Микробиота
этой статьи Начальный раздел может быть слишком коротким, чтобы адекватно суммировать ключевые моменты . ( октябрь 2018 г. ) |
Микробиота представляет собой ряд микроорганизмов , которые могут быть комменсальными , мутуалистическими или патогенными и обнаруживаться внутри и на всех многоклеточных организмах , включая растения . Микробиота включает бактерии , археи , простейшие , грибы и вирусы . [ 2 ] [ 3 ] и было обнаружено, что они имеют решающее значение для иммунологического, гормонального и метаболического гомеостаза хозяина.
Термин «микробиом» описывает либо коллективные геномы микробов, обитающих в экологической нише , либо сами микробы. [ 4 ] [ 5 ] [ 6 ]
Микробиом и хозяин возникли в ходе эволюции как синергетическая единица эпигенетики и генетических характеристик, иногда все вместе называемая холобионтом . [ 7 ] [ 8 ] Присутствие микробиоты в кишечнике человека и других многоклеточных животных имело решающее значение для понимания совместной эволюции многоклеточных животных и бактерий. [ 9 ] [ 10 ] Микробиота играет ключевую роль в иммунных и метаболических реакциях кишечника через продукт ферментации ( короткоцепочечные жирные кислоты ), ацетат . [ 11 ]
Введение
[ редактировать ]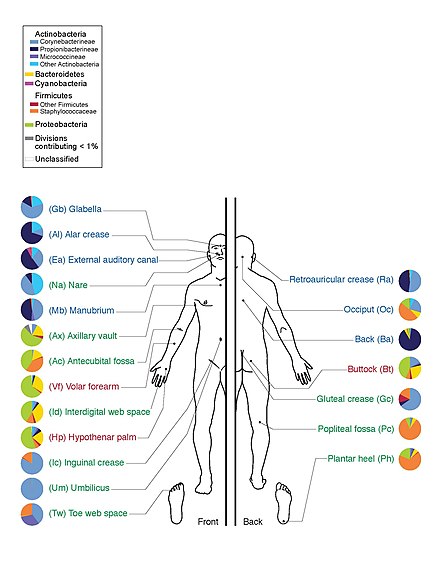
Все растения и животные, от простых форм жизни до человека, живут в тесной связи с микробными организмами. [ 12 ] Несколько достижений способствовали пониманию микробиомов, в том числе:
- способность выполнять геномный анализ и анализ экспрессии генов отдельных клеток и целых микробных сообществ в дисциплинах метагеномика и метатранскриптомика [ 13 ]
- базы данных, доступные исследователям разных дисциплин [ 13 ]
- методы математического анализа, подходящие для сложных наборов данных [ 13 ]
организма Биологи пришли к пониманию того, что микробы составляют важную часть фенотипа , выходящую далеко за рамки случайных симбиотических исследований. [ 13 ]
Типы взаимоотношений микроб-хозяин
[ редактировать ]Комменсализм — концепция, разработанная Пьером-Жозефом ван Бенеденом (1809–1894), бельгийским профессором Лувенского университета в девятнадцатом веке. [ 14 ] занимает центральное место в микробиоме, где микробиота колонизирует хозяина в условиях безвредного сосуществования. Отношения с хозяином называются мутуалистическими , когда организмы выполняют задачи, заведомо полезные для хозяина. [ 15 ] : 700 [ 16 ] паразитический , когда он невыгоден хозяину. Другие авторы определяют ситуацию как мутуалистическую, когда оба получают выгоду, и комменсальную, когда незатронутый хозяин приносит пользу симбионту. [ 17 ] Обмен питательными веществами может быть двунаправленным или однонаправленным, может зависеть от контекста и происходить разными способами. [ 17 ] Микробиота, наличие которой ожидается и которая при нормальных обстоятельствах не вызывает заболевания, считается нормальной флорой или нормальной микробиотой ; [ 15 ] нормальная флора может быть не только безвредной, но и защищать хозяина. [ 18 ]
Приобретение и изменение
[ редактировать ]Первоначальное приобретение микробиоты у животных, от млекопитающих до морских губок , происходит при рождении и может даже происходить через линию зародышевых клеток. У растений процесс колонизации может быть инициирован под землей, в корневой зоне , вокруг прорастающего семени, спермосферы , или исходить из надземных частей, филлосферы и цветочной зоны или антосферы. [ 19 ] Стабильность микробиоты ризосферы на протяжении поколений зависит от типа растения, но еще больше от состава почвы, то есть живой и неживой среды. [ 20 ] Клинически новую микробиоту можно получить посредством трансплантации фекальной микробиоты для лечения таких инфекций, как хроническая инфекция C. difficile . [ 21 ]
Микробиота по хозяину
[ редактировать ]
Люди
[ редактировать ]Микробиота человека включает бактерии , грибы , археи и вирусы. Микроживотные , живущие на теле человека, исключены. Микробиом человека относится к его коллективным геномам . [ 15 ]
Люди колонизированы многими микроорганизмами; традиционная оценка заключалась в том, что у людей в десять раз больше нечеловеческих клеток, чем у человеческих; по более поздним оценкам это соотношение снизилось до 3:1 и даже примерно до 1:1 по количеству (1:350 по массе). [ 22 ] [ 23 ] [ 24 ] [ 25 ] [ 26 ]
На самом деле они настолько малы, что в организме человека насчитывается около 100 триллионов микробиоты. [ 27 ] около 39 триллионов по пересмотренным оценкам, при общей массе всего 0,2 кг в «эталонном» 70-килограммовом человеческом теле. [ 26 ]
Проект «Микробиом человека» секвенировал геном микробиоты человека, уделяя особое внимание микробиоте, которая обычно населяет кожу, рот, нос, пищеварительный тракт и влагалище. [ 15 ] В 2012 году компания достигла важной вехи, когда опубликовала первые результаты. [ 28 ]
Нечеловеческие животные
[ редактировать ]- На коже земноводных имеется микробиота. [ 29 ] Некоторые виды способны переносить грибок Batrachochytrium dendrobatidis , который у других может вызывать смертельную инфекцию хитридиомикоза, в зависимости от их микробиома, сопротивляясь колонизации патогенов или ингибируя их рост с помощью антимикробных пептидов кожи. [ 30 ]
- Новорожденные сумчатые рождаются с гистологически незрелыми иммунными тканями и неспособны создать собственную специфическую иммунную защиту. Поэтому они во многом зависят от иммунной системы матери. [ 31 ] и молоко [ 32 ] для их защиты. У большинства сумчатых есть мешочки, и их собственная микробиота меняется на протяжении репродуктивных стадий: течки, рождения/течки и постэструса. [ 33 ] В некоторых выделениях мешков и кожи были обнаружены антимикробные пептиды, которые, предположительно, поддерживают молодняк в это уязвимое время.
- У млекопитающих травоядные животные, такие как крупный рогатый скот, зависят от микробиома рубца , который преобразует целлюлозу в белки, короткоцепочечные жирные кислоты и газы. Методы культивирования не могут предоставить информацию обо всех присутствующих микроорганизмах. Сравнительные метагеномные исследования дали неожиданный результат: отдельные виды крупного рогатого скота обладают заметно разными структурами сообщества, прогнозируемым фенотипом и метаболическим потенциалом. [ 34 ] даже несмотря на то, что их кормили одинаковой диетой, содержали вместе и, очевидно, функционально идентично использовали ресурсы стенок растительных клеток.
