрации | Органы дыхания | |
|---|---|---|---|---|
| Человек | Всего альвеол [50] = 70–100 м 2 | Альвеола и капилляр (две клетки) | Постоянный кровоток в капиллярах; дыхание | Легкие |
| Рыба | Множество пластинок и нитей на жабре | Обычно одна клетка | Противоток | Жабры |
| Насекомые | Специализированная клетка трахеолы | Одна ячейка | Буккальная накачка | Дыхальца |
| Губки | Остиальные поры | Одна ячейка | Движение воды | Никто |
| Плоские черви | Плоская форма тела | Обычно одна клетка | Противоток | Никто |
| Книдарийцы | Оральные руки | Обычно одна клетка | Движение воды | Никто |
| Рептилии | Множество пластинок и нитей на жабре [ нужны разъяснения ] | Альвеола и капилляр (две клетки) | Противоток | Легкие |
| Земноводные | Множество пластинок и нитей на жабре | Альвеола и капилляр (две клетки) или одна клетка | Противоток | Легкие, кожа и жабры |
| Растения | Высокая плотность устьиц; воздушные пространства внутри листа | Одна ячейка | Постоянный поток воздуха | устьица |
См. также
[ редактировать ]- Дыхательная система - Биологическая система газообмена у животных и растений.
Ссылки
[ редактировать ]- ^ Карлсон, Калифорния; Ингрэм, Дж. Л. (1983). «Сравнение денитрификации Pseudomonas stutzeri , Pseudomonas aeruginosa и Paracoccus denitrificans » . Прил. Окружающая среда. Микробиол . 45 (4): 1247–1253. Бибкод : 1983ApEnM..45.1247C . дои : 10.1128/АЕМ.45.4.1247-1253.1983 . ПМК 242446 . ПМИД 6407395 .
- ^ К.Майкл Хоган. 2011. Сера . Энциклопедия Земли, ред. А. Йоргенсен и К. Дж. Кливленд, Национальный совет по науке и окружающей среде, Вашингтон, округ Колумбия. Архивировано 28 октября 2012 г., в Wayback Machine.
- ^ Пийпер Дж., Дежур П., Хааб П., Ран Х. (1971). «Представления и основные величины в физиологии газообмена». Физиология дыхания . 13 (3): 292–304. дои : 10.1016/0034-5687(71)90034-x . ПМИД 5158848 .
- ^ Перейти обратно: а б Кети С.С. (1951). «Теория и приложения обмена инертных газов в легких и тканях». Фармакологические обзоры . 3 (1): 1–41. ПМИД 14833874 .
- ^ Шнайтер, Р; Брюггер, Б; Сандхофф, Р; Зеллниг, Г; Лебер, А; Лампл, М; Афинштадт, К; Храстник, С; Эдер, С; Даум, Г; Палтауф, Ф; Виланд, штат Форт; Кольвейн, С.Д. (1999). «Тандемный масс-спектрометрический анализ с ионизацией электрораспылением (ESI-MS/MS) молекулярного видового состава липидов субклеточных мембран дрожжей обнаруживает сортировку / ремоделирование на основе ацильной цепи отдельных молекулярных видов на пути к плазматической мембране» . Журнал клеточной биологии . 146 (4): 741–54. дои : 10.1083/jcb.146.4.741 . ПМК 2156145 . ПМИД 10459010 .
- ^ Кокс, Дж.Н. (1 июля 1981 г.). «Кутикула Caenorhabditis elegans : ее выделение и частичная характеристика» . Журнал клеточной биологии . 90 (1): 7–17. дои : 10.1083/jcb.90.1.7 . ПМК 2111847 . ПМИД 7251677 .
- ^ Флегр, Ярослав. «Замороженная эволюция» . Замороженная эволюция . Проверено 21 марта 2017 г.
- ^ «Дыхательная система и прямая диффузия» . Безграничный . Проверено 19 марта 2017 г.
- ^ Перейти обратно: а б с д и ж Кэмпбелл, Нил А. (1990). Биология (Второе изд.). Редвуд-Сити, Калифорния: Benjamin/Cummings Publishing Company, Inc., стр. 836–838. ISBN 978-0-8053-1800-5 .
- ^ Перейти обратно: а б с Хьюз GM (1972). «Морфометрия жабр рыб». Физиология дыхания . 14 (1–2): 1–25. дои : 10.1016/0034-5687(72)90014-x . ПМИД 5042155 .
- ^ Перейти обратно: а б Сторер, Трейси И.; Узергер, Р.Л.; Стеббинс, Роберт С.; Нибаккен, Джеймс В. (1997). Общая зоология (шестое изд.). Нью-Йорк: МакГроу-Хилл. стр. 668–670 . ISBN 978-0-07-061780-3 .
- ^ Перейти обратно: а б с д Скотт, Грэм Р. (2011). «Комментарий: Повышенная производительность: уникальная физиология птиц, летающих на больших высотах» . Журнал экспериментальной биологии . 214 (15): 2455–2462. дои : 10.1242/jeb.052548 . ПМИД 21753038 .
