Горгонопсия
| Горгонопсианцы Временный диапазон: средний перми - поздний пермский ,
| |
|---|---|

| |
| Скелет в космоне Александри в Музее наук , Тренто, Италия | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Клада : | Synapsida |
| Клада : | Терапсида |
| Клада : | Theroidontia |
| Клада : | † Горгонопсия Сили , 1895 |
| Роды | |
Горгонопсия (от греческого горгона , мифологического зверя и аспекта « óps »)-это вымерший клад терапсидов саблезубанных , от середины до верхнего пермца примерно между 265 и 252 миллионами лет назад. Они характеризуются длинным и узким черепом, а также удлиненным верхним, а иногда и нижним собачьим зубами и резцами , которые, вероятно, использовались в качестве рубки и удара оружия. Постканиновые зубы обычно уменьшаются или отсутствуют. Для охоты на большую добычу, они, возможно, использовали тактику укуса и восстановления, ссаживались за засадами и избавленным от цели и следуя за ней на безопасном расстоянии, прежде чем его травмы исчерпали его, после чего горгонопский снял бы животное и доставит Убивая укус. У них был бы непомерный жезл, возможно, превышающий 90 °, без необходимости расстроить челюсть.
Они заметно увеличились в размере с течением времени, растущая с небольших длины черепа 10–15 см (4–6 дюймов) в среднем пермском до пермских до 60 см (2 фута) в верхнем пермском. Последние горгонопсианцы, Rubidgeinae , были самыми надежными из группы и могли производить особенно мощные укусы. Считается, что горгонопсийцы были полностью наземными и могли ходить с полуэлексической походкой, с аналогичной наземной локомоторией, как и современные крокодильцы . Возможно, они были более гибкими, чем их добыча, но, вероятно, были инерционными гомеотермами, а не эндотермами, в отличие от современных терасефалов и цинодонтов , и, таким образом, были сравнительно менее активными. Хотя горгонопсийцы смогли поддерживать довольно высокую температуру тела, неясно, были бы у них также пот или мех (и, по ура, и связанные с ними структуры). Их мозг напоминал современные мозги рептилий , а не мозги живых млекопитающих . Большинство видов могли быть преимущественно Суточная (активная в течение дня), хотя некоторые могли быть сумеречными (активными на рассвете или в сумерках) или ночной (активной ночью). Считается, что у них было бинокулярное зрение , париетальный глаз (который обнаруживает солнечный свет и сохраняет циркадный ритм ), острое обоняние, функциональный рвомероназальный орган («орган Джейкобсона») и, возможно, рудиментарная барабанная собой .
Основные группы TherapsID все развивались к 275 миллионам лет назад от предка « пеликозавра » (плохо определенная группа, включая все синапсиды , которые не являются терапидами). Поглощение TherapsId от пеликозавров произошло у среднего пермца, поскольку мир постепенно стал более сухим. Горгонопсианцы выросли, чтобы стать апелляционными хищниками своей среды после события массового вымирания капитан , которое убило диномефальцев и некоторых крупных тероцефалов после среднего пермца. Несмотря на существование единого континента во время Пермского, Пангея , горгонопсийцы были обнаружены только в супергруппе Кару (в первую очередь в Южной Африке, а также в Танзании, Замбии и Малави), формировании Моради Нигера, Западной России и в Бассейн Турпана в Синьцзяне , Китай , с вероятными останками, известными из формирования Кундарама в бассейне Пранхита -Годавари в Индии . Эти места были полузасушливыми районами с очень сезонным количеством осадков. Горгонопсские роды очень похожи по внешнему виду, и, следовательно, многие виды были названы на основе хрупких и вероятных возрастных различий с момента их открытия в конце 19-го века, и группа подвергалась нескольким таксономическим изменениям.
Большинство горгонопсийцев вымерли во время фазы пермского происхождения, происходящего в самом конце перми, в котором основная вулканическая активность (которая будет производить сибирские ловушки ) и результирующий массивный всплеск в парниковых газах вызвали быстро Спайк, кислотный дождь , частые лесные пожары и потенциальная разбивка озонового слоя . Тем не менее, некоторые меньшие таксоны, такие как Cyonosaurus, могли выжить до раннего триаса . Большие хищнические ниши будут захвачены архозаврами ( а именно крокодильцами и динозаврами ) в мезозое .
Описание
[ редактировать ]
Ранее Gorgonopsids у среднего пермца были довольно маленькими, с длиной черепа 10–15 см (4–6 дюймов), [ 1 ] Принимая во внимание, что некоторые более поздние роды достигли массивных медвежьих, похожих на размеры с наибольшим из которых претендент до 3,5 м (11 футов) в длину и 300 кг (660 фунтов) в массе тела. [ 2 ] Тем не менее, мелкие горгонопсийцы оставались в изобилии до исчезновения (хотя мелкие виды могут фактически представлять юношеские образцы других таксонов). [ 1 ]
Как и другие пермские терапсиды, горгонопсийцы разработали несколько характеристик млекопитающих. Они могли включать парасагиттальную походку (конечности были вертикально ориентированы и перемещены параллельно позвоночнику), в отличие от обширной походки амфибиев и более ранних синапсидов. Это изменение походки в терапсидах было, возможно, было связано с уменьшением размера хвоста и фалангиальной формулы [ 3 ] (Количество костей на цифру, которое для Gorgonopsians было 2,3.4.5.3, как рептилии [ 4 ] ) Другие разработки включали волокнистую пластинную кожу коры и глубоко накрытые зубы. [ 3 ] Как и рептилии, у горгонопсийцев не хватает вторичного неба, отделяющего рот от полости носа, запрещая жевать.
