Гомо
| Гомо Временный диапазон: Piacenzian - присутствует ,
| |
|---|---|

| |
| Notable members of Homo. Clockwise from top left: An approximate reconstruction of a Neanderthal (Homo neanderthalensis) skeleton, a modern human (Homo sapiens) female with a child in India, a reconstructed Homo habilis skull, and a replica skull of Peking Man (subspecies of Homo erectus). | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Primates |
| Suborder: | Haplorhini |
| Infraorder: | Simiiformes |
| Family: | Hominidae |
| Subfamily: | Homininae |
| Tribe: | Hominini |
| Subtribe: | Hominina |
| Genus: | Homo Linnaeus, 1758 |
| Type species | |
| Homo sapiens Linnaeus, 1758
| |
| Species | |
For other species or subspecies suggested, see below. | |
| Synonyms | |
|
Synonyms | |
Homo (от Latin Homō 'Human') - это ( великие род hominidae обезьяны), возникший из рода Australopithecus и охватывает существующие виды Homo sapiens (современные люди ) и ряд вымерших видов (совместно называемые архаичными людьми ), классифицированные как любой наследственные или тесно связанные с современными людьми. К ним относятся Homo erectus и Homo neanderthalensis . Самым старым членом рода является Homo Habilis , с записями чуть более 2 миллионов лет назад. [ А ] Homo вместе с родом Paranthropus , вероятно, наиболее тесно связан с видами Australopithecus africanus в австралопитеке . [ 4 ] Самые близкие живые родственники HOMO имеют род Pan (который включает шимпанзе и бонобо ), а предки Пан и Хомо , по оценкам, расходились около 5,7-11 миллионов лет назад в конце миоцена . [ 5 ]
Homo Erectus появился около 2 миллионов лет назад и распространился по всей Африке (где он называется Homo Ergaster ) и Евразии в нескольких миграциях . Будучи адаптивным и успешным видом, он сохранялся более миллиона лет и постепенно расходился в новые виды примерно к 500 000 лет назад. [b][6]
Anatomically modern humans (Homo sapiens) emerged close to 300,000 to 200,000 years ago,[7] in Africa, and H. neanderthalensis emerged around the same time in Europe and Western Asia. H. sapiens dispersed from Africa in several waves, from possibly as early as 250,000 years ago, and certainly by 130,000 years ago, with the so-called Southern Dispersal beginning about 70–50,000 years ago[8][9][10] leading to the lasting colonisation of Eurasia and Oceania by 50,000 years ago. H. sapiens met and interbred with archaic humans in Africa and in Eurasia.[11][12] Separate archaic (non-sapiens) human species including Neanderthals are thought to have survived until around 40,000 years ago.
Names and taxonomy
[edit]

The Latin noun homō (genitive hominis) means "human being" or "man" in the generic sense of "human being, mankind".[c] The binomial name Homo sapiens was coined by Carl Linnaeus (1758).[d][15] Names for other species of the genus were introduced from the second half of the 19th century (H. neanderthalensis 1864, H. erectus 1892).
The genus Homo has not been strictly defined, even today.[16][17][18] Since the early human fossil record began to slowly emerge from the earth, the boundaries and definitions of the genus have been poorly defined and constantly in flux. Because there was no reason to think it would ever have any additional members, Carl Linnaeus did not even bother to define Homo when he first created it for humans in the 18th century. The discovery of Neanderthal brought the first addition.
The genus Homo was given its taxonomic name to suggest that its member species can be classified as human. And, over the decades of the 20th century, fossil finds of pre-human and early human species from late Miocene and early Pliocene times produced a rich mix for debating classifications. There is continuing debate on delineating Homo from Australopithecus—or, indeed, delineating Homo from Pan. Even so, classifying the fossils of Homo coincides with evidence of: (1) competent human bipedalism in Homo habilis inherited from the earlier Australopithecus of more than four million years ago, as demonstrated by the Laetoli footprints; and (2) human tool culture having begun by 2.5 million years ago to 3 million years ago.[19]
From the late-19th to mid-20th centuries, a number of new taxonomic names, including new generic names, were proposed for early human fossils; most have since been merged with Homo in recognition that Homo erectus was a single species with a large geographic spread of early migrations. Many such names are now regarded as "synonyms" with Homo, including Pithecanthropus,[20] Protanthropus,[21] Sinanthropus,[22] Cyphanthropus,[23] Africanthropus,[24] Telanthropus,[25] Atlanthropus,[26] and Tchadanthropus.[27][28]
Classifying the genus Homo into species and subspecies is subject to incomplete information and remains poorly done. This has led to using common names ("Neanderthal" and "Denisovan"), even in scientific papers, to avoid trinomial names or the ambiguity of classifying groups as incertae sedis (uncertain placement)—for example, H. neanderthalensis vs. H. sapiens neanderthalensis, or H. georgicus vs. H. erectus georgicus.[29] Some recently extinct species in the genus have been discovered only lately and do not as yet have consensus binomial names (see Denisova hominin).[30] Since the beginning of the Holocene, it is likely that Homo sapiens (anatomically modern humans) has been the only extant species of Homo.
John Edward Gray (1825) was an early advocate of classifying taxa by designating tribes and families.[31] Wood and Richmond (2000) proposed that Hominini ("hominins") be designated as a tribe that comprised all species of early humans and pre-humans ancestral to humans back to after the chimpanzee–human last common ancestor, and that Hominina be designated a subtribe of Hominini to include only the genus Homo — that is, not including the earlier upright walking hominins of the Pliocene such as Australopithecus, Orrorin tugenensis, Ardipithecus, or Sahelanthropus.[32] Обозначения Альтернатива Hominina существовала или предлагалась: Australopithecinae (Gregory & Hellman 1939) и Preanthropinae (Cela-Conde & Altaba 2002); [ 33 ] [ 34 ] [ 35 ] А позже, Cela-Conde и Ayala (2003) предположили, что четыре рода Australopithecus , Ardipithecus , Praeantropus и Sahelanthropus были сгруппированы с Homo в гоминини (без пан ). [ 34 ]
Эволюция
[ редактировать ]Australopithecus и появление гомо
[ редактировать ]Несколько видов, в том числе Australopithecus Garhi , Australopithecus sediba , Australopithecus africanus и Australopithecus afarensis , были предложены в качестве предка или сестры линии Homo . [ 36 ] [ 37 ] У этих видов есть морфологические особенности, которые соответствуют им с HOMO , но нет никакого консенсуса, который породил HOMO .