- ^ Перейти обратно: а б с д Ричсон, Г. «BIO 554/754 – Орнитология: дыхание птиц» . Департамент биологических наук Университета Восточного Кентукки . Проверено 23 апреля 2009 г.
- ^ Перейти обратно: а б с д и Уильямс, Питер Л.; Уорик, Роджер; Дайсон, Мэри; Баннистер, Лоуренс Х. (1989). Анатомия Грея (тридцать седьмое изд.). Эдинбург: Черчилль Ливингстон. стр. 1278–1282. ISBN 0443-041776 .
- ^ Перейти обратно: а б с д и ж г час я дж Тортора, Джерард Дж.; Анагностакос, Николас П. (1987). Основы анатомии и физиологии (Пятое изд.). Нью-Йорк: Harper & Row, Издательство. стр. 100-1 570–580 . ISBN 978-0-06-350729-6 .
- ^ «Газообмен у человека» . Проверено 19 марта 2013 г.
- ^ «Том мертвого пространства - Оксфордский справочник» .
- ^ Лавлок, Джеймс (1991). Исцеление Гайи: Практическая медицина для Планеты . Нью-Йорк: Книги Гармонии. стр. 21–34 , 73–88. ISBN 978-0-517-57848-3 .
- ^ Перейти обратно: а б Дим, К.; Лентнер, К. (1970). «Кровь – Неорганические вещества». в: Научные таблицы (Седьмое изд.). Базель, Швейцария: CIBA-GEIGY Ltd. 571.
- ^ Шу, Британская Колумбия; Чанг, ГГ; Ли, штат Нью-Йорк; Ценг, Д.С.; Лин, HY; Лунг, ФРВ (31 октября 2007 г.). «Родительская привязанность, преморбидная личность и психическое здоровье молодых мужчин с синдромом гипервентиляции». Психиатрические исследования . 153 (2): 163–70. doi : 10.1016/j.psychres.2006.05.006 . ПМИД 17659783 . S2CID 3931401 .
- ^ «Электронная медицина — синдром гипервентиляции: статья Эдварда Ньютона, доктора медицины» . Проверено 20 декабря 2007 г.
- ^ Раймонд Х, Свенсон Э (2000). «Распределение и физиологическое значение карбоангидразы в органах газообмена позвоночных». Физиология дыхания . 121 (1): 1–12. дои : 10.1016/s0034-5687(00)00110-9 . ПМИД 10854618 .
- ^ Перейти обратно: а б с д М. Робертс; Майкл Рейсс; Грейс Монгер (2000). Продвинутая биология . Лондон, Великобритания: Нельсон. стр. 164–165.
- ^ Тайсон, продюсер; Престон-Уайт, РА (2013). Погода и климат Южной Африки (второе изд.). Кейптаун, Южная Африка: Издательство Оксфордского университета. п. 14. ISBN 9780195718065 .
- ^ Ньюстед Джеймс Д. (1967). «Тонкое строение дыхательных пластинок костистых жабр». Исследования клеток и тканей . 79 (3): 396–428. дои : 10.1007/bf00335484 . ПМИД 5598734 . S2CID 20771899 .
- ^ Брейнерд, Э.Л. (1999). «Новые взгляды на эволюцию механизмов вентиляции легких у беспозвоночных». Экспериментальная биология онлайн . 4 (2): 1–28. дои : 10.1007/s00898-999-0002-1 . S2CID 35368264 .
- ^ Тейлор, EW; Кэмпбелл, ХА; Лейте, К; Абэ, А.С.; Ван, Т (2007). «Дыхание рептилий». Сравнительная биохимия и физиология. Часть A: Молекулярная и интегративная физиология . 148 : С110–С111. дои : 10.1016/j.cbpa.2007.06.431 .
- ^ Кляйн, Уилфид; Абэ, Аугусто; Андраде, Денис; Перри, Стивен (2003). «Структура послепеченочной перегородки и ее влияние на висцеральную топологию у ящерицы тегу Tupinambis merianae (Teidae: Reptilia)». Журнал морфологии . 258 (2): 151–157. дои : 10.1002/jmor.10136 . ПМИД 14518009 . S2CID 9901649 .
- ^ Оренштейн, Рональд (2001). Черепахи, черепахи и террапины: выжившие в доспехах . Книги Светлячка. ISBN 978-1-55209-605-5 .
- ^ Федер, Мартин Э.; Бурггрен, Уоррен В. (1985). «Кожный газообмен у позвоночных: конструкция, закономерности, контроль и последствия» (PDF) . Биологические обзоры . 60 (1): 1–45. дои : 10.1111/j.1469-185X.1985.tb00416.x . ПМИД 3919777 . S2CID 40158158 .
- ^ Фермер, CG; Сандерс, К. (2010). «Однонаправленный поток воздуха в легких аллигаторов». Наука . 327 (5963): 338–340. Бибкод : 2010Sci...327..338F . дои : 10.1126/science.1180219 . ПМИД 20075253 . S2CID 206522844 .