Череп
[ редактировать ]Анатомия различается невероятно мало между горгонопсинами. [ 5 ] Многие виды отличаются расплывчатыми пропорциональными различиями, и, следовательно, более мелкие виды могут фактически представлять несовершеннолетних с более крупными таксонами. Примечательно, что Вомер на кончике морды варьируется между видами с точки зрения степени его расширения, а также положений, степени раскола и формы 3 гребней. [ 1 ] Обычно они имеют длинный и узкий череп. [ 4 ] У несовершеннолетнего Рубидгея , кажется, имели морду шире, чем долго. [ 6 ] В отличие от Euthryodonts, затылочная кость (в задней части черепа) является прямоугольной (более широкой, чем высокой) и вогнута, в отличие от треугольного. [ 7 ] : 279

Горгонопсский мозг, как и другие не млекопитающие терапсид , не имеет расширения неокортекса , имеет относительно большой задний мозг по сравнению с передним мозгом, большим эпифизиальным нервом (обнаруженного в существах с теменным глазом на верхней части головы), расширенный Гипофиз и общая удлиненная форма; Всего в общем, напоминающем мозг рептилии . [ 8 ] Мозговые ящики также были довольно рептилиями, а также сравнительно меньше и не такой толстый, как у млекопитающих. [ 7 ] : 280 Flocculus вестибуло , доля мозжечка , пропорционально большой и связан с -кокулярным рефлексом (который стабилизирует взгляд при перемещении головы). Судя по ориентации полукруглых каналов в ухе (которые должны быть ориентированы параллельно земле), голова горгонопсийского образца GPIT/RE/7124 будет наклоняться вперед примерно на 41 °, увеличив перекрытие между перекрытием между Визуальные поля двух глаз и улучшение бинокулярного зрения - используются для хищника. В отличие от рептилий или млекопитающих , полукруглые каналы плоские, вероятно, потому, что они были втиснуты между опистотической ( внутренней костью ушей ) и супраоципитальной костью. [ 8 ]
Зубы
[ редактировать ]
Как и многие млекопитающие, горгонопсийцы были гетеродонтами , с четко определенными резцами , собаками и зубами, гомологическими , гомологичными после премоляров и моляров . [ 1 ] У них было пять резцов в верхней челюсти (для большинства первые три были такого же размера, как и друг, и последние два были короче) и четыре на дне. [ 7 ] : 17–18
У большинства горгонопсийцев резцы были большими, а верхние клыки были удлинены в сабли, так же, как и у более поздних саб-зубчатых кошек . У некоторых горгонопсийцев были исключительно длинные верхние клыки, такие как Inostrancevia , и у некоторых из них был фланец на нижней челюсти, чтобы заручить кончик собаки, когда рот был закрыт. Сабли, как правило, интерпретируются как используемые как нанесение удара или срезание оружия, что потребовало бы чрезвычайно широкого защелки. Как верхние, так и нижние клыки Рубидгеи были удлинены, и животному нужно было бы еще больше. [ 9 ] Схема зубца горгонопсийцев была наиболее похожа на тероподные динозавры, чем с другими синапсидами. [ 10 ] Небо также включает в себя бугоральность и хребты, которые часто имеют функциональные зубы, которые, возможно, использовались для удержания борьбы с добычей, отвлекая эти мощные силы от хрупких клыков. Аналогичные хребты были идентифицированы на Мачародонта гомотериуме . [ 7 ] : 280 Постканиновые зубы были уменьшены как по размеру, так и по количеству; Многие Rubidgeines (последние горгонопсианцы) не имели постканинов в нижней челюсти, [ 11 ] И Клеландии не хватало их полностью. [ 6 ]
Горгонопсианцы были полифиодонтами , а зубы непрерывно росли на протяжении всей жизни человека. [ 12 ] Как и некоторые терапсиды, в то время как была одна функциональная собака, другая собака росла, чтобы заменить его, когда он неизбежно разбился. Левая и правая сторона челюстей не должна была быть синхронной, поэтому, например, первая собачья на левой стороне может быть функциональным, в то время как первая собака на правой стороне все еще рос. [ 12 ] Такой метод мог бы быть в игре, так что, как всегда иметь набор функциональных собак, поскольку наличие одного клыков или нет было бы серьезным препятствием для охоты, а выращивание таких больших зубов заняло много времени. С другой стороны, поскольку функциональный собачий обычно встречается в переднем зубном розетке (вместо равного возникновения в любом сокете), возможно, что замена собак произошла конечное количество раз, и животное в конечном итоге осталось одним , Постоянный набор функциональных собак в этих розетках. В 1984 году британские палеонтологи Дорис и Кеннет Кермак предположили, что клыки выросли, чтобы соответствовать размеру черепа, и постоянно отрывались до тех пор, пока животное не перестало расти, и что горгонопсианцы показывают раннюю версию конечных зубов, демонстрируемая у многих млекопитающих. Схемы замены зубов других зубов неясны. [ 9 ] Постканиновые зубы были заменены медленнее, чем другие зубы, вероятно, из -за отсутствия функционального значения. [ 1 ]
Посткраниум
[ редактировать ]
Семь шейных позвонков (на шее) - все то же самое, что и друг, за исключением последнего, что короче и ниже; Есть один атлас и одна ось . Как и кошки с саблезубыми, шея длинная с хорошо развитыми мышцами, что было бы особенно полезны, когда клыки были погружены в животное. Как и другие ранние синапсиды, у горгонопсийцев есть один затылкий мыщел , а артикуляция (суставы) шейных позвонков в целом является рептильной, позволяющей двигаться от стороны в сторону головы, но ограничивая движение вверх и вниз. Последний шейный шейки в форме больше похожа на спинные позвонки . [ 7 ] : 291–293
Дорсалы в форме катушки, и все они появляются примерно так же, как и друг друга. Опистынные процессы круто вытянуты из центра и имеют острые кили спереди и сзади. В отличие от Euthryodonts, горгопсийцы не имеют выдающихся поясничных позвонков . Тем не менее, дорсалы, приравнивающие эту серию, похожи на поясничные поясничные кошки с саблезубыми кошками с круто ориентированными зигопофазами, полезными для стабилизации нижней части спины, особенно при прикреплении добычи. [ 7 ] : 293–295