Тем более, что с 2010 -х годов разграничение HOMO в Australopithecus стало более спорным. Традиционно, появление HOMO было принято, чтобы совпадать с первым использованием каменных инструментов ( индустрия Oldowan ) и, следовательно, по определению с началом нижнего палеолита . Но в 2010 году были представлены доказательства, которые, по -видимому, приписывают использование каменных инструментов Australopithecus afarensis около 3,3 миллиона лет назад, примерно за миллион лет до первого появления Homo . [ 38 ] LD 350-1 , ископаемый челюсть, датированный 2,8 млн лет, обнаруженный в 2013 году в Афар, Эфиопия , был описан как сочетание «примитивных признаков, наблюдаемых в раннем австралопитеке с полученной морфологией, наблюдаемым в более позднем гомо . [ 39 ] Некоторые авторы подтолкнут развитие HOMO близко к или даже за 3 млн лет. [ E ] Он обнаруживает поддержку в недавнем филогенетическом исследовании в гомининах, которое, используя морфологическую, молекулярную и радиометрическую информацию, датируется появлением HOMO при 3,3 млн. Лет (4,30 - 2,56 млн. Лет). [ 40 ] Другие выразили сомнение относительно того, должен ли Homo habilis быть включен в HOMO , предлагая происхождение Homo с Homo erectus примерно в 1,9 млн лет. [ 41 ]
Наиболее важным физиологическим развитием между более ранними австралопитецинными видами и HOMO является увеличение эндокраниального объема (ECV) от 460 см. 3 (28 с вн.) В A. garni до 660 см 3 куб . ( 40 3 (46 с In) в H. erectus , 1250 см 3 (76 куб) в H. heidelbergensis и до 1760 см 3 (107 куб) в H. neanderthalensis . Тем не менее, устойчивый рост числа черепа уже наблюдается в автопитецине и не заканчивается после появления HOMO , так что он не служит объективным критерием для определения появления рода. [ 42 ]
Мужчина под рукой
[ редактировать ]Homo Habilis появился около 2,1 млн лет. Уже до 2010 года были предположения, что Х. Хабилис не должен быть помещен не в род HOMO , а скорее в Australopithecus . [ 43 ] [ 44 ] Основная причина включать H. habilis в Homo , его бесспорное использование инструмента, устарела с обнаружением инструмента Australopithecus, по крайней мере, за миллион лет до Х. Хабилиса . [ 38 ] Кроме того, Х. Хабилис долгое время считался предком более грякового гомо -эргастера ( Homo erectus ). В 2007 году было обнаружено, что Х. Хабилис и Х. Эректус сосуществовали в течение значительного времени, предполагая, что H. erectus сразу получена не от Х. Хабилиса , а вместо этого от общего предка. [ 45 ] С публикацией черепа Dmanisi 5 в 2013 году стало менее уверенным, что Asian H. erectus является потомком африканского H. Ergaster , который, в свою очередь, был получен от H. habilis . Вместо этого H. Ergaster и H. erectus , по -видимому, являются вариантами одного и того же вида, которые могли возникнуть в Африке или в Азии [ 46 ] и широко рассеян по всей Евразии (включая Европу , Индонезию , Китай ) на 0,5 млрд. Мю. [ 47 ]
Человек бдительности
[ редактировать ]Homo Erectus Предполагается, что развивался анагенетически у Х. Хабилиса примерно 2 миллиона лет назад. Этот сценарий был укреплен с открытием Homo erectus Georgicus , ранних образцов H. erectus, найденных на Кавказах , которые, по -видимому, демонстрировали переходные черты с H. habilis . Поскольку самые ранние доказательства H. erectus были найдены за пределами Африки, считалось правдоподобным, что H. erectus развивался в Евразии, а затем мигрировал обратно в Африку. Основываясь на окаменелостях из формации Куби , восток от озера Туркана в Кении, Spoor et al. (2007) утверждали, что Х. Хабилис , возможно, выжил за пределами появления H. erectus , так что эволюция H. erectus не была бы анагенетически, а H. erectus существовал бы вместе с H. habilis около полумиллиона лет ( От 1,9 до 1,4 миллиона лет назад ), во время раннего Калабриана . [ 45 ] 31 августа 2023 года исследователи сообщили, что, основываясь на генетических исследованиях, что человека узкое место населения (от возможных 100 000 до 1000 человек) произошло около 930 000 и 813 000 лет назад ... длился около 117 000 лет и приблизил предков человека к вымирание "." [ 48 ] [ 49 ]
Вайс (1984) подсчитал, что было около 44 миллиардов (короткие) членов рода Homo от его происхождения до эволюции H. erectus , около 56 миллиардов человек от H. erectus до неолита и еще 51 миллиард людей с тех пор неолит. Это дает возможность для огромного количества новых мутационных изменений возникнуть во время эволюции человека. [ 50 ]
Отдельный южноафриканский вид homo gautengensis был постулирован как современный с H. erectus в 2010 году. [ 51 ]
Филогения
[ редактировать ]−10 — – −9 — – −8 — – −7 — – −6 — – −5 — – −4 — – −3 — – −2 — – −1 — – 0 — | ( O. Prenes ) ( O. tuenensis ) ( Ar. Ramidus ) |
| ||||||||||||||||||||||||||||
Таксономия гомо в рамках великих обезьян оценивается следующим образом, когда парантропа и гомо появляются в Австралопитеке (показанный здесь, кладистически предоставляющий парантропу , кенийский и гомо ). [ А ] [ f ] [ 6 ] [ 53 ] [ 52 ] [ 4 ] [ 54 ] [ 55 ] [ 56 ] [ 57 ] [ 58 ] [ 59 ] [ 60 ] [ чрезмерные цитаты ] Точная филогения в Австралопитеке все еще очень спорна. Приблизительные даты радиации дочерних клад показаны миллионами лет назад (MYA). [ 61 ] [ 57 ] Сахелантроп и Оррорин , возможно, сестры в Австралопитек , здесь не показаны. Наименование групп иногда запутывается, так как часто предполагаются определенные группировки до выполнения какого -либо кладистического анализа. [ 55 ]
| Hominoidea |
| ||||||||||||||||||||||||
| (20,4 млн лет) |
Кладограмма на основе Dembo et al. (2016): [ 57 ]
| Австралопитеки |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| (7,3 млн лет) |
Некоторые из линий Homo , по -видимому, выживают потомством через интрогрессию в другие линии. Генетические данные указывают на архаичную линию, отделяющую от других человеческих линий 1,5 миллиона лет назад, возможно, H. erectus , возможно, вмешалась в денизованы около 55 000 лет назад. [ 62 ] [ 54 ] [ 63 ] Фосильские данные показывают, что H. erectus SS выжил по крайней мере до 117 000 лет назад, а еще более базальный H. floresiensis выжил до 50 000 лет назад. Похоже, что 1,5 миллиона лет H. erectus , похоже, попала в современные люди через денисованы и, в частности, в папуас и аборигенов австралийцев. [ 54 ] Геномы не-суб-сахарских африканских людей показывают, что, по-видимому, представляют собой многочисленные независимые интрогрессионные события с участием неандертальца, а в некоторых случаях также денизованы около 45 000 лет назад. [ 64 ] [ 63 ] Генетическая структура некоторых африканских групп к югу от Сахары, по-видимому, свидетельствует о интрогрессии от западной Евразийской популяции около 3000 лет назад. [ 58 ] [ 65 ]
Некоторые данные свидетельствуют о том, что Australopithecus sediba может быть перенесен на род Homo или помещен в свой собственный род из -за его положения в отношении Eg H. Habilis и H. floresiensis . [ 56 ] [ 66 ]
Рассеяние
[ редактировать ]Примерно к 1,8 миллионам лет назад H. erectus присутствует как в Восточной Африке ( H. Ergaster ), так и в Западной Азии ( H. Georgicus ). Предки индонезийского H. floresiensis, возможно, покинули Африку еще раньше. [ G ] [ 56 ]
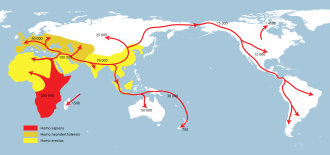
Homo erectus и связанный или полученный архаичный человеческий вид в течение следующих 1,5 миллионов лет распространились по всей Африке и Евразии [ 67 ] [ 68 ] (См.: Недавнее африканское происхождение современных людей ). Европа достигается около 0,5 млн лет Homo Heidelbergensis .
Homo neanderthalensis и H. sapiens развиваются примерно после 300 кья. Homo Naledi присутствует на юге Африки 300 Kya.
H. Sapiens вскоре после его первого появления распространилась по всей Африке и в Западную Азию в нескольких волнах , возможно, еще в 250 Kya, и, конечно, к 130 Kya. В июле 2019 года антропологи сообщили о обнаружении 210 000 -летних остатков H. sapiens и 170 000 -летних остатков H. neanderthalensis в пещере Апидимы , Пелопоннес , Греция , более чем на 150 000 лет старше, чем предыдущие находки H. sapiens в Европе. [ 69 ] [ 70 ] [ 71 ]
Наиболее заметным является южный рассеивание H. sapiens около 60 Kya, что привело к длительному народу Океании и Евразии анатомически современными людьми . [ 11 ] H. sapiens взаимодействует с архаичными людьми как в Африке, так и в Евразии, в Евразии, особенно с неандертальцами и денисованами . [ 72 ] [ 73 ]
Среди существующих групп населения H. sapiens самая глубокая временная подразделение находится в сан -народе Южной Африки, оценивается в почти 130 000 лет, [ 74 ] Или, возможно, более 300 000 лет назад. [ 75 ] Временное разделение между неафриканцами составляет 60 000 лет в случае с австрало-меланзийцами . Отдел европейцев и восточноазиатцев составляет 50 000 лет, с повторными и значительными событиями примесь по всей Евразии во время голоцена .
Архаичный человеческий вид, возможно, выжил до начала голоцена , хотя они в основном вымерли или поглощались расширяющимися популяциями H. sapiens на 40 кя ( неандертальское исчезновение ).
Список линий
[ редактировать ]Статус вида H. rudolfensis , H. Ergaster , H. Georgicus , H. Antecsor , H. Cepranensis , H. Rhodesiensis , H. nanderthalensis , Denisova Hominin и H. floresiensis остаются под дебатами. H. heidelbergensis и H. neanderthalensis тесно связаны друг с другом и считались подвидом H. sapiens .