- Мыши стали наиболее изученными млекопитающими в отношении их микробиомов. Микробиота кишечника изучалась в связи с аллергическими заболеваниями дыхательных путей, ожирением, желудочно-кишечными заболеваниями и диабетом. Перинатальное изменение микробиоты из-за приема низких доз антибиотиков может иметь долгосрочные последствия для будущей восприимчивости к аллергическим заболеваниям дыхательных путей. Частота встречаемости определенных подмножеств микробов связана с тяжестью заболевания. Присутствие специфических микробов в раннем послеродовом периоде определяет будущие иммунные реакции. [ 35 ] [ 36 ] У гнотобиотических мышей было обнаружено, что определенные кишечные бактерии передают определенный фенотип мышам-реципиентам, свободным от микробов, что способствует накоплению регуляторных Т-клеток толстой кишки, а также штаммы, которые модулируют ожирение у мышей и концентрации метаболитов в слепой кишке. [ 37 ] Этот комбинаторный подход позволяет понять вклад микробов в биологию человека на системном уровне. [ 38 ] Но также другие слизистые ткани, такие как легкие и влагалище, изучались в связи с такими заболеваниями, как астма, аллергия и вагиноз. [ 39 ]
- У насекомых есть свои микробиомы. Например, муравьи-листорезы образуют огромные подземные колонии, собирающие сотни килограммов листьев каждый год, и не могут напрямую переваривать целлюлозу, содержащуюся в листьях. Они поддерживают грибные сады как основной источник пищи колонии. Хотя сам гриб не переваривает целлюлозу, это делает микробное сообщество, содержащее множество бактерий. Анализ генома микробной популяции выявил множество генов, участвующих в переваривании целлюлозы. Прогнозируемый профиль ферментов, расщепляющих углеводы, в этом микробиоме аналогичен профилю бычьего рубца, но видовой состав почти полностью отличается. [ 40 ] Микробиота кишечника плодовой мухи может влиять на внешний вид ее кишечника, влияя на скорость обновления эпителия, расстояние между клетками и состав различных типов клеток в эпителии. [ 41 ] Когда моль Spodoptera exigua заражается бакуловирусом, гены, связанные с иммунитетом, подавляются, и количество микробиоты ее кишечника увеличивается. [ 42 ] В кишечнике двукрылых энтероэндокринные клетки воспринимают метаболиты, полученные из микробиоты кишечника, и координируют антибактериальные, механические и метаболические ветви врожденного иммунного ответа кишечника хозяина на комменсальную микробиоту. [ 43 ]
- У рыб есть свои собственные микробиомы, в том числе у недолговечного вида Nothobranchius Furzeri (бирюзовая рыба-киллифиш). Перенос микробиоты кишечника от молодого убийцы к убийце среднего возраста значительно продлевает продолжительность жизни убийцы среднего возраста. [ 44 ]
Растения
[ редактировать ]
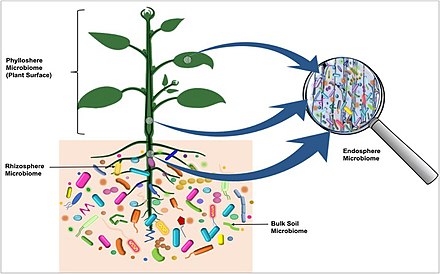
Недавно было обнаружено, что микробиом растения происходит из семян. [ 46 ] Микроорганизмы, передающиеся через семена, мигрируют в развивающийся сеянец определенным путем, при котором одни сообщества перемещаются к листьям, а другие - к корням. [ 46 ] На диаграмме справа микробиота, колонизирующая ризосферу , проникающая в корни и колонизирующая следующее поколение клубней через столоны , визуализирована красным цветом. Бактерии, присутствующие в материнском клубне , проходящие через столоны и мигрирующие в растение, а также в клубни следующего поколения, показаны синим цветом. [ 45 ]
- Почва является основным резервуаром бактерий, колонизирующих клубни картофеля.
- Бактерии рекрутируются из почвы более или менее независимо от сорта картофеля.
- Бактерии могут колонизировать клубни преимущественно изнутри растений через столоны.
- Бактериальная микробиота клубней картофеля состоит из бактерий, передающихся от одного поколения клубней к другому, и бактерий, рекрутированных из почвы, колонизирующих растения картофеля через корень. [ 45 ]

Растения являются привлекательными хозяевами для микроорганизмов, поскольку они предоставляют разнообразные питательные вещества. Микроорганизмы на растениях могут быть эпифитами (находятся на растениях) или эндофитами (находятся внутри тканей растения). [ 47 ] [ 48 ] Оомицеты и грибы в результате конвергентной эволюции развили сходную морфологию и заняли схожие экологические ниши. У них развиваются гифы — нитевидные структуры, проникающие в клетку-хозяина. В мутуалистических ситуациях растение часто обменивает гексозные сахара на неорганический фосфат грибного симбионта. Предполагается, что такие очень древние ассоциации помогли растениям, когда они впервые колонизировали землю. [ 17 ] [ 49 ] Бактерии, стимулирующие рост растений (PGPB), обеспечивают растению важные функции, такие как фиксация азота , солюбилизация минералов, таких как фосфор, синтез растительных гормонов , прямое усиление поглощения минералов и защита от патогенов. [ 50 ] [ 51 ] PGPB могут защищать растения от патогенов, конкурируя с патогеном за экологическую нишу или субстрат, продуцируя ингибирующие аллелохимические вещества или вызывая системную устойчивость растений-хозяев к патогену. [ 19 ]
Исследовать
[ редактировать ]Симбиотические отношения между хозяином и его микробиотой находятся в стадии лабораторных исследований на предмет того, как они могут формировать иммунную систему млекопитающих. [ 52 ] [ 53 ] У многих животных иммунная система и микробиота могут вступать в «перекрестный диалог», обмениваясь химическими сигналами, что может позволить микробиоте влиять на иммунную реактивность и нацеливание. [ 54 ] Бактерии могут передаваться от матери к ребенку при прямом контакте и после рождения . [ 55 ] По мере формирования микробиома младенца комменсальные бактерии быстро заселяют кишечник, вызывая ряд иммунных реакций и «программируя» иммунную систему на долгосрочные эффекты. [ 54 ] Бактерии способны стимулировать лимфоидную ткань, связанную со слизистой оболочкой кишечника, что позволяет ткани вырабатывать антитела к патогенам, которые могут попасть в кишечник. [ 54 ]
Микробиом человека может играть роль в активации toll-подобных рецепторов в кишечнике — типа рецепторов распознавания образов, которые клетки-хозяева используют для распознавания опасностей и устранения повреждений. Патогены могут влиять на это сосуществование, приводя к иммунной дисрегуляции, включая восприимчивость к заболеваниям, механизмам воспаления , иммунной толерантности и аутоиммунным заболеваниям . [ 56 ] [ 57 ]
Коэволюция микробиоты
[ редактировать ]
Организмы развиваются внутри экосистем так, что изменение одного организма влияет на изменение других. Хологеномная теория эволюции предполагает, что объектом естественного отбора является не отдельный организм, а организм вместе со связанными с ним организмами, включая его микробные сообщества.
Коралловые рифы . Теория гологенома возникла в результате исследований коралловых рифов. [ 58 ] Коралловые рифы являются крупнейшими структурами, созданными живыми организмами, и содержат многочисленные и очень сложные микробные сообщества. За последние несколько десятилетий произошло значительное сокращение популяций кораллов. Изменение климата , загрязнение воды и чрезмерный вылов рыбы — три фактора стресса, которые, как было описано, приводят к восприимчивости к болезням. Описано более двадцати различных заболеваний кораллов, но лишь у немногих из них были выделены и охарактеризованы возбудители. Обесцвечивание кораллов является наиболее серьезным из этих заболеваний. В Средиземном море обесцвечивание Oculina patagonica было впервые описано в 1994 году и вскоре было установлено, что оно вызвано заражением Vibrio shiloi . С 1994 по 2002 год бактериальное обесцвечивание O. patagonica происходило каждое лето в восточном Средиземноморье. Удивительно, но после 2003 г. O. patagonica в восточном Средиземноморье стала устойчивой к инфекции V. shiloi , хотя другие заболевания по-прежнему вызывают обесцвечивание. Удивление связано с тем, что кораллы долгоживущие, их продолжительность жизни составляет порядка десятилетий. [ 59 ] и не имеют адаптивной иммунной системы . [ нужна ссылка ] Их врожденная иммунная система не вырабатывает антитела, и, по-видимому, они не смогут реагировать на новые вызовы, кроме как в эволюционных временных масштабах. [ нужна ссылка ]
Загадка того, как кораллам удалось приобрести устойчивость к определенному патогену, привела к предположению в 2007 году о том, что между кораллами и их симбиотическими микробными сообществами существуют динамические отношения. Считается, что, изменяя свой состав, холобионт может адаптироваться к изменяющимся условиям окружающей среды гораздо быстрее, чем с помощью одних лишь генетических мутаций и отбора. Экстраполяция этой гипотезы на другие организмы, включая высшие растения и животных, привела к предложению гологеномной теории эволюции. [ 58 ]
По состоянию на 2007 год [update] теория гологенома все еще обсуждалась. [ 60 ] Серьезной критикой было утверждение о том, что V. shiloi был ошибочно идентифицирован как возбудитель обесцвечивания кораллов и что его присутствие в обесцвеченном O. patagonica было просто результатом оппортунистической колонизации. [ 61 ] Если это правда, то основное наблюдение, ведущее к теории, было бы неверным. Теория приобрела значительную популярность как способ объяснения быстрых изменений в адаптации, которые иначе нельзя объяснить традиционными механизмами естественного отбора. В рамках теории хологенома холобионт стал не только основной единицей естественного отбора, но и результатом другого этапа интеграции, который также наблюдается на клеточном ( симбиогенез , эндосимбиоз ) и геномном уровнях. [ 7 ]
Методы исследования
[ редактировать ]Целевое секвенирование ампликонов
[ редактировать ]| Часть серии о |
| Микробиомы |
|---|
 |
Целевое секвенирование ампликонов основано на некоторых ожиданиях относительно состава изучаемого сообщества. При целевом секвенировании ампликонов для секвенирования выбирают филогенетически информативный маркер. Такой маркер должен присутствовать в идеале у всех ожидаемых организмов. Он также должен развиваться таким образом, чтобы быть достаточно консервативным, чтобы праймеры могли нацеливаться на гены из широкого спектра организмов, при этом развиваясь достаточно быстро, чтобы обеспечить более точное разрешение на таксономическом уровне. Общим маркером для исследований микробиома человека является ген бактериальной 16S рРНК ( т.е. «16S рДНК», последовательность ДНК, которая кодирует молекулу рибосомальной РНК). [ 62 ] Поскольку рибосомы присутствуют во всех живых организмах, использование 16S рДНК позволяет амплифицировать ДНК гораздо большего числа организмов, чем если бы использовался другой маркер. Ген 16S рРНК содержит как медленно развивающиеся области, так и 9 быстро развивающихся областей, также известных как гипервариабельные области (HVR); [ 63 ] первый можно использовать для разработки широких праймеров, а второй позволяет проводить более тонкие таксономические различия. Однако разрешение на видовом уровне с использованием 16S рДНК обычно невозможно. Выбор праймера является важным шагом, поскольку все, что не может быть нацелено на праймер, не будет амплифицировано и, следовательно, не будет обнаружено. Более того, можно выбрать разные наборы праймеров для амплификации разных HVR в гене или их пар. Соответствующий выбор HVR для амплификации должен быть сделан в соответствии с представляющими интерес таксономическими группами, поскольку было показано, что разные целевые регионы влияют на таксономическую классификацию. [ 64 ]
Целевые исследования эукариотических и вирусных сообществ ограничены. [ 65 ] и при условии исключения ДНК хозяина из амплификации и снижения эукариотической и вирусной биомассы в микробиоме человека. [ 66 ]
После секвенирования ампликонов молекулярно-филогенетические методы используются для определения состава микробного сообщества. Это можно сделать с помощью методологий кластеризации , объединяя ампликоны в операционные таксономические единицы (OTU); или, альтернативно, с помощью методологий шумоподавления, идентифицируя варианты последовательностей ампликонов (ASV).
