ДНК-лигаза
| ДНК-лигаза | |||
|---|---|---|---|
 Художественная концепция ДНК-лигазы, восстанавливающей хромосомные повреждения | |||
| Идентификаторы | |||
| Номер ЕС. | 6.5.1.1 | ||
| Номер CAS. | 9015-85-4 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
| лигаза I, ДНК, АТФ-зависимый | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Символ | ЛИГ1 | ||
| ген NCBI | 3978 | ||
| HGNC | 6598 | ||
| МОЙ БОГ | 126391 | ||
| RefSeq | НМ_000234 | ||
| ЮниПрот | P18858 | ||
| Другие данные | |||
| Локус | Хр. 19 [1] | ||
| |||
| лигаза III, ДНК, АТФ-зависимый | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | ЛИГ3 | ||
| ген NCBI | 3980 | ||
| HGNC | 6600 | ||
| МОЙ БОГ | 600940 | ||
| RefSeq | НМ_002311 | ||
| ЮниПрот | P49916 | ||
| Другие данные | |||
| Локус | Хр. 17 q11.2-q12 | ||
| |||
ДНК-лигаза — это тип фермента, который облегчает соединение цепей ДНК , катализируя образование фосфодиэфирной связи . Он играет роль в восстановлении одноцепочечных разрывов дуплексной ДНК в живых организмах, но некоторые формы (например, ДНК-лигаза IV ) могут специфически восстанавливать двухцепочечные разрывы (т.е. разрыв обеих комплементарных цепей ДНК). Однонитевые разрывы восстанавливаются ДНК-лигазой, используя комплементарную цепь двойной спирали в качестве матрицы. [ 1 ] ДНК-лигаза создает последнюю фосфодиэфирную связь для полного восстановления ДНК.
ДНК-лигаза используется как для репарации ДНК , так и для репликации ДНК (см. Лигазы млекопитающих ). Кроме того, ДНК-лигаза широко используется в лабораториях молекулярной биологии для по рекомбинантной ДНК экспериментов (см. «Исследовательские применения »). Очищенная ДНК-лигаза используется при клонировании генов для соединения молекул ДНК с образованием рекомбинантной ДНК .
Ферментативный механизм
[ редактировать ]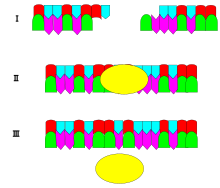
Механизм ДНК-лигазы заключается в образовании двух ковалентных фосфодиэфирных связей между 3'-гидроксильными концами одного нуклеотида («акцептор») и 5'-фосфатным концом другого («донор»). На каждую образовавшуюся фосфодиэфирную связь расходуются две молекулы АТФ. [ нужна ссылка ] AMP необходим для лигазной реакции, которая протекает в четыре этапа:
- Реорганизация места активности, например, разрывы в сегментах ДНК или фрагментах Оказаки и т. д.
- Аденилирование (присоединение АМФ) остатка лизина в активном центре фермента, пирофосфат ; высвобождается
- Перенос АМФ на 5'-фосфат так называемого донора, образование пирофосфатной связи;
- Образование фосфодиэфирной связи между 5'-фосфатом донора и 3'-гидроксилом акцептора. [ 2 ]
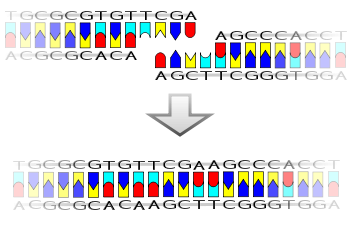
Лигаза также будет работать с тупыми концами , хотя требуются более высокие концентрации фермента и другие условия реакции.
