Волосковая клетка
| Волосковая клетка | |
|---|---|
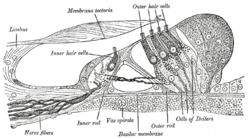 Разрез спирального кортиева органа . Увеличено. («Наружные волосковые клетки» отмечены вверху; «внутренние волосковые клетки» отмечены рядом с центром). | |
 Поперечный разрез улитки . Внутренние волосковые клетки расположены на окончании «нервов внутренних волосковых клеток», а внешние волосковые клетки расположены на окончании «нервов наружных волосковых клеток». | |
| Подробности | |
| Расположение | Улитка |
| Форма | Уникальный (см. текст) |
| Функция | Усиливайте звуковые волны и передайте слуховую информацию в ствол мозга. |
| Нейромедиатор | Глутамат |
| Пресинаптические связи | Никто |
| Постсинаптические связи | Через слуховой нерв к преддверно-улитковому нерву и нижнему холмику. |
| Идентификаторы | |
| НейроЛекс ID | sao1582628662 , sao429277527 |
| Анатомические термины нейроанатомии | |
Волосковые клетки являются сенсорными рецепторами как слуховой, так и вестибулярной системы в ушах всех позвоночных животных , а также в органе боковой линии рыб. Посредством механотрансдукции волосковые клетки обнаруживают движение в окружающей среде. [1]
У млекопитающих расположены внутри спирального кортиева органа на тонкой базилярной мембране улитки слуховые волосковые клетки внутреннего уха . Свое название они получили от пучков стереоцилий, называемых пучками волос , которые выступают из апикальной поверхности клетки в заполненный жидкостью улитковый проток . Стереоцилий насчитывается от пятидесяти до ста в каждой клетке, они плотно упакованы вместе. [2] и уменьшаются в размерах по мере удаления от киноцилии . [3]
Волосковые клетки улитки млекопитающих делятся на два анатомически и функционально различных типа: наружные и внутренние волосковые клетки. Повреждение этих волосковых клеток приводит к снижению слуховой чувствительности , а поскольку волосковые клетки внутреннего уха не могут регенерировать , это повреждение является постоянным. [4] Повреждение волосковых клеток может привести к повреждению вестибулярной системы и, следовательно, вызвать трудности с балансировкой. Однако у других позвоночных, таких как часто изучаемые рыбки данио и птицы , есть волосковые клетки, которые могут регенерировать. [5] [6] Улитка человека при рождении содержит около 3500 внутренних волосковых клеток и 12 000 наружных волосковых клеток. [7]
Наружные волосковые клетки механически усиливают звук низкого уровня, попадающий в улитку . [8] [9] Усиление может осуществляться за счет движения пучков волос или за счет электрической подвижности их клеточных тел. Эта так называемая соматическая электроподвижность усиливает звук у всех четвероногих . На это влияет механизм закрытия механических сенсорных ионных каналов на кончиках пучков волос. [ нужна ссылка ]
Внутренние волосковые клетки преобразуют звуковые вибрации в жидкостях улитки в электрические сигналы, которые затем передаются через слуховой нерв в слуховой ствол мозга и в слуховую кору .
Внутренние волосковые клетки – от звука к нервному сигналу
[ редактировать ]
волосковых клеток Отклонение стереоцилий открывает механически закрытые ионные каналы , которые позволяют любым небольшим положительно заряженным ионам (в первую очередь калию и кальцию ) проникать в клетку. [10] В отличие от многих других электрически активных клеток, волосковая клетка сама по себе не генерирует потенциал действия . Вместо этого приток положительных ионов из эндолимфы в среднюю лестницу деполяризует клетку, что приводит к возникновению рецепторного потенциала . Этот рецепторный потенциал открывает потенциалзависимые кальциевые каналы ; ионы кальция затем проникают в клетку и вызывают высвобождение нейротрансмиттеров на базальном конце клетки. Нейромедиаторы диффундируют через узкое пространство между волосковой клеткой и нервным окончанием, где они затем связываются с рецепторами и, таким образом, запускают потенциалы действия в нерве. Таким образом механический звуковой сигнал преобразуется в электрический нервный сигнал. Реполяризация волосковых клеток осуществляется особым образом. Перилимфа имеет барабанной лестницы очень низкую концентрацию положительных ионов. Электрохимический градиент заставляет положительные ионы течь по каналам в перилимфу.
