гомо

Homo (от латинского homō «человек») — род человекообразных обезьян , который произошел от рода Australopithecus и включает в себя современные виды Homo sapiens (современные люди ) и ряд вымерших видов (вместе называемых архаичными людьми ), классифицируемых как предковые или близкие к ним виды. относящийся к современному человеку. К ним относятся Homo erectus и Homo neanderthalensis . Самым старым представителем рода является Homo habilis , возраст которого составляет чуть более 2 миллионов лет назад. [а] Homo , вместе с родом Paranthropus , вероятно, наиболее тесно связан с видом Australopithecus africanus в составе Australopithecus . [4] Ближайшими ныне живущими родственниками Homo являются представители рода Pan (который включает шимпанзе и бонобо ), а предки Pan и Homo , по оценкам, разошлись примерно 5,7-11 миллионов лет назад во время позднего миоцена . [5]
Homo erectus появился около 2 миллионов лет назад и распространился по Африке (где его называют Homo ergaster ) и Евразии в ходе нескольких миграций . Будучи адаптивным и успешным видом, он существовал более миллиона лет и постепенно выделился в новые виды примерно 500 000 лет назад. [б] [6]
Анатомически современные люди ( Homo sapiens ) появились примерно 300 000–200 000 лет назад. [7] в Африке, а H. neanderthalensis появился примерно в то же время в Европе и Западной Азии . H. sapiens рассеялся из Африки несколькими волнами , возможно, еще 250 000 лет назад и определенно 130 000 лет назад, при этом так называемое южное расселение началось примерно 70–50 000 лет назад. [8] [9] [10] что привело к прочной колонизации Евразии и Океании 50 000 лет назад. H. sapiens встречался и скрещивался с архаичными людьми в Африке и Евразии. [11] [12] отдельные архаичные ( неразумные Считается, что ) виды людей, включая неандертальцев, выжили примерно 40 000 лет назад.
Имена и таксономия
[ редактировать ]
Латинское homō существительное hominis (родительный падеж ) означает «человек» или « человек » в общем смысле «человек, человечество». [с] Биномиальное название Homo sapiens было придумано Карлом Линнеем (1758 г.). [д] [15] Названия остальных видов рода были введены со второй половины XIX в. ( H. neanderthalensis 1864, H. erectus 1892).
Род Homo до сих пор не имеет строгого определения. [16] [17] [18] С тех пор, как ранняя летопись окаменелостей человека начала медленно выходить из земли, границы и определения этого рода были плохо определены и постоянно менялись. Поскольку не было никаких оснований полагать, что у него когда-либо появятся дополнительные члены, Карл Линней даже не удосужился дать определение Homo , когда впервые создал его для людей в 18 веке. Открытие неандертальца принесло первое дополнение.
Род Homo получил свое таксономическое название, чтобы предположить, что его представители могут быть классифицированы как человеческие. И на протяжении десятилетий 20-го века находки окаменелостей дочеловеческих и ранних человеческих видов из позднего миоцена и раннего плиоцена создали богатую смесь для дискуссий по классификациям. Продолжаются споры об отделении Homo от австралопитека — или, более того, об отделении Homo от Пана . Несмотря на это, классификация окаменелостей Homo совпадает со свидетельствами: (1) компетентного человеческого прямохождения у Homo habilis, унаследованного от более раннего австралопитека, жившего более четырех миллионов лет назад, о чем свидетельствуют следы Лаэтоли ; и (2) культура человеческих орудий , зародившаяся от 2,5 до 3 миллионов лет назад. [19]
С конца 19 до середины 20 веков для ранних окаменелостей человека был предложен ряд новых таксономических названий, в том числе новые родовые названия; большинство из них с тех пор были объединены с Homo в знак признания того, что Homo erectus был одним видом с большим географическим распространением ранних миграций. Многие такие имена сейчас рассматриваются как « синонимы » Homo , включая Pithecanthropus , [20] Протантроп , [21] Синантроп , [22] цифантроп , [23] Африкатропус , [24] телантроп , [25] Атлантантроп , [26] и Чадантроп . [27] [28]
Классификация рода Homo на виды и подвиды требует неполной информации и остается плохо сделанной. научных статьях, чтобы избежать трехчленных названий или двусмысленности классификации групп как incertae sedis (неопределенное размещение) — например, H. neanderthalensis vs. Это привело к использованию общих названий («неандерталец» и «денисовец») даже в sapiens neanderthalensis , или H. georgicus против H. erectus georgicus . [29] Некоторые недавно вымершие виды этого рода были открыты лишь недавно и еще не имеют общепринятых биномиальных названий (см. Denisova hominin ). [30] С начала голоцена вполне вероятно, что Homo sapiens (анатомически современный человек) был единственным сохранившимся видом Homo .
Джон Эдвард Грей (1825 г.) был одним из первых сторонников классификации таксонов путем обозначения племен и семейств. [31] Вуд и Ричмонд (2000) предложили обозначить гоминини («гоминины») как племя , включающее в себя все виды ранних людей и дочеловеческих предков человека, начиная с , последнего общего предка человека и шимпанзе гомининов как и обозначить подтриба Hominini , включающая только род Homo , то есть не включая более ранних прямоходящих гомининов плиоцена, таких как австралопитек , Orrorin tugenensis , Ardipithecus или Sahelanthropus . [32] Альтернативные Hominina обозначения существовали или предлагались: Australopithecinae (Gregory & Hellman 1939) и Preanthropinae (Cela-Conde & Altaba 2002); [33] [34] [35] четыре рода Australopithecus , Ardipithecus , Praeanthropus и Sahelanthropus а позже Села-Конде и Аяла (2003) предложили сгруппировать с Homo внутри Hominini (sans Pan ). [34]
Эволюция
[ редактировать ]Австралопитек и появление Homo
[ редактировать ]Несколько видов, в том числе Australopithecus garhi , Australopithecus sediba , Australopithecus africanus и Australopithecus afarensis , были предложены в качестве предка или сестры линии Homo . [36] [37] У этих видов есть морфологические особенности, которые роднят их с Homo , но нет единого мнения относительно того, кто дал начало Homo .
