Фотосистема I

| Фотосистема I | |||
|---|---|---|---|
 Фотосистема растений I с БАК I | |||
| Идентификаторы | |||
| Номер ЕС. | 1.97.1.12 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| |||

Фотосистема I ( PSI , или пластоцианин-ферредоксиноксидоредуктаза — одна из двух фотосистем в фотосинтетических световых реакциях водорослей ) , растений и цианобактерий . Фотосистема I [ 1 ] представляет собой интегральный мембранный белковый комплекс , который использует света энергию для катализа переноса электронов через тилакоидную мембрану от пластоцианина к ферредоксину . В конечном итоге электроны, переносимые Фотосистемой I, используются для производства переносчика водорода средней энергии НАДФН . [ 2 ] Энергия фотонов, поглощаемая Фотосистемой I, также создает протондвижущую силу , которая используется для генерации АТФ . PSI состоит из более чем 110 кофакторов , что значительно больше, чем у Photosystem II . [ 3 ]
История
[ редактировать ]Эта фотосистема известна как PSI, потому что она была открыта до Фотосистемы II, хотя будущие эксперименты показали, что Фотосистема II на самом деле является первым ферментом фотосинтетической цепи переноса электронов. Аспекты PSI были открыты в 1950-х годах, но значение этих открытий в то время еще не было признано. [ 4 ] Луи Дуйсенс впервые предложил концепции фотосистем I и II в 1960 году, и в том же году предложение Фэй Бендалл и Роберта Хилла объединило более ранние открытия в последовательную теорию последовательных фотосинтетических реакций. [ 4 ] Гипотеза Хилла и Бендалла позже была подтверждена в экспериментах, проведенных в 1961 году группами Дуйсенса и Витта. [ 4 ]
Компоненты и действие
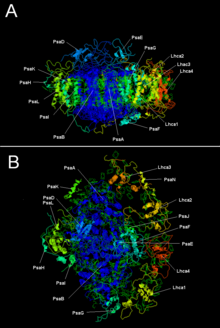
[ редактировать ]Две основные субъединицы PSI, PsaA и PsaB, представляют собой близкородственные белки, участвующие в связывании жизненно важных кофакторов переноса электронов P 700 , Acc, A 0 , A 1 и F x . PsaA и PsaB представляют собой интегральные мембранные белки , содержащие от 730 до 750 аминокислот и содержащие 11 трансмембранных сегментов. Кластер железа и серы [4Fe-4S], называемый F x координируется , четырьмя цистеинами ; по два цистеина обеспечиваются PsaA и PsaB. Два цистеина в каждом расположены проксимально и расположены в петле между девятым и десятым трансмембранными сегментами. лейциновой молнии. мотив Кажется, присутствует [ 5 ] ниже цистеинов и может способствовать димеризации PsaA/PsaB. Концевые акцепторы электронов F A и F B , также железо-серные кластеры [4Fe-4S], расположены в белке массой 9 кДа, называемом PsaC, который связывается с ядром PsaA/PsaB рядом с F X . [ 6 ] [ 7 ]
| Белковые субъединицы | Описание |
|---|---|
| ПсА | Родственные крупные трансмембранные белки, участвующие в связывании P700, A0, A1 и Fx. Входит в семейство белков реакционного центра фотосинтеза . |
| ПСАБ | |
| ПсаК | Железо-серный центр; апопротеин для F a и F b |
| Псад | Необходим для сборки, помогает связывать ферредоксин. ИнтерПро : IPR003685 |
| ПсаЭ | ИнтерПро : IPR003375 |
| ПсаИ | Может стабилизировать PsaL. Стабилизирует связывание светособирающего комплекса II. [ 9 ] ИнтерПро : IPR001302 |
| PsaJ | ИнтерПро : IPR002615 |
| ПсаК | ИнтерПро : IPR035982 |
| ПсаЛ | ИнтерПро : IPR036592 |
| PsaM | ИнтерПро : IPR010010 |
| PsaX | ИнтерПро : IPR012986 |
| цитохрома b 6 f комплекс | Растворимый белок |
| Фа | От ПсаК; В цепи переноса электронов (ETC) |
| Ф б | От ПсаК; В ETC |
| Ф х | От ПсаАБ; В ETC |
| Ферредоксин | Переносчик электронов в внеземных цивилизациях |
| Пластоцианин | Растворимый белок |
| Липиды | Описание |
| МГДГ II | Моногалактозилдиглицеридный липид |
| PG I | Фосфатидилглицерин фосфолипид |
| PG III | Фосфатидилглицерин фосфолипид |
| PG IV | Фосфатидилглицерин фосфолипид |
| Пигменты | Описание |
| Хлорофилл а | 90 молекул пигмента в антенной системе |
| Хлорофилл а | 5 молекул пигмента в ETC |