Волосковые клетки хронически теряют Ca 2+ . Эта утечка вызывает тонизирующее высвобождение нейромедиатора в синапсах. Считается, что именно этот тонизирующий эффект позволяет волосковым клеткам так быстро реагировать на механические раздражители. Быстрота реакции волосковых клеток также может быть связана с тем, что они могут увеличивать количество высвобождаемого нейротрансмиттера в ответ на изменение мембранного потенциала всего на 100 мкВ. [11]
Волосковые клетки также способны различать частоты тонов одним из двух методов. Первый метод, обнаруженный только у немлекопитающих, использует электрический резонанс в базолатеральной мембране волосковой клетки. Электрический резонанс в этом методе проявляется как затухающие колебания мембранного потенциала в ответ на приложенный импульс тока. Второй метод использует тонотопические различия базилярной мембраны. Эта разница возникает из-за разного расположения волосковых клеток. Волосковые клетки, обладающие высокочастотным резонансом, расположены на базальном конце, тогда как волосковые клетки, обладающие значительно более низким частотным резонансом, находятся на апикальном конце эпителия . [12]
Внешние волосковые клетки – акустические предусилители
[ редактировать ]В наружных волосковых клетках млекопитающих изменяющийся рецепторный потенциал преобразуется в активные вибрации тела клетки. Эта механическая реакция на электрические сигналы называется соматической электромобильностью; [13] он управляет изменениями длины ячейки, синхронизируется с входящим звуковым сигналом и обеспечивает механическое усиление за счет обратной связи с бегущей волной. [14]
Наружные волосковые клетки встречаются только у млекопитающих. Хотя слуховая чувствительность млекопитающих аналогична таковой у других классов позвоночных, без функционирующих наружных волосковых клеток чувствительность снижается примерно на 50 дБ. [15] Наружные волосковые клетки расширяют диапазон слуха у некоторых морских млекопитающих примерно до 200 кГц. [16] Они также улучшили частотную избирательность (частотную дискриминацию), что особенно полезно для людей, поскольку позволяет создавать сложные речи и музыку. Наружные волосковые клетки функционируют даже после того, как клеточные запасы АТФ истощены. [13]
Эффект этой системы заключается в нелинейном усилении тихих звуков в большей степени, чем громких, так что широкий диапазон звукового давления может быть уменьшен до гораздо меньшего диапазона смещения волос. [17] Это свойство усиления называется кохлеарным усилителем .
В молекулярной биологии волосковых клеток за последние годы достигнут значительный прогресс благодаря идентификации моторного белка ( престина ), который лежит в основе соматической электроподвижности во внешних волосковых клетках. Было показано, что функция престина зависит от передачи сигналов хлоридных каналов и что она нарушается обычным морским пестицидом трибутилотином . Поскольку этот класс загрязнителей биоконцентрируется вверх по пищевой цепочке, эффект выражен у высших морских хищников, таких как косатки и зубатые киты . [18]
Адаптация сигнала волосковых клеток
[ редактировать ]Приток ионов кальция играет важную роль в адаптации волосковых клеток к усилению сигнала. Это позволяет людям игнорировать постоянные звуки, которые уже не новы, и позволяет нам остро реагировать на другие изменения в нашем окружении. Ключевой механизм адаптации исходит от моторного белка миозина-1c, который обеспечивает медленную адаптацию, обеспечивает напряжение для сенсибилизации каналов трансдукции, а также участвует в аппарате передачи сигналов. [19] [20] Более поздние исследования показывают, что чувствительное к кальцию связывание кальмодулина с миозином-1с может фактически модулировать взаимодействие адаптационного мотора и с другими компонентами аппарата трансдукции. [21] [22]
Быстрая адаптация: во время быстрой адаптации Ca 2+ ионы, которые попадают в стереоцилию через открытый МЕТ-канал, быстро связываются с участком на канале или рядом с ним и вызывают закрытие канала. увеличивается Когда каналы закрываются, напряжение в кончике звена , тяня пучок в противоположном направлении. [19] Быстрая адаптация более заметна в волосковых клетках, распознающих звук и слух, а не в вестибулярных клетках.