Особенно с 2010-х годов определение Homo среди австралопитеков стало более спорным. Традиционно появление человека считалось совпадающим с первым использованием каменных орудий ( олдованская индустрия) и, следовательно, по определению с началом нижнего палеолита . Но в 2010 году были представлены доказательства, которые, по-видимому, приписывают использование каменных орудий Australopithecus afarensis около 3,3 миллиона лет назад, почти за миллион лет до первого появления Homo . [38] LD 350-1 , ископаемый фрагмент нижней челюсти, датируемый 2,8 млн лет назад, обнаруженный в 2013 году в Афаре, Эфиопия , был описан как сочетающий в себе «примитивные черты, наблюдаемые у ранних австралопитеков , с производной морфологией, наблюдаемой у более поздних Homo» . [39] Некоторые авторы считают, что развитие Homo приближается к 3 млн лет назад или даже превышает его. [и] Это находит подтверждение в недавнем филогенетическом исследовании гомининов, которое с использованием морфологической, молекулярной и радиометрической информации датирует появление Homo 3,3 млн лет назад (4,30–2,56 млн лет назад). [40] Другие выразили сомнение относительно того, Homo habilis следует ли включать в Homo , предполагая вместо этого происхождение Homo от Homo erectus примерно 1,9 млн лет назад. [41]
Наиболее заметным физиологическим развитием между более ранними видами австралопитеков и Homo является увеличение эндокраниального объема (ECV) примерно с 460 см3. 3 (28 у.е.) от А. гархи до 660 см. 3 (40 у.е.) у H. habilis и далее до 760 см. 3 (46 куб. Дюймов) у H. erectus , 1250 см 3 (76 куб. дюймов) у H. heidelbergensis и до 1760 см. 3 (107 у.е. дюймов) у H. neanderthalensis . Однако устойчивый рост емкости черепа наблюдается уже у автралопитецин и не прекращается после появления Homo , так что он не служит объективным критерием для определения возникновения рода. [42]
Удобный человек
[ редактировать ]Homo habilis появился около 2,1 млн лет назад. Уже до 2010 года высказывались предложения H. habilis относить не к роду Homo , а к австралопитеку . [43] [44] Основная причина включения H. habilis в число Homo , бесспорное использование им орудий, устарела с открытием использования орудий австралопитеками по крайней мере за миллион лет до H. habilis . [38] Более того, долгое время считалось, что H. habilis является предком более изящного Homo ergaster ( Homo erectus ). В 2007 году было обнаружено, что H. habilis и H. erectus сосуществовали в течение значительного времени, что позволяет предположить, что H. erectus произошел не непосредственно от H. habilis , а от общего предка. [45] После публикации черепа № 5 в Дманиси в 2013 году стало меньше уверенности в том, что азиатский H. erectus является потомком африканского H. ergaster , который, в свою очередь, произошел от H. habilis . Вместо этого H. ergaster и H. erectus кажутся вариантами одного и того же вида, который мог возникнуть либо в Африке, либо в Азии. [46] и широко расселился по Евразии (включая Европу , Индонезию , Китай ) на 0,5 млн лет назад. [47]
Мужчина встал
[ редактировать ]Homo erectus Часто предполагалось, что развился анагенетически из H. habilis примерно 2 миллиона лет назад. Этот сценарий был подкреплен открытием Homo erectus georgicus , ранних экземпляров H. erectus, обнаруженных на Кавказе , которые, по-видимому, проявляли переходные черты с H. habilis . Поскольку самые ранние свидетельства существования H. erectus были обнаружены за пределами Африки, считалось вероятным, что H. erectus развился в Евразии, а затем мигрировал обратно в Африку. На основе окаменелостей из формации Кооби Фора , к востоку от озера Туркана в Кении, Spoor et al. (2007) утверждали, что H. habilis , возможно, выжил после появления H. erectus , так что эволюция H. erectus не была бы анагенетической, и H. erectus существовал бы рядом с H. habilis в течение примерно полумиллиона лет. ( 1,9–1,4 миллиона лет назад ), в период ранней Калабрии . [45] 31 августа 2023 года исследователи на основе генетических исследований сообщили, что предков человека узкое место в популяции (от возможных 100 000 до 1 000 особей) произошло «около 930 000 и 813 000 лет назад… продолжалось около 117 000 лет и приблизило предков человека к вымирание». [48] [49]
Вайс (1984) подсчитал, что насчитывалось около 44 миллиардов (вкратце) представителей рода Homo от его происхождения до эволюции H. erectus , около 56 миллиардов особей от H. erectus до эпохи неолита и еще 51 миллиард особей с тех пор. неолит. Это дает возможность возникнуть огромному количеству новых мутационных вариаций в ходе эволюции человека. [50]
отдельный южноафриканский вид Homo gautengensis В 2010 году был постулирован как современник H. erectus . [51]
Филогения
[ редактировать ]−10 — – −9 — – −8 — – −7 — – −6 — – −5 — – −4 — – −3 — – −2 — – −1 — – 0 — | ( О. praegens ) ( О. тугененсис ) ( Ар. кадабба ) ( Ар. ramidus ) |
| ||||||||||||||||||||||||||||
Таксономия Homo внутри человекообразных обезьян оценивается следующим образом: Paranthropus и Homo появляются внутри Australopithecus (здесь показано кладистически с предоставлением Paranthropus , Kenyanthropus и Homo ). [а] [ф] [6] [53] [52] [4] [54] [55] [56] [57] [58] [59] [60] [ чрезмерное цитирование ] Точная филогения австралопитеков до сих пор остается весьма спорной. Приблизительные даты радиации дочерних клад указаны в миллионах лет назад (млн лет назад). [61] [57] Сахелантроп и Оррорин , возможно, сестры австралопитека , здесь не показаны. Названия группировок иногда бывают запутанными, поскольку часто до проведения кладистического анализа предполагаются определенные группировки. [55]
| Гоминодея |
| ||||||||||||||||||||||||
| (20,4 млн лет назад) |
Кладограмма на основе Dembo et al. (2016): [57]
| Австралопитеки |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| (7,3 млн лет назад) |
Некоторые из линий Homo , по-видимому, имеют выжившее потомство в результате интрогрессии в другие линии. Генетические данные указывают на то, что архаичная линия, отделившаяся от других человеческих линий 1,5 миллиона лет назад, возможно, H. erectus , возможно, скрестилась с денисовцами около 55 000 лет назад. [62] [54] [63] Ископаемые данные показывают, что H. erectus ss дожил по крайней мере до 117 000 лет назад, а еще более базальный H. floresiensis дожил до 50 000 лет назад. возникшая 1,5 миллиона лет назад, Линия, подобная H. erectus, по-видимому, проникла в современных людей через денисовцев и, в частности, в папуасов и аборигенов Австралии. [54] Геномы африканских людей, живущих за пределами Сахары, демонстрируют многочисленные независимые события интрогрессии с участием неандертальцев, а в некоторых случаях и денисовцев, произошедшие около 45 000 лет назад. [64] [63] Генетическая структура некоторых групп Африки к югу от Сахары, по-видимому, указывает на интрогрессию из западно-евразийского населения около 3000 лет назад. [58] [65]
Некоторые данные свидетельствуют о том, что Australopithecus sediba можно было бы отнести к роду Homo или поместить в отдельный род из-за его положения по отношению, например, к H. habilis и H. floresiensis . [56] [66]
Рассредоточение
[ редактировать ]Примерно 1,8 миллиона лет назад H. erectus присутствовал как в Восточной Африке ( H. ergaster ), так и в Западной Азии ( H. georgicus ). Предки индонезийского H. floresiensis, возможно, покинули Африку еще раньше. [г] [56]
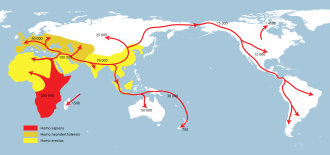
Homo erectus и родственные или производные архаичные виды человека в течение следующих 1,5 миллионов лет распространились по Африке и Евразии. [67] [68] (см.: Недавнее африканское происхождение современного человека ). Европа достигла примерно 0,5 млн лет назад Homo heidelbergensis .
Homo neanderthalensis и H. sapiens развиваются примерно через 300 тыс. лет назад. Homo naledi присутствует в Южной Африке на 300 тыс. лет назад.
H. sapiens вскоре после своего первого появления распространился по Африке и Западной Азии несколькими волнами , возможно, уже через 250 тыс. лет назад и, конечно, к 130 тыс. лет назад. В июле 2019 года антропологи сообщили об обнаружении останков H. sapiens возрастом 210 000 лет и останков H. neanderthalensis возрастом 170 000 лет в пещере Апидима , Пелопоннес , Греция , что более чем на 150 000 лет старше, чем предыдущие находки H. sapiens в Европе. [69] [70] [71]
Наиболее примечательным является южное расселение H. sapiens около 60 тыс. лет назад, которое привело к устойчивому заселению Океании и Евразии анатомически современными людьми . [11] H. sapiens скрещивался с архаичными людьми как в Африке, так и в Евразии, особенно в Евразии с неандертальцами и денисовцами . [72] [73]
Среди существующих популяций H. sapiens самое глубокое временное разделение обнаружено у народа сан в Южной Африке, возраст которого оценивается примерно в 130 000 лет. [74] или, возможно, более 300 000 лет назад. [75] Временное разделение среди неафриканцев составляет порядка 60 000 лет в случае австрало-меланезийцев . Разделение европейцев и аборигенов Восточной Азии длится примерно 50 000 лет, с повторяющимися и значительными событиями смешения по всей Евразии в голоцене .
Архаичные виды человека, возможно, дожили до начала голоцена , хотя в основном они вымерли или были поглощены расширяющейся популяцией H. sapiens к 40 тыс. лет назад ( вымирание неандертальцев ).
Список родословных
[ редактировать ]Видовой статус H. rudolfensis , H. ergaster , H. georgicus , H. antecessor , H. cepranensis , H. rhodesiensis , H. neanderthalensis , Denisova hominin и H. floresiensis остается дискуссионным. H. heidelbergensis и H. neanderthalensis тесно связаны друг с другом и считаются подвидами H. sapiens .