| Хлорофилл а 0 | Ранний акцептор электронов модифицированного хлорофилла в внеземных цивилизациях |
| Хлорофилл а ' | 1 молекула пигмента в ETC |
| β-каротины | 22 молекулы каротиноидного пигмента |
| Коэнзимы и кофакторы | Описание |
| К К -А | Ранний акцептор электронов витамин К 1 филлохинон в внеземных цивилизациях |
| К К -Б | Ранний акцептор электронов витамин К 1 филлохинон в внеземных цивилизациях |
| ФНР | Ферредоксин- НАДФ + фермент оксидоредуктаза |
| Что 2+ |
кальция Ион |
| мг 2+ |
магния Ион |
Фотон
[ редактировать ]Фотовозбуждение молекул пигмента в антенном комплексе вызывает перенос электронов и энергии. [ 10 ]
Антенный комплекс
[ редактировать ]Антенный комплекс состоит из молекул хлорофилла и каротиноидов, закрепленных на двух белках. [ 11 ] Эти молекулы пигмента передают резонансную энергию фотонов, когда они фотовозбуждаются. Молекулы антенны могут поглощать все длины волн света в видимом спектре . [ 12 ] Количество этих молекул пигмента варьируется от организма к организму. Например, цианобактерия Synechococcus elongatus ( Thermynechococcus elongatus ) имеет около 100 хлорофиллов и 20 каротиноидов, тогда как хлоропласты шпината содержат около 200 хлорофиллов и 50 каротиноидов. [ 12 ] [ 3 ] Внутри антенного комплекса PSI расположены молекулы хлорофилла, называемые P700 реакционными центрами . Энергия, передаваемая молекулами антенны, направляется в реакционный центр. В одном P700 может быть от 120 до 25 молекул хлорофилла. [ 13 ]
Реакционный центр P700
[ редактировать ]Реакционный центр P700 состоит из модифицированного хлорофилла а, который лучше всего поглощает свет с длиной волны 700 нм . [ 14 ] P700 получает энергию от молекул антенны и использует энергию каждого фотона, чтобы поднять электрон на более высокий энергетический уровень (P700*). Эти электроны перемещаются парами в процессе окисления/восстановления от P700* к акцепторам электронов, оставляя после себя P700. + . Пара P700* - P700 + имеет электрический потенциал около -1,2 вольта . Реакционный центр состоит из двух молекул хлорофилла и поэтому называется димером . [ 11 ] Считается, что димер состоит из одного хлорофилла на молекулу и одного хлорофилла на молекулу. Однако если P700 образует комплекс с другими молекулами антенны, он уже не может быть димером. [ 13 ]
Модифицированный хлорофилл А 0 и А 1
[ редактировать ]Две модифицированные молекулы хлорофилла являются ранними акцепторами электронов в PSI. Они присутствуют по одному на стороне PsaA/PsaB, образуя две ветви, которые электроны могут пройти, чтобы достичь F x . А 0 принимает электроны от Р700*, передает их А 1 той же стороны, который затем передает электрон хинону на той же стороне. Похоже, что разные виды имеют разные предпочтения в отношении каждой из ветвей A/B. [ 15 ]
Филлохинон
[ редактировать ]Филлохинон витамином , иногда называемый К1 , [ 16 ] является следующим ранним акцептором электронов в PSI. Он окисляет A 1 , чтобы получить электрон, и, в свою очередь, повторно окисляется F x , от которого электрон передается F b и F a . [ 16 ] [ 17 ] Уменьшение Fx , по-видимому, является шагом, ограничивающим скорость. [ 15 ]
Железо-серный комплекс
[ редактировать ]три белковых железо-серных В PSI обнаружено реакционных центра. Обозначенные Fx , Fa и Fb , они служат электронными реле. [ 18 ] F a и F b связаны с белковыми субъединицами комплекса PSI, а F x связан с комплексом PSI. [ 18 ] Различные эксперименты показали некоторое несоответствие между теориями ориентации железо-серного кофактора и порядка действий. [ 18 ] В одной модели F x передает электрон F a , который передает его F b для достижения ферредоксина. [ 15 ]
Ферредоксин
[ редактировать ]Ферредоксин (Fd) — растворимый белок, способствующий восстановлению НАДФ. +
к НАДФН. [ 19 ] Fd перемещается, перенося электрон либо к одинокому тилакоиду, либо к ферменту , восстанавливающему НАДФ. +