Типы
[ редактировать ]кишечная палочка
[ редактировать ]ДНК-лигаза E. coli кодируется геном lig . ДНК-лигаза в E. coli , как и у большинства прокариот, использует энергию, полученную при расщеплении никотинамидадениндинуклеотида (НАД), для создания фосфодиэфирной связи. [ 3 ] Он не лигирует ДНК с тупыми концами, за исключением условий молекулярной скученности полиэтиленгликолем , и не может эффективно соединять РНК с ДНК. [ нужна ссылка ]
Активность ДНК-лигазы E. coli может быть усилена ДНК-полимеразой в правильных концентрациях. Усиление работает только тогда, когда концентрации ДНК-полимеразы 1 намного ниже, чем концентрации фрагментов ДНК, подлежащих лигированию. Когда концентрации ДНК-полимераз Pol I выше, это оказывает неблагоприятное воздействие на ДНК-лигазу E. coli. [ 4 ]
Т4
[ редактировать ]ДНК-лигаза бактериофага Т4 ( бактериофага, инфицирующего бактерии Escherichia coli ). Лигаза Т4 наиболее часто используется в лабораторных исследованиях. [ 5 ] Он может лигировать как слипшиеся, так и тупые концы ДНК, олигонуклеотиды, а также гибриды РНК и РНК-ДНК, но не одноцепочечные нуклеиновые кислоты. Он также может лигировать ДНК с тупыми концами с гораздо большей эффективностью, чем ДНК-лигаза E. coli . В отличие от ДНК-лигазы E. coli , ДНК-лигаза Т4 не может использовать НАД и абсолютно необходима АТФ в качестве кофактора. Были проведены некоторые инженерные разработки для улучшения in vitro активности ДНК-лигазы Т4 ; один успешный подход, например, протестировал ДНК-лигазу Т4, слитую с несколькими альтернативными ДНК-связывающими белками, и обнаружил, что конструкции с p50 или NF-kB в качестве партнеров слияния были более чем на 160% более активны при лигировании тупых концов для целей клонирования, чем конструкции дикого типа. ДНК-лигаза Т4. [ 6 ] В типичной реакции вставки фрагмента в плазмидный вектор используется от 0,01 (липкие концы) до 1 (тупые концы) единицы лигазы. Оптимальная температура инкубации ДНК-лигазы Т4 составляет 16 °С. [ нужна ссылка ]
бактериофага Т4 лигазы Мутанты обладают повышенной чувствительностью как к УФ-облучению, так и к УФ- излучению. [ 7 ] [ 8 ] и алкилирующий агент метилметансульфонат [ 9 ] что указывает на то, что ДНК-лигаза используется для восстановления вызванных повреждений ДНК, этими агентами.
млекопитающее
[ редактировать ]У млекопитающих существует четыре конкретных типа лигазы.
- ДНК-лигаза 1 : лигирует возникающую ДНК отстающей цепи после того, как рибонуклеаза H удалила праймер РНК из фрагментов Оказаки .
- ДНК-лигаза 3 : образует комплексы с репарации ДНК белком XRCC1, помогая запечатывать ДНК в процессе эксцизионной репарации нуклеотидов и рекомбинантных фрагментов. Из всех известных ДНК-лигаз млекопитающих только лигаза 3 присутствует в митохондриях.
- ДНК-лигаза 4 : комплексы с XRCC4 . Он катализирует заключительный этап пути восстановления негомологичного конца двухцепочечного разрыва ДНК. Он также необходим для рекомбинации V(D)J — процесса, который генерирует разнообразие иммуноглобулинов и рецепторов Т-клеток локусов во время развития иммунной системы .
- ДНК-лигаза 2: артефакт очистки, возникающий в результате протеолитической деградации ДНК-лигазы 3. Первоначально ее считали еще одной ДНК-лигазой, и это является причиной необычной номенклатуры ДНК-лигаз. [ 10 ]
ДНК-лигаза эукариот и некоторых микробов использует аденозинтрифосфат (АТФ), а не НАД. [ 3 ]
Термостабильный
[ редактировать ]Полученный из термофильной бактерии, фермент стабилен и активен при гораздо более высоких температурах, чем обычные ДНК-лигазы. Период его полураспада составляет 48 часов при 65°С и более 1 часа при 95°С. Было показано, что амплигаза ДНК-лигаза активна в течение как минимум 500 термических циклов (94 °C/80 °C) или 16 часов езды на велосипеде. 10 Эта исключительная термостабильность обеспечивает чрезвычайно высокую строгость гибридизации и специфичность лигирования. [ 11 ]
Измерение активности
[ редактировать ]Для измерения активности ДНК-лигазы используются как минимум три различные единицы: [ 12 ]
- Единица Вейсса – количество лигазы, катализирующей обмен 1 нмоль 32 Р из неорганического пирофосфата в АТФ за 20 минут при 37 ° C. Это наиболее часто используемый вариант.
