Человеческая таксономия
| Гомо ("люди") Временный диапазон: Piacenzian - присутствует ,
| |
|---|---|
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Сорт: | Млекопитающие |
| Заказ: | Приматы |
| Подотряд: | Гаплорини |
| Инфрамерс: | Simiiformes |
| Семья: | Hominidae |
| Подсемейство: | Гоминин |
| Племя: | Гоминини |
| Подтриб: | Гомин |
| Род: | Гомо Linnaeus , 1758 |
| Тип видов | |
| Homo Sapiens Linnaeus, 1758
| |
| Разновидность | |
Другие виды или подвид предложены | |
| Синонимы | |
|
Синонимы | |
−10 — – −9 — – −8 — – −7 — – −6 — – −5 — – −4 — – −3 — – −2 — – −1 — – 0 — | ( O. Prenes ) ( O. tuenensis ) ( Ar. Ramidus ) |
| ||||||||||||||||||||||||||||
Человеческая таксономия - это классификация человеческого вида (систематическое название Homo Sapiens , латинский : «мудрый человек») в рамках зоологической таксономии. Систематический род , Homo , предназначен для включения как анатомически современных людей , так и вымерших сортов архаичных людей . Нынешние люди были обозначены как подвиды Homo Sapiens Sapiens , дифференцированные, по мнению некоторых, от прямого предка, Homo Sapiens Idaltu (с некоторыми другими исследованиями, вместо этого классифицируя Идальту и нынешние люди как принадлежащие к одинаковым подвиде. [ 1 ] [ 2 ] [ 3 ] ).
С момента введения систематических имен в 18 -м веке знание эволюции человека резко возросло, а в 20 -м и начале 21 -го века было предложено ряд промежуточных таксонов. Наиболее широко принятая группировка таксономии воспринимает род HOMO как начатый между двумя до трех миллионами лет назад, разделенную как минимум на два вида: архаичный Homo erectus и современные Homo Sapiens , с примерно дюжиной дальнейших предложений по видам без универсального признания.
Род Homo помещается в племя Hominini рядом с Pan (шимпанзе). По оценкам, эти два рода расходились в течение длительного времени гибридизации, охватывающего примерно 10-6 миллионов лет назад, с возможной примеси еще 4 миллиона лет назад. Подпекает неопределенной достоверности, группирующей архаичной «до-человеческой» или «парарационных» видов, моложе гомо - панели , является австралопитецина (предложено в 1939 году).
Предложение Wood and Richmond (2000) представит Hominina в качестве подтриб вместе с Australopithecina, а HOMO единственный известный род в Hominina. В качестве альтернативы, следуя Cela-Conde и Ayala (2003), «до человечества» или «Proto-Human» рода австралопитека , Ardipithecus , Praeantropus и, возможно, Sahelanthropus , могут быть помещены на равные ноги вместе с родом Homo . Еще более экстремальная точка зрения отвергает разделение PAN и HOMO как отдельные роды, которые, основываясь на принципе приоритета, будет означать реклассификацию шимпанзе как Paniscus Homo (или аналогично). [ 4 ]
Категоризация людей на основе фенотипов является социально противоречивым предметом. Биологи первоначально классифицировали расы как подвиды , но современные антропологи отвергают концепцию расы как полезный инструмент для понимания человечества, и вместо этого рассматривают человечество как сложный, взаимосвязанный генетический континуум. Таксономия гомининов продолжает развиваться. [ 5 ] [ 6 ]
История
[ редактировать ]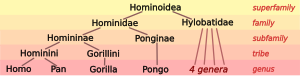
Человеческая таксономия с одной стороны включает в себя размещение людей в таксономии гоминидов ( великих обезьян), а с другой - разделение архаичных и современных людей на виды и, если применимо, подвид . Современная зоологическая таксономия была разработана Карлом Линнеусом в течение 1730-1750 -х годов. Он был первым, кто разрабатывал идею, что, как и другие биологические организации, группы людей могли бы слишком поделиться таксономическими классификациями. [ 7 ] Он назвал человеческий вид Homo Sapiens в 1758 году, как единственный вид члена рода Homo , разделенный на несколько подвидов, соответствующих великим рас . Латинское Homō существительное ( родительный падеж Hominis ) означает «человек». Систематическое название Hominidae для семейства великих обезьян было представлено Джоном Эдвардом Греем (1825). [ 8 ] Грей также предоставил Hominini название племени, включая как шимпанзе (род Pan ), так и людей (род Homo ).
Открытие первого вымершего архаичного человеческого вида из ископаемого записи датируется серединой 19 -го века: Homo neanderthalensis , классифицированный в 1864 году. С тех пор был назван ряд других архаичных видов, но нет универсального консенсуса в отношении их точного число. После открытия H. neanderthalensis , который, даже если «архаика» узнаваем как явно человеческая, антропология с конца 19 -го по начало 20 -го века на некоторое время была занята нахождением предположительно « пропавшего звена » между гомо и панмийтом . « Питтоун -человек » 1912 года был мошенническим представлением такого переходного вида. С середины 20-го века знание развития гоминини стало гораздо более подробным, и таксономическая терминология несколько раз изменялась для этого.
Внедрение Australopithecus в качестве третьего рода, вместе с Homo и Pan , в племени Hominini связано с Raymond Dart (1925). Australopithecina как подтриба, содержащая Australopithecus, а также Paranthropus ( Broom 1938) - это предложение Gregory & Hellman (1939). В последнее время предложенные дополнения к подборибе Australopithecina включают Ardipithecus (1995) и Kenanthropus (2001). Позиция Сахелантропа (2002) относительно Австралопитекины в гоминини неясно. Cela-Conde и Ayala (2003) предлагают распознавание Australopithecus , Ardipithecus , Praeantropus и Sahelanthropus (последний Incertae Sedis ) в качестве отдельных родов. [ 9 ]
Другие предложенные роды, которые в настоящее время в основном считаются частью HOMO , включают в себя: Питекантроп (Дюбуа, 1894), Протантроп (Haeckel, 1895), Сикантроп (черный, 1927), Cyphanthropus (Pycraft, 1928) Африканец (Дрейер, 1935), [ 10 ] Telanthropus (Broom & Anderson 1949), Атлантроп (Arambourg, 1954), Чамантроп (Coppens, 1965).
