Фотосистема я

| Фотосистема я | |||
|---|---|---|---|
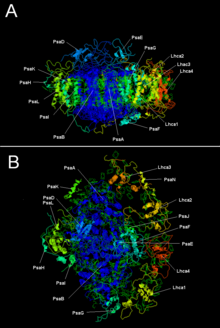 Plant Photogystem I с LHC I | |||
| Идентификаторы | |||
| ЕС №. | 1.97.1.12 | ||
| Базы данных | |||
| Intenz | Intenz View | ||
| Бренда | Бренда вход | ||
| Расширение | Вид Nicezyme | ||
| Кегг | Кегг вход | ||
| Метатический | Метаболический путь | ||
| Напрямую | профиль | ||
| PDB Структуры | RCSB PDB PDBE PDBSUM | ||
| |||

Фотосистема I ( PSI , или пластоцианино -фрередоксинсидоредуктаза является одной из двух фотосистем в фотосинтетических световых реакциях водорослей ) , растений и цианобактерий . Фотосистема я [ 1 ] является интегральным мембранным белковым комплексом , который использует световую энергию для катализации переноса электронов через тилакоидную мембрану от пластоцианина в ферредоксин . В конечном счете, электроны, которые переносятся с помощью фотосистемы I, используются для получения NADPH в средней энергии . [ 2 ] Энергия фотона, поглощенная фотосистемой, я также производит протонную силу , которая используется для генерации АТФ . PSI состоит из более чем 110 кофакторов , что значительно больше, чем фотосистема II . [ 3 ]
История
[ редактировать ]Эта фотосистема известна как PSI, потому что она была обнаружена до Photogystem II, хотя будущие эксперименты показали, что фотосистема II на самом деле является первым ферментом фотосинтетической электронной транспортной цепи. Аспекты PSI были обнаружены в 1950 -х годах, но значение этих открытий еще не было признано в то время. [ 4 ] Луи Дюйзен впервые предложил концепции фотосистем I и II в 1960 году, и в том же году предложение Фэй Бендалла и Роберта Хилла собрали ранее открытия в последовательную теорию последовательных фотосинтетических реакций. [ 4 ] Гипотеза Хилла и Бендалла была позже подтверждена в экспериментах, проведенных в 1961 году группами Duysens и Witt. [ 4 ]
Компоненты и действия
[ редактировать ]Две основные субъединицы PSI, PSAA и PSAB являются тесно связанными белками, участвующими в связывании жизненно важных кофакторов переноса электрона P 700 , ACC, A 0 , A 1 и F x . PSAA и PSAB являются интегральными мембранными белками от 730 до 750 аминокислот , которые содержат 11 трансмембранных сегментов. называемый [4FE-4S] кластер железа-сульфур, F x координируется , четырьмя цистеинами ; Два цистеина предоставляются каждый из PSAA и PSAB. Два цистеина в каждом проксимальны и расположены в петле между девятым и десятым трансмембранным сегментами. лейциновой молнии Похоже, что мотив присутствует [ 5 ] ниже по течению от цистеинов и может способствовать димеризации PSAA/PSAB. Терминальные акцепторы электронов F A и F B , также [4FE-4S] кластеры железа-сульфура, расположены в 9-километровом белке, называемом PSAC, который связывается с ядром PSAA/PSAB вблизи F x . [ 6 ] [ 7 ]
| Белковые субъединицы | Описание |
|---|---|
| Пса | Связанные крупные трансмембранные белки, участвующие в связывании P700, A0, A1 и FX. Часть семейства белков фотосинтетического реакции . |
| Псаб | |
| PSAC | Железный-сулфурский центр; апопротеин для f a и f b |
| Псад | Требуется для сборки, помогает связывать ферредоксин. InterPro : IPR003685 |
| Пса | InterPro : IPR003375 |
| Псаи | Может стабилизировать псал. Стабилизирует светопоглощающее связывание комплекса II. [ 9 ] InterPro : IPR001302 |
| Мочи | InterPro : IPR002615 |