Медленная адаптация: доминирующая модель предполагает, что медленная адаптация происходит, когда миозин-1c скользит вниз по стереоцилии в ответ на повышенное напряжение во время смещения пучка. [19] Результирующее снижение напряжения в концевом звене позволяет пучку двигаться дальше в противоположном направлении. Когда напряжение уменьшается, каналы закрываются, что приводит к снижению тока трансдукции. [19] Медленная адаптация наиболее заметна в вестибулярных волосковых клетках, которые чувствуют пространственное движение, и в меньшей степени в волосковых клетках улитки, которые обнаруживают слуховые сигналы. [20]
Нейронная связь
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( сентябрь 2016 г. ) |
Нейроны слухового или вестибулокохлеарного нерва (восьмой черепной нерв ) иннервируют волосковые клетки улитки и вестибулярного нерва. [23] Нейромедиатором, высвобождаемым волосковыми клетками и стимулирующим концевые нейриты периферических аксонов афферентных ( по направлению к мозгу) нейронов, считается глутамат . В пресинаптическом соединении имеется отчетливое пресинаптическое плотное тело или лента . Это плотное тело окружено синаптическими пузырьками и, как полагают, способствует быстрому высвобождению нейромедиатора.
Иннервация нервных волокон внутренних волосковых клеток гораздо плотнее, чем наружных. Одна внутренняя волосковая клетка иннервируется многочисленными нервными волокнами, тогда как одно нервное волокно иннервирует множество наружных волосковых клеток. Внутренние нервные волокна волосковых клеток также очень сильно миелинизированы, в отличие от немиелинизированных наружных нервных волокон волосковых клеток. можно считать участок базилярной мембраны, подающий входы к определенному афферентному нервному волокну Рецептивным полем .
Эфферентные проекции мозга на улитку также играют роль в восприятии звука. Эфферентные синапсы возникают на внешних волосковых клетках и на афферентных аксонах под внутренними волосковыми клетками. Пресинаптический терминальный бутон заполнен пузырьками, содержащими ацетилхолин и нейропептид, называемый пептидом, родственным гену кальцитонина . Эффекты этих соединений различаются; в некоторых волосковых клетках ацетилхолин гиперполяризует клетку, что локально снижает чувствительность улитки.
возобновление роста
[ редактировать ]Исследования возобновления роста клеток улитки могут привести к разработке методов лечения, восстанавливающих слух. В отличие от птиц и рыб, люди и другие млекопитающие, как правило, не способны вырастить заново клетки внутреннего уха, преобразующие звук в нервные сигналы, когда эти клетки повреждены возрастом или болезнью. [6] [24] Исследователи добиваются прогресса в генной терапии и терапии стволовыми клетками , которые могут позволить регенерировать поврежденные клетки. Поскольку было обнаружено, что волосковые клетки слуховой и вестибулярной систем у птиц и рыб способны к регенерации, их способность изучалась длительное время. [6] [25] Кроме того, было показано, что волосковые клетки боковой линии , которые обладают функцией механотрансдукции и обнаружены у анамниот , снова растут у таких видов, как рыбки данио . [26]
Исследователи идентифицировали ген млекопитающих, который обычно действует как молекулярный переключатель , блокируя повторный рост волосковых клеток улитки у взрослых. [27] Ген Rb1 кодирует белок ретинобластомы , который является супрессором опухоли . Rb останавливает деление клеток, стимулируя их выход из клеточного цикла. [28] [29] Волосковые клетки в культуральной чашке не только регенерируют при удалении гена Rb1, но и у мышей, выведенных с отсутствием этого гена, вырастает больше волосковых клеток, чем у контрольных мышей, у которых есть этот ген. Кроме того, sonic hedgehog было показано, что белок блокирует активность белка ретинобластомы , тем самым индуцируя повторный вход в клеточный цикл и повторный рост новых клеток. [30]
Несколько ингибиторов сигнального пути Notch , включая ингибитор гамма-секретазы LY3056480, изучаются на предмет их потенциальной способности регенерировать волосковые клетки в улитке. [31] [32]
TBX2 Было показано, что (транскрипционный фактор 2 T-box) является главным регулятором дифференцировки внутренних и внешних волосковых клеток. [33] Это открытие позволило исследователям направить волосковые клетки на развитие внутренних или внешних волосковых клеток, что может помочь заменить отмершие волосковые клетки и предотвратить или обратить вспять потерю слуха. [34] [35]
Также было обнаружено, что ингибитор клеточного цикла p27kip1 ( CDKN1B ) стимулирует повторный рост волосковых клеток улитки у мышей после генетической делеции или нокдауна с помощью siRNA, нацеленной на p27. [36] [37] Исследования регенерации волосковых клеток могут приблизить нас к клиническому лечению потери слуха у человека , вызванной повреждением или смертью волосковых клеток.