Род Homo был взят на то, чтобы исчез, около двух миллионов лет назад, так как в 1960 -х годах открытие каменных инструментов в ущелье Олдуваи , Танзания . Homo Habilis (Leakey et al. , 1964) станет первым «человеческим» видом (член рода Homo ) по определению, его типовым образцом является окаменелости OH 7 . Тем не менее, открытие большего количества окаменелостей этого типа открыло дебаты о разграничении H. habilis из Australopithecus . Особенно, LD 350-1 Jawbone Fossil, обнаруженная в 2013 году, датированную 2,8 млн лет, была утверждена как переходная между ними. [ 11 ] Также оспаривается, был ли Х. Хабилис первым гоминином, который использовал каменные инструменты, как австралопитек Гархи , датированный c. 2,5 млн лет было найдено вместе с каменными инструментами. [ 12 ] Fossil KNM-ER 1470 (обнаруженная в 1972 году, обозначенный Pithecanthropus Rudolfensis от Alekseyev 1978) в настоящее время рассматривается как третий ранний вид Homo (вместе с H. habilis и H. erectus ) около 2 миллионов лет назад, либо альтернативно как переходной между Australopithecus и Homo . [ 13 ]
Вуд и Ричмонд (2000) предположили, что племя Грея Гомининини («гоминины») было обозначено как включающее все виды после того, как шимпанзе-Хуман Последний общий предок по определению, к включению австралопитеков и других возможных до человеческих или пара-человеческих видов ( такие как Ardipithecus и Sahelanthropus ) не известны во времена Грея. [ 14 ] В этом предложении новая подтриба Hominina должна была быть обозначена как включающая в себя исключительно род HOMO , так что Hominini будет иметь два подтриба, австралопитецина и гоминину, с единственным известным родом в гоминине - гоме . Оррорин (2001) был предложен в качестве возможного предка гоминины, но не австралопитецина. [ 15 ]
Pregory 1939 (Pregory & Altaba 2002); [ 16 ] [ 17 ] [ 18 ]
Разновидность
[ редактировать ]по крайней мере, дюжина видов гомо , кроме Homo Sapiens Было предложено, , с различной степенью консенсуса. Homo erectus широко признан как вид, непосредственно наследственный для Homo Sapiens . [ Цитация необходима ] Большинство других предлагаемых видов предлагаются в качестве альтернативы принадлежности для Homo erectus или Homo Sapiens в качестве подвида. Это касается гомо Эргастера, в частности. [ 19 ] [ 20 ] Одно предложение разделяет Homo erectus на африканский и азиатский сорт; Африканец - гомо Эргастер, а азиат - это erectus sensu stricto. (Включение Homo Ergaster с азиатским Homo erectus - это homo erectus sensu lato. ) [ 21 ] По-видимому, существует недавняя тенденция, с доступностью все более трудных для классификации окаменелостей, таких как Skulls Dmanisi (2013) или Homo naledi Fossils (2015), чтобы подчеркнуть все архаичные разновидности под Homo erectus . [ 22 ] [ 23 ] [ 24 ]
| Линии | Временный диапазон ( принадлежать ) |
Среда обитания | Взрослый рост | Взрослый масса | Черновая способность (см 3 ) |
Запись ископаемого | Открытие | Публикация имени |
|---|---|---|---|---|---|---|---|---|
| H. Handy Членство в Homo неопределенное |
2,100–1,500 [ А ] [ B ] | Танзания | 110–140 см (3 фута 7 дюймов - 4 фута 7 дюймов) | 33–55 кг (73–121 фунт) | 510–660 | Много | 1960 | 1964 |
| Х. Рудольфенсис Членство в Homo неопределенное |
1,900 | Кения | 700 | 2 сайта | 1972 | 1986 | ||
| H. Gautengensis также классифицируется как Х. Хабилис |
1,900–600 | ЮАР | 100 см (3 фута 3 дюйма) | 3 человека [ 27 ] [ C ] | 2010 | 2010 | ||
| H. Блюдо | 1,900–140 [ 28 ] [ D ] [ 29 ] [ E ] | Африка , Евразия | 180 см (5 футов 11 дюймов) | 60 кг (130 фунтов) | 850 (рано) - 1100 (поздно) | Много [ f ] [ G ] | 1891 | 1892 |
| Х. Эргастер Африканский H. erectus |
1,800–1,300 [ 31 ] | Восточная и южная Африка | 700–850 | Много | 1949 | 1975 | ||
| H. Предшественник | 1,200–800 | Западная Европа | 175 см (5 футов 9 дюймов) | 90 кг (200 фунтов) | 1,000 | 2 сайта | 1994 | 1997 |
| H. Heidelbergensis Ранний H. neanderthalensis |
600–300 [ H ] | Европа, Африка | 180 см (5 футов 11 дюймов) | 90 кг (200 фунтов) | 1,100–1,400 | Много | 1907 | 1908 |