Исторически существовала тенденция постулировать появление новых видов человека, основываясь всего лишь на отдельных окаменелостях. «Минималистский» подход к систематике человека признает не более трех видов: H. habilis (2,1–1,5 млн лет назад, принадлежность к Homo сомнительна), H. erectus (1,8–0,1 млн лет назад, включая большую часть возраста этого рода, и большинство архаичных разновидностей как подвиды , [76] [77] [78] включая H. heidelbergensis как поздний или переходный сорт. [79] [80] [81] ) и Homo sapiens (300 тыс. лет назад, включая H. neanderthalensis и другие разновидности как подвиды ). Последовательные определения и методология определения видов обычно не согласованы в антропологии и палеонтологии. Действительно, видообразующиеся популяции млекопитающих обычно могут скрещиваться в течение нескольких миллионов лет после того, как они начинают генетически расходиться. [82] [83] таким образом, все современные «виды» рода Homo потенциально могли скрещиваться в то время, и интрогрессию из-за пределов рода Homo нельзя априори исключать. [84] Было высказано предположение, что H. naledi мог быть гибридом с поздно выжившим австралипитом (что означает за пределами Homo , ред.), [53] несмотря на то, что эти линии обычно считаются давно вымершими. Как обсуждалось выше, между линиями произошло множество интрогрессий, причем свидетельства интрогрессии произошли после разделения через 1,5 миллиона лет.
| Родословные | Временной диапазон ( принадлежать ) | среда обитания | Рост взрослого | Взрослая масса | Черепная емкость (см 3 ) | Ископаемая запись | Открытие/ публикация имени |
|---|---|---|---|---|---|---|---|
| Х. удобный принадлежность к Homo неясна | 2,100–1,500 [час] [я] | Танзания | 110–140 см (3 фута 7 дюймов – 4 фута 7 дюймов) | 33–55 кг (73–121 фунт) | 510–660 | Много | 1960 1964 |
| Х. рудольфенсис принадлежность к Homo неясна | 1,900 | Кения | 700 | 2 сайта | 1972 1986 | ||
| Х. гаутенгенсис также классифицируется как H. habilis | 1,900–600 | ЮАР | 100 см (3 фута 3 дюйма) | 3 человека [87] [Дж] | 2010 2010 | ||
| Х. прямоходящий | 1,900–140 [88] [к] [89] [л] | Африка , Евразия | 180 см (5 футов 11 дюймов) | 60 кг (130 фунтов) | 850 (рано) – 1100 (поздно) | Много [м] [н] | 1891 1892 |
| Х. эргастер Африканский H. erectus | 1,800–1,300 [91] | Восточная и Южная Африка | 700–850 | Много | 1949 1975 | ||
| Х. предшественник | 1,200–800 | Западная Европа | 175 см (5 футов 9 дюймов) | 90 кг (200 фунтов) | 1,000 | 2 сайта | 1994 1997 |
| Х. гейдельбергенсис ранний H. neanderthalensis | 600–300 [the] | Европа, Африка | 180 см (5 футов 11 дюймов) | 90 кг (200 фунтов) | 1,100–1,400 | Много | 1907 1908 |
| Х. cepranensis единственное ископаемое, возможно, H. heidelbergensis | в. 450 [92] | Италия | 1,000 | 1 тюбетейка | 1994 2003 | ||
| Х. звезда | 335—236 [93] | ЮАР | 150 см (4 фута 11 дюймов) | 45 кг (99 фунтов) | 450 | 15 человек | 2013 2015 |
| Х. длинный | 309–138 [94] | Северо-Восточный Китай | 1,420 [95] | 1 человек | 1933 2021 | ||
| Х. родезийский ранний H. sapiens | в. 300 | Замбия | 1,300 | Один или очень мало | 1921 1921 | ||
| Х. сапиенс (анатомически современный человек) | в. 300 – настоящее время [п] | По всему миру | 150–190 см (4 фута 11 дюймов – 6 футов 3 дюйма) | 50–100 кг (110–220 фунтов) | 950–1,800 | (сохранившийся) | —— 1758 |
| Denisova hominin | в. 285 – ок. 51 | Сибирь | 2 сайта | 2000 2010 [д] | |||
| Х. неандертальский | 240– 40 [98] [р] | Европа, Западная Азия | 170 см (5 футов 7 дюймов) | 55–70 кг (121–154 фунта) (тяжело построенный) | 1,200–1,900 | Много | 1829 1864 |
| Х. флоресский классификация неопределенная | 190–50 | Индонезия | 100 см (3 фута 3 дюйма) | 25 кг (55 фунтов) | 400 | 7 человек | 2003 2004 |
| Нешер Рамла Гомо классификация неопределенная | 140–120 | Израиль | несколько человек | 2021 | |||
| H. tsaichangensis возможно H. erectus или Денисова | в. 100 [с] | Тайвань | 1 человек | 2008(?) 2015 | |||
| Х. лузоненсис | в. 67 [101] [102] | Филиппины | 3 человека | 2007 2019 |
См. также
[ редактировать ]- Список окаменелостей эволюции человека (с изображениями)
- Мультирегиональное происхождение современного человека
Сноски
[ редактировать ]- ^ Jump up to: а б Традиционная оценка возраста H. habilis составляет примерно 2,1–2,3 миллиона лет. [1] [2] Предложения по увеличению возраста до 2,8 млн лет назад были сделаны в 2015 году на основании открытия челюстной кости . [3]
- ^ Homo erectus в узком смысле слова (азиатский вид) вымер 140 000 лет назад; H. erectus soloensis , обнаруженный на Яве , считается последним известным выжившим видом H. erectus . Исследование 2011 года, ранее датированное периодом от 50 000 до 40 000 лет назад, показало, что H. e. Soloensis вымерло самое позднее 143 000 лет назад, но, скорее всего, не позднее 550 000 лет назад. [6]
- ↑ Само слово «человек» происходит от латинского humanus , прилагательного, образованного от корня homo , которое, как полагают, происходит от протоиндоевропейского слова «земля», реконструированного как *dhǵhem- . [13]
- ↑ В 1959 году Карл Линней был признан лектотипом Homo sapiens . [14] это означает, что, следуя номенклатурным правилам , Homo sapiens был обоснованно определен как вид животных, к которому принадлежал Линней.
- ^ Cela-Conde & Ayala (2003) выделяют пять родов внутри Hominina: Ardipithecus , Australopithecus (включая Paranthropus ), Homo (включая Kenyanthropus ), Praeanthropus (включая Orrorin ) и Sahelanthropus . [34]
- ^ Линия самых ранних представителей Homo произошла от австралопитеков , рода, который отделился от последнего общего предка шимпанзе и человека в конце миоцена или начале плиоцена . [52]
- ^ В филогенетическом исследовании 2015 года H. floresiensis был помещен вместе с Australopithecus sediba , H. habilis и Dmanisi Man , что повышает вероятность того, что предки H. floresiensis покинули Африку до появления H. erectus , возможно, даже став первыми гомининами, которые сделали это и получили дальнейшее развитие в Азии. [56]
- ^ Подтвержденные окаменелости H. habilis датируются периодом от 2,1 до 1,5 миллионов лет назад. Этот диапазон дат совпадает с появлением Homo erectus . [85] [86]
- ^ Гоминины с признаками «прото-гомо», возможно, жили еще 2,8 миллиона лет назад, о чем свидетельствует ископаемая челюстная кость, классифицированная как переходная между австралопитеком и человеком, обнаруженная в 2015 году.
- ^ Вид, предложенный в 2010 году на основе ископаемых останков трех особей, датированных периодом от 1,9 до 0,6 миллиона лет назад. Те же окаменелости были классифицированы как H. habilis , H. ergaster или Australopithecus . другими антропологами
- ^ H. erectus, возможно, появился около 2 миллионов лет назад. Окаменелости, датированные 1,8 миллиона лет назад, были найдены как в Африке, так и в Юго-Восточной Азии, а самые древние окаменелости с небольшим отрывом (от 1,85 до 1,77 миллионов лет назад) были найдены на Кавказе, так что неясно, был ли H . erectus возник в Африке и мигрировал в Евразию, или, наоборот, эволюционировал в Евразию и мигрировал обратно в Африку.
- ^ Homo erectus Soloensis , найденный на Яве , считается последним известным выжившим видом H. erectus . Ранее датированное периодом от 50 000 до 40 000 лет назад, исследование 2011 года отодвинуло дату исчезновения H. e. soloensis самое позднее 143 000 лет назад, но, скорее всего, не позднее 550 000 лет назад. [90]
- ^ Теперь в H. erectus также включены пекинский человек (ранее Sinanthropus pekinensis ) и яванский человек (ранее Pithecanthropus erectus ).
- ^ H. erectus теперь сгруппирован в различные подвиды, в том числе Homo erectus erectus , Homo erectus yuanmouensis Homo , , erectus lantianensis Homo erectus nankinensis , Homo erectus pekinensis , Homo erectus palaeojavanicus , Homo erectus Soloensis , Homo erectus tautavelensis , Homo erectus georgicus . Отличие от видов-потомков, таких как Homo ergaster , Homo floresiensis , Homo antecessor , Homo heidelbergensis. да и вообще Homo sapiens не совсем ясно.
- ^ Тип ископаемого — Мауэр 1 , датированный ок. 0,6 миллиона лет назад.Переход от H. heidelbergensis к H. neanderthalensis между 300 и 243 тысячами лет назад является общепринятым и основывается на том факте, что в этот период не существует известных ископаемых. Примерами H. heidelbergensis являются окаменелости, найденные в Билцингслебене (также классифицируемые как Homo erectus bilzingslebensis ).