Есть три сакральных позвонка , и сериал, прикрепленный к тазу первого позвонка. Таз - это рептилий, с разделенным подвздошным подвздошной кости , ишимом и лобком . Бедра плечевая слегка S и короткая, но длинная и тонкая, чем кость . Для большинства большеберцовая кость и малоберцовая кость сильно кричат друг в друга, а голени более надежны, чем малоберцовая кость. [ 7 ] : 295–299 Сустав между лодыжкой и костями пятки, возможно, был несколько мобильным. Пятая цифра как для рук, так и для ног не была прикреплена к карпу / лапку и вместо этого соединялась непосредственно к кости локтевой кости. [ 13 ]
Таксономия
[ редактировать ]Стоимость ископаемого подшипника
[ редактировать ]
В 1876 году первые горгонопсские останки были идентифицированы в группе Бофорта в супергруппы Кару Южной Африке, биологом и палеонтологом Ричардом Оуэном . Он классифицировал окаменелости как Gorgonops Torvus , объединив греческий Горгон , мифологический зверь, со словом óps ( ὄὄ ), что означает «аспект». [ 14 ] В Африке горгонопсианцы также были найдены в обнаженных показаниях Кару в долине Рухуху в Танзании, долине Верхней Луангва в Замбии и Чивете , Малави . [ 7 ] : 7
Горгонопсийцы были впервые идентифицированы в России в 1890 -х годах в местности Соколки на северной Двине в Сибири под надзором русского палеонтолога Владимира Прохоровича Амалицкого . В посмертной публикации она была описана как Inostrancevia Alexandri , и это один из самых известных и крупнейших горгонопсийцев. С тех пор было описано только несколько российских таксонов: Pravoslavlevia , Sauroctonus , Viatkogorgon , Sugogorgon и Nochnitsa . [ 15 ] Одно из рода, названных русскими родами, Леогоргон , теперь считается номенем Дубий . [ 6 ]
Горгонопсии явно отсутствуют за пределами этих двух областей. [ 15 ] В 1979 году китайский палеонтолог Ян Чжунцзян описал китайский горгонопский « wangwusaurus tayuensis » на основе зубов из позднего пермского образования джиюань , [ 16 ] Но в 1981 году палеонтологи Дениз Сигогно-Русселл и AI-Lin Sun обнаружили, что назначенный материал является случайным совокупностью, из которых только два имеют даже отдаленное сходство с горгонопсией. [ 17 ] В 2011 году был обнаружен неполный набор зубов в местности в депрессии Турпана , Синьцзян , на северо -западе страны. Окаменечный материал, хотя и тонкий, описывается в 2022 году палеонтологами Джун Лю и Ван Ян и подтверждает, что он исходит от горгонопсийского свидания от верхнего пермца, который фактически жил в современном Китае. [ 18 ] В 2003 году индийские палеонтологи Сангхамитра Рэй и Сасвати Бандиопадхьяй назначили несколько фрагментов черепа от позднего пермского образования Кундарам для среднего размера горгонопсиана, [ 19 ] Хотя горгонопсские характеристики также были задокументированы у некоторых тероцефалов. [ 15 ] В 2008 году был идентифицирован большой фрагмент верхней челюсти и собаки рубиджина и собака в позднем пермском моради в Нигере (одно из немногих низкоистогот-пермских, несущих тетрапод, и является первым доказательством низколостных горгопсовцев. Полем [ 20 ]
Классификация
[ редактировать ]
После открытия Оуэн предположил, что Gorgonops и несколько других таксонов, которые он описал в супергруппе Кару, были хладнокровными рептилиями, несмотря на то, что они носили зубы, напоминающие зубы, плотоядных млекопитающих. Он предложил классифицировать их всех в рамках недавно приуманенного приказа Theroidontia (который он поместил в классную рептилия). Он решил подразделять Theroidontia в семьи , основанные на анатомии ноздрей (кости нариалс) - «Mononarialia» для тех, у кого есть одно открытие в черепе для носа, как у млекопитающих, «Binarialia» для тех, у кого два отверстия, как в рептилетах, как у млекопитающих, «Binarialia» для тех, у кого два отверстия, как в рептилетах, как у млекопитающих, «Binarialia». и "Tectinarialia" для Gorgonops , потому что его отверстие было омрачено толстой костяной крышей [ 14 ] ( Tectus является латинским для «покрытого, крытого, украшенного»). [ 21 ] В 1890 году английский натуралист Ричард Лидеккер сделал Gorgonops типовым видом семейства Gorgonopsidae. [ B ] [ 22 ] Британский палеонтолог Гарри Сили в 1895 году считал, что у Горгонопса не было отверстия во временной кости (височная фенестра), которая является диагностической особенностью Theroidontia, и, таким образом, повышенным Gorgonopsidae до Gorgonopsia, отличной от Theroidontia. Он классифицировал все южноафриканские материалы с чертами рептилий, так и млекопитающих в порядке «Териоохия», и считал подчинения Горгонопсии и Териодонтии. его [ 23 ] Американский палеонтолог Генри Фэйрфилд Осборн полностью переработал классификацию рептилии в 1903 году и построил две основные группы: Diapsida и Synapsida , [ 24 ] А в 1905 году южноафриканский палеонтолог Роберт Брум создал третью группу Therapsida , чтобы разместить «рептилии, похожие на млекопитающие», включая Theriodontia. Он также бросил вызов требованиям Сили и отменил Горгонопс обратно в Терадонтию, но он поместил его в свою недавно возведенную подгруппную тероцефалию , растворяя горгонопсию. [ 25 ] В 1913 году, особенно в свете почти полного черепа Дж. Торвуса, обнаруженного преподобным Джоном Х. Варсом, метла восстановила Горгонопсию. [ 26 ]

Число южноафриканских родов быстро росло в 20-м веке, возглавляемое главным образом Брумом, чья обширная работа над кару-терапсидами-от начала его карьеры в стране в 1897 году до его смерти в 1951 году-под его описанием 57 Горгонопсские образцы голотипа и 29 родов. Многие из таксонов Брулы позже будут признаны недействительными. [ 27 ] Многие другие современные работники создали совершенно новые виды или роды на основе отдельных образцов. [ 7 ] : 57 Следовательно, Gorgonopsia была предметом большой таксономической суматохи и является одной из самых проблемных синапсид. Поскольку анатомия черепа отличается очень мало по всем таксонам, многие определяются на основе расплывчатых пропорциональных различий, включая даже известных членов. Номинальные виды различаются преимущественно по признакам, которые, как известно, являются весьма переменными в зависимости от возраста человека, включая размер орбиты глаз , длину морды и количество почтовых зубов. Таким образом, возможно, что некоторые таксоны являются синонимом друг друга и представляют собой разные стадии развития. [ 28 ]