- Единица Модрича-Лемана - используется редко, и одна единица определяется как количество фермента, необходимое для преобразования 100 нмолей d(AT) n в форму, устойчивую к экзонуклеазе-III, за 30 минут в стандартных условиях.
- Многие коммерческие поставщики лигаз используют произвольную единицу измерения, основанную на способности лигазы лигировать сплоченные концы. Эти единицы зачастую более субъективны, чем количественные, и им не хватает точности.
Исследовательские приложения
[ редактировать ]ДНК-лигазы стали незаменимыми инструментами в современных молекулярно-биологических исследованиях для создания рекомбинантных последовательностей ДНК. Например, ДНК-лигазы используются с ферментами рестрикции для встраивания фрагментов ДНК, часто генов , в плазмиды .
Контроль оптимальной температуры является жизненно важным аспектом проведения эффективных экспериментов по рекомбинации, включающих лигирование фрагментов со слипшимися концами. В большинстве экспериментов используется ДНК-лигаза Т4 (выделенная из бактериофага Т4 ), которая наиболее активна при 37 °C. [ 13 ] Однако для оптимальной эффективности лигирования фрагментов со сцепленными концами («липких концов») оптимальная температура фермента должна быть сбалансирована с температурой плавления T m лигируемых липких концов. [ 14 ] гомологичное спаривание липких концов не будет стабильным, поскольку высокая температура разрушает водородные связи . Реакция лигирования наиболее эффективна, когда липкие концы уже стабильно отожжены, и поэтому разрушение отжигаемых концов может привести к низкой эффективности лигирования. Чем короче свес , тем ниже T m .
Поскольку фрагменты ДНК с тупыми концами не имеют сцепленных концов, которые можно было бы отжигать, температура плавления не является фактором, который следует учитывать в нормальном температурном диапазоне реакции лигирования. Ограничивающим фактором при лигировании тупых концов является не активность лигазы, а количество происходящих выравниваний между концами фрагментов ДНК. Таким образом, наиболее эффективной температурой лигирования ДНК с тупыми концами будет температура, при которой может произойти наибольшее количество выравниваний. Большинство перевязок с тупыми концами проводят при температуре 14–25 °C в течение ночи. Отсутствие стабильно отожженных концов также означает, что эффективность лигирования снижается, что требует использования более высокой концентрации лигазы. [ 14 ]
Новое применение ДНК-лигазы можно увидеть в области нанохимии, особенно в ДНК-оригами. Принципы самосборки, основанные на ДНК, оказались полезными для организации наноразмерных объектов, таких как биомолекулы, наномашины, наноэлектронные и фотонные компоненты. Сборка такой наноструктуры требует создания сложной сети молекул ДНК. Хотя самосборка ДНК возможна без какой-либо внешней помощи с использованием различных субстратов, таких как кататоническая поверхность алюминиевой фольги, ДНК-лигаза может оказать ферментативную помощь, необходимую для создания решетчатой структуры ДНК из остатков ДНК. [ 15 ]
История
[ редактировать ]Первая ДНК-лигаза была очищена и охарактеризована в 1967 году лабораториями Геллерта, Лемана, Ричардсона и Гурвица. [ 16 ] Впервые он был очищен и охарактеризован Вайсом и Ричардсоном с использованием шестиэтапного процесса хроматографического фракционирования, начинающегося с удаления остатков клеток и добавления стрептомицина, за которым следовали несколько промывок колонки диэтиламиноэтил (DEAE)-целлюлозой и окончательное фракционирование фосфоцеллюлозы. Конечный экстракт содержал 10% активности, первоначально зафиксированной в среде E. coli ; В ходе процесса было обнаружено, что АТФ и Mg++ необходимы для оптимизации реакции. Обычные коммерчески доступные ДНК-лигазы были первоначально обнаружены в бактериофаге Т4 , E.coli и других бактериях . [ 17 ]
расстройства
[ редактировать ]Генетические дефициты ДНК-лигаз человека связаны с клиническими синдромами, характеризующимися иммунодефицитом, чувствительностью к радиации и аномалиями развития. [ 16 ] Синдром LIG4 (синдром лигазы IV) — редкое заболевание, связанное с мутациями ДНК-лигазы 4 и препятствующее механизмам восстановления разрывов дцДНК. Синдром лигазы IV вызывает у людей иммунодефицит и обычно связан с микроцефалией и гипоплазией костного мозга. [ 18 ] Список распространенных заболеваний, вызванных отсутствием или нарушением работы ДНК-лигазы, следующий.