- ^ Долгое время считалось, что возраст H. sapiens близок к 200 000 лет, но с 2017 года появился ряд предположений о продлении этого времени до 300 000 лет.Окаменелости, найденные в 2017 году в Джебель-Ирхуде ( Марокко ), позволяют предположить, что Homo sapiens мог появиться еще 315 000 лет назад. [96] Генетические данные были представлены для возраста примерно 270 000 лет. [97]
- ^ предварительные названия Homo sp. Алтайский или Homo sapiens ssp. Денисова .
- ^ Первые люди с «чертами протонеандертальца» жили в Евразии еще от 0,6 до 0,35 миллиона лет назад (классифицируются как H. heidelbergensis , также называемая хроновидом , поскольку она представляет собой хронологическую группировку, а не основана на четких морфологических различиях от любого из них). H. erectus или H. neanderthalensis ). В Европе существует ископаемый разрыв между300 и 243 тыс. лет назад, а окаменелости моложе 243 тыс. лет назад принято называть «неандертальцами». [99]
- ^ моложе 450 тысяч лет назад, либо между 190–130, либо между 70–10 тысячами лет назад. [100]
Ссылки
[ редактировать ]- ^ Стрингер, CB (1994). «Эволюция первых людей». В Джонсе, С.; Мартин, Р.; Пилбим, Д. (ред.). Кембриджская энциклопедия эволюции человека . Кембридж: Издательство Кембриджского университета . п. 242.
- ^ Шренк, Ф.; Куллмер, О.; Бромедж, Т. (2007). «Глава 9: Самые ранние предполагаемые окаменелости человека ». Ин Хенке, В.; Таттерсолл, И. (ред.). Справочник палеоантропологии . стр. 1611–1631. дои : 10.1007/978-3-540-33761-4_52 .
- ^ Спур, Ф.; Гунц, П.; Нойбауэр, С.; Стельцер, С.; Скотт, Н.; Квекасон, А.; Дин, MC (март 2015 г.). «Реконструированный тип OH 7 Homo habilis предполагает глубоко укоренившееся видовое разнообразие раннего Homo». Природа . 519 (7541): 83–86. Бибкод : 2015Natur.519...83S . дои : 10.1038/nature14224 . ПМИД 25739632 . S2CID 4470282 .
- ^ Jump up to: а б Хайле-Селассие И., Гиберт Л., Мелилло С.М., Райан Т.М., Ален М., Дейно А. и др. (май 2015 г.). «Новые виды из Эфиопии еще больше расширяют разнообразие гомининов среднего плиоцена». Природа . 521 (7553): 483–8. Бибкод : 2015Natur.521..483H . дои : 10.1038/nature14448 . ПМИД 26017448 . S2CID 4455029 .
- ^ Фоли, Роберт А.; Мирасон Лар, Марта (январь 2024 г.). «Призраки вымерших обезьян: геномный взгляд на эволюцию африканских гоминид» . Тенденции в экологии и эволюции . 39 (5): 456–466. Бибкод : 2024TEcoE..39..456F . дои : 10.1016/j.tree.2023.12.009 . ПМИД 38302324 .
- ^ Jump up to: а б с Индриати Э., Свишер С.С., Лепроси С., Куинн Р.Л., Суриянто Р.А., Хаскарио А.Т. и др. (2011). «Возраст 20-метровой террасы реки Соло, Ява, Индонезия и выживание человека прямоходящего в Азии» . ПЛОС Один . 6 (6): e21562. Бибкод : 2011PLoSO...621562I . дои : 10.1371/journal.pone.0021562 . ПМК 3126814 . ПМИД 21738710 . .
- ^ Каллауэй, Э. (7 июня 2017 г.). «Древнейшие ископаемые останки Homo sapiens переписывают историю нашего вида » Природа . дои : 10.1038/nature.2017.22114 . Проверено 11 июня 2017 г.
- ^ Пост С., Рено Г., Миттник А., Друкер Д.Г., Ружье Х., Купийяр С. и др. (март 2016 г.). «Митохондриальные геномы плейстоцена предполагают единое крупное расселение неафриканцев и смену населения в конце ледникового периода в Европе». Современная биология . 26 (6): 827–33. Бибкод : 2016CBio...26..827P . дои : 10.1016/j.cub.2016.01.037 . hdl : 2440/114930 . ПМИД 26853362 . S2CID 140098861 .
- ^ См.:
- Кармин М., Сааг Л., Висенте М., Уилсон Сейрес М.А., Ярве М., Талас У.Г. и др. (апрель 2015 г.). «Недавнее ограничение разнообразия Y-хромосомы совпадает с глобальными изменениями в культуре» . Геномные исследования . 25 (4): 459–66. дои : 10.1101/гр.186684.114 . ПМЦ 4381518 . ПМИД 25770088 .
- Пагани Л., Лоусон Д.Д., Ягода Э., Мёрзебург А., Эрикссон А., Митт М. и др. (октябрь 2016 г.). «Геномный анализ дает информацию о миграционных событиях во время заселения Евразии» . Природа . 538 (7624): 238–242. Бибкод : 2016Natur.538..238P . дои : 10.1038/nature19792 . ПМК 5164938 . ПМИД 27654910 .
- ^ Хабер М., Джонс А.Л., Коннелл Б.А., Арсьеро Э., Ян Х., Томас М.Г. и др. (август 2019 г.). «Редкая глубоко укоренившаяся африканская гаплогруппа Y-хромосомы D0 и ее значение для расселения современного человека за пределы Африки» . Генетика . 212 (4): 1421–1428. дои : 10.1534/genetics.119.302368 . ПМК 6707464 . ПМИД 31196864 .
- ^ Jump up to: а б Грин Р.Э., Краузе Дж., Бриггс А.В., Маричич Т., Стензель У., Кирхер М. и др. (май 2010 г.). «Проект последовательности генома неандертальца» . Наука . 328 (5979): 710–722. Бибкод : 2010Sci...328..710G . дои : 10.1126/science.1188021 . ПМК 5100745 . ПМИД 20448178 .
- ^ Лоури Р.К., Урибе Дж., Хименес Э.Б., Вайс М.А., Эррера К.Дж., Регейру М., Эррера Р.Дж. (ноябрь 2013 г.). «Генетическое сходство неандертальцев и денисовых с современными людьми: интрогрессия против общих предковых полиморфизмов». Джин . 530 (1): 83–94. дои : 10.1016/j.gene.2013.06.005 . ПМИД 23872234 . Это исследование повышает вероятность того, что наблюдаемое генетическое родство между архаичными и современными человеческими популяциями в основном связано с общим предковым полиморфизмом.
- ^ «Словарь английского языка американского наследия» (4-е изд.). 2000.
- ^ Стерн, WT (1959). «История вклада Линнея в номенклатуру и методы систематической биологии». Систематическая зоология . 8 (1): 4–22. дои : 10.2307/2411603 . JSTOR 2411603 .
- ^ фон Линне, К. (1758). Система природы. Животный мир (10-е изд.). Правительство Гильельми Энгельмана. стр. 18, 20 Проверено 19 ноября 2012 г.
- ^ Шварц, Дж. Х.; Таттерсолл, И. (август 2015 г.). «Определение рода Homo ». Наука . 349 (6251): 931–932. Бибкод : 2015Sci...349..931S . doi : 10.1126/science.aac6182 . ПМИД 26315422 . S2CID 206639783 .
- ^ Ленц, Н. (4 октября 2014 г.). « Homo naledi и проблемы рода Homo » . Вильдернист . Архивировано из оригинала 18 ноября 2015 года . Проверено 2 ноября 2015 г.
- ^ Вуд, Б.; Коллард, М. (апрель 1999 г.). «Род человеческий». Наука . 284 (5411): 65–71. Бибкод : 1999Наука...284...65. . дои : 10.1126/science.284.5411.65 . ПМИД 10102822 . S2CID 7018418 .
- ^ Пламмер, Томас (9 февраля 2023 г.). «Расширенное географическое распространение и диетические стратегии первых олдованских гомининов и парантропов» . Наука . 379 (6632): 561–566. Бибкод : 2023Sci...379..561P . дои : 10.1126/science.abo7452 . ПМИД 36758076 . S2CID 256697931 .
- ^ «человек-обезьяна», от Pithecanthropus erectus ( Яванский человек ), Эжен Дюбуа, Pithecanthropus erectus: человекоподобная переходная форма с Явы (1894 г.), отождествляемая с Pithecanthropus alalus (т. е. «не говорящим человеком-обезьяной»), выдвинул гипотезу. ранее Эрнст Геккель
- ^ Геккель, Эрнст (1895). «Протантроп первородный». Систематическая филогения . 3 : с. 625 .