. [ 19 ] Мембраны тилакоидов имеют по одному сайту связывания для каждой функции Fd. [ 19 ] Основная функция Fd – перенос электрона от железосерного комплекса к ферменту ферредоксин– НАДФ. +
редуктаза . [ 19 ]
Ферредоксин – НАДФ +
редуктаза (ФНР)
[ редактировать ] Этот фермент переносит электрон от восстановленного ферредоксина к НАДФ. +
для завершения восстановления до НАДФН. [ 20 ] FNR также может принимать электрон от НАДФН, связываясь с ним. [ 20 ]
Пластоцианин
[ редактировать ]Пластоцианин является переносчиком электронов, который переносит электрон от цитохрома b6f к кофактору P700 PSI в его ионизированном состоянии P700. + . [ 10 ] [ 21 ]
Белковый домен Ycf4
[ редактировать ]Белковый домен Ycf4, обнаруженный на тилакоидной мембране, жизненно важен для фотосистемы I. Этот трансмембранный белок тилакоида помогает собирать компоненты фотосистемы I. Без него фотосинтез был бы неэффективным. [ 22 ]
Эволюция
[ редактировать ]Молекулярные данные показывают, что PSI, вероятно, развился из фотосистем зеленых серобактерий . Фотосистемы зеленых серобактерий и фотосистем цианобактерий , водорослей и высших растений не одинаковы, но имеют много аналогичных функций и сходных структур. Три основные особенности разных фотосистем схожи. [ 23 ] Во-первых, окислительно-восстановительный потенциал достаточно отрицателен, чтобы восстановить ферредоксин. [ 23 ] Далее к электронопринимающим реакционным центрам относятся железо-серные белки. [ 23 ] Наконец, окислительно-восстановительные центры в комплексах обеих фотосистем построены на димере белковой субъединицы. [ 23 ] Фотосистема зеленых серобактерий содержит даже все те же кофакторы электрон-транспортной цепи PSI. [ 23 ] Количество и степень сходства между двумя фотосистемами убедительно указывают на то, что PSI и аналогичная фотосистема зеленых серных бактерий произошли от общей предковой фотосистемы.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Гольбек Дж. Х. (1987). «Структура, функции и организация комплекса реакционного центра Фотосистема I». Biochimica et Biophysical Acta (BBA) - Обзоры по биоэнергетике . 895 (3): 167–204. дои : 10.1016/s0304-4173(87)80002-2 . ПМИД 3333014 .
- ^ Ямори В., Шиканай Т. (апрель 2016 г.). «Физиологические функции циклического транспорта электронов вокруг фотосистемы I в поддержании фотосинтеза и роста растений» . Ежегодный обзор биологии растений . 67 : 81–106. doi : 10.1146/annurev-arplant-043015-112002 . ПМИД 26927905 .
- ^ Jump up to: а б Нельсон Н., Yocum CF (2006). «Строение и функции фотосистем I и II». Ежегодный обзор биологии растений . 57 : 521–65. doi : 10.1146/annurev.arplant.57.032905.105350 . ПМИД 16669773 .
- ^ Jump up to: а б с Фромм П., Матис П. (2004). «Раскрытие фотосистемы I реакционного центра: история, или сумма многих усилий». Исследования фотосинтеза . 80 (1–3): 109–24. Бибкод : 2004PhoRe..80..109F . doi : 10.1023/B:PRES.0000030657.88242.e1 . ПМИД 16328814 . S2CID 13832448 .
- ^ Уэббер А.Н., Малкин Р. (май 1990 г.). «Белки реакционного центра фотосистемы I содержат мотивы лейциновой молнии. Предполагаемая роль в образовании димеров». Письма ФЭБС . 264 (1): 1–4. Бибкод : 1990FEBSL.264....1W . дои : 10.1016/0014-5793(90)80749-9 . ПМИД 2186925 . S2CID 42294700 .
- ^ Джаганнатан Б., Гольбек Дж. Х. (апрель 2009 г.). «Нарушение биологической симметрии мембранных белков: асимметричная ориентация PsaC на симметричном ядре псевдо-C2 Фотосистемы I» . Клеточные и молекулярные науки о жизни . 66 (7): 1257–70. дои : 10.1007/s00018-009-8673-x . ПМЦ 11131447 . ПМИД 19132290 . S2CID 32418758 .
- ^ Джаганнатан Б., Гольбек Дж. Х. (июнь 2009 г.). «Понимание интерфейса связывания между PsaC и гетеродимером PsaA/PsaB в фотосистеме I». Биохимия . 48 (23): 5405–16. дои : 10.1021/bi900243f . ПМИД 19432395 .
- ^ Сенгер В., Джордан П., Краусс Н. (апрель 2002 г.). «Сборка белковых субъединиц и кофакторов в фотосистеме I». Современное мнение в области структурной биологии . 12 (2): 244–54. дои : 10.1016/S0959-440X(02)00317-2 . ПМИД 11959504 .