Затем между последовательностями делаются выводы о филогенетических отношениях. Из-за сложности данных между образцами микробиома обычно определяются меры расстояния, такие как расстояния UniFrac , а на матрицах расстояний применяются последующие многомерные методы. Важным моментом является то, что масштаб данных обширен, и необходимо использовать дополнительные подходы для выявления закономерностей на основе доступной информации. Инструменты, используемые для анализа данных, включают VAMPS, [ 67 ] ЦЕНА , [ 68 ] мать [ 69 ] и ДАДА2 [ 70 ] или УНОАЗЕ3 [ 71 ] для шумоподавления.
Метагеномное секвенирование
[ редактировать ]Метагеномика также широко используется для изучения микробных сообществ. [ 72 ] [ 73 ] [ 74 ] При метагеномном секвенировании ДНК извлекается непосредственно из образцов окружающей среды нецелевым образом с целью получения объективного образца всех генов всех членов сообщества. использовалось секвенирование по Сэнгеру или пиросеквенирование . В недавних исследованиях для восстановления последовательностей считываний [ 75 ] Чтения затем могут быть собраны в контиги . Чтобы определить филогенетическую идентичность последовательности, ее сравнивают с доступными полногеномными последовательностями с использованием таких методов, как BLAST . Одним из недостатков этого подхода является то, что многие члены микробных сообществ не имеют репрезентативного секвенированного генома, но это применимо и к секвенированию ампликона 16S рРНК и является фундаментальной проблемой. [ 62 ] С помощью дробового секвенирования эту проблему можно решить, если обеспечить высокий охват (50-100 раз) неизвестного генома, эффективно выполнив сборку генома de novo . Как только станет доступен полный геном неизвестного организма, его можно будет сравнить филогенетически и поставить организм на свое место на древе жизни , создав новые таксоны . Новый подход заключается в сочетании дробового секвенирования с данными бесконтактного лигирования ( Hi-C ) для сбора полных микробных геномов без культивирования. [ 76 ]
Несмотря на то, что метагеномика ограничена доступностью эталонных последовательностей, одним из существенных преимуществ метагеномики перед целевым секвенированием ампликонов является то, что данные метагеномики могут пролить свет на функциональный потенциал ДНК сообщества. [ 77 ] [ 78 ] Целевые генные исследования не могут этого сделать, поскольку они лишь выявляют филогенетические отношения между одним и тем же геном из разных организмов. Функциональный анализ проводится путем сравнения восстановленных последовательностей с базами данных метагеномных аннотаций, такими как KEGG . Метаболические пути, в которых участвуют эти гены, можно затем предсказать с помощью таких инструментов, как MG-RAST. [ 79 ] КАМЕРА [ 80 ] и ИМГ/М . [ 81 ]
Подходы на основе РНК и белков
[ редактировать ]Метатранскриптомические исследования были проведены для изучения экспрессии генов микробных сообществ с помощью таких методов, как пиросеквенирование извлеченной РНК. [ 82 ] Исследования, основанные на структуре, также выявили некодирующие РНК (нкРНК), такие как рибозимы из микробиоты. [ 83 ] Метапротеомика — это подход, изучающий белки, экспрессируемые микробиотой, что дает представление о ее функциональном потенциале. [ 84 ]
Проекты
[ редактировать ]Проект «Микробиом человека», США запущенный в 2008 году, представлял собой инициативу Национальных институтов здравоохранения по выявлению и характеристике микроорганизмов, обнаруженных как у здоровых, так и у больных людей. [ 85 ] Пятилетний проект, который лучше всего можно охарактеризовать как технико-экономическое обоснование с бюджетом в 115 миллионов долларов, проверял, как изменения в микробиоме человека связаны со здоровьем или болезнями человека. [ 85 ]
Проект «Микробиом Земли» (EMP) — это инициатива по сбору природных образцов и анализу микробного сообщества по всему миру. Микробы очень многочисленны, разнообразны и играют важную роль в экологической системе. Тем не менее, по состоянию на 2010 г. [update], было подсчитано, что в результате общих глобальных усилий по секвенированию ДНК окружающей среды было получено менее 1 процента от общего количества ДНК, обнаруженной в литре морской воды или грамме почвы, [ 86 ] а конкретные взаимодействия между микробами в значительной степени неизвестны. Целью EMP является обработка до 200 000 образцов в различных биомах, создание полной базы данных микробов на Земле для характеристики окружающей среды и экосистем по микробному составу и взаимодействию. Используя эти данные, можно предложить и проверить новые экологические и эволюционные теории. [ 87 ]
Микробиота кишечника и диабет 2 типа
[ редактировать ]Микробиота кишечника очень важна для здоровья хозяина, поскольку она играет роль в деградации неперевариваемых полисахаридов (ферментация резистентного крахмала, олигосахаридов, инулина), укреплении целостности кишечника или формировании кишечного эпителия, сборе энергии, защите от патогенов и регулировании организма хозяина. иммунитет. [ 88 ] [ 89 ]
Несколько исследований показали, что бактериальный состав кишечника у пациентов с диабетом изменяется с увеличением уровней Lactobacillus gasseri , Streptococcus mutans и Clostridiales, а также с уменьшением количества бактерий, продуцирующих бутират, таких как Roseburia кишечная и Faecalibacterium prausnitzii. [ 90 ] [ 91 ] Это изменение связано со многими факторами, такими как злоупотребление антибиотиками, диета и возраст .
Снижение выработки бутирата связано с дефектами кишечной проницаемости, что может привести к эндотоксемии , то есть повышению уровня циркулирующих липополисахаридов из стенки грамотрицательных бактериальных клеток. Установлено, что эндотоксемия связана с развитием инсулинорезистентности. [ 90 ]
Кроме того, выработка бутирата влияет на уровень серотонина. [ 90 ] Повышенный уровень серотонина способствует ожирению, которое, как известно, является фактором риска развития диабета.