| Псак | InterPro : IPR035982 |
| Он написал | InterPro : IPR036592 |
| PsaM | InterPro : IPR010010 |
| PSAX | InterPro : IPR012986 |
| Цитохром B 6 F комплекс | Растворимый белок |
| F a | Из PSAC; В цепи переноса электронов (и т. Д.) |
| F B. | Из PSAC; В т. Д |
| F x | Из PSAAB; В т. Д |
| Ферредоксин | Электронный носитель в т.е. |
| Пластоцианин | Растворимый белок |
| Липиды | Описание |
| Mgdg II | Моногалактозилдиглицерид липид |
| Pg i | Фосфатидилглицерол фосфолипид |
| PG III | Фосфатидилглицерол фосфолипид |
| PG IV | Фосфатидилглицерол фосфолипид |
| Пигменты | Описание |
| Хлорофилл а | 90 пигментных молекул в антенной системе |
| Хлорофилл а | 5 пигментных молекул в т. Д. |
| Хлорофилл A 0 | Ранний акцептор электронов модифицированного хлорофилла в т.е. |
| Хлорофилл А ' | 1 пигментная молекула в т.е. |
| β-каротизм | 22 каротиноидных молекулы пигментов |
| Коэнзименты и кофакторы | Описание |
| Q K -a | Ранний акцепторный электронный витамин K 1 Филлохинон в ETC |
| Q K -B | Ранний акцепторный электронный витамин K 1 Филлохинон в ETC |
| Фнр | Ferredoxin- nadp + Оксидоредуктаза фермент |
| Что 2+ |
кальция Ион |
| Мг 2+ |
Магний ион |
Фотон
[ редактировать ]Фотоэкссификация пигментных молекул в антеннном комплексе вызывает перенос электронов и энергии. [ 10 ]
Комплекс антенны
[ редактировать ]Комплекс антенны состоит из молекул хлорофилла и каротиноидов, установленных на двух белках. [ 11 ] Эти пигментные молекулы передают резонансную энергию от фотонов, когда они становятся фотоэксценами. Молекулы антенны могут поглощать все длины волны света в видимом спектре . [ 12 ] Число этих пигментных молекул варьируется от организма к организму. Например, Cyanobacterium synechoccus elongatus ( Thermosynechococcus elongatus ) имеет около 100 хлорофиллов и 20 каротиноидов, тогда как хлоропласты шпината имеют около 200 хлорофиллов и 50 каротиноидов. [ 12 ] [ 3 ] В рамках антенного комплекса PSI расположены молекулы хлорофилла, называемые p700 реакционными центрами . Энергия, передаваемая молекулами антенны, направлена на реакционный центр. Там может быть до 120 или всего 25 молекул хлорофилла на P700. [ 13 ]
P700 Реакционный центр
[ редактировать ]Реакционный центр P700 состоит из модифицированного хлорофилла A , который лучше всего поглощает свет на длине волны 700 нм . [ 14 ] P700 получает энергию от молекул антенны и использует энергию от каждого фотона, чтобы поднять электрон до более высокого уровня энергии (P700*). Эти электроны перемещаются парами в процессе окисления/восстановления от P700* до акцепторов электронов, оставляя после себя P700 + Полем Пара P700* - P700 + имеет электрический потенциал около -1,2 вольт . Реакционный центр состоит из двух молекул хлорофилла и поэтому называется димером . [ 11 ] Считается, что димер состоит из одного хлорофилла молекулы и одной молекулы хлорофилла . Однако, если P700 образует комплекс с другими молекулами антенны, он больше не может быть димером. [ 13 ]
Модифицированный хлорофилл A 0 и 1
[ редактировать ]Две модифицированные молекулы хлорофилла являются ранними акцепторами электронов в PSI. Они присутствуют по одному на PSAA/PSAB, образуя две ветви, которые могут взять, чтобы достичь F x . A 0 принимает электроны из P700*, передает его 1 с той же стороны, которая затем передает электрон к хинону с той же стороны. Различные виды, по -видимому, имеют разные предпочтения для любой ветви A/B. [ 15 ]