Среди первых попыток организовать кладу были выполнены британским зоологом Дэвидом Мередитом Сиаром Уотсона и американским палеонтологом Альфредом Ромером в 1956 году, которые разделили его на двадцать семей, из которых три (Burnetiidae, Hipposauridae и Phthinosuchae) не рассматриваются Горгонопсианцы больше. [ 6 ] В 1970 году и снова в 1989 году, в основном рассматривая африканские таксоны, Sigogneau-Russell опубликовал комплексную монографию о Gorgonopsia (определяя его как недостаток ) и признал только две семьи: Watongiidae и Gorgonopidae. [ 29 ] Ватонгия была перенесена в Варанопиды в 2004 году канадскими палеонтологами Робертом Р. Рейшом и Мишелем Лаурином . [ 30 ] Sigogneau-Russell Split Gorgonopidae на три подсемейства-Gorgonopsinae, Rubidgeinae и Inostranceviinae-и уменьшили количество родов до двадцати трех. [ 29 ] В 2002 году российские таксоны проводятся подрядкой Горгонопсией . Русская (Phyridgeoida) была поверхностной Gorgonopioida . [ 31 ] В 2007 году биолог Ева VI Gebauer, в своем всестороннем обзоре Gorgonopsia (ее диссертация PhD), отвергла модель Ивахненко в пользу Sigogneau-Russell's, [ 7 ] : 57 Arcthonnathus Aelugonhathus , , , Eoarchops Saurccognadus , " Gorgonps Cyopadus , , " , Dexeya " nasuta (under the informal nomen nudum "Njalila"), Lycaenops, Arctognathus, Aelurognathus, Sycosurus , Clelanda и Rubidge [ 7 ] : 244 В целом, модель Сигогно-Русселла поддерживается, но существует мало консенсуса, по которому можно назначить роды, на которые можно назначить подсемейства. [ 6 ] В 2015 году американский палеонтолог Кристиан Ф. Каммерер и коллеги переписали эрифхостому (который был помечен как неопределенный Theriodont) как горгонопский [ 32 ] и потопл Scylacognathus и в следующем году в это. [ 6 ]

Первая филогения (семейное древо) членов горгонопсии была опубликована в 2016 году американским палеонтологом Кристианом Ф. Каммером, который специально исследовал Rubidgeinae и переписывал как подсемейство, так и девять видов, которые он назначил (уменьшив число из Тридцати. -Сикс виды). Kammerer также воскресил Dinogorgon , Leontosaurus , Ruhuhucerberus и Smilesaurus . Каммерер не был уверен, представляют ли Леонтозавр , Клеландина , Диногоргон и Рубидгея один и тот же таксон или нет (для которого Диногоргон имеет приоритет ), но он решил классифицировать их все в племени Рубиджини в ожидании дальнейшего изучения. [ 6 ] В 2018 году Каммерер и российский палеонтолог Владимир Масиутин определил нового рода Nochnitsa как наиболее известный базал -горгонопсианцы и обнаружили, что все российские таксоны (кроме Viatkogorgon , который находится в переоборудовании) образуют совершенно отдельный клад от африканских таксонов. [ 15 ] Также в 2018 году палеобиолог Ева-Мария Бендель, Каммерер и его коллеги воскрешали Cynariops . [ 1 ] В 2022 году Каммерер и коллега -палеонтолог Брюс С. Рубидж описали Phorcys из Южной Африки. [ 33 ]
| |||||||||||||||||||||
| Филогения горгонопсии в соответствии с Каммеррером и Рубиджем (2022) [ 33 ] |
Эволюция
[ редактировать ]Synapsida традиционно разделяется на базальную « Пеликозаврию » и полученную Therapsida. Первый включает в себя хладнокровные существа с обширной походкой и, по-видимому, более низким метаболизмом, который развивался в верхнем углероде . В середине до конца 20-го века американский палеонтолог Эверетт С. Олсон исследовал синапсидное разнообразие в средних пермских формациях Сан-Анджело , Цветочника и Чикаша в Северной Америке и отметил, что разнообразие пеликозавра сокращается с шести до трех в этих формациях и что Они сосуществовали с несколькими фрагментарными образцами, которые он интерпретировал как терапиды. Затем он предложил адаптивный переход от класса пеликозавра к терапидному классу произошел во время среднего перми ( вымирание Олсона ); Тем не менее, классификация этих «терапидов» и возраста образования с тех пор была оспорена. Таким образом, точное время поглощения TherapsId неясно, но шесть основных групп TherapsID ( Biarmosuchia , Dinocephalia , Anomodontia , Gorgonopsia, Throcephalia и Cynodontia) эволюционировали к 265 миллионам лет назад во время Вордиан . [ 34 ] Самый старый горгонопский образец - это частичная морда, принадлежащая неопределяемому роду из эодицинодон зоны сборки в бассейне Кару, примерно знакомства с Фордианом. Phorcys, Karoo Tapinocephalus известный с самого нижнего конца зоны сборки , примерно немного позже до границы Ворджина/Капитан, является самым старым идентифицируемым горгонопсовым таксоном. [ 33 ]
| Synapsida [ 34 ] |

Перми постепенно стал сушилкой и сушилкой. В верхней части каменноугольной и нижней перми, пеликозавры, по -видимому, цеплялись за средние обитания на угольном болоте возле экватора (окаменелости, известные в пределах 10 ° с обеих сторон палеоеватора); За этим примерно до 30 ° была обширная пустыня, которая простиралась до побережья, отделяя болота от умеренных областей. По среднему перми, экваториальные леса перешли на сезонную влажную/сухой систему, но болота были связаны с умеренными зонами через прибрежные проходы вдоль Восточной Пангеи, что позволило перекрестно-континентальную миграцию из того, что сейчас является Южной Африкой на то, что сейчас является Россией. Полем Терапсиды, по -видимому, развивались в этом сезонно влажном ландшафте, расширяясь даже в умеренные зоны. На этом этапе синапсиды были единственными крупными наземными животными в окружающей среде; и пеликозавры, возможно, не смогли адаптироваться к аридификации. Примерно во время вымирания пеликозавра, TherapsIds испытал серьезную адаптацию Радиация (все плотоядные) продолжается в верхнем пермском. [ 34 ]