- ^ Плёхингер, Магдалена; Тораби, Салар; Рантала, Марджаана; Тикканен, Микко; Суорса, Марьяана; Йенсен, Пол-Эрик; Аро, Ева Мари; Мойрер, Йорг (сентябрь 2016 г.). «Низкомолекулярный белок PsaI стабилизирует место стыковки светособирающего комплекса II фотосистемы I» . Физиология растений . 172 (1): 450–463. дои : 10.1104/стр.16.00647 . ПМК 5074619 . ПМИД 27406169 .
- ^ Jump up to: а б Рэйвен П.Х., Эверт Р.Ф., Эйххорн С.Е. (2005). «Фотосинтез, свет и жизнь». Биология растений (7-е изд.). Нью-Йорк: WH Freeman. стр. 121–127 . ISBN 978-0-7167-1007-3 .
- ^ Jump up to: а б Зейгер Э., Таиз Л. (2006). «Гл. 7: Тема 7.8: Фотосистема I» . Физиология растений (4-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 0-87893-856-7 . [ постоянная мертвая ссылка ]
- ^ Jump up to: а б «Процесс фотосинтеза» . Архивировано из оригинала 19 февраля 2009 г.
- ^ Jump up to: а б Шубин В.В., Карапетян Н.В., Красновский А.А. (январь 1986 г.). «Молекулярное устройство пигментно-белкового комплекса фотосистемы 1». Исследования фотосинтеза . 9 (1–2): 3–12. Бибкод : 1986PhoRe...9....3S . дои : 10.1007/BF00029726 . ПМИД 24442279 . S2CID 26158482 .
- ^ Резерфорд А.В., Хиткот П. (декабрь 1985 г.). «Первичная фотохимия в фотосистеме-I». Исследования фотосинтеза . 6 (4): 295–316. Бибкод : 1985PhoRe...6..295R . дои : 10.1007/BF00054105 . ПМИД 24442951 . S2CID 21845584 .
- ^ Jump up to: а б с Гротьоханн, я; Фромм, П. (2013). «Фотосистема I». Энциклопедия биологической химии (Второе изд.). Лондон. стр. 503–507. дои : 10.1016/B978-0-12-378630-2.00287-5 . ISBN 978-0-12-378630-2 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Jump up to: а б Ито С., Иваки М. (1989). «Витамин К 1 (филлохинон) восстанавливает круговорот FeS-центров экстрагированных эфиром частиц PSI шпината» . Письма ФЭБС . 243 (1): 47–52. дои : 10.1016/0014-5793(89)81215-3 . S2CID 84602152 .
- ^ Palace GP, Франке Дж. Э., Уорден Дж. Т. (май 1987 г.). «Является ли филлохинон обязательным переносчиком электронов в фотосистеме I?» . Письма ФЭБС . 215 (1): 58–62. Бибкод : 1987FEBSL.215...58P . дои : 10.1016/0014-5793(87)80113-8 . ПМИД 3552735 . S2CID 42983611 .
- ^ Jump up to: а б с Васильев И.Р., Антонкин М.Л., Гольбек Ю.Х. (октябрь 2001 г.). «Железо-серные кластеры в реакционных центрах I типа». Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1507 (1–3): 139–60. дои : 10.1016/S0005-2728(01)00197-9 . ПМИД 11687212 .
- ^ Jump up to: а б с д Форти Дж., Мария П., Грубас Г. (1985). «Два места взаимодействия ферредоксина с тилакоидами» . Письма ФЭБС . 186 (2): 149–152. Бибкод : 1985FEBSL.186..149F . дои : 10.1016/0014-5793(85)80698-0 . S2CID 83495051 .
- ^ Jump up to: а б Мадос Х., Фернандес Ресио Х., Гомес Морено С., Фернандес В.М. (ноябрь 1998 г.). «Исследование диафоразной реакции ферредоксина– НАДФ». +
Редуктаза электрохимическими методами » (PDF) . Биоэлектрохимия и биоэнергетика . 47 (1): 179–183. doi : 10.1016/S0302-4598(98)00175-5 . - ^ Хоуп А.Б. (январь 2000 г.). «Перенос электронов между цитохромом f, пластоцианином и фотосистемой I: кинетика и механизмы». Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1456 (1): 5–26. дои : 10.1016/S0005-2728(99)00101-2 . ПМИД 10611452 .
- ^ Будро Э., Такахаси Ю., Лемье С., Тюрмель М., Роше Ж.Д. (октябрь 1997 г.). «Открытые рамки считывания хлоропластов ycf3 и ycf4 Chlamydomonas reinhardtii необходимы для накопления комплекса фотосистемы I» . Журнал ЭМБО . 16 (20): 6095–104. дои : 10.1093/emboj/16.20.6095 . ПМЦ 1326293 . ПМИД 9321389 .
- ^ Jump up to: а б с д и Локау В., Ничке В. (1993). «Фотосистема I и ее бактериальные аналоги». Физиология Плантарум . 88 (2): 372–381. дои : 10.1111/j.1399-3054.1993.tb05512.x .