Phylloquione
[ редактировать ]Филлохинон К , иногда называемый витамином 1 , [ 16 ] это следующий ранний акцептор электронов в PSI. Он окисляет 1 повторно окисляется F x , из которого электрон передается F B и F A. , чтобы получить электрон и, в свою очередь , [ 16 ] [ 17 ] Сокращение F x , по-видимому, является ограничивающим скоростью шаг. [ 15 ]
Железный комплекс
[ редактировать ]Три белковых центров реакции железа и мульчарного обслуживания обнаружены в PSI. Маркированные F x , F A и F B , они служат электронным реле. [ 18 ] F A и F B связаны с белковыми субъединицами комплекса PSI, а F X связаны с комплексом PSI. [ 18 ] Различные эксперименты показали некоторую неравенство между теориями ориентации кофакторов железа и сульфы и порядка эксплуатации. [ 18 ] В одной модели F X передает электрон к F A , который передает его F B, чтобы достичь ферредоксина. [ 15 ]
Ферредоксин
[ редактировать ]Ферредоксин (FD) является растворимым белком, который облегчает восстановление NADP +
в NADPH. [ 19 ] FD движется, чтобы переносить электрон либо в одинокий тилакоид, либо в фермент , который уменьшает NADP +
. [ 19 ] Тилакоидные мембраны имеют один сайт связывания для каждой функции FD. [ 19 ] Основная функция FD заключается в переносе электрона из комплекса железной и-сульфы в фермент ферредоксин- NADP +
редуктаза . [ 19 ]
Ферредоксин - NADP +
редуктаза (FNR)
[ редактировать ] Этот фермент передает электрон от уменьшенного ферредоксина в NADP +
Чтобы завершить сокращение до NADPH. [ 20 ] FNR также может принять электрон из NADPH, связываясь с ним. [ 20 ]
Пластоцианин
[ редактировать ]Пластоцианин является электронным носителем, который переносит электрон от цитохрома B6F на кофактор P700 PSI в его ионизированном состоянии P700 + . [ 10 ] [ 21 ]
YCF4 белковый домен
[ редактировать ]Белковый домен YCF4, обнаруженный на тилакоидной мембране, жизненно важен для фотосистемы I. Этот тилакоидный трансмембранный белок помогает собирать компоненты фотосистемы I. Без нее фотосинтез был бы неэффективным. [ 22 ]
Эволюция
[ редактировать ]Молекулярные данные показывают, что PSI, вероятно, развивался из фотосистем зеленых бактерий серы . Фотосистемы бактерий зеленого серы и бактерий цианобактерий , водорослей и более высоких растений не одинаковы, но есть много аналогичных функций и аналогичных структур. Три основных функции похожи между различными фотосистемами. [ 23 ] Во -первых, окислительно -восстановительный потенциал достаточно отрицательный, чтобы уменьшить ферредоксин. [ 23 ] Далее, центры реакции электронов включают белки железа и мульти. [ 23 ] Наконец, окислительно -восстановительные центры в комплексах обеих фотосистем построены на димере субъединицы белка. [ 23 ] Фотосистема бактерий зеленого серы даже содержит все одинаковые кофакторы цепи переноса электронов в PSI. [ 23 ] Количество и степень сходства между двумя фотосистемами убедительно указывают на то, что PSI и аналогичная фотосистема бактерий зеленого серы развивались из общей цестральной фотосистемы.
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Гольбек Дж. Х. (1987). «Структура, функция и организация комплекса реакционного центра Photosystem I». Biochimica et Biophysica Acta (BBA) - обзоры биоэнергетики . 895 (3): 167–204. doi : 10.1016/s0304-4173 (87) 80002-2 . PMID 3333014 .