Развитие кишечной микробиоты и антибиотики
[ редактировать ]Колонизация микробиоты кишечника человека может начаться еще до рождения. [ 92 ] В окружающей среде существует множество факторов, влияющих на развитие микробиоты, причем способ рождения является одним из наиболее влиятельных. [ 93 ]
Еще одним фактором, который, как было замечено, вызывает огромные изменения в микробиоте кишечника, особенно у детей, является использование антибиотиков, что связано с такими проблемами со здоровьем, как более высокий ИМТ, [ 94 ] [ 95 ] и, кроме того, повышенный риск метаболических заболеваний, таких как ожирение. [ 96 ] У детей грудного возраста наблюдалось, что амоксициллин и макролиды вызывают значительные изменения в микробиоте кишечника, характеризующиеся изменением классов бактерий Bifidobacteria, Enterobacteria и Clostridia. [ 97 ] Однократный курс антибиотиков у взрослых вызывает изменения как бактериальной, так и грибковой микробиоты, причем в грибковых сообществах происходят еще более стойкие изменения. [ 98 ] Бактерии и грибы живут вместе в кишечнике, и, скорее всего, существует конкуренция за источники питательных веществ. [ 99 ] [ 100 ] Зеебиндер и др . обнаружили, что комменсальные бактерии в кишечнике регулируют рост и патогенность Candida albicans с помощью своих метаболитов, особенно пропионата, уксусной кислоты и 5-додеценоата. [ 98 ] Кандида ранее была связана с ВЗК. [ 101 ] Кроме того, наблюдалось увеличение этого показателя у лиц, не ответивших на биологический препарат инфликсимаб, назначаемый пациентам с ВЗК с тяжелым течением ВЗК. [ 102 ] Пропионат и уксусная кислота представляют собой короткоцепочечные жирные кислоты (КЦЖК), которые, как было замечено, полезны для здоровья микробиоты кишечника. [ 103 ] [ 104 ] [ 105 ] Когда антибиотики влияют на рост бактерий в кишечнике, может произойти чрезмерный рост определенных грибов, которые могут быть патогенными, если их не регулировать. [ 98 ]
Проблемы конфиденциальности
[ редактировать ]ДНК микробов, населяющая человеческое тело человека, может однозначно идентифицировать человека. Конфиденциальность человека может быть поставлена под угрозу, если он анонимно пожертвует данные ДНК микроба. Их состояние здоровья и личность могут быть раскрыты. [ 106 ] [ 107 ] [ 108 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Дастогер, К.М., Тумпа, Ф.Х., Султана, А., Актер, М.А. и Чакраборти, А. (2020) «Микробиом растений - отчет о факторах, которые формируют состав и разнообразие сообщества». Текущая биология растений , 23 : 100161. дои : 10.1016/j.cpb.2020.100161 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Де Сорди, Луиза; Лоренсу, Марта; Дебарбье, Лоран (2019). «Внутренняя битва: взаимодействие бактериофагов и бактерий в желудочно-кишечном тракте» . Клетка-хозяин и микроб . 25 (2): 210–218. дои : 10.1016/j.chom.2019.01.018 . ПМИД 30763535 . S2CID 73455329 .
- ^ Петерсон, Дж; Гаргес, С; и др. (2009). «Проект NIH по микробиому человека» . Геномные исследования . 19 (12). Рабочая группа NIH HMP: 2317–2323. дои : 10.1101/гр.096651.109 . ПМЦ 2792171 . ПМИД 19819907 .
- ^ Бэкхед, Ф.; Лей, RE; Зонненбург, JL; Петерсон, Д.А.; Гордон, Дж.И. (2005). «Хозяин-бактериальный мутуализм в кишечнике человека» . Наука . 307 (5717): 1915–1920. Бибкод : 2005Sci...307.1915B . дои : 10.1126/science.1104816 . ПМИД 15790844 . S2CID 6332272 .
- ^ Тернбо, П.Дж.; Лей, RE; Хамади, М.; Фрейзер-Лиггетт, CM; Найт, Р.; Гордон, Дж.И. (2007). «Проект микробиома человека» . Природа . 449 (7164): 804–810. Бибкод : 2007Natur.449..804T . дои : 10.1038/nature06244 . ПМЦ 3709439 . ПМИД 17943116 .
- ^ Лей, RE; Петерсон, Д.А.; Гордон, Дж.И. (2006). «Экологические и эволюционные силы, формирующие микробное разнообразие в кишечнике человека» . Клетка . 124 (4): 837–848. дои : 10.1016/j.cell.2006.02.017 . ПМИД 16497592 . S2CID 17203181 .
- ^ Jump up to: а б Сальвуччи, Э. (2016). «Микробиом, холобионт и сеть жизни». Критические обзоры по микробиологии . 42 (3): 485–494. дои : 10.3109/1040841X.2014.962478 . hdl : 11336/33456 . ПМИД 25430522 . S2CID 30677140 .
- ^ Герреро, Р.; Маргулис, Линн ; Берланга, М. (2013). «Симбиогенез: холобионт как единица эволюции». Международная микробиология . 16 (3): 133–143. дои : 10.2436/20.1501.01.188 . ПМИД 24568029 .
- ^ Давенпорт, Эмили Р. и др. «Микробиом человека в эволюции». БМК Биология . том. 15,1 127. 27 дек. 2017 г., два : 10.1186/s12915-017-0454-7
- ^ «Эволюция кишечной флоры человека». Эндрю Х. Мёллер, Инъинг Ли, Эйтель Мпуди Нголе, Стив Ахука-Мундеке, Элизабет В. Лонсдорф, Энн Э. Пьюзи, Мартин Питерс, Беатрис Х. Хан, Ховард Охман. Труды Национальной академии наук . ноябрь 2014 г., 111 (46) 16431–16435; два : 10.1073/pnas.1419136111
- ^ Джагдер, Бат-Эрдэне; Камареддин, Лейла; Уотник, Паула И. (2021). «Ацетат, полученный из микробиоты, активирует врожденный иммунитет кишечника через комплекс гистонацетилтрансферазы Tip60» . Иммунитет . 54 (8): 1683–1697.e3. doi : 10.1016/j.immuni.2021.05.017 . ISSN 1074-7613 . ПМЦ 8363570 . ПМИД 34107298 .
- ^ Мендес, Р.; Рааймейкерс, Дж. М. (2015). «Сходства в функциях микробиома между королевствами» . Журнал ISME . 9 (9): 1905–1907. Бибкод : 2015ISMEJ...9.1905M . дои : 10.1038/ismej.2015.7 . ПМЦ 4542044 . ПМИД 25647346 .
- ^ Jump up to: а б с д Бош, ТКГ; Макфолл-Нгай, MJ (2011). «Метаорганизмы как новый рубеж» . Зоология . 114 (4): 185–190. Бибкод : 2011Zool..114..185B . дои : 10.1016/j.zool.2011.04.001 . ПМЦ 3992624 . ПМИД 21737250 .
- ^ Поро Б., Биология и сложность: история и модели комменсализма . Кандидатская диссертация, Лионский университет, Франция, 2014 г.
- ^ Jump up to: а б с д Шервуд, Линда; Уилли, Джоанн; Вулвертон, Кристофер (2013). Микробиология Прескотта (9-е изд.). Нью-Йорк: МакГроу Хилл. стр. 713–721. ISBN 9780073402406 . ОСЛК 886600661 .
- ^ Куигли, Э.М. (сентябрь 2013 г.). «Кишечные бактерии в здоровье и болезни» . Гастроэнтерол Гепатол (Нью-Йорк) . 9 (9): 560–569. ПМЦ 3983973 . ПМИД 24729765 .
- ^ Jump up to: а б с Реми В., Тейлор Т.Н., Хасс Х., Керп Х. (1994). «Везикулярная арбускулярная микориза возрастом четыреста миллионов лет» . Учеб. Натл. акад. наук. США . 91 (25): 11841–11843. Бибкод : 1994PNAS...9111841R . дои : 10.1073/pnas.91.25.11841 . ПМЦ 45331 . ПМИД 11607500 .
- ^ Коупленд, CS (сентябрь – октябрь 2017 г.). «Мир внутри нас» . Журнал здравоохранения Нового Орлеана . Архивировано из оригинала 07.12.2019 . Проверено 7 декабря 2019 г.
- ^ Jump up to: а б Компант С., Даффи Б., Новак Дж., Клеман С., Барка Э.А. (2005). «Использование бактерий, стимулирующих рост растений, для биоконтроля болезней растений: принципы, механизмы действия и перспективы» . Appl Environ Microbiol . 71 (9): 4951–4959. Бибкод : 2005ApEnM..71.4951C . дои : 10.1128/АЕМ.71.9.4951-4959.2005 . ПМЦ 1214602 . ПМИД 16151072 .
- ^ Ткач, Анджей; Чима, Джитендер; Чандра, Говинд; Грант, Аластер; Пул, Филип С. (ноябрь 2015 г.). «Стабильность и преемственность микробиоты ризосферы зависят от типа растений и состава почвы» . ИСМЕ Дж . 9 (11): 2349–2359. Бибкод : 2015ISMEJ...9.2349T . дои : 10.1038/ismej.2015.41 . ПМЦ 4611498 . ПМИД 25909975 .