Пигментная ксеродерма
[ редактировать ]Пигментная ксеродерма , широко известная как XP, представляет собой наследственное заболевание, характеризующееся крайней чувствительностью к ультрафиолетовым (УФ) лучам солнечного света. Это заболевание чаще всего поражает глаза и участки кожи, подвергающиеся воздействию солнца. У некоторых пострадавших также наблюдаются проблемы с нервной системой. [ 19 ]
Атаксия-телеангиэктазия
[ редактировать ]Мутации в гене ATM вызывают атаксию-телеангиэктазию . Ген ATM дает инструкции по созданию белка, который помогает контролировать деление клеток и участвует в восстановлении ДНК. Этот белок играет важную роль в нормальном развитии и деятельности ряда систем организма, включая нервную и иммунную системы. Белок АТМ помогает клеткам распознавать поврежденные или сломанные цепи ДНК и координирует восстановление ДНК, активируя ферменты, которые восстанавливают сломанные цепи. Эффективное восстановление поврежденных цепей ДНК помогает поддерживать стабильность генетической информации клетки. У больных детей обычно возникают трудности при ходьбе, проблемы с балансом и координацией рук, непроизвольные подергивания (хорея), мышечные подергивания (миоклонус) и нарушения функции нервов (нейропатия). Проблемы с передвижением обычно приводят к тому, что в подростковом возрасте людям требуется помощь инвалидной коляски. Люди с этим расстройством также имеют невнятную речь и проблемы с перемещением глаз, чтобы посмотреть из стороны в сторону (глазодвигательная апраксия). [ 20 ]
Анемия Фанкони
[ редактировать ]Анемия Фанкони (ФА) — редкое наследственное заболевание крови, приводящее к недостаточности костного мозга. FA не позволяет костному мозгу производить достаточно новых клеток крови, чтобы организм мог нормально работать. FA также может привести к тому, что костный мозг выработает много дефектных клеток крови. Это может привести к серьезным проблемам со здоровьем, например, лейкемии . [ 21 ]
Синдром Блума
[ редактировать ]Синдром Блума приводит к тому, что кожа становится чувствительной к воздействию солнечных лучей, и обычно появляется пятно покрасневшей кожи в форме бабочки на носу и щеках. Кожная сыпь может также появиться на других участках, которые обычно подвергаются воздействию солнца, например, на тыльной стороне кистей и предплечьях. В сыпи часто появляются небольшие скопления расширенных кровеносных сосудов (телеангиэктазии); телеангиэктазии также могут возникать в глазах. Другие особенности кожи включают участки кожи, которые светлее или темнее окружающих участков (гипопигментация или гиперпигментация соответственно). Эти пятна появляются на участках кожи, не подвергающихся воздействию солнца, и их развитие не связано с высыпаниями.