| H. cepranensis Один ископаемый, возможно, H. heidelbergensis |
в 450 [ 32 ] | Италия | 1,000 | 1 крышка черепа | 1994 | 2003 | ||
| H. Long | 309–138 [ 33 ] | Северо -восточный Китай | 1,420 [ 34 ] | 1 человек | 1933 | 2021 | ||
| H. Rhodesiensis Ранний Х. Сапиенс |
в 300 | Замбия | 1,300 | Одинокий или очень немногие | 1921 | 1921 | ||
| Х. Налед | в 300 [ 35 ] | ЮАР | 150 см (4 фута 11 дюймов) | 45 кг (99 фунтов) | 450 | 15 человек | 2013 | 2015 |
| H. Sapiens (анатомически современные люди) |
в 300 -Present [ я ] | Во всем мире | 150–190 см (4 фута 11 дюймов - 6 футов 3 дюйма) | 50–100 кг (110–220 фунтов) | 950–1,800 | (существует) | —— | 1758 |
| H. neanderthalensis |
240– 40 [ 38 ] [ J ] | Европа, Западная Азия | 170 см (5 футов 7 дюймов) | 55–70 кг (121–154 фунта) (сильно построенный) |
1,200–1,900 | Много | 1829 | 1864 |
| H. floresiensis классификация неопределенна |
190–50 | Индонезия | 100 см (3 фута 3 дюйма) | 25 кг (55 фунтов) | 400 | 7 человек | 2003 | 2004 |
| Несер Рамла Хомо классификация неопределенна |
140–120 | Израиль | Несколько человек | 2021 | ||||
| H. tsaichangensis possibly H. erectus or Denisova |
в 100 [ k ] | Тайвань | 1 человек | 2008(?) | 2015 | |||
| H. luzonensis |
в 67 [ 41 ] [ 42 ] | Филиппины | 3 человека | 2007 | 2019 | |||
| Denisova hominin | 40 | Сибирь | 2 сайта | 2000 |
2010 [ L ] |
Подвид
[ редактировать ]Homo Sapiens Subspecies
[ редактировать ]
Признание или несознание подвида Homo Sapiens имеет сложную историю. Ранг подвидов в зоологии вводится для удобства, а не по объективным критериям, основанным на прагматическом рассмотрении таких факторов, как географическая изоляция и половой отбор . Неформальный таксономический ранг расы по -разному считается эквивалентным или подчиненным званию подвида, а разделение анатомически современных людей ( H. sapiens ) на подвид тесно связано с признанием основных расовых групп на основе генетических изменений человека .
Подвид не может быть распознан независимо: вид будет либо не иметь никаких подвидов вообще, либо, по крайней мере, двух (включая любой вымерший). Следовательно, обозначение существующих подвидов Homo Sapiens Sapiens имеет смысл только в том случае, если будет распознается хотя бы один другой подвид. Х. С. Sapiens приписывается «Линнеус (1758)» по таксономическому принципу координации . [ 44 ] В течение 19-го по середину 20-го века была обычная практика классифицировать основные подразделения существующих H. sapiens как подвиды после Линнеуса (1758), который признал H. s. Americanus , H. s. Europaeus , H. s. Азиат и Х. С. Как группировка местного населения Америки , Западной Евразии , Восточной Азии и Африки к югу от Сахары , соответственно. Linnaeus также включал H. s. Ферус , для «дикой» формы, которую он идентифицировал с дикими детьми , и две другие «дикие» формы для зарегистрированных образцов, которые теперь считаются очень сомнительными (см. Cryptozoology ), H. s. Monstrosus и H. s. Троглодиты . [ 45 ]
Были вариации и дополнения к категориям Линнеуса, таких как H. s. Тасманиан для коренного населения Австралии. [ 46 ] Bory de St. Винсент в своем Essai Sur L'homme Линнеуса 1825) расширил « расовые до пятнадцати » категории « гладких волос »): japeticus (с Ираником , Neptunianus , Индиком , Sinicus , Hyperboreus , ( , , Australasicus American Columbicus , American , American ( Патагоника ; Oilotrichi (« хрустящий »): Aethio- , Cafer , Hottentotus , Melaninus . [ 47 ] Аналогичным образом, Жорж Вахер де Лапуж (1899) также имел категории, основанные на расе, такие как Прискус , Спелей (и т. Д.).
Homo sapiens neanderthalensis был предложен Кингом (1864) в качестве альтернативы Homo neanderthalensis . [ 48 ] Были «таксономические войны» в отношении того, были ли неандертальцы отдельным видом с момента их открытия в 1860 -х годах. Pääbo (2014) создает это как дебаты, которые в принципе неразрешены, «поскольку нет никакого определения видов, идеально описывающих этот случай». [ 49 ] Луи Лартет (1869) предложил Homo Sapiens Fossilis на основе окаменелостей Cro-Magnon .