См. также
[ редактировать ]Дополнительные изображения
[ редактировать ] Сетчатая пластинка и нижележащие структуры.
Сетчатая пластинка и нижележащие структуры. Стереоцилии внутреннего уха лягушки
Стереоцилии внутреннего уха лягушки


Ссылки
[ редактировать ]- ^ Лампкин, Эллен А.; Маршалл, Кара Л.; Нельсон, Эйслин М. (2010). «Клеточная биология прикосновения» . Журнал клеточной биологии . 191 (2): 237–248. дои : 10.1083/jcb.201006074 . ПМЦ 2958478 . ПМИД 20956378 .
- ^ Макферсон, Дуэйн (18 июня 2018 г.). «Сенсорные волосковые клетки: введение в структуру и физиологию» . Интегративная и сравнительная биология . 58 (2): 282–300. дои : 10.1093/icb/icy064 . ПМК 6104712 . ПМИД 29917041 .
- ^ Шлоссер, Герхард (1 июня 2018 г.). «Краткая история почти всех чувств – эволюционная история типов сенсорных клеток позвоночных» . Интегративная и сравнительная биология . 58 (2): 301–316. дои : 10.1093/icb/icy024 . ПМИД 29741623 .
- ^ Надол, Джозеф Б. (1993). «Потеря слуха». Медицинский журнал Новой Англии . 329 (15): 1092–1102. дои : 10.1056/nejm199310073291507 . ПМИД 8371732 .
- ^ Лаш, Марк Э.; Пиотровский, Татьяна (2013). «Регенерация сенсорных волосковых клеток в боковой линии рыбок данио» . Динамика развития . 243 (10): 1187–1202. дои : 10.1002/dvdy.24167 . ПМК 4177345 . ПМИД 25045019 .
- ^ Перейти обратно: а б с Котанш, Дуглас А. (1994). «Регенерация волосковых клеток в улитке птиц после повреждения шумом или ототоксическими лекарствами». Анатомия и эмбриология . 189 (1): 1–18. дои : 10.1007/bf00193125 . ПМИД 8192233 . S2CID 25619337 .
- ^ Реми Пужоль, Режис Нувиан, Марк Ленуар, « Волосковые клетки (cochlea.eu)
- ^ Эшмор, Джонатан Феликс (1987). «Быстрая подвижная реакция в наружных волосковых клетках морской свинки: клеточная основа улиткового усилителя» . Журнал физиологии . 388 (1): 323–347. doi : 10.1113/jphysicalol.1987.sp016617 . ISSN 1469-7793 . ПМЦ 1192551 . ПМИД 3656195 .
- ^ Эшмор, Джонатан (2008). «Подвижность наружных волосковых клеток улитки». Физиологические обзоры . 88 (1): 173–210. doi : 10.1152/physrev.00044.2006 . ISSN 0031-9333 . ПМИД 18195086 . S2CID 17722638 .
- ^ Мюллер, У (октябрь 2008 г.). «Кадгерины и механотрансдукция волосковыми клетками» . Современное мнение в области клеточной биологии . 20 (5): 557–566. дои : 10.1016/j.ceb.2008.06.004 . ПМЦ 2692626 . ПМИД 18619539 .
- ^ Чан Д.К., Хадспет А.Дж. (февраль 2005 г.). «Нелинейная амплификация, управляемая током Ca2+, в улитке млекопитающих in vitro» . Природа Нейронауки . 8 (2): 149–155. дои : 10.1038/nn1385 . ПМК 2151387 . ПМИД 15643426 .