- ^ Фермер, CG; Перевозчик ДР (2000). «Тазовая аспирация у американского аллигатора ( Alligator Mississippiensis )». Журнал экспериментальной биологии . 203 (11): 1679–1687. дои : 10.1242/jeb.203.11.1679 . ПМИД 10804158 .
- ^ Маннс, СЛ; Оверкович, Т.; Эндрюарта, SJ; Фраппелл, ПБ (2012). «Вспомогательная роль диафрагмальной мышцы в вентиляции легких у эстуарного крокодила Crocodylus porosus » . Журнал экспериментальной биологии . 215 (5): 845–852. дои : 10.1242/jeb.061952 . ПМИД 22323207 .
- ^ Фермер, CG; Сандерс, К. (2010). «Однонаправленный поток воздуха в легких аллигаторов» (PDF) . Наука . 327 (5963): 338–340. Бибкод : 2010Sci...327..338F . дои : 10.1126/science.1180219 . ПМИД 20075253 . S2CID 206522844 . Архивировано из оригинала (PDF) 24 июня 2016 г. Проверено 20 апреля 2017 г.
- ^ Шахнер, ER; Хатчинсон-младший; Фармер, К. (2013). «Легочная анатомия нильского крокодила и эволюция однонаправленного воздушного потока у архозаврий» . ПерДж . 1 : е60. дои : 10.7717/peerj.60 . ПМЦ 3628916 . ПМИД 23638399 .
- ^ Уитмарш Дж., Говинджи (1999). «Глава 2: Основной процесс фотосинтеза» . В: Сингхал Г.С., Ренгер Г., Сопори С.К., Иррганг К.Д., Говинджи (ред.). Концепции фотобиологии: фотосинтез и фотоморфогенез . Бостон: Академическое издательство Kluwer. п. 13. ISBN 978-0-7923-5519-9 .
- ^ Маркес, Диего А.; Стюарт-Уильямс, Хилари; Фаркухар, Грэм Д. (01 марта 2021 г.). «Улучшенная теория для более точного расчета газообмена листьев с учетом малых потоков» . Природные растения . 7 (3): 317–326. doi : 10.1038/s41477-021-00861-w . hdl : 1885/238421 . ISSN 2055-0278 . ПМИД 33649595 . S2CID 232090898 .
- ^ К. Рашке (1976). «Как устьица решают дилемму противоположных приоритетов» . Фил. Пер. Р. Сок. Лонд. Б. 273 (927): 551–560. Бибкод : 1976РСТБ.273..551Р . дои : 10.1098/rstb.1976.0031 .
- ^ Тинг, ИП (1985). «Метаболизм крассуловой кислоты». Ежегодный обзор физиологии растений . 36 (1): 595–622. дои : 10.1146/annurev.pp.36.060185.003115 . HDL : 10150/552219 .
- ^ С. фон Кеммерер; Г. Д. Фаркуар (1981). «Некоторые связи биохимии фотосинтеза и газообмена листьев» . Планта . 153 (4): 376–387. дои : 10.1007/bf00384257 . ПМИД 24276943 . S2CID 22760377 .
- ^ Портативная система газообменной флуоресценции GFS-3000. Руководство по эксплуатации (PDF) , 20 марта 2013 г., заархивировано из оригинала (PDF) 15 декабря 2017 г. , получено 21 октября 2014 г.
- ^ BBC Bitesize - GCSE Biology - Газообмен в растениях
- ^ Андерсон, Д. (2001) Зоология беспозвоночных Oxford University Press
- ^ «Книдарийная дыхательная система» . исследование.com . Проверено 20 марта 2017 г.
- ^ «Дыхательная система нематод» . исследование.com . Проверено 20 марта 2017 г.
- ^ «Дыхательная система платихельминтов» . rspp.weebly.com . Проверено 20 марта 2017 г.
- ^ Лейн, Нью-Джерси; Харрисон, Дж. Б. (1986). «Соединения и цитоскелет в тканях насекомых». Журнал клеточной биологии . 103 (5): А69.
- ^ Клоуден, MJ 2007. Физиологические системы насекомых. Эльзевир/Академическая пресса. стр. 440-442
- ^ Гарвуд, Рассел Дж. и Эджкомб, Грегори Д. (сентябрь 2011 г.). «Ранние наземные животные, эволюция и неопределенность» . Эволюция: образование и информационно-пропагандистская деятельность . 4 (3): 489–501. дои : 10.1007/s12052-011-0357-y .
- ^ Бассет Дж., Кроун С., Саумон Дж. (1987). «Значение активного транспорта ионов в трансальвеолярном поглощении воды: исследование на изолированных легких крысы» . Журнал физиологии . 384 : 311–324. дои : 10.1113/jphysicalol.1987.sp016456 . ПМЦ 1192264 . ПМИД 3656149 .