- ^ Коупленд, CS (19 апреля 2019 г.). «Что такое Clostridium difficile?» . Жизнеспособность .
- ^ Часто задаваемые вопросы Американской академии микробиологии : Микробиом человека. Архивировано 31 декабря 2016 г. в Wayback Machine, январь 2014 г.
- ^ Джуда Л. Рознер для журнала Microbe, февраль 2014 г. Микробных клеток в десять раз больше, чем клеток тела у людей?
- ^ Элисон Эбботт для Nature News. 8 января 2016 г. Ученые развенчивают миф о том, что в нашем организме бактерий больше, чем клеток человека
- ^ Отправитель, Р; Фукс, С; Майло, Р. (январь 2016 г.). «Действительно ли нас значительно превосходит численность? Пересматривая соотношение бактерий и клеток-хозяев у людей» . Клетка . 164 (3): 337–340. дои : 10.1016/j.cell.2016.01.013 . ПМИД 26824647 . S2CID 1790146 .
- ^ Jump up to: а б Отправитель, Рон; Фукс, Шай; Майло, Рон (19 августа 2016 г.). «Пересмотренные оценки количества клеток человека и бактерий в организме» . ПЛОС Биология . 14 (8): e1002533. дои : 10.1371/journal.pbio.1002533 . ПМЦ 4991899 . ПМИД 27541692 .
- ^ «На и в тебе». Micropia, www.micropia.nl/en/discover/stories/on-and-in-you/#:~:text=Они представляют собой%20on%20you%2C%20in,re%20known%20as%20human%20микробиота.
- ^ «Проект NIH Human Microbiome Project определяет нормальный бактериальный состав организма» . Новости НИЗ. 13 июня 2012 г.
- ^ Батай, А; Ли-Круз, Л; Трипати, Б; Ким, Х; Уолдман, Б. (январь 2016 г.). «Вариации микробиома в разных участках кожи амфибий: значение для усилий по смягчению последствий хитридиомикоза». Микроб. Экол . 71 (1): 221–232. Бибкод : 2016MicEc..71..221B . дои : 10.1007/s00248-015-0653-0 . ПМИД 26271741 . S2CID 12951957 .
- ^ Вудхэмс, округ Колумбия, Роллинз-Смит Л.А., Алфорд Р.А., Саймон М.А., Харрис Р.Н. (2007). «Врожденная иммунная защита кожи амфибий: антимикробные пептиды и многое другое» . Охрана животных . 10 (4): 425–428. Бибкод : 2007AnCon..10..425W . дои : 10.1111/j.1469-1795.2007.00150.x . S2CID 84293044 .
- ^ Старый Дж. М., Дин Э. М. (2000). «Развитие иммунной системы и иммунологической защиты у молодняка сумчатой сумки». Развивающая и сравнительная иммунология . 24 (5): 445–454. дои : 10.1016/S0145-305X(00)00008-2 . ПМИД 10785270 .
- ^ Стэннард Х.Дж., Миллер Р.Д., Олд Дж.М. (2020). «Молоко сумчатых и однопроходных – обзор его питательных веществ и иммунных свойств» . ПерДж . 8 : е9335. дои : 10.7717/peerj.9335 . ПМК 7319036 . ПМИД 32612884 .
- ^ Старый Дж. М., Дин Э. М. (1998). «Влияние течки и присутствия молодняка на аэробные бактерии, выделенные из сумки таммарного валлаби Macropus eugenii ». Сравнительная иммунология, микробиология и инфекционные болезни . 21 (4): 237–245. дои : 10.1016/s0147-9571(98)00022-8 . ПМИД 9775355 .
- ^ Брюльк Дж.М.; Антонопулос Д.А.; Миллер МЭБ; и др. (2009). «Генецентрическая метагеномика микробиома рубца крупного рогатого скота, прикрепившегося к волокнам, выявляет специфические для корма гликозидгидролазы» . Учеб. Натл. акад. наук. США . 106 (6): 1948–1953. Бибкод : 2009PNAS..106.1948B . дои : 10.1073/pnas.0806191105 . ПМЦ 2633212 . ПМИД 19181843 .
- ^ Рассел С.Л., Голд МДж; и др. (май 2012 г.). «Изменения микробиоты, вызванные приемом антибиотиков в раннем возрасте, повышают восприимчивость к аллергической астме» . Представитель ЭМБО . 13 (5): 440–447. дои : 10.1038/embor.2012.32 . ПМЦ 3343350 . ПМИД 22422004 .
- ^ Рассел С.Л., Голд М.Дж. и др. (август 2014 г.). «Перинатальные изменения в микробиоте кишечника, вызванные антибиотиками, оказывают различное влияние на воспалительные заболевания легких». J Аллергия Клин Иммунол . 135 (1): 100–109. дои : 10.1016/j.jaci.2014.06.027 . ПМИД 25145536 .
- ^ Тернбо П.Дж. и др. (декабрь 2006 г.). «Микробиом кишечника, связанный с ожирением, с повышенной способностью собирать энергию». Природа . 444 (7122): 1027–1031. Бибкод : 2006Natur.444.1027T . дои : 10.1038/nature05414 . ПМИД 17183312 . S2CID 4400297 .
- ^ Фейт Дж.Дж., Ахерн П.П., Ридаура В.К. и др. (январь 2014 г.). «Идентификация фенотипических отношений между кишечным микробом и хозяином с использованием комбинаторных сообществ у гнотобиотических мышей» . наук. Перевод Мед . 6 (220): 220. doi : 10.1126/scitranslmed.3008051 . ПМЦ 3973144 . ПМИД 24452263 .
- ^ Барфод, КК; Роггенбак, М; Хансен, Л.Х.; Шёрринг, С; Ларсен, СТ; Соренсен, С.Дж.; Крогфельт, К.А. (2013). «Микробиом легких мышей по отношению к кишечным и вагинальным бактериальным сообществам» . БМК Микробиол . 13 :303. дои : 10.1186/1471-2180-13-303 . ПМЦ 3878784 . ПМИД 24373613 .
- ^ Суен; Скотт Джей-Джей; Эйлуорд ФО; и др. (2010). Зонненбург, Джастин (ред.). «Микробиом насекомых-травоядных с высокой способностью к разложению биомассы растений» . ПЛОС Генет . 6 (9): e1001129. дои : 10.1371/journal.pgen.1001129 . ПМЦ 2944797 . ПМИД 20885794 .
- ^ Бродерик, Николь А.; Бюшон, Николя; Леметр, Бруно (2014). «Вызванные микробиотой изменения в экспрессии генов-хозяев Drosophila melanogaster и морфологии кишечника» . мБио . 5 (3): e01117–14. дои : 10.1128/mBio.01117-14 . ПМК 4045073 . ПМИД 24865556 .
- ^ Якубовская, Агата К.; Фогель, Хейко; Эрреро, Сальвадор (май 2013 г.). «Увеличение микробиоты кишечника после подавления иммунитета у личинок, инфицированных бакуловирусом» . ПЛОС Патог . 9 (5): e1003379. дои : 10.1371/journal.ppat.1003379 . ПМЦ 3662647 . ПМИД 23717206 .
- ^ Уотник, Паула И.; Джагдер, Бат-Эрдэне (01 февраля 2020 г.). «Микробный контроль гомеостаза кишечника посредством передачи сигналов врожденного иммунитета энтероэндокринных клеток» . Тенденции в микробиологии . 28 (2): 141–149. дои : 10.1016/j.tim.2019.09.005 . ISSN 0966-842X . ПМК 6980660 . ПМИД 31699645 .
- ^ Тиббс Т.Н., Лопес Л.Р., Артур Дж.К. (2019). «Влияние микробиоты на развитие иммунитета, хроническое воспаление и рак в контексте старения» . Микробная клетка . 6 (8): 324–334. doi : 10.15698/mic2019.08.685 (неактивен 15 апреля 2024 г.). ПМК 6685047 . ПМИД 31403049 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на апрель 2024 г. ( ссылка ) - ^ Jump up to: а б с Бухгольц Ф., Антониелли Л., Костич Т., Сессич А. и Миттер Б. (2019) «Бактериальное сообщество картофеля пополняется из почвы и частично наследуется из поколения в поколение». PLOS One , 14 (11): e0223691. дои : 10.1371/journal.pone.0223691 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Jump up to: а б Абдельфаттах, Ахмед; Вишневский, Майкл; Шена, Леонардо; Тэк, Айко Дж. М. (2021). «Экспериментальные доказательства наследования микробов у растений и путей передачи от семян к филлосфере и корням» . Экологическая микробиология . 23 (4): 2199–2214. Бибкод : 2021EnvMi..23.2199A . дои : 10.1111/1462-2920.15392 . ISSN 1462-2920 . ПМИД 33427409 . S2CID 231576517 .