На протяжении всего среднего пермья часто гигантские диномефальцы были доминирующими животными их экосистем. Они исчезают из ископаемых записей во время события массового вымирания капитан, вызванного вулканической деятельностью, которая сформировала китайские ловушки Эмейшана . Точная причина их вымирания неясна, но их заменили горгонопсийцы и дицинодонты (которые начали значительно увеличивать размеры) и меньших тероцефалов. [ 35 ] Рубиджцы были наиболее полученными горгонопсинами, и, следовательно, самыми массивными и сильно построенными. [ 6 ]
Палеобиология
[ редактировать ]Кусать
[ редактировать ]
Горгонопсианцы, вероятно, были активными хищниками. У Rubidgeines есть особенно надежный череп среди горгонопсин, сравнимых с огромными макропредаторами, которые используют свои черепа в качестве основного оружия, например, мозасавры или некоторые с теропод динозавры . Менее надежные горгонопсийцы с более длинными клыками и гораздо более слабым укусом, такими как Smilesaurus или Inostrancevia , вместо этого, вероятно, использовали свои собаки для рубки, гораздо более похожих на саб-зубчатых кошек. Постканины Клеланды были заменены гладким гребнем, в отличие от дицинодонтов, которые имеют кератиновый хребет, похожий на лезвия, и это могло быть преимущественно после добычи, он мог проглотить целое. Горгонопсские таксоны действительно сосуществовали друг с другом - как и семь лет одновременно - и тот факт, что некоторые рубиджины обладают постканинами, в то время как некоторые другие современные не предполагают, что они практиковали нишевые распределения и преследовали различные добычи. [ 6 ]
Считалось, что удлиненные клыки, как правило, сыграли важную роль в их тактике охоты. Горгонопсская шарнирная шарнир была двойной соединенной и состоит из несколько мобильных и вращающихся костей, что позволило бы им невероятно широко открывать рты - возможно, превышает 90 ° - без необходимости расстроить челюсть. [ 36 ] В качестве альтернативы было предложено (сначала в 2002 году биологами Блэр Ван Валькенбург и Тайсон Секко, хотя в связи с кошками), которые сабли развивали в первую очередь из -за сексуального отбора в качестве формы спаривания . Это демонстрируется у некоторых современных видов оленей, но трудно проверить, учитывая отсутствие живых саб-зубчатых синапсид-хищников. Считается, что у кошек с сабрезубыми, длинноноводными («дирк-зубцами») таксонами хищниками , тогда как считается, что таксоны с короткими зубами («Scimitar-Toothed») были засадными . [ 37 ] Предполагается, что среди кошек с дирк-зубцами эти хищники убили с хорошо расположенной чертой до горла после захватывания добычи, но горгонопсийцы могли быть менее точными с размещением укусов, вооруженными рептитными челюстями и аранжировками зубов. Вместо этого горгонопсийцы, возможно, использовали тактику укуса и восстановления: хищник засадил бы за упущенную засаду и займет из нее значительный и изнурительный укус, а затем следовать, как добыча пыталась сбежать, прежде чем поддаться травме, после чего горгонопский доставит убийственный укус. Поскольку постканины уменьшаются или полностью отсутствуют, мясо было бы насильственно оторванным от туши и проглочено целым. [ 36 ] Эта стратегия «прокола -пуль» также предполагается, что они использовались динозаврами Theropod. [ 10 ]

Горгонопсианцы, наряду с другими ранними плотоядными, а также крокодилами, в основном полагались на «кинетическую инерциальную систему» (KI), кусая добычу, в которой птеригоидные и височные мышцы быстро зажали челюсти, используя импульс и энергию кинетическую Челюсти и зубы, чтобы схватить жертву. Утолочные животные млекопитающих, в том числе саблезубые кошек, вместо этого полагаются в основном на «систему статического давления» (SP), где временные и масс-мышцы вызывают сильную силу укуса, чтобы убить добычу. [ 11 ] Второлис и массатер разделились только у млекопитающих, и у горгонопсийцев была мышца, простирающаяся от нижней стороны крыши черепа , обратно к сквомозной кости (в задней части черепа) и через скулы. Часть, закрепленная на щеках, стабилизировала челюстную конус и позволила ей перемещаться из стороны в сторону во время закрытия. Возможно, это было очень важно при укусе, так как скулы становятся сильнее в тандеме, когда собаки становятся длиннее. [ 7 ] : 278
Меньшие горгонопсийцы, такие как Cyonosaurus (который на самом деле может представлять несовершеннолетний другой вид), имели гранильные черепа и сабли, и, возможно, действовали так же, как шакалы и лисы . У более крупных горгонопсийцев, таких как Gorgonops , были давно надежные морды с сильно расклешенными щеками, которые поддерживали бы сильные птеригоиды и мощный укус KI. среднего размера У Arctognathus был ящик, похожий на коробку и, в результате, мощную морду, которая позволила бы сильные изгибы и кручение, а также сочетание элементов укуса KI и SP. Еще более крупные горгонопсийцы, такие как Arctops , имели более короткую и более выпуклую морду, как более ранний диметродон Спенкодонта , и смог бы быстро закрепить челюсти от широкого зазора (что было бы необходимо, учитывая длинные клыки). У еще более крупных Rubidgeinae были чрезвычайно мощные, сильно построенные, стимулированные черепа, с широкими мордами, сильно фланцевыми щеками и чрезвычайно длинными зубами; Сабли Рубидгея Атрокс длиннее зубов Тираннозавр . [ 11 ]
В отличие от плотоядных животных млекопитающих, горгонопсийцам (и тероцефальцам) уменьшалось или полностью не имело постканинов, и челюсть, вероятно, не смогла оказывать давление сдвига, необходимое для раздавливания костей, открытых для доступа к костному мозгу . [ 38 ] этого в сегодня не развивался ли костный мозг еще у пермских синапсидов ( рыбы и многим амфибиям не хватает В значительной степени было неясно , ) костный мозг в их конечностях. [ 39 ]
Локомоция
[ редактировать ]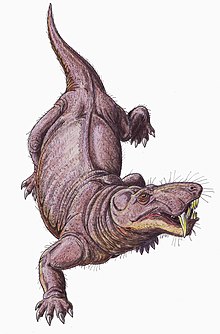
Считается, что горгонопсийцы были строго наземными . [ 40 ] Считается, что они смогли перемещаться с прямой походкой, подобной той, которая использовалась крокодильцами, конечности расположены почти вертикально, в отличие от горизонтали, как в обширной походке ящериц. Глоноидная полость на плечевом лопасти сильно наклоняется, поэтому конечности имели ограниченное движение вперед, и у них, возможно, была короткая длина шага. Ящерицы часто перемещают свои колючки в сторону, чтобы увеличить длину шага, но более вертикально ориентированные фасеточные суставы, соединяющие позвонки у горгонопсинцев, сделали бы позвоночник более жестким и стабильным, обременяя такое движение. [ 7 ] : 259–260