Исторически была тенденция постулировать новые человеческие виды, основанные всего лишь как отдельный окаменелостей. «Минималистский» подход к таксономии человека признает не более трех видов, H. habilis (2,1–1,5 млн лет, членство в Homo сомнительно), H. erectus (1,8–0,1 млн лет, включая большую часть эпохи рода и большинство архаичных сортов в качестве подвида , [ 76 ] [ 77 ] [ 78 ] в том числе H. heidelbergensis в качестве позднего или переходного разнообразия [ 79 ] [ 80 ] [ 81 ] ) и Homo Sapiens (300 KYA, в том числе H. neanderthalensis и другие разновидности в качестве подвида ). Последовательные определения и методология разграничения видов обычно не согласованы в антропологии или палеонтологии. Действительно, спецификация популяций млекопитающих может обычно переходить в течение нескольких миллионов лет после того, как они начинают генетически расходиться, [ 82 ] [ 83 ] Таким образом, все современные «виды» в роде HOMO потенциально смогли бы скрещиваться в то время, и интрогрессия из -за пределов рода Homo не может быть априори . [ 84 ] Было высказано предположение, что H. naledi , возможно, был гибридом с поздним выжившим австралипитом (воспринимается за Homo Homo , ed.), [ 53 ] Несмотря на то, что эти линии обычно считаются долго вымершими. Как обсуждалось выше, между линиями происходило много интрогрессий с признаками интрогрессии после разделения 1,5 миллиона лет.
| Линии | Временный диапазон ( принадлежать ) |
Среда обитания | Взрослый рост | Взрослый масса | Черновая способность (см 3 ) |
Запись ископаемого | Открытие/ публикация имени |
|---|---|---|---|---|---|---|---|
| H. Handy Членство в Homo неопределенное |
2,100–1,500 [ H ] [ я ] | Танзания | 110–140 см (3 фута 7 дюймов - 4 фута 7 дюймов) | 33–55 кг (73–121 фунт) | 510–660 | Много | 1960 1964 |
| Х. Рудольфенсис Членство в Homo неопределенное |
1,900 | Кения | 700 | 2 сайта | 1972 1986 | ||
| H. Gautengensis также классифицируется как Х. Хабилис |
1,900–600 | ЮАР | 100 см (3 фута 3 дюйма) | 3 человека [ 87 ] [ J ] | 2010 2010 | ||
| H. Блюдо | 1,900–140 [ 88 ] [ k ] [ 89 ] [ L ] | Африка , Евразия | 180 см (5 футов 11 дюймов) | 60 кг (130 фунтов) | 850 (рано) - 1100 (поздно) | Много [ м ] [ n ] | 1891 1892 |
| Х. Эргастер Африканский H. erectus |
1,800–1,300 [ 91 ] | Восточная и южная Африка | 700–850 | Много | 1949 1975 | ||
| H. Предшественник | 1,200–800 | Западная Европа | 175 см (5 футов 9 дюймов) | 90 кг (200 фунтов) | 1,000 | 2 сайта | 1994 1997 |
| H. floresiensis классификация неопределенна |
1,000–50 | Индонезия | 100 см (3 фута 3 дюйма) | 25 кг (55 фунтов) | 400 | 7 человек | 2003 2004 |
| H. Heidelbergensis Ранний H. neanderthalensis |
600–300 [ O ] | Европа, Африка | 180 см (5 футов 11 дюймов) | 90 кг (200 фунтов) | 1,100–1,400 | Много | 1907 1908 |
| H. cepranensis Один ископаемый, возможно, H. heidelbergensis |
в 450 [ 92 ] | Италия | 1,000 | 1 крышка черепа | 1994 2003 | ||
| Х. Налед | 335—236 [ 93 ] | ЮАР | 150 см (4 фута 11 дюймов) | 45 кг (99 фунтов) | 450 | 15 человек | 2013 2015 |
| H. Long | 309–138 [ 94 ] | Северо -восточный Китай | 1,420 [ 95 ] | 1 человек | 1933 2021 | ||
| H. Rhodesiensis Ранний Х. Сапиенс |
в 300 | Замбия | 1,300 | Одинокий или очень немногие | 1921 1921 | ||
| H. Sapiens (анатомически современные люди) |
в 300 -Present [ P ] | Во всем мире | 150–190 см (4 фута 11 дюймов - 6 футов 3 дюйма) | 50–100 кг (110–220 фунтов) | 950–1,800 | (существует) | —— 1758 |
| Denisova hominin | в 285 - c. 51 | Сибирь | 2 сайта | 2000 2010 [ Q ] | |||
| H. neanderthalensis |
240– 40 [ 98 ] [ r ] | Европа, Западная Азия | 170 см (5 футов 7 дюймов) | 55–70 кг (121–154 фунта) (сильно построенный) |
1,200–1,900 | Много | 1829 1864 |
| Несер Рамла Хомо классификация неопределенна |
140–120 | Израиль | Несколько человек | 2021 | |||
| H. tsaichangensis possibly H. erectus or Denisova |
в 100 [ s ] | Тайвань | 1 человек | 2008(?) 2015 | |||
| H. luzonensis |
в 67 [ 101 ] [ 102 ] | Филиппины | 3 человека | 2007 2019 |
Смотрите также
[ редактировать ]- Список окаменелостей эволюции человека (с изображениями)
- Многорегиональное происхождение современных людей
Сноски
[ редактировать ]- ^ Jump up to: а беременный Обычная оценка возраста Х. Хабилиса составляет примерно от 2,1 до 2,3 миллиона лет. [ 1 ] [ 2 ] Предложения по возвращению возраста до 2,8 млн лет были сделаны в 2015 году на основе открытия челюсти . [ 3 ]
- ^ Homo erectus в узком смысле (азиатские виды) вымерли к 140 000 лет назад; H. erectus soloensis , найденный в Java , считается последним известным выживанием H. erectus . Ранее от 50 000 до 40 000 лет назад исследование 2011 года выдвинуло H. e. Солоэндис датируется до 143 000 лет назад по последним, более вероятном до 550 000 лет назад. [ 6 ]
- ^ Слово «человеческое» само по себе происходит от латинского гумана , прилагательное, сформированное на корне гомо , которое, по мнению прото-индоевропейского слова для «Земли», реконструированного как *dhǵem- . [ 13 ]
- ^ В 1959 году Карл Линнеус был обозначен как лектотип для Homo Sapiens , [ 14 ] Это означает, что следуя номенклатурным правилам Homo Sapiens был обоснованно определен как виды животных, к которым принадлежал Линнеус.
- ^ Cela-Conde & Ayala (2003) распознает пять родов в гоминине: Ardipithecus , Australopithecus (включая парантроп ), Homo (включая кенийский ), Praeantropus (включая Orrorin ) и Sahelanthropus . [ 34 ]
- ^ Линия к самым ранним членам HOMO была получена от Australopithecus , рода, который отделился от шимпанзе -Хумана последнего общего предка по последним миоценом или ранним временам плиоцена . [ 52 ]
- ^ В филогенетическом исследовании 2015 года H. floresiensis был помещен с Australopithecus sediba , H. habilis и Dmanisi Man , что повысило вероятность того, что предки H. floresiensis покинули Африку до появления H. erectus , возможно, даже становятся первыми гомининами Сделайте это и развивались дальше в Азии. [ 56 ]
- ^ Подтвержденные окаменелости H. habilis датируются от 2,1 до 1,5 миллионов лет назад. Этот диапазон дат перекрывается с появлением Homo erectus . [ 85 ] [ 86 ]
- ^ Гоминины с чертами «прото-гомо», возможно, проживали еще 2,8 миллиона лет назад, как было предложено ископаемой челюсти, классифицированной как переходная между Австралопитеком и гомо , обнаруженным в 2015 году.
- ^ Вид, предложенный в 2010 году на основе ископаемых остатков трех особей, от 1,9 до 0,6 миллиона лет назад. Те же окаменелости также были классифицированы как Х. Хабилис , Х. Эргастер или Австралопитек другими антропологами.
- ^ H. Erectus , возможно, появился около 2 миллионов лет назад. Окаменелости от 1,8 миллионов лет назад были обнаружены как в Африке, так и в Юго -Восточной Азии, и самые старые окаменелости с узким отрывом (от 1,85 до 1,77 миллиона лет назад были обнаружены в Кавказе, так что неясно, не четко ли H. . Erectus появился в Африке и мигрировал в Евразию, или, наоборот, он развивался в Евразии и мигрировал обратно в Африку.