- ^ Берлец, Алеш (01 сентября 2012 г.). «Новые методы и результаты изучения микробиоты растений: поиск растительных пробиотиков». Наука о растениях . 193–194: 96–102. Бибкод : 2012PlnSc.193...96B . doi : 10.1016/j.plantsci.2012.05.010 . ПМИД 22794922 .
- ^ Уиппс, Дж. М.; Хэнд, П.; Пинк, Д.; Изгиб, Б-г (01 декабря 2008 г.). «Микробиология филлосферы с особым упором на разнообразие и генотип растений» (PDF) . Журнал прикладной микробиологии . 105 (6): 1744–1755. дои : 10.1111/j.1365-2672.2008.03906.x . ISSN 1365-2672 . ПМИД 19120625 . S2CID 35055151 .
- ^ Чибукос MC, Тайлер Б.М. (2009). «Общие темы получения питательных веществ симбиотическими микробами растений, описанные Онтологией генов» . БМК Микробиология . 9(Дополнение 1) (Дополнение 1): S6. дои : 10.1186/1471-2180-9-S1-S6 . ПМЦ 2654666 . ПМИД 19278554 .
- ^ Клоппер, Дж. В. (1993). «Ризобактерии, стимулирующие рост растений, как средства биологической борьбы». В Меттинге, Ф.Б.-младшем (ред.). Почвенная микробная экология: применение в сельском хозяйстве и управлении окружающей средой . Нью-Йорк: Marcel Dekker Inc., стр. 255–274. ISBN 978-0-8247-8737-0 .
- ^ Блумберг, Г.В.; Лугтенберг, БЖЖ (2001). «Молекулярные основы стимуляции роста растений и биоконтроля с помощью ризобактерий». Современное мнение в области биологии растений . 4 (4): 343–350. Бибкод : 2001COPB....4..343B . дои : 10.1016/S1369-5266(00)00183-7 . ПМИД 11418345 .
- ^ Палм, Ной В.; де Зете, Марсель Р.; Флавелл, Ричард А. (30 июня 2015 г.). «Взаимодействие иммунитета и микробиоты в здоровье и болезни» . Клиническая иммунология . 159 (2): 122–127. дои : 10.1016/j.clim.2015.05.014 . ISSN 1521-6616 . ПМЦ 4943041 . ПМИД 26141651 .
- ^ Раунд, Джун Л.; О'Коннелл, Райан М.; Мазманян, Саркис К. (2010). «Координация толерогенных иммунных ответов комменсальной микробиотой» . Журнал аутоиммунитета . 34 (3): J220–J225. дои : 10.1016/j.jaut.2009.11.007 . ПМК 3155383 . ПМИД 19963349 .
- ^ Jump up to: а б с Кахенцли, Джулия; Балмер, Мария Л.; Маккой, Кэти Д. (2012). «Микробно-иммунные перекрестные взаимодействия и регуляция иммунной системы» . Иммунология . 138 (1): 12–22. дои : 10.1111/j.1365-2567.2012.03624.x . ПМЦ 3533697 . ПМИД 22804726 .
- ^ Розенберг, Юджин; Зильбер-Розенберг, Илана (2016). «Микробы управляют эволюцией животных и растений: концепция гологенома» . мБио . 7 (2): e01395–15. дои : 10.1128/mbio.01395-15 . ПМЦ 4817260 . ПМИД 27034283 .
- ^ Бландер, Дж. Магарян; Лонгман, Рэнди С; Илиев, Илиян Д; Зонненберг, Грегори Ф; Артис, Дэвид (19 июля 2017 г.). «Регуляция воспаления путем взаимодействия микробиоты с хозяином» . Природная иммунология . 18 (8): 851–860. дои : 10.1038/ni.3780 . ISSN 1529-2908 . ПМК 5800875 . ПМИД 28722709 .
- ^ Никоопур, Э; Сингх, Б. (2014). «Взаимность во взаимодействии микробиома и иммунной системы и ее последствия для болезней и здоровья» . Целевые лекарства от аллергии при воспалении . 13 (2): 94–104. дои : 10.2174/1871528113666140330201056 . ПМИД 24678760 .
- ^ Jump up to: а б Розенберг Э., Корен О., Решеф Л., Эфрони Р., Зильбер-Розенберг И. (2007). «Роль микроорганизмов в здоровье, болезнях и эволюции кораллов». Обзоры природы Микробиология . 5 (5): 355–362. дои : 10.1038/nrmicro1635 . ПМИД 17384666 . S2CID 2967190 .
- ^ Бэрд А.Х., Бхагули Р., Ральф П.Дж., Такахаши С. (2009). «Обесцвечивание кораллов: роль хозяина» (PDF) . Тенденции экологии и эволюции . 24 (1): 16–20. Бибкод : 2009TEcoE..24...16B . дои : 10.1016/j.tree.2008.09.005 . ПМИД 19022522 .
- ^ Леггат В., Эйнсворт Т., Байтелл Дж., Дав С., Гейтс Р., Хоег-Гульдберг О., Иглесиас-Прието Р., Йеллоулис Д. (2007). «Теория гологенома не учитывает коралловый холобионт» . Обзоры природы Микробиология . 5 (10): Интернет-переписка. дои : 10.1038/nrmicro1635-c1 . S2CID 9031305 .
- ^ Эйнсворт Т.Д., Файн М., Рофф Г., Хог-Гульдберг О. (2008). «Бактерии не являются основной причиной обесцвечивания средиземноморского коралла Oculina patagonica » . Журнал ISME . 2 (1): 67–73. Бибкод : 2008ISMEJ...2...67A . дои : 10.1038/ismej.2007.88 . ПМИД 18059488 . S2CID 1032896 .
- ^ Jump up to: а б Кучински, Дж.; Лаубер, CL; Уолтерс, Вашингтон; Парфри, LW; Клементе, Дж. К.; Геверс, Д.; Найт, Р. (2011). «Экспериментально-аналитический инструментарий для изучения микробиома человека» . Обзоры природы Генетика . 13 (1): 47–58. дои : 10.1038/nrg3129 . ПМК 5119550 . ПМИД 22179717 .
- ^ Чакраворти, Сумитеш; Хелб, Даника; Бердей, Мишель; Коннелл, Нэнси; Алланд, Дэвид (май 2007 г.). «Детальный анализ сегментов гена 16S рибосомальной РНК для диагностики патогенных бактерий» . Журнал микробиологических методов . 69 (2): 330–339. дои : 10.1016/j.mimet.2007.02.005 . ПМК 2562909 . ПМИД 17391789 .
- ^ Сориано-Лерма, Ана; Перес-Карраско, Вирджиния; Санчес-Мараньон, Мануэль; Ортис-Гонсалес, Матильда; Санчес-Мартин, Виктория; Хихон, Хуан; Наварро-Мари, Хосе Мария; ГАРСИА-САЛЬСЕДО, Хосе Антонио; Сориано, Мигель (декабрь 2020 г.). «Влияние целевой области 16S рРНК на результаты исследований микробиома в образцах почвы и слюны» . Научные отчеты . 10 (1): 13637. Бибкод : 2020NatSR..1013637S . дои : 10.1038/s41598-020-70141-8 . ISSN 2045-2322 . ПМЦ 7423937 . ПМИД 32788589 .
- ^ Маркези, младший (2010). «Прокариотическое и эукариотическое разнообразие кишечника человека». Достижения прикладной микробиологии, том 72 . Том. 72. стр. 43–62. дои : 10.1016/S0065-2164(10)72002-5 . ISBN 9780123809896 . ПМИД 20602987 .
- ^ Вестхайм, Х.; Джарман, С.Н. (2008). «Блокирующие праймеры для усиления ПЦР-амплификации редких последовательностей в смешанных образцах – тематическое исследование ДНК добычи в желудках антарктического криля» . Границы в зоологии . 5:12 . дои : 10.1186/1742-9994-5-12 . ПМЦ 2517594 . ПМИД 18638418 .
- ^ «VAMPS: визуализация и анализ структур микробных популяций» . Центр Бэй Пола, MBL, Вудс-Хоул . Проверено 11 марта 2012 г.