Есть ряд предложений вымерших разновидностей гомо -сапинов, сделанных в 20 -м веке. Многие из первоначальных предложений не использовали явную триномиальную номенклатуру , хотя они все еще называются действительными синонимами H. sapiens от Wilson & Reeder (2005). [ 50 ] К ним относятся: Grimaldii (Lapouge, 1906); Hauser aurignacensis (Klaatsch & Hauser, 1910), Notathropus eurarfricanus (Sergi, 1911), Человек по высыпению инфрас. Прото-аэтио- (Giuffrida-Ruggeri, 1915) Telanthropus capensis (Broom, 1917); [ 51 ] Человек Ваджакенсис (Дюбуа, 1921) Homo sapiens cro-magnonensis , homo sapiens grimaldiensis (Gregory, 1921) Homo Drennan (Kleinschmidt, 1931) [ 52 ] Homo galiLensis (Joleaud, 1931) = палестинус палестина (McCown & Keith, 1932). [ 53 ] Rightmire (1983) предложил Homo Sapiens Rhodesiensis . [ 54 ]
После Второй мировой войны практика разделения существующих групп населения Homo Sapiens на подвид уменьшилась. Раннее власть, явно избегая разделения H. sapiens на подвид, был Grzimeks Tierleben , опубликованная в 1967–1972 годах. [ 55 ] Поздним примером академического органа, предлагающего, чтобы человеческие расовые группы считались таксономическими подвидом, является Джон Бейкер (1974). [ 56 ] Триномиальная номенклатура Homo Sapiens Sapiens стала популярной для «современных людей» в контексте неандертальцев, которые считаются подвидом H. sapiens во второй половине 20 -го века. Полученная из съезда, широко распространенная в 1980 -х годах, рассмотрение двух подвидов, H. s. Неандерталенсис и Х. С. Sapiens , явное утверждение о том, что « H. s. Sapiens является единственным существующим человеческим подвидом», появляется в начале 1990 -х годов. [ 57 ]
С 2000 -х годов вымерший Homo Sapiens Idaltu (White et al. , 2003) получила широкое признание как подвид Homo sapiens , но даже в этом случае существует несогласное мнение, утверждая, что «черепа могут быть недостаточно отличительными, чтобы гарантировать Новое название подвида ". [ 58 ] Х. С. Неандерталенсис и Х. С. Rhodesiensis по -прежнему считаются отдельными видами некоторыми властями, но открытие генетических доказательств архаичной человеческой примеси с современными людьми в 2010 -х годах вновь открыло детали таксономии архаичных людей. [ 59 ]
Человеческий подвид
[ редактировать ]Homo Erectus с момента его введения в 1892 году был разделен на многочисленные подвиды, многие из которых раньше считали отдельные виды HOMO . Ни один из этих подвидов не имеет универсального консенсуса среди палеонтологов.
- Homo erectus erectus ( Java Man ) (1970 -е годы) [ 60 ]
- Homo erectus yuanmouensis ( юаньмоу человек ) (Li et al., 1977)
- Гомо, возведенный латиньеры ( Lantian Man ) (Woo Ju-Kang, 1964)
- Homo erectus nankinensis ( Nanjing Man ) (1993)
- Homo erectus pekinensis ( Peking Man ) (1970 -е годы) [ 60 ]
- Man erectus palaejavanicus ( Мегантроп ) (Тайлер, 2001)
- Homo erectus soloensis ( Solo Man ) (Oppenoorth, 1932)
- Homo erectus tautavelensis ( Tautavel Man ) (De Lumley and De Lumley, 1971)
- Man Erectus Georgian (1991)
- Homo erectus bilzingslebenensis (Vlček, 2002) [ 61 ]
Смотрите также
[ редактировать ]Сноски
[ редактировать ]- ^ Подтвержденные окаменелости H. habilis датируются от 2,1 до 1,5 миллионов лет назад. Этот диапазон дат перекрывается с появлением Homo erectus . [ 25 ] [ 26 ]
- ^ Гоминины с чертами «прото-гомо», возможно, проживали еще 2,8 миллиона лет назад, как было предложено ископаемой челюсти, классифицированной как переходная между Австралопитеком и гомо , обнаруженным в 2015 году.
- ^ Вид, предложенный в 2010 году на основе ископаемых остатков трех особей, от 1,9 до 0,6 миллиона лет назад. Те же окаменелости также были классифицированы как Х. Хабилис , Х. Эргастер или Австралопитек другими антропологами.
- ^ H. Erectus , возможно, появился около 2 миллионов лет назад. Окаменелости от 1,8 миллионов лет назад были обнаружены как в Африке, так и в Юго -Восточной Азии, и самые старые окаменелости с узким отрывом (от 1,85 до 1,77 миллиона лет назад были обнаружены в Кавказе, так что неясно, не четко ли H. . Erectus появился в Африке и мигрировал в Евразию, или, наоборот, он развивался в Евразии и мигрировал обратно в Африку.
- ^ Homo erectus soloensis , найденный в Java , считается последним известным выживанием H. erectus . Ранее, датированное от 50 000 до 40 000 лет назад, исследование 2011 года оттолкнуло дату его исчезновения H. e. Soloensis до 143 000 лет назад по последним, более вероятном до 550 000 лет назад. [ 30 ]
- ^ Теперь также включены в H. erectus - Peking Man (ранее Sinantropus Pekinensis ) и Java Man (ранее Pithecanthropus Erectus ).
- ^ H. Эркт в том числе Alert erectus , , Man lantianensis в настоящее время сгруппируется в различные подвиды , erectus , Lantianensis Man erector Pekinensis Palaeojavanicus , палеоджаванику , мужчина эректаж Soloensis , человеческий ярость Tautavelensis , Человек Георгика . Различие от качества потомков, такого как человек, эргастер , человек Floresiensis , предок , человек Heidelbergensis И действительно Sapiens не совсем ясен.
- ^ Тип окаменелости - Mauer 1 , датированное ок. 0,6 миллиона лет назад. Переход от H. heidelbergensis в H. neanderthalensis между 300 и 243 тысячами лет назад является обычным и использует тот факт, что в этот период нет известных ископаемых. Примерами H. heidelbergensis являются окаменелости, найденные в Bilzingsleben (также классифицируется как Homo erectus bilzingslebensis ).
- ^ Возраст H. sapiens уже давно приближается к 200 000 лет, но с 2017 года на этот раз продлил ряд предложений до 300 000 лет. В 2017 году окаменелости, обнаруженные в Джебель Ирхоуд ( Марокко ), предполагают, что Homo Sapiens , возможно, поощрялся еще в 315 000 лет назад. [ 36 ] Генетические данные были представлены в возрасте около 270 000 лет. [ 37 ]
- ^ Первые люди с «прото-неандертальскими чертами» жили в Евразии уже с 0,6 до 0,35 миллиона лет назад (классифицируется как H. heidelbergensis , также называемый хроноспецифическими , потому что она представляет хронологическую группу, а не основан на четких морфологических различиях из них. H. erectus или H. neanderthalensis ). В Европе есть ископаемый разрыв между 300 и 243 Kya, а по конвенции окаменелости моложе 243 Kya называются «неандертальцем». [ 39 ]
- ^ моложе 450 KYA, либо между 190–130, либо между 70–10 кА [ 40 ]
- ^ Временные названия Homo sp. Altai или Homo Sapiens Ssp. Денисова .