- ^ Homo erectus soloensis , найденный в Java , считается последним известным выживанием H. erectus . Ранее, датированное от 50 000 до 40 000 лет назад, исследование 2011 года оттолкнуло дату его исчезновения H. e. Soloensis до 143 000 лет назад по последним, более вероятном до 550 000 лет назад. [ 90 ]
- ^ Теперь также включены в H. erectus - Peking Man (ранее Sinantropus Pekinensis ) и Java Man (ранее Pithecanthropus Erectus ).
- ^ H. Эркт в том числе Alert erectus , , Man lantianensis в настоящее время сгруппируется в различные подвиды , erectus , Lantianensis Man erector Pekinensis Palaeojavanicus , палеоджаванику , мужчина эректаж Soloensis , человеческий ярость Tautavelensis , Человек Георгика . Различие от качества потомков, такого как человек, эргастер , человек Floresiensis , предок , человек Heidelbergensis И действительно Sapiens не совсем ясен.
- ^ Тип окаменелости - Mauer 1 , датированное ок. 0,6 миллиона лет назад. Переход от H. heidelbergensis в H. neanderthalensis между 300 и 243 тысячами лет назад является обычным и использует тот факт, что в этот период нет известных ископаемых. Примерами H. heidelbergensis являются окаменелости, найденные в Bilzingsleben (также классифицируется как Homo erectus bilzingslebensis ).
- ^ Возраст H. sapiens уже давно приближается к 200 000 лет, но с 2017 года на этот раз продлил ряд предложений до 300 000 лет. В 2017 году окаменелости, обнаруженные в Джебель Ирхоуд ( Марокко ), предполагают, что Homo Sapiens , возможно, поощрялся еще в 315 000 лет назад. [ 96 ] Генетические данные были представлены в возрасте около 270 000 лет. [ 97 ]
- ^ Временные названия Homo sp. Altai или Homo Sapiens Ssp. Денисова .
- ^ Первые люди с «прото-неандертальскими чертами» жили в Евразии уже с 0,6 до 0,35 миллиона лет назад (классифицируется как H. heidelbergensis , также называемый хроноспецифическими , потому что она представляет хронологическую группу, а не основан на четких морфологических различиях из них. H. erectus или H. neanderthalensis ). В Европе есть ископаемый разрыв между 300 и 243 Kya, а по конвенции окаменелости моложе 243 Kya называются «неандертальцем». [ 99 ]
- ^ моложе 450 KYA, либо между 190–130, либо между 70–10 кА [ 100 ]
Ссылки
[ редактировать ]- ^ Стрингер, CB (1994). «Эволюция ранних людей». В Джонс, с.; Martin, R.; Pilbeam, D. (Eds.). Кембриджская энциклопедия эволюции человека . Кембридж: издательство Кембриджского университета . п. 242
- ^ Schrenk, F.; Куллмер, О.; Bromage, T. (2007). «Глава 9: самые ранние предполагаемые гомо окаменелости». В Хенке, W.; Tattersall, I. (ред.). Справочник по палеоантропологии . С. 1611–1631. doi : 10.1007/978-3-540-33761-4_52 .
- ^ Spoor, F.; Gunz, P.; Neubauer, S.; Stelzer, S.; Скотт, Н.; Kwekason, A.; Дин, MC (март 2015 г.). «Реконструированный Homo Habilis Type OH 7 предлагает глубоко укоренившееся видовое разнообразие в раннем гомо». Природа . 519 (7541): 83–86. Bibcode : 2015natur.519 ... 83S . doi : 10.1038/nature14224 . PMID 25739632 . S2CID 4470282 .
- ^ Jump up to: а беременный Haile-Selassie Y, Gibert L, Melillo SM, Ryan TM, Alene M, Deino A, et al. (Май 2015). «Новые виды из Эфиопии еще больше расширяют разнообразие гоминина среднего плиоцена». Природа . 521 (7553): 483–8. Bibcode : 2015natur.521..483h . doi : 10.1038/nature14448 . PMID 26017448 . S2CID 4455029 .
- ^ Фоли, Роберт А.; Миразон Лар, Марта (январь 2024 г.). «Призраки вымерших обезьян: геномное понимание африканской эволюции гоминидов» . Тенденции в экологии и эволюции . 39 (5): 456–466. Bibcode : 2024tecoe..39..456f . doi : 10.1016/j.tree.2023.12.009 . PMID 38302324 .
- ^ Jump up to: а беременный в Indriati E, Swisser CC, Leplace C, Quinn RL, Suriyanto RA, Hascaryo AT, et al. (2011). «Возраст 20 -метровой Solo River Terrace, Java, Индонезия и выживание Homo erectus в Азии » Plos один 6 (6): E2 Bibcode : 2011ploso ... 621562i Doi : 10.1371/ journal.pone.0 3126814PMC 21738710PMID Полем
- ^ Callaway, E. (7 июня 2017 г.). «Старейшие ископаемые претензии Homo Sapiens переписывают историю нашего вида» . Природа . doi : 10.1038/nature.2017.22114 . Получено 11 июня 2017 года .
- ^ Пост С., Рено Г., Миттник А., Друкер Д.Г., Ружье Х., Кубок С. и др. (Март 2016 г.). «Плейстоценовые митохондриальные геномы предполагают единый серьезный рассеяние неафриканцев и поздний оборот ледниковой популяции в Европе». Текущая биология . 26 (6): 827–33. Bibcode : 2016cbio ... 26..827p . doi : 10.1016/j.cub.2016.01.037 . HDL : 2440/114930 . PMID 26853362 . S2CID 140098861 .
- ^ См.:
- Кармин М., Сааг Л., Висенте М., Уилсон Сайрес М.А., Ярв М., Талас Уг и др. (Апрель 2015). «Недавнее узкое место разнообразия Y -хромосом совпадает с глобальными изменениями в культуре» . Исследование генома . 25 (4): 459–66. doi : 10.1101/gr.186684.114 . PMC 4381518 . PMID 25770088 .
- Pagani L, Lawson DJ, Jagoda E, Mörseburg A, Eriksson A, Mitt M, et al. (Октябрь 2016). «Геномный анализ информирует о событиях миграции во время народа Евразии» . Природа . 538 (7624): 238–242. Bibcode : 2016natur.538..238p . doi : 10.1038/nature19792 . PMC 5164938 . PMID 27654910 .
- ^ Haber M, Jones AL, Connell BA, Arciero E, Yang H, Thomas MG, et al. (Август 2019). «Редкая глубоко укоренившаяся африканская гаплогруппа D0 D0 и ее последствия для расширения современных людей из Африки» . Генетика . 212 (4): 1421–1428. doi : 10.1534/Genetics.119.302368 . PMC 6707464 . PMID 31196864 .
- ^ Jump up to: а беременный Green Re, Krause J, Briggs AW, Maricic T, Stenzel U, Kircher M, et al. (Май 2010). «Проектная последовательность неандертального генома» . Наука . 328 (5979): 710–722. Bibcode : 2010sci ... 328..710G . doi : 10.1126/science.1188021 . PMC 5100745 . PMID 20448178 .
- ^ Лоури Р.К., Урибе Г., Хименес Э.Б., Вайс М.А., Эррера К.Дж., Регюро М., Эррера Р.Дж. (ноябрь 2013). «Неандертальские и Денисова генетические сродства с современными людьми: интрогрессия и общие наследственные полиморфизмы». Ген . 530 (1): 83–94. doi : 10.1016/j.gene.2013.06.005 . PMID 23872234 . Это исследование повышает возможность наблюдаемого генетического сродства между архаическим и современным человеческим популяциями, в основном из -за общих наследственных полиморфизмов.
- ^ «Американский словарь английского языка» (4 -е изд.). 2000.
- ^ Stearn, WT (1959). «Фон вкладов Линнеуса в номенклатуру и методы систематической биологии». Систематическая зоология . 8 (1): 4–22. doi : 10.2307/2411603 . JSTOR 2411603 .
- ^ Линн, С. (1758). Система природы. Царство животных (10 изд.). Ex Guilielmi Engelmann. стр. 18, 20 . Получено 19 ноября 2012 года .
- ^ Schwartz, JH; Tattersall, I. (август 2015). «Определение рода гомо ». Наука . 349 (6251): 931–932. Bibcode : 2015sci ... 349..931S . doi : 10.1126/science.aac6182 . PMID 26315422 . S2CID 206639783 .
- ^ Лентс, Н. (4 октября 2014 г.). « Homo naledi и проблемы с родом Homo » . Вильдернист . Архивировано с оригинала 18 ноября 2015 года . Получено 2 ноября 2015 года .