- ^ Капорасо, Дж.Г.; Кучински, Дж.; Стомбо, Дж.; Биттингер, К.; Бушман, Федеральный округ; Костелло, ЕК; Фирер, Н.; Пенья, АГ; Гудрич, Дж. К.; Гордон, Дж.И.; Хаттли, Джорджия; Келли, ST; Найтс, Д.; Кениг, Дж. Э.; Лей, RE; Лозупоне, Калифорния; Макдональд, Д.; Мюгге, Б.Д.; Пиррунг, М.; Ридер, Дж.; Севинский-младший; Тернбо, П.Дж.; Уолтерс, Вашингтон; Видманн, Дж.; Яцуненко Т.; Заневельд, Дж.; Найт, Р. (2010). «QIIME позволяет анализировать данные общественного секвенирования с высокой пропускной способностью» . Природные методы . 7 (5): 335–336. дои : 10.1038/nmeth.f.303 . ПМК 3156573 . ПМИД 20383131 .
- ^ Шлосс, PD; Уэсткотт, СЛ; Рябин Т.; Холл, младший; Хартманн, М.; Холлистер, Э.Б.; Лесневский, РА; Окли, BB; Паркс, ДХ; Робинсон, CJ; Сахл, JW; Стрес, Б.; Таллинджер, Г.Г.; Ван Хорн, диджей; Вебер, CF (2009). «Представляем mothur: программное обеспечение с открытым исходным кодом, независимое от платформы и поддерживаемое сообществом, для описания и сравнения микробных сообществ» . Прикладная и экологическая микробиология . 75 (23): 7537–7541. Бибкод : 2009ApEnM..75.7537S . дои : 10.1128/АЕМ.01541-09 . ПМЦ 2786419 . ПМИД 19801464 .
- ^ Каллахан, Бенджамин Дж.; Макмерди, Пол Дж.; Розен, Майкл Дж.; Хан, Эндрю В.; Джонсон, Эми Джо А.; Холмс, Сьюзен П. (июль 2016 г.). «DADA2: Вывод образцов высокого разрешения на основе данных ампликона Illumina» . Природные методы . 13 (7): 581–583. дои : 10.1038/nmeth.3869 . ISSN 1548-7105 . ПМЦ 4927377 . ПМИД 27214047 .
- ^ Эдгар, Роберт К. (15 октября 2016 г.). «UNOISE2: улучшенное исправление ошибок для секвенирования ампликонов Illumina 16S и ITS» : 081257. doi : 10.1101/081257 . S2CID 784388 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Тернбо, П.Дж.; Хамади, М.; Яцуненко Т.; Кантарель, БЛ; Дункан, А.; Лей, RE; Согин, М.Л.; Джонс, WJ; Роу, бакалавр; Аффуртит, JP; Эгхольм, М.; Генриссат, Б.; Хит, AC; Найт, Р.; Гордон, Дж.И. (2008). «Основной микробиом кишечника у тучных и худых близнецов» . Природа . 457 (7228): 480–484. Бибкод : 2009Natur.457..480T . дои : 10.1038/nature07540 . ПМЦ 2677729 . ПМИД 19043404 .
- ^ Цинь, Дж.; Ли, Р.; Раес, Дж.; Арумугам, М.; Бургдорф, Канзас; Маничан, К.; Нильсен, Т.; Понс, Н.; Левенес, Ф.; Ямада, Т.; У меня есть, ДР; Ли, Дж.; Сюй, Дж.; Гладкий.; Ли, Д.; Цао, Дж.; Ван, Б.; Лян, Х.; Чжэн, Х.; Се, Ю.; Тэп, Дж.; Лепаж, П.; Берталан, М.; Батто, Дж. М.; Хансен, Т.; Ле Паслье, Д.; Линнеберг, А.; Нильсен, HBR; Пеллетье, Э.; Рено, П. (2010). «Каталог генов микробов кишечника человека, созданный с помощью метагеномного секвенирования» . Природа . 464 (7285): 59–65. Бибкод : 2010Natur.464...59. . дои : 10.1038/nature08821 . ПМЦ 3779803 . ПМИД 20203603 .
- ^ Тринге, СГ; Фон Меринг, К.; Кобаяши, А.; Саламов А.А.; Чен, К.; Чанг, HW; Подар, М.; Шорт, Дж. М.; Матур, Э.Дж.; Деттер, Дж.К.; Борк, П.; Гугенгольц, П.; Рубин, Э.М. (2005). «Сравнительная метагеномика микробных сообществ». Наука . 308 (5721): 554–557. Бибкод : 2005Sci...308..554T . CiteSeerX 10.1.1.377.2288 . дои : 10.1126/science.1107851 . ПМИД 15845853 . S2CID 161283 .
- ^ Вули, Джей Си; Годзик, А.; Фридберг, И. (2010). Борн, Филип Э. (ред.). «Букварь по метагеномике» . PLOS Вычислительная биология . 6 (2): e1000667. Бибкод : 2010PLSCB...6E0667W . дои : 10.1371/journal.pcbi.1000667 . ПМК 2829047 . ПМИД 20195499 .
- ^ Уотсон, Мик; Роэ, Райнер; Уокер, Алан В.; Дьюхерст, Ричард Дж.; Снеллинг, Тимоти Дж.; Иван Лячко; Лэнгфорд, Кайл В.; Пресс, Максимилиан О.; Уайзер, Эндрю Х. (28 февраля 2018 г.). «Сборка 913 микробных геномов в результате метагеномного секвенирования рубца коровы» . Природные коммуникации . 9 (1): 870. Бибкод : 2018NatCo...9..870S . дои : 10.1038/s41467-018-03317-6 . ISSN 2041-1723 . ПМК 5830445 . ПМИД 29491419 .
- ^ Мюллер, Дж.; Шклярчик, Д.; Жюльен, П.; Летунич, И.; Рот, А.; Кун, М.; Пауэлл, С.; Фон Меринг, К.; Доркс, Т.; Дженсен, LJ; Борк, П. (2009). «EggNOG v2.0: Расширение эволюционной генеалогии генов за счет расширенных неконтролируемых ортологичных групп, видов и функциональных аннотаций» . Исследования нуклеиновых кислот . 38 (Проблема с базой данных): D190–D195. дои : 10.1093/нар/gkp951 . ПМК 2808932 . ПМИД 19900971 .
- ^ Канехиса, М.; Гото, С.; Фурумичи, М.; Танабэ, М.; Хиракава, М. (2009). «KEGG для представления и анализа молекулярных сетей, связанных с болезнями и лекарствами» . Исследования нуклеиновых кислот . 38 (Проблема с базой данных): D355–D360. дои : 10.1093/нар/gkp896 . ПМК 2808910 . ПМИД 19880382 .
- ^ Мейер, Ф.; Паарманн, Д.; д'Суза, М.; Олсон, Р.; Стекло, ЭМ; Кубал, М.; Пачиан, Т.; Родригес, А.; Стивенс, Р.; Вилке, А.; Уилкенинг, Дж.; Эдвардс, РА (2008). «Метагеномный RAST-сервер – общедоступный ресурс для автоматического филогенетического и функционального анализа метагеномов» . БМК Биоинформатика . 9 : 386. дои : 10.1186/1471-2105-9-386 . ПМК 2563014 . ПМИД 18803844 .
- ^ Солнце, С.; Чен, Дж.; Ли, В.; Алтинтас, И.; Лин, А.; Пельтье, С.; Стокс, К.; Аллен, Э.Э.; Эллисман, М.; Грета, Дж.; Вули, Дж. (2010). «Киберинфраструктура сообщества для передовых исследований и анализа микробной экологии: ресурс CAMERA» . Исследования нуклеиновых кислот . 39 (Проблема с базой данных): D546–D551. дои : 10.1093/нар/gkq1102 . ПМК 3013694 . ПМИД 21045053 .
- ^ Марковиц, В.М.; Иванова, Н.Н.; Сзето, Э.; Паланиаппан, К.; Чу, К.; Далеви, Д.; Чен, IMA; Гречкин Ю.; Дубчак И.; Андерсон, И.; Ликидис, А.; Мавроматис, К.; Гугенгольц, П.; Кирпидес, Северная Каролина (2007). «IMG/M: Система управления и анализа данных метагеномов» . Исследования нуклеиновых кислот . 36 (Проблема с базой данных): D534–D538. дои : 10.1093/нар/gkm869 . ПМК 2238950 . ПМИД 17932063 .
- ^ Ши, Ю.; Тайсон, GW; Делонг, EF (2009). «Метатранскриптомика обнаруживает уникальные микробные малые РНК в толще воды океана». Природа . 459 (7244): 266–269. Бибкод : 2009Natur.459..266S . дои : 10.1038/nature08055 . ПМИД 19444216 . S2CID 4340144 .