Как мишень для наркотиков
[ редактировать ]В недавних исследованиях ДНК-лигаза I человека использовалась в компьютерной разработке лекарств для идентификации ингибиторов ДНК-лигазы в качестве возможных терапевтических средств для лечения рака. [ 22 ] Поскольку чрезмерный рост клеток является признаком развития рака, таргетная химиотерапия, нарушающая функционирование ДНК-лигазы, может препятствовать развитию адъювантных форм рака. Кроме того, было показано, что ДНК-лигазы можно разделить на две категории, а именно АТФ- и НАД. + -зависимый. Предыдущие исследования показали, что хотя НАД + -зависимые ДНК-лигазы были обнаружены в спорадических клеточных или вирусных нишах за пределами бактериальной области жизни, нет ни одного случая, когда НАД + -зависимая лигаза присутствует в эукариотическом организме. Присутствие исключительно у неэукариотических организмов, уникальная субстратная специфичность и отличительная доменная структура НАД+-зависимых по сравнению с АТФ-зависимыми ДНК-лигазами человека вместе делают НАД + -зависимые лигазы являются идеальными мишенями для разработки новых антибактериальных препаратов. [ 16 ]
См. также
[ редактировать ]- Конец ДНК
- Отстающая прядь
- репликация ДНК
- Фрагмент Окадзаки
- ДНК-полимераза
- Секвенирование путем лигирования
Ссылки
[ редактировать ]- ^ Паскаль Дж.М., О'Брайен П.Дж., Томкинсон А.Е., Элленбергер Т. (ноябрь 2004 г.). «ДНК-лигаза человека I полностью окружает и частично раскручивает разорванную ДНК». Природа . 432 (7016): 473–8. Бибкод : 2004Natur.432..473P . дои : 10.1038/nature03082 . ПМИД 15565146 . S2CID 3105417 .
- ^ Lehman IR (ноябрь 1974 г.). «ДНК-лигаза: структура, механизм и функции». Наука . 186 (4166): 790–7. Бибкод : 1974Sci...186..790L . дои : 10.1126/science.186.4166.790 . ПМИД 4377758 . S2CID 86549159 .
- ^ Jump up to: а б Фостер Дж. Б., Слончевски Дж. (2010). Микробиология: развивающаяся наука (второе изд.). Нью-Йорк: WW Norton & Company. ISBN 978-0-393-93447-2 .
- ^ Ян Ю, LiCata VJ (февраль 2018 г.). «ДНК-полимеразы Pol I стимулируют соединение концов ДНК с помощью ДНК-лигазы Escherichia coli» . Связь с биохимическими и биофизическими исследованиями . 497 (1): 13–18. дои : 10.1016/j.bbrc.2018.01.165 . ПМИД 29409896 .
- ^ «Лигазы» (PDF) . Руководство по ферментным ресурсам . Корпорация Промега. стр. 8–14.
- ^ Уилсон Р.Х., Мортон С.К., Дейдерик Х., Герт М.Л., Пол Х.А., Гербер И., Патель А., Эллингтон А.Д., Ханике-Смит С.П., Патрик В.М. (июль 2013 г.). «Инженерные ДНК-лигазы с улучшенной активностью in vitro» . Белковая инженерия, проектирование и отбор . 26 (7): 471–8. дои : 10.1093/протеин/gzt024 . ПМИД 23754529 .
- ^ Болди М.В. (1968). «Репарация и рекомбинация в фаге Т4. II. Гены, влияющие на чувствительность к ультрафиолетовому излучению». Симпозиумы Колд-Спринг-Харбор по количественной биологии . 33 : 333–8. дои : 10.1101/sqb.1968.033.01.038 . ПМИД 4891973 .
- ^ Болди М.В. (февраль 1970 г.). «Чувствительность к УФ-излучению некоторых чувствительных к температуре мутантов фага Т4 с ранней функцией». Вирусология . 40 (2): 272–87. дои : 10.1016/0042-6822(70)90403-4 . ПМИД 4909413 .
- ^ Болди М.В., Стром Б., Бернштейн Х. (март 1971 г.). «Репарация дезоксирибонуклеиновой кислоты алкилированного бактериофага Т4 по механизму с участием полинуклеотидлигазы» . Журнал вирусологии . 7 (3): 407–8. doi : 10.1128/JVI.7.3.407-408.1971 . ПМК 356131 . ПМИД 4927528 .