Ссылки
[ редактировать ]- ^ Стрингер, Крис (12 июня 2003 г.). «Эволюция человека: из Эфиопии». Природа . 423 (6941): 693–695. Bibcode : 2003natur.423..692S . doi : 10.1038/423692a . PMID 12802315 . S2CID 26693109 .
- ^ "Herto Skulls (Homo Sapiens Idaltu)" . Talkorigins org . Получено 7 июня 2016 года .
- ^ Стрингер, C. (2016). «Происхождение и эволюция Homo Sapiens» . Философские транзакции Королевского общества B: биологические науки . 371 (1698): 20150237. DOI : 10.1098/rstb.2015.0237 . PMC 4920294 . PMID 27298468 .
- ^ Джаред Даймонд в третьем шимпанзе (1991) и Морриса Гудмана (2003) Хехт, Джефф (19 мая 2003 г.). «Чимпанза - это человек, подразумевает исследование генов» . Новый ученый . Получено 8 декабря 2011 года .
- ^ К. Вагнер, Дженнифер (2016). «Взгляды антропологов на расу, происхождение и генетику» . Американский журнал физической антропологии . 162 (2): 318–327. doi : 10.1002/ajpa.23120 . PMC 5299519 . PMID 27874171 .
- ^ «Заявление AAA о гонке» . Американская антропологическая ассоциация .
- ^ Маркс, Джонатан (май 2007 г.). «Длинная тень человеческой таксономии Линнеуса» . Природа . 447 (7140): 28. Bibcode : 2007natur.447q..28m . doi : 10.1038/447028a . PMID 17476243 . S2CID 33066198 .
- ^ Грей, JE (1825). «Схема попытки распоряжения млекопитающих в племенах и семьях, со списком родов, по -видимому, аппетитным к каждому племени». Анналы философии . Новая серия: 337–344. Найд 10026538759 .
- ^ Cela-Conde, CJ; Ayala, FJ (2003). «Роды человеческой родословной» . Труды Национальной академии наук . 100 (13): 7684–7689. Bibcode : 2003pnas..100.7684c . doi : 10.1073/pnas.0832372100 . PMC 164648 . PMID 12794185 .
- ^ Представлено для черепа Флорисбада (обнаруженного в 1932 году, Homo florisbadensis или Homo helmei ). Также род, предложенный для ряда архаичных человеческих черепов, найденных на озере Эйаси Вайнертом (1938). Утечка, Журнал Восточной Африки Общества естественной истории (1942), с. 43
- ^ Villmoare, B. (2015). «Ранний гомо в 2,8 млн. Лет от Леджи-Герару, издалека, Эфиопия» . Наука . 347 (6228): 1352–1355. Bibcode : 2015sci ... 347.1352V . doi : 10.1126/science.aaa1343 . PMID 25739410 . Полем Некоторые палеоантропологи считают таксон Х. Хабилиса недействительным, состоящим из ископаемых образцов Австралопитека и гомо . Tattersall, I. & Schwartz, JH, Feminct Renions , Westview Press, Нью -Йорк, 2001, с. 111.
- ^ De heinzelin, J; Кларк, JD; Белый, т; Харт, w; Ренне, P; Woldegabriel, g; Beyene, y; VRBA, E (1999). «Окружающая среда и поведение 2,5-миллионных гоминидов Бури». Наука . 284 (5414): 625–9. Bibcode : 1999sci ... 284..625d . doi : 10.1126/science.284.5414.625 . PMID 10213682 .
- ^ Каплан, Мэтт (8 августа 2012 г.). «Окаменелости указывают на большую семью для человеческих предков». Природа . doi : 10.1038/nature.2012.11144 . S2CID 87482930 .
- ^ Вуд, Бернард; Ричмонд, Брайан Г. (2000). «Эволюция человека: таксономия и палеобиология» . Журнал анатомии . 197 (Pt 1): 19–60. doi : 10.1046/j.1469-7580.2000.19710019.x . PMC 1468107 . PMID 10999270 .
- ^ Рейнольдс, Салли С; Галлахер, Эндрю (29 марта 2012 г.). Африканский генезис: перспективы эволюции гоминина . Издательство Кембриджского университета. ISBN 9781107019959 .
- ^ Бруне, Мишель; Парень, Франк; Pilbeam, Дэвид; Маккей, Хассан Таиссо; ЛИМИЙС, Андосси; Ахаунта, Джимдумалбай; Бовилен, Ален; Блондель, Сециле; Bocherens, Hervé; Boisserie, Жан-Ррена; Де Бонис, Луи; Коппенс, Ив; Дежакс, Джин; Денис, Кристиан; Во время Филиппа; Эйзенманн, Вера; Фаноне, Гонддибе; Фронти, Пьер; Гераадс, Денис; Леманн, Томас; Лихоро, Фабрис; Лачарт, Антуан; Махамат, Адум; Merceron, Gildas; Мушелин, парень; Отеро, Ольга; Кампоманс, Пабло Пелаз; Де Леон, Марсия Понсе; Ярость, Жан-Клод; Сапанет, Мишель; Шустер, Матье; Судре, Джин; Тасси, Паскаль; Валентин, Ксавье; Виньо, Патрик; Виру, Лоран; Заззо, Антуан; Zollikofer, Christoph (11 июля 2002 г.). «Новый гоминид из верхнего миоцена Чада, Центральная Африка». Природа . 418 (6894): 145–151. Bibcode : 2002nnatur.418..145b . Doi : 10.1038/nature00879 . PMID 12110880 . S2CID 1316969 .