Горгонопский плечевой сустав имеет очень необычную конфигурацию. Голова плеча, которая соединяется с плечом, длиннее гленоида, поэтому она не может вписаться в полость. Следовательно, они, возможно, были прикреплены с большой массой хряща, с плечевой костью, выполняющим холмуе по гленоиду. Теоретически это может сделать угол между плечевой кости и гленоидом где -то от 80 до 145 °, когда они обращаются к животному. Если бы угол находился на нижнем конце, это было бы довольно твердое соединение, что позволило бы дельтоидам оказывать большую силу через переднюю конебу, например, когда прикололо добычу или удерживая тушу, когда срывая плоть. Если плечевая кость была расположена под более высоким углом, это могло бы позволить усилить вперед разгибания вперед и назад (вдоль длинной оси) и, следовательно, большую длину шага, полезную в атаке или в коротких погонах. Плечевое лезвие расширяется по бокам животного (выступает в поперечном направлении), также обеспечивая большое прикрепление для дельтоидов. Все Scapulohumeral мышцы имели сильно развитые привязанности, особенно дельтоиды. При расширении передних конечностей дельтоиды могли поднять переднюю сторону (передний край) плечевой кости, а мышца Coracobrachialis опустила заднюю сторону (задний край). При отказании передней передней части мышца грудной клетки могла толкать передний край, а мышца Subcapularis подняла задний край. [ 7 ] : 260–264
Связь таза имеет обычную конфигурацию соединения с шариком и сокетами . Несколько сплюснутая головка бедренной кости теоретически могла бы вписаться в тазобедренную гнездо под широким диапазоном углов. В 1982 году палеонтолог Том С. Кемп предположил, что ранние териоды, в том числе горгонопсийцы, могут поместить бедру под горизонтальным углом в обширной походке, так и более вертикальный угол в прямой походке. Он сравнил локомоторивые привычки этих существ с привычками крокодилов, которые используют обширную походку на короткие расстояния, но переключаются на прямое, работая или перемещаясь на более длительные расстояния. Хотя бедра образца GPIT/RE/7113, по -видимому, является анатомически промежуточным между диметродоном и млекопитающими - с подвздошной кости , расширяемой в направлении встречи, чем хвосты, и лобик несколько уменьшен - Puboischiofemoris рептилии, которые бегают от таза до бедренной кости), широко прикрепленные к нижней стороне лобкового лобка и ишия , что позволило бы ему произвести сильное Аддикационная сила (притягивание ног ближе к телу), полезная в обширной походке. Также возможно, что горгонопсийцы в первую очередь задействовали эту мышцу, во время борьбы с добычей. [ 7 ] : 264–270 Голоки относительно короткие по сравнению с бедром, что предполагает, что горгонопсийцы не были хорошо адаптированы для проведения больших расстояний. [ 7 ] : 298–299

Что касается того, как ноги были помещены на землю, горгонопсийцы являются единственными ранними терапидами, которые представляют этатаксонию (последняя цифра имеет наибольший вес), гомоподия (следы и отпечатки рук выглядят одинаково) полуспа и Ноги были помещены плоскими на землю). [ 41 ] Эти адаптации, возможно, сделали Gorgonopsians Swifter и более гибкими, чем их добыча. [ 36 ] У горгонопсийцев были довольно ловкие цифры, что свидетельствует о возможности захвата как для рук, так и для ног, возможно, для борьбы с добычей, которая борется с трудностями, чтобы предотвратить чрезмерную нагрузку, а также косвенное разрушение или разрушение, клыков, пока они погрузились в жертву. [ 13 ]
Чувства
[ редактировать ]
В отличие от эвриодонтов, но, как и некоторые эктотермические существа сегодня, все горгонопсийцы обладали шишковидным глазом на макушке головы, который используется для обнаружения дневного света (и, таким образом, оптимальной температурой для активности). Вполне возможно, что другие териодоны потеряли это из -за эволюции любой эндотермии, по сути фоточувствительных ганглиозных клеток сетчатки в глазах - в тандеме с потерей цветового зрения и переходом к ночной жизни - или оба. [ 42 ] Предполагается, что ночное поведение возникало у млекопитающих ( ночное узкое место ), но большой размер орбиты и наличие склеротических колец во многих ранних синапсидах, растянувшись накапливаемой на какол. Условия освещения развивались намного раньше. Основываясь на этих аспектах, образец SAM-PK-K10034, возможно, имел мезопическое зрение , а Cyonosaurus Scotopic или фотопейское зрение . [ 43 ] Диаметры склеротических колец для небольшого виаткогоргона пропорционально большие, с внутренним диаметром 1,5 см (0,6 дюйма) и наружным диаметром 2,3 см (0,9 дюйма) по сравнению с диаметром 2,8 см (1,1 дюйма) для орбиты) по сравнению с диаметром 2,8 см (1,1 дюйма) для орбиты (0,9 дюйма), по сравнению с диаметром 2,8 см (1,1 дюйма) для орбиты. Сам, что говорит о том, что это сделало преимущественно ночные экскурсии. [ 15 ] Среди горгопсийцев у Рубиджин Клеландии есть необычайно маленькие склеротические кольца, что указывает на фотопное зрение и было строго суточным ; Каммерер предположил, что нишевое разделение среди рубиджинов (поскольку в районе сосуществовало до семи различных таксонов), частично, приняли форму различных видов, которые были активны в разное время дня, но склеротические кольца только из Clelandina среди Эта подсемейство была идентифицирована, что делает эту гипотезу очень спекулятивной. [ 6 ]

Горгонопсианцы имеют довольно короткую носовую полость, например, пеликозавры, но в нем есть обильные продольные хребты за внутренними ноздрями (которые соединяют полость носа с горлом); Поскольку дыхательный воздух не проходил бы через них, они, как правило, интерпретируются как обонятельные турбинаты и давали бы горгонопсианцам довольно высоко развитый обоняние. [ 44 ] Горгонопсианцы обладали вомероназальным органом («орган Джейкобсона») - часть аксессуальной обонятельной системы , которая была бы помещена у основания носовой перегородки ; В отличие от дицинододонтов и тероцефалов, кажется, был канал, соединяющий орган с ртом, что указывает на то, что он был функциональным у горгопсинцев. [ 45 ]