- ^ Томкинсон, Алан Э; Саллмир, Аннахита (5 сентября 2013 г.). «Структура и функция ДНК-лигаз, кодируемых геном LIG3 млекопитающих» . Джин . 531 (2): 150–157. дои : 10.1016/j.gene.2013.08.061 . ПМЦ 3881560 . ПМИД 24013086 .
- ^ «Амплигаза-термостабильная ДНК-лигаза» . www.epibio.com . Архивировано из оригинала 19 июня 2017 г. Проверено 15 мая 2017 г.
- ^ Рассел Д.В., Сэмбрук Дж. (2001). «Глава 1: Плазмиды и их полезность в молекулярном клонировании». Молекулярное клонирование: лабораторное пособие . Том. 1 (3-е изд.). Колд-Спринг-Харбор, Нью-Йорк: Лаборатория Колд-Спринг-Харбор. стр. 1–159. ISBN 978-0-87969-577-4 .
- ^ Бэнейкс Ф., Люкотт Дж. (1993). Введение в методы молекулярного клонирования . Чичестер: Джон Уайли и сыновья. п. 156. ИСБН 978-0-471-18849-0 .
- ^ Jump up to: а б Табор С. (май 2001 г.). «ДНК-лигазы». Современные протоколы молекулярной биологии . Глава 3: Раздел 3.14. дои : 10.1002/0471142727.mb0314s08 . ISBN 978-0-471-14272-0 . ПМИД 18265223 . S2CID 23944826 .
- ^ Бханджадео М.М., Наяк АК, Субудхи У (2017). «Самосборка ДНК с помощью поверхности: безэнзимная стратегия формирования разветвленной решетки ДНК». Связь с биохимическими и биофизическими исследованиями . 485 (2): 492–498. дои : 10.1016/j.bbrc.2017.02.024 . ПМИД 28189681 .
- ^ Jump up to: а б с Шуман С (июнь 2009 г.). «ДНК-лигазы: прогресс и перспективы» . Журнал биологической химии . 284 (26): 17365–9. дои : 10.1074/jbc.R900017200 . ПМК 2719376 . ПМИД 19329793 .
- ^ Вайс Б., Ричардсон CC (апрель 1967 г.). «Ферментативный разрыв и присоединение дезоксирибонуклеиновой кислоты. I. Репарация одноцепочечных разрывов ДНК ферментной системой Escherichia coli, инфицированной бактериофагом Т4» . Труды Национальной академии наук Соединенных Штатов Америки . 57 (4): 1021–8. Бибкод : 1967PNAS...57.1021W . дои : 10.1073/pnas.57.4.1021 . ПМК 224649 . ПМИД 5340583 .
- ^ Альтманн Т., Дженнери А.Р. (октябрь 2016 г.). «Синдром ДНК-лигазы IV; обзор» . Сиротский журнал редких заболеваний . 11 (1): 137. дои : 10.1186/s13023-016-0520-1 . ПМК 5055698 . ПМИД 27717373 .
- ^ «пигментная ксеродерма» . Домашний справочник по генетике . Проверено 15 мая 2017 г.
- ^ «атаксия-телеангиэктазия» . Домашний справочник по генетике . Проверено 15 мая 2017 г.
- ^ «Что такое анемия Фанкони?» . НХЛБИ, НИЗ . Проверено 15 мая 2017 г.
- ^ Томкинсон А.Е., Хоуз Т.Р., Уист Н.Е. (июнь 2013 г.). «ДНК-лигазы как терапевтические мишени» . Трансляционное исследование рака . 2 (3). ПМЦ 3819426 . ПМИД 24224145 .
Внешние ссылки
[ редактировать ]- ДНК-лигаза: молекула месяца PDB
- Общая информация о лигазе Дэвидсон-колледжа
- Протокол лигирования ДНК OpenWetWare
- Обзор всей структурной информации, доступной в PDB для UniProt : P00970 (ДНК-лигаза) в PDBe-KB .
- Обзор всей структурной информации, доступной в PDB для UniProt : P18858 (ДНК-лигаза 1) в PDBe-KB .
- Обзор всей структурной информации, доступной в PDB для UniProt : P49916 (ДНК-лигаза 3) в PDBe-KB .