- ^ Cela-Conde, CJ; Ayala, FJ (2003). «Роды человеческой родословной» . ПНА . 100 (13): 7684–7689. Bibcode : 2003pnas..100.7684c . doi : 10.1073/pnas.0832372100 . PMC 164648 . PMID 12794185 .
- ^ Вуд, б.; Lonergan, N. (2008). «Запись ископаемого гоминина: таксоны, оценки и клады» . Журнал анатомии . 212 (4): 354–376. doi : 10.1111/j.1469-7580.2008.00871.x . PMC 2409102 . PMID 18380861 .
- ^ Хазарика, Манджи (16–30 июня 2007 г.). « Homo erectus / Ergaster и из Африки: недавние события в палеоантропологии и доисторической археологии» (PDF) . Архивировано с оригинала 15 октября 2013 года.
{{cite news}}: Cs1 maint: непредвзятый URL ( ссылка ) - ^ Кляйн Р. (1999). Человеческая карьера: человеческое биологическое и культурное происхождение . Чикаго, Иллинойс: Университет Чикагской Прессы . ISBN 0226439631 .
- ^ Антон, СК (2003). «Естественная история гомо -эректа» . Американский журнал физической антропологии . 122 : 126–170. doi : 10.1002/ajpa.10399 . PMID 14666536 .
К 1980 -м годам растущее число образцов H. erectus , особенно в Африке, привело к осознанию того, что азиатский H. erectus ( H. erectus sensu Stricto ), когда -то считался таким примитивным, на самом деле более полученным, чем его африканские коллеги. Эти морфологические различия были интерпретированы некоторыми как доказательство того, что более одного вида могут быть включены в H. erectus sensu lato (например, Stringer, 1984; Andrews, 1984; Tattersall, 1986; Wood, 1984, 1991a, B; Schwartz and Tattersall, 2000) ... в отличие от европейской линии, по моему мнению, таксономические проблемы, связанные с азиатским и африканским H. erectus, более неразрешимы. Проблема была наиболее остроодана с именом H. ergaster на основе типа нижней челюсти Knm-Er 992, но также включая частичный скелет и изолированные зубы Knm-Er 803 среди других остается коби (Groves and Mazak, 1975. ) Недавно это конкретное название было применено к большинству ранних африканских и грузинских H. erectus в знак признания менее происходящей природы этих остатков в условиях азиатского H. erectus (см. Wood, 1991a, p. 268; Gabunia et al. , 2000a). По крайней мере, части паратипа H. Ergaster (например, KNM-ER 1805) не включены в большинство современных концепций этого таксона. А H. Ergaster Вопрос остается известным неразрешенным (например, Stringer, 1984; Tattersall, 1986; Wood, 1991a, 1994; Rightmire, 1998b; Gabunia et al., 2000a; Schwartz and Tattersall, 2000), в немалой части, потому что первоначальный диагноз; не при условии сравнения с азиатской ископаемой записи.
- ^ Перкинс, Сид (17 октября 2013 г.). «Череп предполагает, что три ранних человеческих вида были одним из них». Природа . doi : 10.1038/nature.2013.13972 .
- ^ Лордкипанидзе, Давид; Ponce de Leòn, Marcia S.; Margvelashvili, Ann; Рак, Йоэль; Rightmire, G. Philip; Vekua, Abesalom; Zollikofer, Christoph PE (18 октября 2013 г.). «Полный череп из Дманиси, Джорджии и эволюционная биология раннего гомо». Наука . 342 (6156): 326–331. Bibcode : 2013sci ... 342..326L . doi : 10.1126/science.1238484 . PMID 24136960 . S2CID 20435482 .
- ^ Switek, Брайан (17 октября 2013 г.). «Красивые черепа шпоры дебаты по истории человечества» . National Geographic . Архивировано из оригинала 17 октября 2013 года . Получено 22 сентября 2014 года .
- ^ Schrenk F, Kullmer O, Bromage T (2007). «Самые ранние предполагаемые гомо окаменелости». В Henke W, Tattersall I (Eds.). Справочник по палеоантропологии . Тол. 1. В сотрудничестве с Торольфом Хардтом. Берлин, Гейдельберг: Спрингер . С. 1611–1631. doi : 10.1007/978-3-540-33761-4_52 . ISBN 978-3-540-32474-4 .
- ^ Dimaggio EN, Campisano CJ, Rowan J, Dupont-Nivet G, Deino AL, Bibi F, et al. (Март 2015 г.). «Пауантропология. Поздний плиоценовый осадочный отчет и экологический контекст раннего гомо издалека, Эфиопия» Наука . 347 (6228): 1355–9 Bibcode : 2015sci ... 347.1355d Doi : 10.1126/ science.aaa1 25739409PMID S2CID 43455561
- ^ Curnoe D (июнь 2010 г.). «Обзор раннего HOMO в южной части Африки, посвященный черепно -нижнечелюстному и зубным остаткам, с описанием нового вида (Homo gautengensis sp. Nov.)». Гомо . 61 (3): 151–77. doi : 10.1016/j.jchb.2010.04.002 . PMID 20466364 .
- ^ Haviland WA, Walrath D, Prins HE , McBride B (2007). Эволюция и предыстория: Человеческий вызов (8 -е изд.). Белмонт, Калифорния: Томсон Уодсворт . п. 162. ISBN 978-0-495-38190-7 .