Ранние териоды (включая горгонопсийцев), возможно, обладали барабанной трансляцией , в отличие от более ранних пеликозавров, о которых указывалось восстановление связи между квадратной костью (в шарнире челюсти) и птеригоидной костью (во вкусе ), позволяя квадрату независимо вибрировать в степень. Это могло позволить обнаружить воздушные звуки с низкой амплитудой менее 1 мм (0,039 дюйма), но барабанная перепонка была бы поддержана хрящом или связками вместо кости. Если правильно, то постдентарные кости (которые у ранних млекопитающих образуют кости среднего уха ) должны были бы стать отделенными от зубной кости (Jawbone); Горгонопсская ископаемая запись, по-видимому, указывает на постдентар-дентационное соединение, которое было уменьшено. Несмотря на то, что учитывая специализацию, необходимые для укуса, состояние изолирующего квадрата у горгонопсийцев можно альтернативно объяснить как стрептостически (вращающийся квадрат), чтобы расширить Gape, а не облегчить слух. [ 3 ]
Терморегуляция
[ редактировать ]Основной анатомический сдвиг произошел между более ранними пеликозаврами и терапидами, которые, как предполагается, были связаны с растущим метаболизмом и происхождением гомеотермии (поддержание высокой температуры тела). Эволюция вторичного неба и отделение рта от полости носа может обладать повышенной эффективностью вентиляции, связанной с высоким уровнем аэробной активности; У горгонопсий не было костинного вторичного неба, но, возможно, у нее была одна мягкая ткань. Тем не менее, вторичное небо могло бы вместо этого помочь в употреблении большого количества пищи одновременно, а не в вентиляции. Было постулирована реорганизация скелета (от обширного до паразагиттальной походки), как показать присутствие диафрагмы , и, таким образом, также усилила вентиляцию для аэробной активности; Но вместо этого это могло бы состоять в увеличении ускорения или ловкости, что не обязательно соответствует интенсивной аэробной активности, как у крокодилов. Фиброзной пластинчатой коры, которую имели все ранние терапсиды, указывают на увеличение скорости роста, но это не может быть связано с скоростью метаболизма. [ 46 ]
Современные большие рептилии естественным образом выделяют тепло тела с более медленной скоростью, чем меньшие, и считаются «инерционными гомеотермами», но они поддерживают низкую температуру тела 25–30 ° C (77–86 ° F). Если бы терапиды потребовались более высокую температуру тела 35–40 ° C (95–104 ° F), им нужно было бы либо эндотермы (генерируя собственное тепло тела), либо имели больший контроль над потерей тепла (то есть лучше гомеотермия). Парасагиттальная походка могла помочь последнему, так как она удерживала бы большую часть тела от земли, а также позволила бы кровь оставаться в животе, а не циркулировать через придатки, оба из которых уменьшили бы теплообмен на землю и стабилизировать температуру ядра . Снижение хвоста также уменьшило бы общую площадь поверхности животного, что еще больше минимизировало потерю тепла. [ 47 ] Среди терапсидов только эвриодонты (не горгонопсианцы) имеют дыхательные носовые турбинаты , которые помогают сохранить влагу, втягивая большие количества воздуха, и его эволюция обычно связана с началом показателей потребления кислорода и происхождения эндотермии. [ 44 ]
Если горгонопсийцы были инерционными гомеотермами, невозможно, чтобы у них были волосы. Мвина, как правило, пронизана отверстием (маленькие отверстия, которые поддаются кровеносным сосудам), что потенциально может указывать на существование свободной кожи (в отличие от чешуйчатых), волос, различных кожных желез (таких как потные железы ) и усы ; Тем не менее, некоторые рептилии представляют аналогичный паттерн отверстия, который вместо этого связан с развитием зубов, а не с кожей. [ 48 ]
Палеопатология
[ редактировать ]

Переднелатеральный аспект левого радиуса (кость предплечья) горгонопсийского образца NHCC LB396 представляет собой круговое костяное поражение с нерегулярными и радиальными шипами из коры, окруженной тонким слоем субперостеальной кости , которые быстро растут над одинокой костью вегетационный сезон. Это согласуется с периоститом , скорее всего, вытекает из субпериостальной гематомы . Это конкретное состояние, а также быстрые темпы роста более напоминают млекопитающих и динозавров, чем крокодильцы или ящериц . Среди ранних синапсидов единственная другая отмеченная патология - это остеомиелит в нескольких группах пеликозавра. [ 49 ]
Лабиальная (губа/щека) сторона корня зуба из функционального собачьего клыка RB382 представляет собой целых 8 поражений, кластеризующихся вдоль средней линии зуба, что напоминает миниатюрные зубы с мякотью , дентином и тонким эмалью . Они примерно круглые - с диаметрами, варьирующими от 0,3 до 3,9 мм (0,012–0,154 дюйма) - хотя они становятся менее круглыми в средней точке корня до прохождения шейки матки зуба . заболевания человека Это примерно согласуется с одонтомой , наиболее частым типом одонтогенной опухоли , которая ранее распространялась только на несколько миллионов лет назад в ископаемом отчете. В 255 миллионов лет RB382 представляет собой старейший известный случай одонтомы. [ 50 ]
У взрослая морда SAM-PK-11490 из неопределенного среднего пермского горгонопского вида имеет встроенный зуб от неопознаваемого животного. Кость разработала мозоль вокруг зуба, что указывает на его заживление, а человек пережил атаку. Это либо из хищника, а не биармосухинского , тероцефального или другого горгонопса - либо внутривидового укуса лица, как это обычно выступает в социальных хищниках, таких как крупные кошки или мониторинг ящериц, и это было предложено для нескольких вымерших промышленности, таких как теропод , водные рептилии и саблезубые кошки. Социальный укус предназначен для утверждения доминирования или облегчения размножения, и, если это правильно, предполагает, что, по крайней мере, некоторые средние пермирские горгонопсии были социальными плотоядными. Изначально зуб был упущен, поэтому неясно, насколько распространена эта патология на самом деле. [ 51 ]