- ^ Макферсон, Дуэйн Р. (01 августа 2018 г.). «Сенсорные волосковые клетки: введение в структуру и физиологию» . Интегративная и сравнительная биология . 58 (2): 282–300. дои : 10.1093/icb/icy064 . ISSN 1540-7063 . ПМК 6104712 . ПМИД 29917041 .
- ^ Перейти обратно: а б Браунелл В.Е., Бадер Ч.Р., Бертран Д., де Рибопьер Ю. (11 января 1985 г.). «Вызванные механические реакции изолированных наружных волосковых клеток улитки». Наука . 227 (4683): 194–196. Бибкод : 1985Sci...227..194B . дои : 10.1126/science.3966153 . ПМИД 3966153 .
- ↑ Видеоклип, показывающий, как изолированная внешняя волосковая клетка движется в ответ на электрическую стимуляцию, можно посмотреть здесь (physical.ox.ac.uk). Архивировано 7 марта 2012 г. в Wayback Machine.
- ^ Гелеок Г.С., Холт-младший (2003). «Слуховое усиление: внешние волосковые клетки решают проблему» . Тенденции нейробиологии . 26 (3): 115–117. дои : 10.1016/S0166-2236(03)00030-4 . ПМЦ 2724262 . ПМИД 12591210 .
- ^ Варцог Д., Кеттен Д.Р. (1999). «Сенсорные системы морских млекопитающих» (PDF) . В Рейнольдсе Дж., Роммеле С. (ред.). Биология морских млекопитающих . Издательство Смитсоновского института . п. 132. S2CID 48867300 . Архивировано из оригинала (PDF) 19 сентября 2018 г.
- ^ Хадспет Эй Джей (28 августа 2008 г.). «Прилагая усилие, чтобы слушать: механическое усиление в ухе» . Нейрон . 59 (4): 530–545. дои : 10.1016/j.neuron.2008.07.012 . ПМЦ 2724262 . ПМИД 18760690 .
- ^ Сантос-Сакки Джозеф; Сун Лэй; Чжэн Цзефу; Наттолл Альфред Л. (12 апреля 2006 г.). «Контроль кохлеарной амплификации млекопитающих с помощью хлорид-анионов» . Журнал неврологии . 26 (15): 3992–3998. doi : 10.1523/JNEUROSCI.4548-05.2006 . ПМК 6673883 . ПМИД 16611815 .
- ^ Перейти обратно: а б с д Гиллеспи, PG; Сир, JL (2004). «Миозин-1c, двигатель адаптации волосковых клеток». Ежегодный обзор физиологии . 66 : 521–545. doi : 10.1146/annurev.phyol.66.032102.112842 . ПМИД 14977412 .
- ^ Перейти обратно: а б Стауффер, Э.А.; Холт, младший (2007). «Сенсорная трансдукция и адаптация во внутренних и внешних волосковых клетках слуховой системы мыши» . Журнал нейрофизиологии . 98 (6): 3360–3369. дои : 10.1152/jn.00914.2007 . ПМЦ 2647849 . ПМИД 17942617 .
- ^ Сир, Дж.Л.; Дюмон, РА; Гиллеспи, П.Г. (2002). «Миозин-1c взаимодействует с рецепторами волосковых клеток через свои IQ-домены, связывающие кальмодулин» . Журнал неврологии . 22 (7): 2487–2495. doi : 10.1523/JNEUROSCI.22-07-02487.2002 . ПМК 6758312 . ПМИД 11923413 .
- ^ Хаусли, Джорджия; Эшмор, Дж. Ф. (1992). «Ионные токи наружных волосковых клеток, выделенных из улитки морской свинки» . Журнал физиологии . 448 (1): 73–98. doi : 10.1113/jphysical.1992.sp019030 . ISSN 1469-7793 . ПМК 1176188 . ПМИД 1593487 .
- ^ «Чепной нерв VIII. Вестибулокохлеарный нерв» . Меддин . Университет Лойолы в Чикаго . Проверено 4 июня 2008 г.
- ^ Эдж А.С., Чен Цзы (2008). «Регенерация волосковых клеток» . Современное мнение в нейробиологии . 18 (4): 377–382. дои : 10.1016/j.conb.2008.10.001 . ПМЦ 5653255 . ПМИД 18929656 .