- ^ Ferring R, Oms O, Agustí J, Berna F, Nioradze M, Shelia T, et al. (Июнь 2011 г.). «Самые ранние человеческие занятия в Дманиси (Грузинский Кавказ) от 1,85-1,78 млн. Лет» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (26): 10432–6. BIBCODE : 2011PNAS..10810432F . doi : 10.1073/pnas.1106638108 . PMC 3127884 . PMID 21646521 .
- ^ Indriati E, Swisser CC, Leplace C, Quinn RL, Suriyanto RA, Hascaryo AT, et al. (2011). «Возраст 20 -метровой Solo River Terrace, Java, Индонезия и выживание Homo erectus в Азии » Plos один 6 (6): E2 Bibcode : 2011ploso ... 621562i Doi : 10.1371/ journal.pone.0 3126814PMC 21738710PMID
- ^ Хазарика М. (2007). « Homo erectus/Ergaster и из Африки: недавние события в палеоантропологии и доисторической археологии» (PDF) . EAA Summer School Ebook . Тол. 1. Европейская антропологическая ассоциация. С. 35–41.
Интенсивный курс по биологической антрологии, 1 -я летняя школа Европейской антропологической ассоциации, 16–30 июня 2007 г., Прага, Чешская Республика
- ^ Muttoni G, Scardia G, Kent DV, Swisher CC, Manzi G (2009). «Плейстоценовая магнитохронология ранних участков гоминина в Сепрано и Фонтане Рануччо, Италия». Земля и планетарные научные письма . 286 (1–2): 255–268. Bibcode : 2009e & psl.286..255m . doi : 10.1016/j.epsl.2009.06.032 .
- ^ Ji Q, Wu W, Ji Y, Li Q, Ni X (25 июня 2021 г.). «Поздний средний плейстоцен Харбин череп представляет собой новый вид гомо» . Инновация . 2 (3): 100132. DOI : 10.1016/j.xinn.2021.100132 . PMC 8454552 . PMID 34557772 .
- ^ Ni X, Ji Q, Wu W, Shao Q, Ji Y, Zhang C, Liang L, Ge J, Guo Z, Li J, Li Q, Grün R, Stringer C (25 июня 2021 г.). «Массовый череп из Харбина на северо -восточном Китае создает новую человеческую линию среднего плейстоцена» . Инновация . 2 (3): 100130. DOI : 10.1016/j.xinn.2021.100130 . PMC 8454562 . PMID 34557770 .
- ^ Dirks PH, Roberts EM, Hilbert-Wolf H, Kramers JD, Hawks J, Dosseto A, et al. (Май 2017). «Homo naledi и связанные с ними отложения в пещере Rising Star, Южная Африка» . элиф . 6 : E24231. doi : 10.7554/elife.24231 . PMC 5423772 . PMID 28483040 .
- ^ Каллавей, Эван (7 июня 2017 г.). «Старейшие ископаемые претензии Homo Sapiens переписывают историю нашего вида» . Природа . doi : 10.1038/nature.2017.22114 . Получено 11 июня 2017 года .
- ^ Poth C, Wißing C, Kitagawa K, Pagani L, Van Holstein L, Racimo F, et al. (Июль 2017). «Глубокий дивергентный архаичный митохондриальный геном обеспечивает нижнюю границу времени для африканского потока генов в неандертальца» . Природная связь . 8 : 16046. Bibcode : 2017natco ... 816046p . doi : 10.1038/ncomms16046 . PMC 5500885 . PMID 28675384 .
- ^ Bischoff JL, Shamp DD, Aramburu A, et al. (Март 2003 г.). «Гоминиды Sima de Los Huesos датируются равенством U/TH (> 350 Kyr) и, возможно, до 400–500 Kyr: новые радиометрические даты». Журнал археологической науки . 30 (3): 275–280. doi : 10.1006/jasc.2002.0834 . ISSN 0305-4403 .
- ^ Дин Д., Хублин Дж.Дж., Холлоуэй Р., Зиглер Р. (май 1998). «На филогенетическом положении до-нендертальского образца из переигрывателя, Германия». Журнал человеческой эволюции . 34 (5): 485–508. doi : 10.1006/JHEV.1998.0214 . PMID 9614635 .
- ^ Чанг Ч., Кайфу Ю., Такай М., Коно Р.Т., Грюн Р., Мацура С. и др. (Январь 2015). «Первый архаичный гомо из Тайваня» . Природная связь . 6 : 6037. Bibcode : 2015natco ... 6.6037c . doi : 10.1038/ncomms7037 . PMC 4316746 . PMID 25625212 .
- ^ Détroit F, Mijares AS, Corny J, Daver G, Zanolli C, Dizon E и др. (Апрель 2019). «Новый вид гомо из покойного плейстоцена Филиппин» (PDF) . Природа . 568 (7751): 181–186. Bibcode : 2019natur.568..181d . doi : 10.1038/s41586-019-1067-9 . PMID 30971845 . S2CID 106411053 .
- ^ Циммер С (10 апреля 2019 г.). «Новый человеческий вид, когда -то живший в этой Филиппинской пещере - археологи на острове Лусон выявили кости отдаленно связанных видов, Homo luzonensis, еще больше расширяя человеческое семейное древо» . New York Times . Получено 10 апреля 2019 года .
- ^ Ральф, Боб (19 февраля 1987 г.). «Соответствие типу» . Новый ученый . № 1548. с. 59.
Насколько я знаю, для Homo Sapiens нет материала типа . Чтобы быть справедливым по отношению к Линнеусу, практика отмены образцов типа в стороне, похоже, не развивалась до сих пор или около того.
- ^ «Статья 46.1». Глоссарий ICZN (4 -е изд.). Международный кодекс зоологической номенклатуры . Архивировано с оригинала 2016-03-03 . Получено 2018-06-04 .