- ^ Вуд, б.; Коллард, М. (апрель 1999 г.). "Человеческий род". Наука . 284 (5411): 65–71. Bibcode : 1999sci ... 284 ... 65. Полем doi : 10.1126/science.284.5411.65 . PMID 10102822 . S2CID 7018418 .
- ^ Пламмер, Томас (9 февраля 2023 г.). «Расширенное географическое распределение и диетические стратегии самых ранних гомининов и парантропа Олдоуэна» . Наука . 379 (6632): 561–566. Bibcode : 2023sci ... 379..561p . doi : 10.1126/science.abo7452 . PMID 36758076 . S2CID 256697931 .
- ^ "Ape-Man", от Pithecanthropus erectus ( Java Man ), Eugène Dubois, Pithecanthropus erectus: человеческий переход из Java (1894), идентифицированный с Pithecanthropus alalus (т.е. "не говорящий Ape-Man") Гиплеры эреки. Гекель
- ^ Хейкель, Эрнст (1895). «Протантроп Примагениус». Систематическая филогения . 3 : с. 625 .
ранний человек
- ^ "Синитан", от Money , Synanthropus Davidson Black (1927).
- ^ «Кривой человек», от Cyphanthropus rodesiensis ( Rhodesian ) William Pycraft (1928).
- ^ «Африканский человек», используемый TF Dreyer (1935) для черепа Флорисбада , который он нашел в 1932 году (также Homo florisbadensis или Homo helmei ). Также род, предложенный для ряда архаичных человеческих черепов, найденных на озере Эйаси Вайнертом (1938). Утечка, журнал Общества естественной истории Восточной Африки »(1942), с. 43
- ^ "удаленный человек"; От Telanthropus capensis (Broom and Robinson 1949), см. (1961), с. 487 .
- ^ из Атлантропа Мавританик , Название дано видам ископаемых (три нижних костях челюсти и теменная кость черепа), обнаруженная в 1954 году по 1955 год Камиллом Арамбургом в Тигеннифе , Алжир. Arambourg, C. (1955). «Недавнее открытие в палеонтологии человека: Атлантроп тернифина (Алжир)». Американский журнал физической антропологии . 13 (2): 191–201. doi : 10.1002/ajpa.1330130203 .
- ^ Coppens, Y. (1965). «Гоминиан Чеда». Деяния V Конг. Ppest . Я : 329f.
- ^ Coppens, Y. (1966). "Le Chadantropus". Антропология . 70 : 5–16.
- ^ Вивело, Александра (25 августа 2013 г.). Характеристика уникальных особенностей денизованского экзома (тезис магистров). Калифорнийский государственный университет . HDL : 10211.3/47490 . Архивировано с оригинала 29 октября 2013 года.
- ^ Баррас, С. (14 марта 2012 г.). «Китайские человеческие окаменелости в отличие от каких -либо известных видов» . Новый ученый . Получено 15 марта 2012 года .
- ^ Грей, JE (1825). «Схема попытки распоряжения млекопитающих в племенах и семьях, со списком родов, по -видимому, аппетитным к каждому племени». Анналы философии . Новая серия: 337–344.
- ^ Wood & Richmond (2000) , с. 19–60
- ^ Brunet, M.; Парень, ф.; Pilbeam, D.; Mackaye, HT; ЛИМИЙС, А.; Ahounta, D.; и др. (Июль 2002 г.). «Новый гоминид из верхнего миоцена Чада, Центральная Африка» (PDF) . Природа . 418 (6894): 145–51. Bibcode : 2002natur.418..145b . doi : 10.1038/nature00879 . PMID 12110880 . S2CID 1316969 .
- ^ Jump up to: а беременный в Cela-Conde, CJ; Айала, FJ (июнь 2003 г.). «Роды человеческой родословной» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (13): 7684–7689. Bibcode : 2003pnas..100.7684c . doi : 10.1073/pnas.0832372100 . PMC 164648 . PMID 12794185 .
- ^ Вуд, б.; Lonergan, N. (апрель 2008 г.). «Запись ископаемого гоминина: таксоны, оценки и клады» (PDF) . Журнал анатомии . 212 (4): 354–76. doi : 10.1111/j.1469-7580.2008.00871.x . PMC 2409102 . PMID 18380861 .
- ^ Пикеринг, Р.; Dirks, ph; Jinnah, Z.; де Руйтер, диджей; Черчилль, SE; Херрис, ИИ; и др. (Сентябрь 2011). «Australopithecus sediba на 1,977 млн. Лет и последствия для происхождения рода Homo» . Наука . 333 (6048): 1421–1423. Bibcode : 2011sci ... 333.1421p . doi : 10.1126/science.1203697 . PMID 21903808 . S2CID 22633702 .
- ^ Asfaw, B.; White, T.; Lovejoy, O.; Latyer, B.; Симпсон, с.; Сува, Г. (апрель 1999 г.). «Австралопитек Гархи: новый вид раннего гоминида из Эфиопии» Наука . 284 (5414): 629–6 Bibcode : 1999sci ... 284..629a Doi : 10.1126/ science.284.5414.6 10213683PMID
- ^ Jump up to: а беременный McPherron, SP; Alemseged, Z.; Мариан, CW; Wynn, JG; Рид, Д.; Geraads, D.; и др. (Август 2010 г.). «Свидетельство о потреблении тканей животных с помощью каменного инструмента до 3,39 миллионов лет назад в Дикике, Эфиопия». Природа . 466 (7308): 857–860. Bibcode : 2010natur.466..857m . doi : 10.1038/nature09248 . PMID 20703305 . S2CID 4356816 .
Самое старое прямое доказательство производства каменных инструментов поступает от GONA (Эфиопия) и датируется от 2,6 до 2,5 миллионов лет (MYR) назад. [...] Здесь мы сообщаем о отметках с каменными наборами на костях, обнаруженных во время недавней работы в области обследования в Дикике, Эфиопии [... показывают] однозначные отметки из камня для удаления плоти [..., датируемой] до 3,42 и 3,24 млн лет назад [...] Наше открытие простирается примерно на 800 000 лет древности каменных инструментов и потребления с использованием каменного инструмента, содержащихся в гонинах; Кроме того, это поведение теперь можно отнести к Australopithecus afarensis.
- ^ См.:
- Villmoare, B.; Kimbel, WH; Seyoum, C.; Лагерь, CJ; Dimaggio, en; J. и др. (Март 2015 г.). «Палуоантропология. Ранний гомо в 2,8 млн. Леджерар, издалека, Эфиопия» . Нанесение 347 (6228): 1352–1355. Bibcode : 2015sci ... 347.1352V . doi : 10.1126/ science.aaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaaa PMID 25739410 . Полем
- Dimaggio EN, Campisano CJ, Rowan J, Dupont-Nivet G, Deino AL, Bibi F, et al. (Март 2015 г.). «Палоантропология. Поздние плиоценовые fossileferous отложение осадок и экологический контекст раннего гомо издалека, Эфиопия » Наука . 347 (6228): 1355–1 Bibcode : 2015sci ... 347.1355d Doi : 10.1126/ science.aaa1 25739409PMID
- ^ Пюшель, Ганс П.; Бертран, Орнелла С.; О'Рейли, Джозеф Э.; Бобе, Рене; Пюшель, Томас А. (июнь 2021 г.). «Оценки дивергенции для гомининов дают представление о энцефализации и тенденциях массы тела в эволюции человека» . Природа экология и эволюция . 5 (6): 808–819. Bibcode : 2021natee ... 5..808p . doi : 10.1038/s41559-021-01431-1 . PMID 33795855 . S2CID 232764044 .
- ^ Вуд, Бернард (28 июня 2011 г.). «Ранний гомо мигрировал» из »или« в «Африке?» Полем Труды Национальной академии наук . 108 (26): 10375–10376. BIBCODE : 2011PNAS..10810375W . doi : 10.1073/pnas.1107724108 . ISSN 0027-8424 . PMC 3127876 . PMID 21677194 .
Адаптивная когерентность гомо будет скомпрометирована, если Х. Хабилис будет включен в HOMO . Таким образом, если эти аргументы принимаются, происхождение рода Homo совпадает со временем и в месте с появлением H. erectus , а не H. habilis .
- ^ Kimbel, WH; Villmoare, B. (июль 2016 г.). «От австралопитека до гомо: переход, который был не» . Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 371 (1698): 20150248. DOI : 10.1098/rstb.2015.0248 . PMC 4920303 . PMID 27298460 .