ранний человек
- ^ «Китайский человек», из Sinanthropus pekinensis ( Пекинский человек ), Дэвидсон Блэк (1927).
- ^ «кривой человек», из Cyphanthropus rhodesiensis ( родезийский человек ) Уильяма Плейна Пайкрафта (1928).
- ^ «Африканский человек», использованный Т. Ф. Дрейером (1935) для обозначения флорисбадского черепа , который он нашел в 1932 году (также Homo florisbadensis или Homo helmei ). Также этот род предполагается для ряда архаичных человеческих черепов, найденных на озере Эяси Вайнертом (1938) . Лики, Журнал Восточноафриканского общества естествознания» (1942), с. 43 .
- ^ "удаленный человек"; от Telanthropus capensis (Broom and Robinson 1949), см. (1961), с. 487 .
- ^ из Atlanthropus mauritanicus ,название, данное виду окаменелостей (три кости нижней челюсти и теменная кость черепа), обнаруженных в 1954–1955 годах Камиллой Арамбургом в Тигеннифе , Алжир. Арамбург, К. (1955). «Недавнее открытие в палеонтологии человека: атлантроп тернифинский (Алжир)». Американский журнал физической антропологии . 13 (2): 191–201. дои : 10.1002/ajpa.1330130203 .
- ^ Коппенс, Ю. (1965). «Оминиен дю Чад». Акты V Конгр. ППЭК . Я : 329ф.
- ^ Коппенс, Ю. (1966). «Чадантроп». Антропология . 70 : 5–16.
- ^ Вивело, Александра (25 августа 2013 г.). Характеристика уникальных особенностей денисовского экзома (магистерская диссертация). Калифорнийский государственный университет . hdl : 10211.3/47490 . Архивировано из оригинала 29 октября 2013 года.
- ^ Баррас, К. (14 марта 2012 г.). «Китайские окаменелости человека не похожи ни на один известный вид» . Новый учёный . Проверено 15 марта 2012 г.
- ^ Грей, Дж. Э. (1825). «Описание попытки разделения млекопитающих на племена и семейства со списком родов, очевидно принадлежащих каждому племени». Анналы философии . новая серия: 337–344.
- ^ Вуд и Ричмонд (2000) , стр. 19–60.
- ^ Брюне, М.; Гай, Ф.; Пилбим, Д.; Маккей, ХТ; Ликиус, А.; Ахаунта, Д.; и др. (июль 2002 г.). «Новый гоминид из верхнего миоцена Чада, Центральная Африка» (PDF) . Природа . 418 (6894): 145–51. Бибкод : 2002Natur.418..145B . дои : 10.1038/nature00879 . ПМИД 12110880 . S2CID 1316969 .
- ^ Jump up to: а б с Села-Конде, CJ; Аяла, Ф.Дж. (июнь 2003 г.). «Роды человеческого рода» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (13): 7684–7689. Бибкод : 2003PNAS..100.7684C . дои : 10.1073/pnas.0832372100 . ПМК 164648 . ПМИД 12794185 .
- ^ Вуд, Б.; Лонерган, Н. (апрель 2008 г.). «Летопись окаменелостей гомининов: таксоны, степени и клады» (PDF) . Журнал анатомии . 212 (4): 354–76. дои : 10.1111/j.1469-7580.2008.00871.x . ПМК 2409102 . ПМИД 18380861 .
- ^ Пикеринг, Р.; Диркс, PH; Джинна, З.; де Рюитер, диджей; Черчилль, SE; Херрис, А.И.; и др. (сентябрь 2011 г.). «Австралопитек седиба, возраст 1,977 млн лет назад, и значение происхождения рода Homo» . Наука . 333 (6048): 1421–1423. Бибкод : 2011Sci...333.1421P . дои : 10.1126/science.1203697 . ПМИД 21903808 . S2CID 22633702 .
- ^ Асфау, Б.; Уайт, Т.; Лавджой, О.; Латимер, Б.; Симпсон, С.; Сува, Г. (апрель 1999 г.). «Австралопитек гархи: новый вид ранних гоминидов из Эфиопии». Наука . 284 (5414): 629–635. Бибкод : 1999Sci...284..629A . дои : 10.1126/science.284.5414.629 . ПМИД 10213683 .
- ^ Jump up to: а б Макферрон, СП; Алемсегед, З.; Мэриан, CW; Винн, Дж. Г.; Рид, Д.; Гераадс, Д.; и др. (август 2010 г.). «Доказательства потребления тканей животных с помощью каменных орудий до 3,39 миллиона лет назад в Дикике, Эфиопия». Природа . 466 (7308): 857–860. Бибкод : 2010Natur.466..857M . дои : 10.1038/nature09248 . ПМИД 20703305 . S2CID 4356816 .
Самые древние прямые свидетельства производства каменных орудий происходят из Гоны (Эфиопия) и датируются периодом от 2,6 до 2,5 миллионов лет назад. [...] Здесь мы сообщаем о следах, оставленных каменными орудиями на костях, обнаруженных во время недавних исследовательских работ в Дикике, Эфиопия, [... показывающих] недвусмысленные следы от порезов каменными орудиями для удаления плоти [..., датированные] периодом 3.42 и 3,24 млн лет назад [...] Наше открытие продлевает примерно на 800 000 лет древность каменных орудий и поедания копытных гоминидами с помощью каменных орудий; более того, теперь такое поведение можно приписать Australopithecus afarensis.
- ^ См.:
- Вильмоаре, Б.; Кимбел, Вашингтон; Сейюм, К.; Кампизано, CJ; ДиМаджио, EN; Роуэн, Дж.; и др. (март 2015 г.). «Палеоантропология. Ранний человек возрастом 2,8 млн лет назад из Леди-Герару, Афар, Эфиопия» . Наука . 347 (6228): 1352–1355. Бибкод : 2015Sci...347.1352V . дои : 10.1126/science.aaa1343 . ПМИД 25739410 . .
- ДиМаджио Э.Н., Камписано С.Дж., Роуэн Дж., Дюпон-Ниве Дж., Дейно А.Л., Биби Ф. и др. (март 2015 г.). «Палеоантропология. Летопись окаменелостей позднего плиоцена и экологический контекст раннего человека издалека, Эфиопия» . Наука . 347 (6228): 1355–1359. Бибкод : 2015Sci...347.1355D . дои : 10.1126/science.aaa1415 . ПМИД 25739409 .
- ^ Пюшель, Ханс П.; Бертран, Орнелла К.; О'Рейли, Джозеф Э.; Бобе, Рене; Пюшель, Томас А. (июнь 2021 г.). «Оценки времени расхождения гомининов дают представление о тенденциях энцефализации и массы тела в эволюции человека» . Экология и эволюция природы . 5 (6): 808–819. Бибкод : 2021NatEE...5..808P . дои : 10.1038/s41559-021-01431-1 . ПМИД 33795855 . S2CID 232764044 .
- ^ Вуд, Бернард (28 июня 2011 г.). «Мигрировали ли ранние люди «из» или «в» Африку?» . Труды Национальной академии наук . 108 (26): 10375–10376. Бибкод : 2011PNAS..10810375W . дои : 10.1073/pnas.1107724108 . ISSN 0027-8424 . ПМК 3127876 . ПМИД 21677194 .
адаптивная когерентность Homo будет нарушена, если H. habilis будет включен в Homo . Таким образом, если принять эти аргументы, то происхождение рода Homo совпадает по времени и месту с появлением H. erectus , а не H. habilis .
- ^ Кимбел, Вашингтон; Вильмоаре, Б. (июль 2016 г.). «От австралопитека к человеку: переход, которого не было» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 371 (1698): 20150248. doi : 10.1098/rstb.2015.0248 . ПМЦ 4920303 . ПМИД 27298460 .
Свежий взгляд на размер мозга, морфологию рук и самые ранние технологии позволяет предположить, что ряд ключевых признаков Homo , возможно, уже присутствует у обобщенных видов австралопитеков, и что адаптивные различия у Homo являются просто усилением или расширением тенденций древних гоминидов. [...] адаптивный сдвиг, представленный ECV австралопитеков, по крайней мере, столь же значителен, как и тот, который представлен ECV ранних Homo , и что с появлением H. erectus произошел значительный скачок в размерах мозга «на уровне класса». вероятно, иллюзорно.
- ^ Вуд и Ричмонд (2000) , с. мало, если таковые вообще имеются 41: «Недавняя переоценка кладистических и функциональных данных пришла к выводу, что оснований для сохранения H. habilis в составе Homo , и рекомендовала передать материал (или, в некоторых случаях, вернуть) австралопитекам (Wood & Collard) , 1999).»