Заявление принципа координации применяется к названиям видовой группы. Название, установленное для таксона в любом ранге в видовой группе, считается одновременно установленным одним и тем же автором для таксона в другом ранге в группе; Оба номинальных таксона имеют одинаковый тип именного подготовки, независимо от того, был ли этот тип фиксированным изначально или впоследствии.
Homo Sapiens Sapiens редко используется до 1940 -х годов. В 1946 году Джон Венделл Бейли приписывает название Линнеууусу (1758) явно: «Линнеус. Сист. Нат. Изд. 10, том 1. С. 20, 21, 22, перечисляет пять рас, а именно: Homo sapiens sapiens (Белый - Кавказский) [...] », это ошибка, но H. s. Sapiens с тех пор часто приписывается Linnaeus. На самом деле, Linnaeus, Syst. НАТ редакция 10 тот. 1. с. 21 не имеет Homo Sapiens Sapiens , «белая» или «кавказская» гонка вместо этого называется Homo Sapiens Europaeus . Это явно указывается в Бюллетене швейцарского общества по антропологии и этнологии Том 21 (1944), с. 18 (Аргумент не против H. s. Sapiens , а против " H. s. Albus L. ", предложенного фон Эйкшстедтом и Петерсом): «Европейская расовая группа, как подвид, [будет] Homo Sapiens Eurpoaeus L. Heissen» (« «Расовая группа европидала, рассматриваемая подвидом, будет названа H. s. Europeaneus L. »). См.: Джон Р. Бейкер , Race , Oxford University Press (1974), 205 . - ^ Линн, Карл Фон (1758). Система природы. Царство животных (10 изд.). стр. 18
- ^ См. EG Джон Уэнделл Бейли, млекопитающие Вирджинии (1946), с. 356;; Журнал млекопитающих 26-27 (1945), с. 359; Дж. Десмонд Кларк (ред.), Кембриджская история Африки , издательство Кембриджского университета (1982), с. 141 (со ссылками).
- ^ Анналы философии 11, Лондон (1826), с. 71
- ^ Фредерик С. Шсалай, Эрик Делсон, Эволюционная история приматов (2013), 508
- ^ Пябо, Сванте (2014). Неандертальский человек: в поисках потерянных геномов . Нью -Йорк: Основные книги . п. 237
- ^ Groves, CP (2005). Уилсон, де ; Ридер, DM (ред.). Виды млекопитающих мира: таксономическая и географическая ссылка (3 -е изд.). Балтимор: издательство Джона Хопкинса. ISBN 0-801-88221-4 Полем OCLC 62265494 .
- ^ Харрисон, Т. (2013). Кимбел, Уильям Х.; Мартин, Лоуренс Б. (ред.). Виды, видовые концепции и эволюция приматов . Спрингер. п. 361. ISBN 9781489937452 .
- ^ Г -н Дреннан, «Австралоидный череп из Кейп -Флэтс», журнал Королевского антропологического института Великобритании и Ирландии . 59 (июль - декабрь, 1929), 417-427.
- ^ Среди других имен, предложенных для окаменелостей, впоследствии подключенных под NeanderThalensis , см. Дельсон, Эрик; Tattersall, Ian; Couvering, Джон Ван; Брукс, Элисон С. (2004). Энциклопедия эволюции человека и предыстории (второе изд.). Routledge . ISBN 9781135582272 .
- ^ Rightmire, GP (3 июня 1983 г.). «Озеро Ndutu Cranium и Rano Homo Sapiens в Африке». Американский журнал физической антропологии . 61 (2): 245–54. doi : 10.1002/ajpa.1330610214 . PMID 6410925 .
- ^ Перевод английского языка (1972–1975): Энциклопедия животной жизни Грзимека , том 11, с. 55
- ^ Джон Р. Бейкер, Race , Oxford University Press (1974).
- ^ Китахара, Мичио (1991). Трагедия эволюции: человеческое животное противостоит современному обществу . п. xi.
Мы единственные выжившие подвиды Homo Sapiens
- ^ Стрингер, Крис (12 июня 2003 г.). «Эволюция человека: из Эфиопии» . Природа . 423 (6941): 692–3, 695. Bibcode : 2003natur.423..692S . doi : 10.1038/423692a . PMID 12802315 . S2CID 26693109 .
- ^ Hublin, JJ (2009). «Происхождение неандерталов» . Труды Национальной академии наук . 106 (38): 16022–7. Bibcode : 2009pnas..10616022H . doi : 10.1073/pnas.0904119106 . JSTOR 40485013 . PMC 2752594 . PMID 19805257 . Harvati, K.; Фрост, ср; McNulty, KP (2004). «Неандертальская таксономия пересмотрена: последствия трехмерных моделей приматов внутри- и межвидовых различий» . Труды Национальной академии наук . 101 (5): 1147–52. Bibcode : 2004pnas..101.1147h . doi : 10.1073/pnas.0308085100 . PMC 337021 . PMID 14745010 . " Homo neanderthalensis King, 1864". Уайли-Блэквелл Энциклопедия эволюции человека . Чичестер, Западный Суссекс: Wiley-Blackwell . 2013. С. 328–331.
- ^ Jump up to: а беременный В 1970 -х годах была разработана тенденция рассматривать яванское разнообразие H. erectus как подвид, Homo erectus erectus , причем китайский сорт называется Homo erectus pekinensis . Видеть: Сартоно С. (12 мая 2011 г.). Причастный от Pitoricantropus VIII " В Таттле, Рассел Х. (ред.). ́phology Палология: Mor Уолтер де Грютер. п. 328. ISBN 9783110810691 .
- ^ Emanuel Vlček: Fossil Man of Bilzingsleben (= Bilzingsleben. Vol. 6 = Вклад в первоначальную и раннюю историю Центральной Европы 35). Beier & Beran, Langenweißbach 2002.