Свежий взгляд на размер мозга, морфология рук и самая ранняя технология предполагает, что ряд ключевых атрибутов HOMO уже могут присутствовать у обобщенных видов австралопитека, и что адаптивными различиями в HOMO являются просто амплификации или расширения древних тенденций гоминина. [...] Адаптивный сдвиг, представленный ECV австралопитека, по крайней мере, такой же значитель, как и тот, который представлен ECV раннего HOMO , и что основной «скачок» в размере мозга с появлением H. erectus Вероятно, иллюзорно.
- ^ Вуд и Ричмонд (2000) , с. есть мало оснований для сохранения H. habilis 41: «Недавняя переоценка кладистических и функциональных доказательств пришло к выводу, что в HOMO , и рекомендовал перевести материал (или, для некоторых, возвращен) в Australopithecus (Wood & Colrard , 1999). "
- ^ Миллер, JM (май 2000). «Чернофоциальные различия в Homo habilis: анализ доказательств для нескольких видов». Американский журнал физической антропологии . 112 (1): 103–128. doi : 10.1002/(SICI) 1096-8644 (200005) 112: 1 <103 :: AID-AJPA10> 3.0.CO; 2-6 . PMID 10766947 .
- ^ Jump up to: а беременный
Spoor, F.; Leakey, Mg ; Гатого, Пн; Браун, FH; Антон, Южная Каролина; McDougall, я.; и др. (Август 2007 г.). «Последствия новых ранних окаменелостей HOMO от Илерета, к востоку от озера Туркана, Кения». Природа . 448 (7154): 688–691. Bibcode : 2007natur.448..688s . doi : 10.1038/nature05986 . PMID 17687323 . S2CID 35845 .
Частичная верхняя челюсть, назначенная Х. Хабилису, надежно демонстрирует, что этот вид выжил до тех пор, пока не узнал ранее, что маловероятно, что анагенетическая связь с H. erectus. Открытие особенно небольшой кальварии H. erectus указывает на то, что этот таксон перекрывается по размеру с H. habilis, и, возможно, показал заметный половой диморфизм. Новые окаменелости подтверждают отличительность Х. Хабилиса и Х. Эректа, независимо от общего размера черепа, и предполагают, что эти два ранних таксона жили в целом симпатически в одном и том же бассейне озера в течение почти полумиллиона лет.
- ^ Agustí J, Lordkipanidze D (июнь 2011 г.). «Как« африканский »был ранним человеческим рассеиванием из Африки?». Кватернарные науки обзоры . 30 (11–12): 1338–1342. Bibcode : 2011qsrv ... 30.1338a . doi : 10.1016/j.quascirev.2010.04.012 .
- ^ Принс, он; Walrath, D.; McBride, B. (2007). Эволюция и предыстория: человеческий вызов . Wadsworth Publishing. п. 162. ISBN 978-0-495-38190-7 . .
- ^ Циммер, Карл (31 августа 2023 г.). «Предки человечества чуть не погибли, как предполагает генетическое исследование - популяция разбилась после изменения климата около 930 000 лет назад, заключили ученые. Другие эксперты не убеждены в анализе» . New York Times . Архивировано из оригинала 31 августа 2023 года . Получено 2 сентября 2023 года .
- ^ Ху, Ванджи; и др. (31 августа 2023 г.). «Геномный вывод тяжелого узкого места человека во время первого и среднего плейстоценового перехода» . Наука . 381 (6661): 979–984. Bibcode : 2023sci ... 381..979H . doi : 10.1126/science.abq7487 . PMID 37651513 . S2CID 261396309 . Архивировано из оригинала 1 сентября 2023 года . Получено 2 сентября 2023 года .
- ^ Вайс, Кеннет М. (декабрь 1984 г.). «По количеству членов рода Homo, которые когда -либо жили, и некоторые эволюционные последствия» . Человеческая биология . 56 (4): 637–649. JSTOR 41463610 . PMID 6442261 . Получено 12 июня 2024 года .
- ^ Curnoe, D. (июнь 2010 г.). «Обзор раннего HOMO в южной части Африки, посвященный черепно -нижнечелюстному и зубным остаткам, с описанием нового вида (Homo gautengensis sp. Nov.)». Гомо . 61 (3): 151–77. doi : 10.1016/j.jchb.2010.04.002 . PMID 20466364 .
- ^ Jump up to: а беременный Шустер А.М. (1997). «Самые ранние останки рода Homo » . Археология . 50 (1) . Получено 5 марта 2015 года .
- ^ Jump up to: а беременный Berger, LR; Ястребы, Дж.; Dirks, ph; Elliott, M.; Робертс, Эм (май 2017 г.). Перри, Г.Х. (ред.). «Эволюция гомоминина Homo Naledi и плейстоцена в субикэкейской Африке» . элиф . 6 : E24234. doi : 10.7554/elife.24234 . PMC 5423770 . PMID 28483041 .
- ^ Jump up to: а беременный в Mondal, M.; Bertranpetit, J.; Лао, О. (январь 2019). «Примерные байесовские вычисления с глубоким обучением поддерживают третью архаичную интрогрессию в Азии и Океании» . Природная связь . 10 (1): 246. Bibcode : 2019natco..10..246m . doi : 10.1038/s41467-018-08089-7 . PMC 6335398 . PMID 30651539 .
- ^ Jump up to: а беременный Zeitoun, V. (сентябрь 2003 г.). «Высокое происшествие основной функции в Homo erectus: анатомическое описание прегленоидного бугорка» . Анатомическая запись Часть B: новый анатомист . 274 (1): 148–156. doi : 10.1002/ar.b.10028 . PMID 12964205 .
- ^ Jump up to: а беременный в дюймовый Дембо М., Мацке Н.Дж., Муерс Аё, Коллард М (август 2015 г.). «Байесовский анализ морфологической суперматрицы проливает свет на противоречивые отношения ископаемых гомининов» . Разбирательство. Биологические науки . 282 (1812): 20150943. DOI : 10.1098/rspb.2015.0943 . PMC 4528516 . PMID 26202999 .
- ^ Jump up to: а беременный в Дембо М., Радавчич Д., Гарвин Х.М., Лейрд М.Ф., Шредер Л., Скотт Дж. И. и др. (Август 2016 г.). «Эволюционные отношения и возраст Homo Naledi: оценка с использованием датированных байесовских филогенетических методов». Журнал человеческой эволюции . 97 : 17–26. Bibcode : 2016jhume..97 ... 17d . doi : 10.1016/j.jhevol.2016.04.008 . HDL : 2164/8796 . PMID 27457542 .
- ^ Jump up to: а беременный KO, KH (декабрь 2016 г.). «Внутреннее скрещивание гоминина и эволюция человеческих вариаций» . Журнал биологических исследований . 23 (1): 17. doi : 10.1186/s40709-016-0054-7 . PMC 4947341 . PMID 27429943 .
- ^ Харрисон, Н. (1 мая 2019 г.). Происхождение европейцев и их доисторические инновации с 6 миллионов до 10 000 г. до н.э.: от 6 миллионов до 10 000 г. до н.э. Algora Publishing. ISBN 9781628943795 .
- ^ См.:
- Пролив D, Grine F, Fleagle J (2015). Анализ филогения гоминина гоминина: кладистический подход . Спрингер. С. 1989–2014 (Cladogram p. 2006). ISBN 978-3-642-39978-7 . .
- Mounier, A.; Caparros, M. (2015). «Филогенетический статус Homo heidelbergensis - кладистское исследование гомининов среднего плейстоцена». BMSAP (по -французски). 27 (3–4): 110–134. doi : 10.1007/s13219-015-0127-4 . ISSN 0037-8984 . S2CID 17449909 .
- Rogers AR, Harris NS, Achenbach AA (февраль 2020 г.). «Неандертальские предки-денисовановые предки перестановлены с отдаленно связанным гоминином» . Наука достижения . 6 (8): eaay5483. Bibcode : 2020scia .... 6.5483r . doi : 10.1126/sciadv.aay5483 . PMC 7032934 . PMID 32128408 .
- ^ Пюшель, Ганс П.; Бертран, Орнелла С.; О'Рейли, Джозеф Э.; Бобе, Рене; Пюшель, Томас А. (июнь 2021 г.). «Оценки дивергенции для гомининов дают представление о энцефализации и тенденциях массы тела в эволюции человека» . Природа экология и эволюция . 5 (6): 808–819. Bibcode : 2021natee ... 5..808p . doi : 10.1038/s41559-021-01431-1 . PMID 33795855 . S2CID 232764044 .