- ^ Ломбарте А., Ян Х.И., Поппер А.Н., Чанг Дж.С., Платт С. (январь 1993 г.). «Повреждение и регенерация пучков волосковых клеток в ушах рыб после лечения гентамицином». Слышать. Рез . 64 (2): 166–174. дои : 10.1016/0378-5955(93)90002-i . ПМИД 8432687 . S2CID 4766481 .
- ^ Уитфилд, ТТ (2002). «Рыба данио как модель слуха и глухоты» . Журнал нейробиологии . 53 (2): 157–171. дои : 10.1002/neu.10123 . ПМИД 12382273 .
- ^ Хендерсон М (15 января 2005 г.). «Ген, который больше не может оставаться глухим к старости». Таймс онлайн .
- ^ Сейдж, Кирилл; Хуан, Минцянь; Воллрат, Мелисса А.; Браун, М. Кристиан; Хиндс, Филип В.; Кори, Дэвид П.; Веттер, Дуглас Э.; Чжэн-И, Чен (2005). «Основная роль белка ретинобластомы в развитии и слухе волосковых клеток млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (19): 7345–7350. Бибкод : 2006PNAS..103.7345S . дои : 10.1073/pnas.0510631103 . ПМК 1450112 . ПМИД 16648263 .
- ^ Рафаэль Ю., Мартин Д.М. (июль 2005 г.). «Глухота: отсутствие регулирования способствует росту волосковых клеток» . Джин Тер . 12 (13): 1021–1022. дои : 10.1038/sj.gt.3302523 . ПМИД 19202631 . S2CID 28974038 .
- ^ Лу, На; Чен, Ян; Ван, Чжэнминь; Чен, Гуолин; Линь, Цинь; Чен, Чжэн-И; Ли, Huawei (2013). «Sonic hedgehog инициирует регенерацию волосковых клеток улитки посредством подавления белка ретинобластомы» . Связь с биохимическими и биофизическими исследованиями . 430 (2). Эльзевир: 700–705. дои : 10.1016/j.bbrc.2012.11.088 . ПМЦ 3579567 . ПМИД 23211596 .
- ^ Эрни, Сильвия Т.; Гилл, Джон К.; Палаферри, Карлотта; Фернандес, Габриэлла; Бури, Мишель; Лазаридес, Кэтрин; Гранжирар, Дени; Эдж, Альберт С.Б.; Лейб, Стивен Л.; Роччио, Марта (13 августа 2021 г.). «Поколение волосковых клеток в моделях кохлеарной культуры, опосредованное новыми ингибиторами γ-секретазы» . Границы клеточной биологии и биологии развития . 9 . Frontiers Media SA: 710159. doi : 10.3389/fcell.2021.710159 . ISSN 2296-634X . ПМЦ 8414802 . ПМИД 34485296 .
- ^ Самараджива, Аншула; Жак, Бонни Э.; Дабдуб, Ален (8 мая 2019 г.). «Терапевтический потенциал передачи сигналов Wnt и Notch и эпигенетической регуляции в регенерации сенсорных волосковых клеток млекопитающих» . Молекулярная терапия . 27 (5). Эльзевир Б.В.: 904–911. дои : 10.1016/j.ymthe.2019.03.017 . ISSN 1525-0016 . ПМК 6520458 . ПМИД 30982678 .
- ^ Гарсиа-Аньоверос, Хайме; Клэнси, Джон К.; Фу, Чуан Чжи; Гарсиа-Гомес, Игнасио; Чжоу, Инцзе; Хомма, Казуаки; Читэм, Мэри Энн; Дагган, Энн (04 мая 2022 г.). «Tbx2 является главным регулятором дифференциации внутренних и внешних волосковых клеток» . Природа . 605 (7909): 298–303. Бибкод : 2022Natur.605..298G . дои : 10.1038/s41586-022-04668-3 . ISSN 1476-4687 . ПМЦ 9803360 . ПМИД 35508658 .
- ^ Пол, Марла (04 мая 2022 г.). «Новый инструмент для создания клеток слуха, утраченных с возрастом» . Центр новостей Северо-Западной медицины . Проверено 11 мая 2022 г.
- ^ Хэндсли-Дэвис, Матильда (5 мая 2022 г.). «Генетическое открытие может помочь ученым обратить вспять потерю слуха» . Космос . Королевский институт Австралии . Проверено 11 мая 2022 г.