- ^ Хименес, РМ; Делварт, Э.; Луптак, А (2011). «Структурный поиск выявил рибозимы-молоты в микробиоме человека» . Журнал биологической химии . 286 (10): 7737–7743. дои : 10.1074/jbc.C110.209288 . ПМК 3048661 . ПМИД 21257745 .
- ^ Марон, Пенсильвания; Ранджард, Л.; Мугель, К.; Лемансо, П. (2007). «Метапротеомика: новый подход к изучению функциональной микробной экологии». Микробная экология . 53 (3): 486–493. Бибкод : 2007MicEc..53..486M . дои : 10.1007/s00248-006-9196-8 . ПМИД 17431707 . S2CID 26953155 .
- ^ Jump up to: а б «Проект NIH по микробиому человека» . Национальные институты здравоохранения США, Министерство здравоохранения и социальных служб, Правительство США. 2016. Архивировано из оригинала 11 июня 2016 года . Проверено 14 июня 2016 г.
- ^ Гилберт, Дж.А.; Мейер, Ф.; Антонопулос, Д.; и др. (2010). «Отчет о встрече: Семинар по метагеномике Terabase и видение проекта земного микробиома» . Стандарты в геномных науках . 3 (3): 243–248. дои : 10.4056/sigs.1433550 . ПМК 3035311 . ПМИД 21304727 .
- ^ Гилберт, Дж.А.; О'Дор, Р.; Кинг, Н.; Фогель, ТМ (2011). «Важность метагеномных исследований для микробной экологии: или почему Дарвин стал метагеномным ученым» . Микробная информатика и экспериментирование . 1 (1): 5. дои : 10.1186/2042-5783-1-5 . ПМЦ 3348666 . ПМИД 22587826 .
- ^ Ибрагим, Несма (01 июля 2018 г.). «Микробиота кишечника и сахарный диабет 2 типа: какая связь?» . Афро-египетский журнал инфекционных и эндемических заболеваний . 6 (2): 112–119. дои : 10.21608/aeji.2018.9950 . ISSN 2090-7184 . S2CID 3900880 .
- ^ Терсби, Элизабет; Джуг, Натали (01 июня 2017 г.). «Знакомство с микробиотой кишечника человека» . Биохимический журнал . 474 (11): 1823–1836. дои : 10.1042/BCJ20160510 . ISSN 0264-6021 . ПМЦ 5433529 . ПМИД 28512250 .
- ^ Jump up to: а б с Муньос-Гарач, Арасели; Диас-Пердигонес, Кристина; Тинахонес, Франсиско Дж. (декабрь 2016 г.). «Микробиота и сахарный диабет 2 типа». Эндокринология и питание (на испанском языке). 63 (10): 560–568. дои : 10.1016/j.indonu.2016.07.008 . ПМИД 27633134 .
- ^ Бландино, Г.; Интурри, Р.; Лаззара, Ф.; ДиРоса, М.; Малагуарнера, Л. (1 ноября 2016 г.). «Влияние микробиоты кишечника на сахарный диабет». Диабет и обмен веществ . 42 (5): 303–315. дои : 10.1016/j.diabet.2016.04.004 . ISSN 1262-3636 . ПМИД 27179626 .
- ^ Ванденплас, Ю., Карниелли, В.П., Ксязык, Дж., Луна, М.С., Мигачева, Н., Моссельманс, Дж.М., ... и Вабич, М. (2020), Факторы, влияющие на развитие микробиоты кишечника в раннем возрасте. Питание, 78, 110812.
- ^ Корпела К., Хельве О., Колхо К.Л., Сайсто Т., Скогберг К., Дикарева Е., Стефанович В., Салонен А., Андерссон С., де Вос В.М. Трансплантация материнской фекальной микробиоты у младенцев, рожденных с помощью кесарева сечения, быстро восстанавливает нормальное развитие микробов в кишечнике: исследование, подтверждающее концепцию. Клетка. 15 октября 2020 г.;183(2):324-334.e5. doi: 10.1016/j.cell.2020.08.047. Epub, 1 октября 2020 г. PMID 33007265.
- ^ Корпела, К., Салонен, А., Саксен, Х., Никконен, А., Пелтола, В., Яаккола, Т., ... и Колхо, К.Л. (2020). Антибиотики в раннем возрасте связаны со специфическими признаками кишечной микробиоты в проспективной когорте младенцев. Педиатрические исследования, 1-6
- ^ Шей, К., Симпсон, М.Р., Авершина, Э., Руди, К., Ойен, Т., Юлиуссон, П.Б., ... и Эдегард, РА (2020). Грибковая и бактериальная микробиота раннего кишечника и рост в детском возрасте. Границы в педиатрии, 8, 658
- ^ Корпела К., Салонен А., Вирта Л.Дж., Кекконен Р.А., Форслунд К., Борк П. и Де Вос ВМ (2016). Кишечный микробиом связан с пожизненным применением антибиотиков у финских детей дошкольного возраста. Природные коммуникации, 7, 10410
- ^ Корпела, К., Салонен, А., Саксен, Х., Никконен, А., Пелтола, В., Яаккола, Т., ... и Колхо, К.Л. (2020). Антибиотики в раннем возрасте связаны со специфическими признаками кишечной микробиоты в проспективной когорте младенцев. Педиатрические исследования, 1-6.
- ^ Jump up to: а б с Зилбиндер, Б., Чен, Дж., Брунке, С., Васкес-Урибе, Р., Сантаман, Р., Мейер, А.С., ... и Панайоту, Г. (2020). Антибиотики вызывают переход от мутуализма к конкуренции в кишечных сообществах человека, оказывая более продолжительное воздействие на грибы, чем на бактерии. Микробиом, 8(1), 1-20
- ^ Кабрал, DJ, Пенумутчу, С., Норрис, К., Моронес-Рамирес, младший, и Беленький, П. (2018). Микробная конкуренция между Escherichia coli и Candida albicans выявляет растворимый фунгицидный фактор. Микробная клетка, 5(5), 249
- ^ Пелег, А.Ю., Хоган, Д.А., и Милонакис, Э. (2010). Медицински важные бактериально-грибковые взаимодействия. Nature Reviews Микробиология, 8 (5), 340–349
- ^ Сокол Х, Ледюк В, Ашард Х, Фам Х.П., Жегу С., Ландман С, Коэн Д., Лигуори Дж., Буррье А., Нион-Лармурье I, Коснес Дж., Сексик П., Ланджелла П., Скурник Д., Ричард М.Л., Божери Л. Грибковый дисбиоз микробиоты при ВЗК. Гут 2017;66:1039–1048. дои: 10.1136/gutjnl-2015-310746
- ^ Ребекка Вентин-Хольмберг, Аня Эберл, Шахзад Сакиб, Катри Корпела, Сеппо Виртанен, Тайна Сиппонен, Анне Салонен, Пяйви Саавалайнен, Эйя Ниссила, Бактериальные и грибковые профили как маркеры реакции на препарат инфликсимаб при воспалительном заболевании кишечника, Журнал Крона и колита , 2020;, jjaa252, https://doi.org/10.1093/ecco-jcc/jjaa252
- ^ Эль Хаге, Р., Эрнандес-Санабриа, Э., Калатаюд Арройо, М., Реквизит, Р., и Ван де Виле, Т. (2019). Консорциум по производству пропионата восстанавливает дисбактериоз, вызванный антибиотиками, в динамической модели микробной экосистемы кишечника человека in vitro. Границы микробиологии, 10, 1206.
- ^ Тиан, X., Хеллман, Дж., Хорсвилл, А.Р., Кросби, Х.А., Фрэнсис, К.П., и Пракаш, А. (2019). Повышенные уровни пропионата, получаемого из микробиома кишечника, связаны со снижением стерильного воспаления легких и бактериального иммунитета у мышей. Границы микробиологии, 10, 159.
- ^ Ли, Ю., Фаден, Х.С., и Чжу, Л. (2020). Реакция микробиоты кишечника на изменения в питании в первые два года жизни. Границы фармакологии, 11, 334.
- ^ журнал, Юэн. «Микробная ДНК в организме человека может быть использована для идентификации личности» . Научный американец . Проверено 17 мая 2015 г.
- ^ Каллауэй, Юэн (2015). «Микробиомы вызывают проблемы конфиденциальности» . Природа . 521 (7551): 136. Бибкод : 2015Natur.521..136C . дои : 10.1038/521136а . ПМИД 25971486 . S2CID 4393347 .
- ^ Йонг, Эд (11 мая 2015 г.). «Могут ли микробы, которые вы оставляете после себя, использовать для вашей идентификации?» . Нэшнл Географик . Архивировано из оригинала 30 мая 2015 года . Проверено 17 мая 2015 г.