- ^ См.:
- DEDIU, D.; Левинсон, Южная Каролина (1 июня 2018 года). «Пересмотр неандертальского языка: не только мы». Текущее мнение в поведенческих науках . Эволюция языка. 21 : 49–55. doi : 10.1016/j.cobeha.2018.01.001 . HDL : 21.11116/0000-0000-1667-4 . ISSN 2352-1546 . S2CID 54391128 .
- Hubisz, MJ; Уильямс, Ал; Зипель, А. (август 2020 г.). «Картирование потока генов между древними гомининами посредством демографии с учетом графа наследственной рекомбинации» . PLOS Genetics . 16 (8): E1008895. doi : 10.1371/journal.pgen.1008895 . PMC 7410169 . PMID 32760067 .
- Kuhlwilm M, Gronau I, Hubisz MJ, De Filippo C, Prado-Martinez J, Kircher M, et al. (Февраль 2016 г.). «Древний поток генов от ранних современных людей в восточные неандертальцы» . Природа . 530 (7591): 429–33. Bibcode : 2016natur.530..429k . doi : 10.1038/nature16544 . PMC 4933530 . PMID 26886800 .
- Prüfer K, Ricimo F, Patterson N, Jay F, Sankararaman S, Sawyer S, et al. (Январь 2014). «Полная последовательность генома неандертальца из Алтайских гор» . Природа . 505 (7481): 43–9. Bibcode : 2014natur.505 ... 43 с . doi : 10.1038/nature12886 . PMC 4031459 . PMID 24352235 .
- ^ Jump up to: а беременный Callaway, E. (2016). «Свидетельством для межрединирования бонанзы у древних человеческих видов» . Nature News . doi : 10.1038/nature.2016.19394 . S2CID 87029139 .
- ^ Варки А. (апрель 2016 г.). «Почему нет постоянных гибридов людей с денисованами, неандертальцами или кем -либо еще?» Полем Труды Национальной академии наук Соединенных Штатов Америки . 113 (17): E2354. BIBCODE : 2016PNAS..113E2354V . doi : 10.1073/pnas.1602270113 . PMC 48555598 . PMID 27044111 .
- ^ Пикрелл Дж.К., Паттерсон Н., Лох П.Р., Липсон М., Бергер Б., Стоункинг М. и др. (Февраль 2014 г.). «Древнее Западное Евразийское происхождение на юге и восточной Африке» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (7): 2632–2637. Arxiv : 1307.8014 . Bibcode : 2014pnas..111.2632p . doi : 10.1073/pnas.1313787111 . PMC 3932865 . PMID 24550290 .
- ^ Groves C (2017). «Прогресс в человеческой систематике. Обзор» . Paradigmi (2): 59–74. doi : 10.3280/para2017-002005 . ISSN 1120-3404 .
- ^ Бейин А. (2011). «Верхний плейстоценовый человеческий рассеивание из Африки: обзор текущего состояния дебатов» . Международный журнал эволюционной биологии . 2011 (615094): 615094. DOI : 10.4061/2011/615094 . PMC 3119552 . PMID 21716744 .
- ^ Callaway, E. (март 2016 г.). «Старейшая ДНК древняя человеческая ДНК рассвета неандертальцев» . Природа . 531 (7594): 286. Bibcode : 2016nater.531..296c . doi : 10.1038/531286a . PMID 26983523 . S2CID 4459329 .
- ^ Циммер, С. (10 июля 2019 г.). «Кость черепа, обнаруженная в Греции, может изменить историю человеческой предыстории - кость, обнаруженная в пещере, является самой старой современной человеческой ископаемым, когда -либо обнаруженным в Европе. Намекает на то, что люди начали оставлять Африку намного раньше, чем когда -то думали» . New York Times . Получено 11 июля 2019 года .
- ^ Персонал (10 июля 2019 г.). « Самые старые останки» за пределами Африки сброшены человеческие миграционные часы » . Phys.org . Получено 10 июля 2019 года .
- ^ Harvati, K . ; Röding, C.; Босман, Ам; Каракостис, Фа; Грюн, Р.; Стрингер, С .; и др. (Июль 2019). «Окаменелости пещеры апидимы предоставляют самые ранние доказательства Homo Sapiens в Евразии» . Природа . 571 (7766): 500–504. doi : 10.1038/s41586-019-1376-z . PMID 31292546 . S2CID 195873640 .
- ^ Reich D, Green Re, Kircher M, Krause J, Patterson N, Durand EY, et al. (Декабрь 2010). «Генетическая история архаичной группы гоминина из пещеры Денисовой в Сибири» (PDF) . Природа . 468 (7327): 1053–1060. BIBCODE : 2010NATR.468.1053R . doi : 10.1038/nature09710 . HDL : 10230/25596 . PMC 4306417 . PMID 21179161 .
- ^ Рейх Д., Паттер Н., Дельфин М., Дельфин Ф., Нандини М.Р., Паггах I, Ал. (Октябрь 2011). Первый современный человек и Океания " Журнал человеческой генетики 89 (4): 516–5 doi : 10.1016/jhg . PMC 31188841 . PMID 21944045 .
- ^ Хенн Б.М., Гиньу К.Р., Джобин М., Гранка Дж.М., Макферсон Дж. М., Кидд Дж. М. и др. (Март 2011 г.). «Геномное разнообразие охотников-собирателей предполагает южное африканское происхождение для современных людей» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (13): 5154–5162. Bibcode : 2011pnas..108.5154H . doi : 10.1073/pnas.1017511108 . PMC 3069156 . PMID 21383195 .
- ^ Schlebusch CM, Malmström H, Günther T, Sjödin P, Coutinho A, Edlund H, et al. (Ноябрь 2017). «Древние геномы южной африки оценивают современную дивергенцию человека от 350 000 до 260 000 лет назад» . Наука . 358 (6363): 652–655. Bibcode : 2017sci ... 358..652S . doi : 10.1126/science.aao6266 . PMID 28971970 .
- ^ Перкинс, Сид (17 октября 2013 г.). «Череп предполагает, что три ранних человеческих вида были одним из них» . Nature News & Comment .
- ^ Switek, B. (17 октября 2013 г.). «Красивые черепа шпоры дебаты по истории человечества» . National Geographic . Архивировано с оригинала 17 октября 2013 года . Получено 22 сентября 2014 года .
- ^ Lordkipanidze D, Ponce de León MS, Margvelashvili A, Rak Y, Rightmire GP, Vekua A, Zollikofer CP (октябрь 2013). «Полный череп из Дманиси, Джорджии и эволюционная биология раннего гомо». Наука . 342 (6156): 326–31. Bibcode : 2013sci ... 342..326L . doi : 10.1126/science.1238484 . PMID 24136960 . S2CID 20435482 .
- ^ « Homo heidelbergensis - эволюционная разделительная линия между Homo erectus и современными людьми не была резкой» . Деннис О'Нил. Архивировано с оригинала 9 марта 2016 года . Получено 29 ноября 2015 года .
- ^ Mounier, A.; Marchal, F.; Condemi, S. (март 2009 г.). «Homo heidelbergensis отличным видом? Новое понимание челюсти Мауэра» . Журнал человеческой эволюции . 56 (3): 219–246. Bibcode : 2009jhume..56..219m . doi : 10.1016/j.jhevol.2008.12.006 . PMID 19249816 .
- ^ Либерман, де; McBratney, BM; Кровиц, Г. (февраль 2002 г.). «Эволюция и развитие черепной формы у гомосапинов» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (3): 1134–1139. Bibcode : 2002pnas ... 99.1134L . doi : 10.1073/pnas.022440799 . PMC 122156 . PMID 11805284 .
- ^ Уилсон, AC; Максон, LR; Сарич, Вирджиния (июль 1974 г.). «Два типа молекулярной эволюции. Данные из исследований межвидовой гибридизации» . Труды Национальной академии наук Соединенных Штатов Америки . 71 (7): 2843–2847. Bibcode : 1974pnas ... 71.2843w . doi : 10.1073/pnas.71.7.2843 . PMC 388568 . PMID 4212492 .
- ^ Попадин, К.; Gunbin, K.; Пешкин, Л.; Annis, S.; Kraytsberg, Y.; Markuzon, N.; и др. (19 октября 2017 г.). «Митохондриальные псевдогены предполагают повторную межвидовую гибридизацию среди прямых человеческих предков» . Biorxiv . 134502. DOI : 10.1101/134502 . HDL : 11427/36660 .