- ^ Левенхайм Х., Фернесс Д.Н., Кил Дж., Зинн С., Гюльтиг К., Феро М.Л., Фрост Д., Гаммер А.В., Робертс Дж.М., Рубель Э.В., Хакни К.М., Зеннер Х.П. (30 марта 1999 г.). «Нарушение гена p27(Kip1) обеспечивает пролиферацию клеток в кортизоновом органе постнатального и взрослого человека» . Proc Natl Acad Sci США . 96 (7): 4084–4088. Бибкод : 1999PNAS...96.4084L . дои : 10.1073/pnas.96.7.4084 . ПМК 22424 . ПМИД 10097167 . (основной источник)
- ^ Оно К., Накагава Т., Кодзима К., Мацумото М., Каваучи Т., Хосино М., Ито Дж. (декабрь 2009 г.). «Замалчивание p27 меняет постмитотическое состояние поддерживающих клеток в улитках неонатальных мышей» (PDF) . Мол клеточные нейроны . 42 (4): 391–398. дои : 10.1016/j.mcn.2009.08.011 . hdl : 2433/87734 . ПМИД 19733668 . S2CID 206831997 .
Библиография
[ редактировать ]- Гроб А., Келли М., Мэнли Г.А., Поппер А.Н. (2004). «Эволюция сенсорных волосковых клеток». В Мэнли и др. (ред.). Эволюция слуховой системы позвоночных . стр. 55–94.
- Феттиплейс Р., Хакни СМ (2006). «Сенсорная и моторная роль слуховых волосковых клеток». Обзоры природы. Нейронаука . 7 (1): 19–29. дои : 10.1038/nrn1828 . ПМИД 16371947 . S2CID 10155096 .
- Кандель Э.Р. , Шварц Дж.Х., Джесселл Т.М. (2000). Принципы нейронауки (4-е изд.). Нью-Йорк: МакГроу-Хилл. стр. 590–594 . ISBN 0-8385-7701-6 .
- Мэнли Г.А., Поппер А.Н., Фэй Р.Р. (2004). Эволюция слуховой системы позвоночных . Нью-Йорк: Springer-Verlag. ISBN 0-387-21093-8 .
- Мэнли Джорджия (2004). «Достижения и перспективы в изучении эволюции слуховой системы позвоночных». В Мэнли и др. (ред.). Эволюция слуховой системы позвоночных . стр. 360–368.
- Рэббитт Р.Д., Бойл Р., Хайстейн С.М. (1–5 февраля 2010 г.). «Механическая амплификация волосковыми клетками полукружных каналов» . Труды Национальной академии наук . 107 (8): 3864–3869. Бибкод : 2010PNAS..107.3864R . дои : 10.1073/pnas.0906765107 . ПМК 2840494 . ПМИД 20133682 .
- «Встроенные усилители: как легкие движения головы и тихие звуки передаются мозгу» . Медицинский Экспресс . 9 февраля 2010 г.
- Бренеман К.Д., Браунелл В.Е., Рэббитт Р.Д. (22 апреля 2009 г.). Брезина В. (ред.). «Пучки волосковых клеток: флексоэлектрические моторы внутреннего уха» . ПЛОС ОДИН . 4 (4): е5201. Бибкод : 2009PLoSO...4.5201B . дои : 10.1371/journal.pone.0005201 . ПМЦ 2668172 . ПМИД 19384413 .
- «Усилитель руля для вашего слуха: в ушах есть крошечные «флексоэлектрические» моторы для усиления звука» . Phys.org (пресс-релиз). 22 апреля 2009 г.
Внешние ссылки
[ редактировать ]- Молекулярная основа слуха
- Наружные волосковые клетки танцуют «рок круглосуточно»
- Видео о танцах OHC Yale Ear Lab
- Поиск NIF - волосковые клетки , заархивировано 3 марта 2016 г. в Wayback Machine через информационную структуру нейронауки.
- Hair-Tuning-Sound-Sensor. Архивировано 26 августа 2021 г. в Wayback Machine. Краткий отчет о недавней разработке звуковых датчиков на основе настройки волос студентами SMMEE, IIT Ropar.