- ^ Yamori W, Shikanai T (апрель 2016 г.). «Физиологические функции циклического транспорта электронов вокруг фотосистемы I в поддержании фотосинтеза и роста растений» . Ежегодный обзор биологии растений . 67 : 81–106. doi : 10.1146/annurev-arplant-043015-12002 . PMID 26927905 .
- ^ Jump up to: а беременный Нельсон Н., Йокум CF (2006). «Структура и функция фотосистем I и II». Ежегодный обзор биологии растений . 57 : 521–65. doi : 10.1146/annurev.arplant.57.032905.105350 . PMID 16669773 .
- ^ Jump up to: а беременный в Fromme P, Mathis P (2004). «Раскрытие фотосистемы I реакционного центра: история или сумма многих усилий». Фотосинтез исследования . 80 (1–3): 109–24. Bibcode : 2004phore..80..109f . doi : 10.1023/b: pres.0000030657.88242.e1 . PMID 16328814 . S2CID 13832448 .
- ^ Уэббер А.Н., Малкин Р (май 1990 г.). «Белки, ориентированные на реакцию, ориентированную на фотосистему I, содержат мотивы лейциновой молнии. Предложенная роль в формировании димера». Письма Febs . 264 (1): 1–4. Bibcode : 1990febsl.264 .... 1w . doi : 10.1016/0014-5793 (90) 80749-9 . PMID 2186925 . S2CID 42294700 .
- ^ Джаганнатан Б., Голбек Дж.Х. (апрель 2009 г.). «Разрушение биологической симметрии в мембранных белках: асимметричная ориентация PSAC на ядре Pseudo-C2 симметричной фотосистемы I» . Клеточные и молекулярные науки о жизни . 66 (7): 1257–70. doi : 10.1007/s00018-009-8673-x . PMC 11131447 . PMID 19132290 . S2CID 32418758 .
- ^ Джаганнатан Б., Голбек Дж. Х. (июнь 2009 г.). «Понимание границы связывания между PSAC и гетеродимером PSAA/PSAB в фотосистеме I». Биохимия . 48 (23): 5405–16. doi : 10.1021/bi900243f . PMID 19432395 .
- ^ Saenger W, Jordan P, Krauss N (апрель 2002 г.). «Сборка белковых субъединиц и кофакторов в фотосистеме I». Современное мнение в структурной биологии . 12 (2): 244–54. doi : 10.1016/s0959-440x (02) 00317-2 . PMID 11959504 .
- ^ Плочингер, Магдалена; Тораби, Салар; Рантала, Марджана; Тикканен, Микко; Suorsa, Marjaana; Дженсен, Пул-Эрик; Аро, Ева Мари; Meurer, Jörg (сентябрь 2016 г.). «Низкий молекулярный белок Psai стабилизирует светлополивающий комплекс II сайта стыковки Photosystem I» . Физиология растений . 172 (1): 450–463. doi : 10.1104/pp.16.00647 . PMC 5074619 . PMID 27406169 .
- ^ Jump up to: а беременный Raven PH, Evert RF, Eichhorn SE (2005). «Фотосинтез, свет и жизнь». Биология растений (7 -е изд.). Нью -Йорк: WH Freeman. С. 121–127 . ISBN 978-0-7167-1007-3 .
- ^ Jump up to: а беременный Zeiger E, Taiz L (2006). "Ch. 7: Тема 7.8: фотосистема I" . Физиология растений (4 -е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 0-87893-856-7 . [ Постоянная мертвая ссылка ]
- ^ Jump up to: а беременный «Фотосинтетический процесс» . Архивировано из оригинала на 2009-02-19.