- ^ Миллер, Дж. М. (май 2000 г.). «Черепно-лицевые вариации Homo habilis: анализ доказательств существования нескольких видов». Американский журнал физической антропологии . 112 (1): 103–128. doi : 10.1002/(SICI)1096-8644(200005)112:1<103::AID-AJPA10>3.0.CO;2-6 . ПМИД 10766947 .
- ^ Jump up to: а б Спур, Ф.; Лики, Миннесота ; Гатого, Пенсильвания; Браун, Ф.Х.; Антон, Южная Каролина; Макдугалл, И.; и др. (август 2007 г.). «Последствия новых окаменелостей раннего человека из Илерета, к востоку от озера Туркана, Кения». Природа . 448 (7154): 688–691. Бибкод : 2007Natur.448..688S . дои : 10.1038/nature05986 . ПМИД 17687323 . S2CID 35845 .
Частичная верхняя челюсть, отнесенная к H. habilis, достоверно демонстрирует, что этот вид выживал позже, чем считалось ранее, что делает маловероятным анагенетическое родство с H. erectus. Обнаружение особенно маленького черепа H. erectus указывает на то, что этот таксон по размеру перекрывался с H. habilis и, возможно, демонстрировал выраженный половой диморфизм. Новые окаменелости подтверждают различия H. habilis и H. erectus, независимо от общего размера черепа, и позволяют предположить, что эти два ранних таксона жили в основном симпатрически в одном и том же озерном бассейне в течение почти полумиллиона лет.
- ^ Агусти Дж., Лордкипанидзе Д. (июнь 2011 г.). «Насколько «африканским» было раннее расселение людей из Африки?». Четвертичные научные обзоры . 30 (11–12): 1338–1342. Бибкод : 2011QSRv...30.1338A . doi : 10.1016/j.quascirev.2010.04.012 .
- ^ Принс, Х.Э.; Уолрат, Д.; Макбрайд, Б. (2007). Эволюция и предыстория: человеческий вызов . Издательство Уодсворт. п. 162. ИСБН 978-0-495-38190-7 . .
- ^ Циммер, Карл (31 августа 2023 г.). «Предки человечества почти вымерли, как предполагает генетическое исследование. Численность населения сократилась в результате изменения климата около 930 000 лет назад, заключили ученые. Других экспертов этот анализ не убедил» . «Нью-Йорк Таймс» . Архивировано из оригинала 31 августа 2023 года . Проверено 2 сентября 2023 г.
- ^ Ху, Ванцзе; и др. (31 августа 2023 г.). «Геномный вывод о серьезном узком месте человека во время перехода от раннего к среднему плейстоцену» . Наука . 381 (6661): 979–984. Бибкод : 2023Sci...381..979H . дои : 10.1126/science.abq7487 . ПМИД 37651513 . S2CID 261396309 . Архивировано из оригинала 1 сентября 2023 года . Проверено 2 сентября 2023 г.
- ^ Вайс, Кеннет М. (декабрь 1984 г.). «О количестве когда-либо живших представителей рода Homo и некоторых эволюционных последствиях» . Биология человека . 56 (4): 637–649. JSTOR 41463610 . ПМИД 6442261 . Проверено 12 июня 2024 г.
- ^ Курно, Д. (июнь 2010 г.). «Обзор ранних Homo на юге Африки с упором на остатки черепа, нижней челюсти и зубов, с описанием нового вида (Homo gautengensis sp. nov.)». ГОМО . 61 (3): 151–77. дои : 10.1016/j.jchb.2010.04.002 . ПМИД 20466364 .
- ^ Jump up to: а б Шустер А.М. (1997). «Древние остатки рода Homo » . Археология . 50 (1) . Проверено 5 марта 2015 г.
- ^ Jump up to: а б Бергер, ЛР; Хоукс, Дж.; Диркс, PH; Эллиотт, М.; Робертс, EM (май 2017 г.). Перри, GH (ред.). «Homo naledi и эволюция гоминидов плейстоцена в субэкваториальной Африке» . электронная жизнь . 6 : е24234. doi : 10.7554/eLife.24234 . ПМЦ 5423770 . ПМИД 28483041 .
- ^ Jump up to: а б с Мондаль, М.; Бертранпети, Ж.; Лао, О. (январь 2019 г.). «Приблизительные байесовские вычисления с глубоким обучением поддерживают третью архаичную интрогрессию в Азии и Океании» . Природные коммуникации . 10 (1): 246. Бибкод : 2019NatCo..10..246M . дои : 10.1038/s41467-018-08089-7 . ПМК 6335398 . ПМИД 30651539 .
- ^ Jump up to: а б Зейтун, В. (сентябрь 2003 г.). «Высокая распространенность базикраниальных особенностей у человека прямоходящего: анатомическое описание прегленоидного бугорка» . Анатомические записи, часть B: Новый анатом . 274 (1): 148–156. дои : 10.1002/ar.b.10028 . ПМИД 12964205 .
- ^ Jump up to: а б с д Дембо М., Мацке, Нью-Джерси, Мурс А.О., Коллард М. (август 2015 г.). «Байесовский анализ морфологической суперматрицы проливает свет на спорные взаимоотношения ископаемых гомининов» . Слушания. Биологические науки . 282 (1812): 20150943. doi : 10.1098/rspb.2015.0943 . ПМЦ 4528516 . ПМИД 26202999 .
- ^ Jump up to: а б с Дембо М., Радовчич Д., Гарвин Х.М., Лэрд М.Ф., Шредер Л., Скотт Дж.Э. и др. (август 2016 г.). «Эволюционные взаимоотношения и возраст Homo naledi: оценка с использованием устаревших байесовских филогенетических методов». Журнал эволюции человека . 97 : 17–26. Бибкод : 2016JHumE..97...17D . дои : 10.1016/j.jhevol.2016.04.008 . hdl : 2164/8796 . ПМИД 27457542 .
- ^ Jump up to: а б Ко, К.Х. (декабрь 2016 г.). «Скрещивание гомининов и эволюция человеческих вариаций» . Журнал биологических исследований . 23 (1): 17. дои : 10.1186/s40709-016-0054-7 . ПМЦ 4947341 . ПМИД 27429943 .
- ^ Харрисон, Н. (1 мая 2019 г.). Происхождение европейцев и их доисторические инновации от 6 миллионов до 10 000 лет до нашей эры: от 6 миллионов до 10 000 лет до нашей эры . Издательство Алгора. ISBN 9781628943795 .
- ^ См.:
- Пролив D, Грин Ф, Флигл Дж (2015). Анализ филогении гомининов: кладистический подход . Спрингер. стр. 1989–2014 гг. (кладограмма стр. 2006 г.). ISBN 978-3-642-39978-7 . .
- Мунье, А.; Капаррос, М. (2015). «Филогенетический статус Homo heidelbergensis - кладистическое исследование гомининов среднего плейстоцена». БМСАП (на французском языке). 27 (3–4): 110–134. дои : 10.1007/s13219-015-0127-4 . ISSN 0037-8984 . S2CID 17449909 .
- Роджерс А.Р., Харрис Н.С., Ахенбах А.А. (февраль 2020 г.). «Предки неандертальцев-денисовцев скрещивались с дальним родственником гоминина» . Достижения науки . 6 (8): eaay5483. Бибкод : 2020SciA....6.5483R . дои : 10.1126/sciadv.aay5483 . ПМК 7032934 . ПМИД 32128408 .
- ^ Пюшель, Ханс П.; Бертран, Орнелла К.; О'Рейли, Джозеф Э.; Бобе, Рене; Пюшель, Томас А. (июнь 2021 г.). «Оценки времени расхождения гомининов дают представление о тенденциях энцефализации и массы тела в эволюции человека» . Экология и эволюция природы . 5 (6): 808–819. Бибкод : 2021NatEE...5..808P . дои : 10.1038/s41559-021-01431-1 . ПМИД 33795855 . S2CID 232764044 .
- ^ См.:
- Дедиу, Д.; Левинсон, Южная Каролина (1 июня 2018 г.). «Возвращение к неандертальскому языку: не только мы». Современное мнение в области поведенческих наук . Эволюция языка. 21 : 49–55. дои : 10.1016/j.cobeha.2018.01.001 . hdl : 21.11116/0000-0000-1667-4 . ISSN 2352-1546 . S2CID 54391128 .