- ^ Акерманн, RR; Арнольд, ML; Baiz, MD; Cahill, Ja; Cortés-Ortiz, L.; Эванс, BJ; и др. (Июль 2019). «Гибридизация в эволюции человека: понимание других организмов» . Эволюционная антропология . 28 (4): 189–209. doi : 10.1002/evan.21787 . HDL : 2027.42/151330 . PMC 6980311 . PMID 31222847 .
- ^ Schrenk F, Kullmer O, Bromage T (2007). «9 самые ранние предполагаемые гомо окаменелости». В Henke W, Tattersall I (Eds.). Справочник по палеоантропологии . Тол. 1. В сотрудничестве с Торольфом Хардтом. Берлин, Гейдельберг: Спрингер . С. 1611–1631. doi : 10.1007/978-3-540-33761-4_52 . ISBN 978-3-540-32474-4 .
- ^ Dimaggio, en; Campisano, CJ; Rowan, J.; Dupont-Nivet, G.; Деино, Ал; Bibi, F.; и др. (Март 2015 г.). «Палоантропология. Поздние плиоценовые fossileferous отложение осадок и экологический контекст раннего гомо издалека, Эфиопия » Наука . 347 (6228): 1355–9 Bibcode : 2015sci ... 347.1355d Doi : 10.1126/ science.aaa1 25739409PMID S2CID 43455561
- ^ Curnoe, D. (июнь 2010 г.). «Обзор раннего HOMO в южной части Африки, посвященный черепно -нижнечелюстному и зубным остаткам, с описанием нового вида (Homo gautengensis sp. Nov.)». Гомо . 61 (3): 151–177. doi : 10.1016/j.jchb.2010.04.002 . PMID 20466364 .
- ^ Haviland WA, Walrath D, Prins HE , McBride B (2007). Эволюция и предыстория: Человеческий вызов (8 -е изд.). Белмонт, Калифорния: Томсон Уодсворт . п. 162. ISBN 978-0-495-38190-7 .
- ^ Ferring R, Oms O, Agustí J, Berna F, Nioradze M, Shelia T, et al. (Июнь 2011 г.). «Самые ранние человеческие занятия в Дманиси (Грузинский Кавказ) от 1,85-1,78 млн. Лет» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (26): 10432–6. BIBCODE : 2011PNAS..10810432F . doi : 10.1073/pnas.1106638108 . PMC 3127884 . PMID 21646521 .
- ^ Indriati, E.; Swisher, CC; Lepre, C.; Куинн, RL; Суриянто, Ра; Hascaryo, AT; и др. (2011). «Возраст 20 -метровой Solo River Terrace, Java, Индонезия и выживание Homo erectus в Азии » Plos один 6 (6): E2 Bibcode : 2011ploso ... 621562i Doi : 10.1371/ journal.pone.0 3126814PMC 21738710PMID
- ^ Хазарика М. (2007). « Homo erectus/Ergaster и из Африки: недавние события в палеоантропологии и доисторической археологии» (PDF) . EAA Summer School Ebook . Тол. 1. Европейская антропологическая ассоциация . С. 35–41.
Интенсивный курс по биологической антрологии, 1 -я летняя школа Европейской антропологической ассоциации, 16–30 июня 2007 г., Прага, Чешская Республика
- ^ Muttoni G, Scardia G, Kent DV, Swisher CC, Manzi G (2009). «Плейстоценовая магнитохронология ранних участков гоминина в Сепрано и Фонтане Рануччо, Италия». Земля и планетарные научные письма . 286 (1–2): 255–268. Bibcode : 2009e & psl.286..255m . doi : 10.1016/j.epsl.2009.06.032 . HDL : 2434/164132 .
- ^ Dirks, ph; Робертс, Эм; Hilbert-Wolf, H.; Крамерс, JD; Ястребы, Дж.; Dosseto A.; и др. (Май 2017). «Homo naledi и связанные с ними отложения в пещере Rising Star, Южная Африка» . элиф . 6 : E24231. doi : 10.7554/elife.24231 . PMC 5423772 . PMID 28483040 .
- ^ Ji, Q.; Wu, W.; Ji, y.; Li, Q.; Ni, X. (25 июня 2021 г.). «Поздний средний плейстоцен Харбин череп представляет собой новый вид гомо» . Инновация . 2 (3): 100132. Bibcode : 2021innnov ... 200132j . doi : 10.1016/j.xinn.2021.100132 . PMC 8454552 . PMID 34557772 .
- ^ Ni X, Ji Q, Wu W, Shao Q, Ji Y, Zhang C, Liang L, Ge J, Guo Z, Li J, Li Q, Grün R, Stringer C (25 июня 2021 г.). «Массовый череп из Харбина на северо -восточном Китае создает новую человеческую линию среднего плейстоцена» . Инновация . 2 (3): 100130. Bibcode : 2021innnov ... 200130n . doi : 10.1016/j.xinn.2021.100130 . PMC 8454562 . PMID 34557770 .
- ^ Каллавей, Эван (7 июня 2017 г.). «Старейшие ископаемые претензии Homo Sapiens переписывают историю нашего вида» . Природа . doi : 10.1038/nature.2017.22114 . Получено 11 июня 2017 года .
- ^ Пост, C.; Wißing, C.; Китагава, К.; Pagani, L.; Ван Холштейн, Л.; Расимо, Ф.; и др. (Июль 2017). «Глубокий дивергентный архаичный митохондриальный геном обеспечивает нижнюю границу времени для африканского потока генов в неандертальца» . Природная связь . 8 : 16046. Bibcode : 2017natco ... 816046p . doi : 10.1038/ncomms16046 . PMC 5500885 . PMID 28675384 .
- ^ Bischoff JL, Shamp DD, Aramburu A, et al. (Март 2003 г.). «Гоминиды Sima de Los Huesos датируются равенством U/TH (> 350 Kyr) и, возможно, до 400–500 Kyr: новые радиометрические даты». Журнал археологической науки . 30 (3): 275–280. Bibcode : 2003jarsc..30..275b . doi : 10.1006/jasc.2002.0834 . ISSN 0305-4403 .
- ^ Дин, Д.; Hublin, JJ; Holloway, R.; Зиглер Р. (май 1998). «На филогенетическом положении до-нендертальского образца из переигрывателя, Германия» . Журнал человеческой эволюции . 34 (5): 485–508. Bibcode : 1998jhume..34..485d . doi : 10.1006/JHEV.1998.0214 . PMID 9614635 .
- ^ Чанг, г; Kaifu, Y.; Takai, M.; Коно, RT; Грюн, Р.; Matsu'ura, S.; и др. (Январь 2015). «Первый архаичный гомо из Тайваня» . Природная связь . 6 : 6037. Bibcode : 2015natco ... 6.6037c . doi : 10.1038/ncomms7037 . PMC 4316746 . PMID 25625212 .
- ^ Détroit, F.; Миджарес, как; Корни, Дж.; Daver, G.; Zanolli, C.; Dizon, E.; и др. (Апрель 2019). «Новый вид гомо из покойного плейстоцена Филиппин» (PDF) . Природа . 568 (7751): 181–186. Bibcode : 2019natur.568..181d . doi : 10.1038/s41586-019-1067-9 . PMID 30971845 . S2CID 106411053 .
- ^ Циммер С (10 апреля 2019 г.). «Новый человеческий вид, когда -то живший в этой Филиппинской пещере - археологи на острове Лусон выявили кости отдаленно связанных видов, Homo luzonensis, еще больше расширяя человеческое семейное древо» . New York Times . Получено 10 апреля 2019 года .
Библиография
[ редактировать ]- Вуд, Бернард; Ричмонд, Брайан Г. (июль 2000 г.). «Эволюция человека: таксономия и палеобиология» . Журнал анатомии . 197 (1): 19–60. doi : 10.1046/j.1469-7580.2000.19710019.x . PMC 1468107 . PMID 10999270 .
Внешние ссылки
[ редактировать ]- Изучение записей окаменелостей гоминид ( Центр продвинутого изучения палеобиологии гоминидов в Университете Джорджа Вашингтона )
- Виды гоминид
- Выдающиеся гоминидные окаменелости
- Микко архив филогения
- "Homo" в энциклопедии жизни
- Главная временная шкала (интерактивная) - Смитсоновский институт , Национальный музей естественной истории (август 2016 г.).



| Базы данных управления авторитетом : национальный |
|---|