- ^ Jump up to: а беременный Shubin VV, Karapetyan NV, Krasnovsky AA (январь 1986). «Молекулярное расположение пигментного белкового комплекса фотосистемы 1». Фотосинтез исследования . 9 (1–2): 3–12. Bibcode : 1986 Phore ... 9 .... 3S . doi : 10.1007/bf00029726 . PMID 24442279 . S2CID 26158482 .
- ^ Резерфорд AW, Heathcote P (декабрь 1985 г.). «Первичная фотохимия в фотосистеме-I». Фотосинтез исследования . 6 (4): 295–316. Bibcode : 1985 Phore ... 6..295r . doi : 10.1007/bf00054105 . PMID 24442951 . S2CID 21845584 .
- ^ Jump up to: а беременный в Гротжоханн, я; Fromme, P (2013). «Фотосистема я». Энциклопедия биологической химии (второе изд.). Лондон С. 503–507. doi : 10.1016/b978-0-12-378630-2.00287-5 . ISBN 978-0-12-378630-2 .
{{cite book}}: CS1 Maint: местоположение отсутствует издатель ( ссылка ) - ^ Jump up to: а беременный Itoh S, Iwaki M (1989). «Витамин K 1 (филлохинон) восстанавливает оборот центров FES, экспрессируемых эфиром частиц Spinach PSI» . Письма Febs . 243 (1): 47–52. doi : 10.1016/0014-5793 (89) 81215-3 . S2CID 84602152 .
- ^ Palace GP, Franke JE, Warden JT (май 1987). "Является ли филлохинон облигатным электронным носителем в фотосистеме I?" Полем Письма Febs . 215 (1): 58–62. Bibcode : 1987febsl.215 ... 58p . doi : 10.1016/0014-5793 (87) 80113-8 . PMID 3552735 . S2CID 42983611 .
- ^ Jump up to: а беременный в Vassiliev IR, Antonkine ML, Golbeck JH (октябрь 2001 г.). «Железные кластеры в центрах реакции типа I». Biochimica et Biophysica Acta (BBA) - Биоэнергетика . 1507 (1–3): 139–60. doi : 10.1016/s0005-2728 (01) 00197-9 . PMID 11687212 .
- ^ Jump up to: а беременный в дюймовый Forti G, Maria P, Grubas G (1985). «Два участка взаимодействия ферредоксина с тилакоидами» . Письма Febs . 186 (2): 149–152. Bibcode : 1985 Phebsl.186..149f . doi : 10.1016/0014-5793 (85) 80698-0 . S2CID 83495051 .
- ^ Jump up to: а беременный Madoz J, Fernandez Recio J, Witch More C, Fernández VM (ноябрь 1998 г.). NADP +
Редуктаза с помощью электрохимических методов » (PDF) . Биоэлектрохимия и биоэнергетика . 47 (1): 179–183. DOI : 10.1016/S0302-4598 (98) 00175-5 . - ^ Надежда А.Б. (январь 2000 г.). «Электронные передачи среди цитохрома F, пластоцианина и фотосистемы I: кинетика и механизмы». Biochimica et Biophysica Acta (BBA) - Биоэнергетика . 1456 (1): 5–26. doi : 10.1016/s0005-2728 (99) 00101-2 . PMID 10611452 .
- ^ Будро Е., Такахаши Ю., Лемье С., Турмель М., Рочайкс Дж.Д. (октябрь 1997 г.). «Открытые кадры чтения Chloroplast YCF3 и YCF4 Chlamydomonas reinhardtii необходимы для накопления комплекса Photosystem I» . Embo Journal . 16 (20): 6095–104. doi : 10.1093/emboj/16.20.6095 . PMC 1326293 . PMID 9321389 .
- ^ Jump up to: а беременный в дюймовый и Lockau W, Nitschke W (1993). «Фотосистема I и ее бактериальные аналоги». Physiologia plantarum . 88 (2): 372–381. doi : 10.1111/j.1399-3054.1993.tb05512.x .