- Хубиш, MJ; Уильямс, Алабама; Сипель, А. (август 2020 г.). «Картирование потока генов между древними гомининами посредством демографического вывода графа рекомбинации предков» . ПЛОС Генетика . 16 (8): e1008895. дои : 10.1371/journal.pgen.1008895 . ПМК 7410169 . ПМИД 32760067 .
- Кулвильм М., Гронау И., Хубиш М.Дж., де Филиппо С., Прадо-Мартинес Дж., Кирхер М. и др. (февраль 2016 г.). «Древний поток генов от ранних современных людей к восточным неандертальцам» . Природа . 530 (7591): 429–33. Бибкод : 2016Natur.530..429K . дои : 10.1038/nature16544 . ПМЦ 4933530 . ПМИД 26886800 .
- Прюфер К., Расимо Ф., Паттерсон Н., Джей Ф., Санкарараман С., Сойер С. и др. (январь 2014 г.). «Полная последовательность генома неандертальца с Горного Алтая» . Природа . 505 (7481): 43–9. Бибкод : 2014Natur.505...43P . дои : 10.1038/nature12886 . ПМК 4031459 . ПМИД 24352235 .
- ^ Jump up to: а б Каллауэй, Э. (2016). «Появляется все больше свидетельств того, что скрещивание древних человеческих видов принесло удачу» . Новости природы . дои : 10.1038/nature.2016.19394 . S2CID 87029139 .
- ^ Варки, А. (апрель 2016 г.). «Почему не существует устойчивых гибридов человека с денисовцами, неандертальцами или кем-либо еще?» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (17): Е2354. Бибкод : 2016PNAS..113E2354V . дои : 10.1073/pnas.1602270113 . ПМЦ 4855598 . ПМИД 27044111 .
- ^ Пикрелл Дж.К., Паттерсон Н., Лох П.Р., Липсон М., Бергер Б., Стоункинг М. и др. (февраль 2014 г.). «Древнее западно-евразийское происхождение в южной и восточной Африке» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (7): 2632–2637. arXiv : 1307.8014 . Бибкод : 2014PNAS..111.2632P . дои : 10.1073/pnas.1313787111 . ПМЦ 3932865 . ПМИД 24550290 .
- ^ Гроувс С. (2017). «Прогресс в систематике человека. Обзор» . Парадигми (2): 59–74. дои : 10.3280/PARA2017-002005 . ISSN 1120-3404 .
- ^ Бейин, А. (2011). «Расселение людей из Африки в верхнем плейстоцене: обзор текущего состояния дебатов» . Международный журнал эволюционной биологии . 2011 (615094): 615094. doi : 10.4061/2011/615094 . ПМК 3119552 . ПМИД 21716744 .
- ^ Каллауэй, Э. (март 2016 г.). «ДНК древнейшего человека детализирует рассвет неандертальцев» . Природа . 531 (7594): 286. Бибкод : 2016Natur.531..296C . дои : 10.1038/531286a . ПМИД 26983523 . S2CID 4459329 .
- ^ Циммер, К. (10 июля 2019 г.). «Кость черепа, обнаруженная в Греции, может изменить историю предыстории человечества. Кость, найденная в пещере, является старейшим ископаемым современного человека, когда-либо обнаруженным в Европе. Она намекает на то, что люди начали покидать Африку гораздо раньше, чем когда-то считалось» . Нью-Йорк Таймс . Проверено 11 июля 2019 г.
- ^ Персонал (10 июля 2019 г.). « Самые старые останки за пределами Африки обнуляют часы миграции людей» . Физика.орг . Проверено 10 июля 2019 г.
- ^ Харвати, К. ; Рёдинг, К.; Босман, AM; Каракостис, Ф.А.; Грюн, Р.; Стрингер, К. ; и др. (июль 2019 г.). «Окаменелости пещеры Апидима являются самыми ранними свидетельствами существования Homo sapiens в Евразии» . Природа . 571 (7766): 500–504. дои : 10.1038/s41586-019-1376-z . ПМИД 31292546 . S2CID 195873640 .
- ^ Райх Д., Грин Р.Э., Кирхер М., Краузе Дж., Паттерсон Н., Дюран Э.Ю. и др. (декабрь 2010 г.). «Генетическая история архаической группы гомининов из Денисовой пещеры в Сибири» (PDF) . Природа . 468 (7327): 1053–1060. Бибкод : 2010Natur.468.1053R . дои : 10.1038/nature09710 . hdl : 10230/25596 . ПМК 4306417 . ПМИД 21179161 .
- ^ Райх Д., Паттерсон Н., Кирхер М., Дельфин Ф., Нандинени М.Р., Пугач И. и др. (октябрь 2011 г.). «Денисова примесь и первые расселения современного человека в Юго-Восточную Азию и Океанию» . Американский журнал генетики человека . 89 (4): 516–528. дои : 10.1016/j.ajhg.2011.09.005 . ПМК 3188841 . ПМИД 21944045 .
- ^ Хенн Б.М., Жинью С.Р., Джобин М., Гранка Дж.М., Макферсон Дж.М., Кидд Дж.М. и др. (март 2011 г.). «Геномное разнообразие охотников-собирателей предполагает южноафриканское происхождение современных людей» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (13): 5154–5162. Бибкод : 2011PNAS..108.5154H . дои : 10.1073/pnas.1017511108 . ПМК 3069156 . ПМИД 21383195 .
- ^ Шлебуш К.М., Мальмстрем Х., Гюнтер Т., Сьёдин П., Коутиньо А., Эдлунд Х. и др. (ноябрь 2017 г.). «Древние геномы Южной Африки оценивают расхождение современного человека от 350 000 до 260 000 лет назад» . Наука . 358 (6363): 652–655. Бибкод : 2017Sci...358..652S . дои : 10.1126/science.aao6266 . ПМИД 28971970 .
- ^ Перкинс, Сид (17 октября 2013 г.). «Череп предполагает, что три ранних человеческих вида были одним» . Новости природы и комментарии .
- ^ Свитек, Б. (17 октября 2013 г.). «Красивый череп стимулирует дебаты об истории человечества» . Нэшнл Географик . Архивировано из оригинала 17 октября 2013 года . Проверено 22 сентября 2014 г.
- ^ Лордкипанидзе Д., Понсе де Леон М.С., Маргвелашвили А., Рак Ю., Райтмайр Г.П., Векуа А., Золликофер К.П. (октябрь 2013 г.). «Полный череп из Дманиси, Грузия, и эволюционная биология раннего человека». Наука . 342 (6156): 326–31. Бибкод : 2013Sci...342..326L . дои : 10.1126/science.1238484 . ПМИД 24136960 . S2CID 20435482 .
- ^ « Homo heidelbergensis . Эволюционная разделительная линия между Homo erectus и современным человеком не была резкой» . Деннис О'Нил. Архивировано из оригинала 9 марта 2016 года . Проверено 29 ноября 2015 г.
- ^ Мунье, А.; Маршал, Ф.; Кондеми, С. (март 2009 г.). «Является ли Homo heidelbergensis отдельным видом? Новый взгляд на нижнюю челюсть Мауэра» . Журнал эволюции человека . 56 (3): 219–246. Бибкод : 2009JHumE..56..219M . дои : 10.1016/j.jhevol.2008.12.006 . ПМИД 19249816 .
- ^ Либерман, Делавэр; МакБрэтни, Б.М.; Кровитц, Г. (февраль 2002 г.). «Эволюция и развитие черепной формы у Homosapiens» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (3): 1134–1139. Бибкод : 2002PNAS...99.1134L . дои : 10.1073/pnas.022440799 . ПМК 122156 . ПМИД 11805284 .
- ^ Уилсон, AC; Мэксон, ЛР; Сарич, В.М. (июль 1974 г.). «Два типа молекулярной эволюции. Данные исследований межвидовой гибридизации» . Труды Национальной академии наук Соединенных Штатов Америки . 71 (7): 2843–2847. Бибкод : 1974PNAS...71.2843W . дои : 10.1073/pnas.71.7.2843 . ПМЦ 388568 . ПМИД 4212492 .
- ^ Попадин, К.; Гунбин, К.; Пешкин Л.; Аннис, С.; Крайцберг, Ю.; Маркузон, Н.; и др. (19 октября 2017 г.). «Митохондриальные псевдогены предполагают повторную межвидовую гибридизацию среди прямых предков человека» . биоRxiv . 134502. дои : 10.1101/134502 . hdl : 11427/36660 .
- ^ Акерманн, Р.Р.; Арнольд, ML; Байз, доктор медицины; Кэхилл, Дж.А.; Кортес-Ортис, Л.; Эванс, Би Джей; и др. (июль 2019 г.). «Гибридизация в эволюции человека: опыт других организмов» . Эволюционная антропология . 28 (4): 189–209. дои : 10.1002/evan.21787 . hdl : 2027.42/151330 . ПМК 6980311 . ПМИД 31222847 .
- ^ Шренк Ф., Куллмер О., Бромейдж Т. (2007). «9 самых ранних предполагаемых окаменелостей человека». В Henke W, Tattersall I (ред.). Справочник палеоантропологии . Том. 1. В сотрудничестве с Торольфом Хардтом. Берлин, Гейдельберг: Springer . стр. 1611–1631. дои : 10.1007/978-3-540-33761-4_52 . ISBN 978-3-540-32474-4 .
- ^ ДиМаджио, EN; Кампизано, CJ; Роуэн, Дж.; Дюпон-Ниве, Ж.; Дейно, Алабама; Биби, Ф.; и др. (март 2015 г.). «Палеоантропология. Летопись окаменелостей позднего плиоцена и экологический контекст раннего человека издалека, Эфиопия» . Наука . 347 (6228): 1355–9. Бибкод : 2015Sci...347.1355D . дои : 10.1126/science.aaa1415 . ПМИД 25739409 . S2CID 43455561 .
- ^ Курно, Д. (июнь 2010 г.). «Обзор ранних Homo на юге Африки с упором на остатки черепа, нижней челюсти и зубов, с описанием нового вида (Homo gautengensis sp. nov.)». ГОМО . 61 (3): 151–177. дои : 10.1016/j.jchb.2010.04.002 . ПМИД 20466364 .
- ^ Хэвиленд В.А., Уолрат Д., Принс Х.Э. , Макбрайд Б. (2007). Эволюция и предыстория: человеческий вызов (8-е изд.). Бельмонт, Калифорния: Томсон Уодсворт . п. 162. ИСБН 978-0-495-38190-7 .
- ^ Ферринг Р., Омс О., Агусти Дж., Берна Ф., Ниорадзе М., Шелия Т. и др. (июнь 2011 г.). «Самые ранние заселения человека в Дманиси (Грузинский Кавказ) датируются 1,85-1,78 млн лет назад» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (26): 10432–6. Бибкод : 2011PNAS..10810432F . дои : 10.1073/pnas.1106638108 . ПМК 3127884 . ПМИД 21646521 .
- ^ Индриати, Э.; Свишер, CC; Лепре, К.; Куинн, РЛ; Суриянто, РА; Хаскарьо, Арканзас; и др. (2011). «Возраст 20-метровой террасы реки Соло, Ява, Индонезия и выживание человека прямоходящего в Азии» . ПЛОС Один . 6 (6): e21562. Бибкод : 2011PLoSO...621562I . дои : 10.1371/journal.pone.0021562 . ПМК 3126814 . ПМИД 21738710 .
- ^ Хазарика М (2007). « Homo erectus/ergaster и выходцы из Африки: последние достижения в палеоантропологии и доисторической археологии» (PDF) . Электронная книга Летней школы EAA . Том. 1. Европейская антропологическая ассоциация . стр. 35–41.
Интенсивный курс биологической антропологии, 1-я летняя школа Европейской антропологической ассоциации, 16–30 июня 2007 г., Прага, Чехия.
- ^ Муттони Дж., Скардиа Дж., Кент Д.В., Свишер CC, Манзи Дж. (2009). «Плейстоценовая магнитохронология стоянок ранних гомининов в Чепрано и Фонтана Рануччо, Италия». Письма о Земле и планетологии . 286 (1–2): 255–268. Бибкод : 2009E&PSL.286..255M . дои : 10.1016/j.epsl.2009.06.032 . hdl : 2434/164132 .
- ^ Диркс, PH; Робертс, EM; Гильберт-Вольф, Х.; Крамерс, доктор медицинских наук; Хоукс, Дж.; Доссето А.; и др. (май 2017 г.). «Homo naledi и связанные с ним отложения в пещере Восходящей звезды, Южная Африка» . электронная жизнь . 6 : е24231. doi : 10.7554/eLife.24231 . ПМЦ 5423772 . ПМИД 28483040 .
- ^ Джи, К.; Ву, В.; Джи, Ю.; Ли, К.; Ни, X. (25 июня 2021 г.). «Череп Харбина позднего среднего плейстоцена представляет собой новый вид Homo» . Инновация . 2 (3): 100132. Бибкод : 2021Innov...200132J . дои : 10.1016/j.xinn.2021.100132 . ПМЦ 8454552 . ПМИД 34557772 .
- ^ Ni X, Ji Q, Wu W, Шао Q, Ji Y, Чжан C, Лян L, Ge J, Го Z, Ли J, Ли Q, Грюн Р, Стрингер C (25 июня 2021 г.). «Массивный череп из Харбина на северо-востоке Китая устанавливает новую человеческую линию среднего плейстоцена» . Инновация . 2 (3): 100130. Бибкод : 2021Innov...200130N . дои : 10.1016/j.xinn.2021.100130 . ПМЦ 8454562 . ПМИД 34557770 .
- ^ Каллауэй, Юэн (7 июня 2017 г.). «Древнейшие ископаемые останки Homo sapiens переписывают историю нашего вида » Природа . дои : 10.1038/nature.2017.22114 . Проверено 11 июня 2017 г.
- ^ Пост, К.; Виссинг, К.; Китагава, К.; Пагани, Л.; ван Гольштейн, Л.; Расимо, Ф.; и др. (июль 2017 г.). «Глубоко расходящийся архаичный митохондриальный геном обеспечивает нижнюю временную границу для потока африканских генов в неандертальцев» . Природные коммуникации . 8 : 16046. Бибкод : 2017NatCo...816046P . дои : 10.1038/ncomms16046 . ПМК 5500885 . ПМИД 28675384 .
- ^ Бишофф Дж.Л., Шамп Д.Д., Арамбуру А. и др. (март 2003 г.). «Дата гоминид Сима-де-лос-Уэсос до равновесия U/Th (>350 тыс. лет) и, возможно, до 400–500 тыс. лет: новые радиометрические даты». Журнал археологической науки . 30 (3): 275–280. Бибкод : 2003JArSc..30..275B . дои : 10.1006/jasc.2002.0834 . ISSN 0305-4403 .
- ^ Дин, Д.; Хаблин, Джей-Джей; Холлоуэй, Р.; Зиглер, Р. (май 1998 г.). «О филогенетическом положении пренеандертальского экземпляра из Райлингена, Германия» . Журнал эволюции человека . 34 (5): 485–508. Бибкод : 1998JHumE..34..485D . дои : 10.1006/jhev.1998.0214 . ПМИД 9614635 .
- ^ Чанг, Швейцария; Кайфу, Ю.; Такай, М.; Коно, РТ; Грюн, Р.; Мацуура, С.; и др. (январь 2015 г.). «Первый архаичный человек из Тайваня» . Природные коммуникации . 6 : 6037. Бибкод : 2015NatCo...6.6037C . дои : 10.1038/ncomms7037 . ПМК 4316746 . ПМИД 25625212 .
- ^ Детройт, Ф.; Михарес, А.С.; Корни, Дж.; Дэйвер, Г.; Занолли, К.; Дизон, Э.; и др. (апрель 2019 г.). «Новый вид Homo из позднего плейстоцена Филиппин» (PDF) . Природа . 568 (7751): 181–186. Бибкод : 2019Natur.568..181D . дои : 10.1038/s41586-019-1067-9 . ПМИД 30971845 . S2CID 106411053 .
- ^ Циммер С (10 апреля 2019 г.). «В этой филиппинской пещере когда-то жил новый человеческий вид – археологи на острове Лусон обнаружили кости дальнего родственного вида, Homo luzonensis, что еще больше расширило генеалогическое древо человека» . Нью-Йорк Таймс . Проверено 10 апреля 2019 г.
Библиография
[ редактировать ]- Вуд, Бернард; Ричмонд, Брайан Г. (июль 2000 г.). «Эволюция человека: систематика и палеобиология» . Журнал анатомии . 197 (1): 19–60. дои : 10.1046/j.1469-7580.2000.19710019.x . ПМЦ 1468107 . ПМИД 10999270 .
Внешние ссылки
[ редактировать ]- Изучение летописи окаменелостей гоминид ( Центр перспективных исследований палеобиологии гоминид при Университете Джорджа Вашингтона )
- Виды гоминидов
- Выдающиеся окаменелости гоминидов
- Архив филогении Микко
- «Гомо» в Энциклопедии жизни
- Хронология человечества (интерактивная) – Смитсоновский институт , Национальный музей естественной истории (август 2016 г.).



| Базы данных органов управления : Национальные |
|---|