Палеоэкология
[ редактировать ]Палеоэкола
[ редактировать ]
После исчезновения диномефальцев и (в Южной Африке) базальные тероцефальцы Scylacosauridae и Lycosuchidae , горгонопсинцы эволюционировали из малых и необычных форм в большие хищники вершины . [ C ] Через середину до верхнего перми в Южной Африке дицинодонты были наиболее распространенными животными, тогда как пареайавры Дельтавжация и Скутозавр были наиболее распространенными в российских образованиях с горгонопсии. [ 52 ] Во время верхней пермья южноафриканская группа Бофорта была полузасушливой холодной степи с большими сезонными ( эфемерными ) реками и поймами, истощающими источники воды, гораздо дальше на север в море Кару, с некоторыми случаями внезапных наводнений после внезапного, сильного дождя; [ 53 ] [ 54 ] Распределение карбонатов согласуется с современными отложениями калиша , которые образуются в климате со средней температурой 16–20 ° C (61–68 ° F) и 100–500 мм (3,9–19,7 дюйма) сезонных осадков. [ 54 ] Горгонопсово, несущая саларевская формация в Западной России, также, вероятно, была нанесена в полузасушливой среде с очень сезонными осадками и имела растения гигрофитов и галофитов в прибрежных районах, а также более устойчивые к засухе хвойные деревья на более высоких возвышениях. [ 55 ] Формирование Моради была засушливой пустыней, в которой в основном доминировали Captorhinid рептилия Moradisaurus и Pareiasaur Bunostegos . [ 20 ] Он имел вольтские хвойные деревья и сравнивался с внутренней пустыней Намиб или бассейном озера Эйр . [ 56 ]
Вымирание
[ редактировать ]Большинство горгонопсийцев вымерли в конце верхнего пермца во время пермского происхождения , которое было в первую очередь вызвано вулканизмом, который образовал сибирские ловушки . Результирующий массивный всплеск в парниковых газах вызвал быстрое засущество из -за: температурного всплеска (целых 8–10 ° C на экваторе со средними экваториальными температурами 32–35 ° C или 90–95 ° F, в начале триасы), кислотный дождь (с рН всего 2 или 3 во время извержения и 4 во всем мире, и последующая нехватка лесов в течение первых 10 миллионов лет триаса), частые лесные пожары (хотя они уже были довольно распространенными во всем пермском ) и потенциальное разрушение слоя озона (возможно, кратко увеличивая бомбардировку ультрафиолетового излучения на 400% на экваторе и 5000% на полюсах). [ 57 ] Возможный образец Cyonosaurus предполагает, что некоторые меньшие таксоны, возможно, выжили до последнего перми или раннего триаса . [ 58 ] Среди терапсидов мелким терасефальцам и крупным травоядным аномодонтам сумели пересечь пермс-триассическую границу и выжили соответственно до среднего и верхнего триаса , но только мелкие виды цинодонтов выжили в юридическую болезнь, потомки, потомки, потомки будут включать млекопитающие. Нишские горгонопсианцы , оставленные открытыми, были в конечном итоге заполнены архозаврами ( включая крокодилов и динозавров ) на ранних стадиях триаса . [ 59 ]
Смотрите также
[ редактировать ]Примечания
[ редактировать ]- ^ ce, cerebellum ; CNI, обонятельный нерв ; CNV +VCM - тройничный нерв и вена Capitis Medialis ; CNVI, Abducens нерв ; CNVII, лицевой нерв ; Cnix-Xi, глянцевая и вагоакссовые нервы ; CNXII, гипоглоссальный нерв ; EN, эпифизарный нерв ; FB, Forebrain ; FCL, Flocculus ; Ибик, внутренняя ветвь внутренней сонной артерии ; LOB, оставлена обонятельной лампочкой ; OB, обонятельная лампочка; ОТ, обонятельный тракт ; PG, гипофиз ; PGLL, боковая доли гипофиза; PF, Pontine Flexure ; Роб, правая обонятельная лампочка; VC, Видианский канал ; VC = Спа, видианский канал, где спенопалатиновая артерия проходит ; VCD, Vena Capitis Dorsalis
- ^ В более поздних классификациях семейство Gorgonopsidae, как правило, определяется, чтобы исключить базальных представителей горгонопсии. [ 7 ] : 242–244
- ^ Публикация в 2022 году официального описания рода Phorcys и аналогичного неопределенного образца показывает, что некоторые горгонопсийцы уже были больше по размеру во время среднего перми, без достижения аналогичных размеров до Inostrancevia или Rubidgeines . Тем не менее, крупные горгонопсийцы, такие как Phorcys и другие крупные терапсиды, могли оставить нишевое разделение для более мелких представителей после вымирания капитан . [ 33 ]
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый и фон Бендель, Ева-Мария; Kammerer, Christian F.; Кардджилов, Николай; Фернандес, Винсент; Fröbisch, Jörg (2018). «Черноальная анатомия горгонопсийского Cynariops robustus на основе CT-реконструкции» . Plos один . 13 (11): E0207367. BIBCODE : 2018PLOSO..1307367B . doi : 10.1371/journal.pone.0207367 . PMC 6261584 . PMID 30485338 .
- ^ Протеро, Дональд Р. (18 апреля 2022 г.). «20. Синапсиды: происхождение млекопитающих». Эволюция позвоночных: от происхождения до динозавров и за ее пределами . Boca Raton: CRC Press. doi : 10.1201/9781003128205-4 . ISBN 9780367473167 Полем S2CID 246318785 .
- ^ Jump up to: а беременный в Лаурин, Мишель (1998). «Новые данные об анатомии черепа ликанопов (Synapsida, Gorgonopsidae) и размышления о возможном присутствии стрептостильного у горгопсинцев». Журнал палеонтологии позвоночных . 18 (4): 765–776. Bibcode : 1998jvpal..18..765L . doi : 10.1080/02724634.1998.10011105 .
- ^ Jump up to: а беременный Суинтон, мы (1954). Ископаемые амфибии и рептилии . Британский музей (естественная история). п. 33.
- ^ Angielczyk, Kenneth D.; Kammerer, Christian F. (2018). «Не млекопитающие синапсиды: глубокие корни семейного древа млекопитающих». В Зачосе, Фрэнк Э.; Ашер, Роберт Дж. (Ред.). Эволюция млекопитающих, разнообразие и систематика . Берлин: де Грютер. ISBN 9783110275902 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k Kammerer, Christian F. (2016). Суды, Ганс-Дитер (ред.). «Систематика Rubidgeinae (Therapsida: Gorgonopsia)» . ПЕРЕЙ . 4 : E1608. doi : 10.7717/peerj.1608 . PMC 4730894 . PMID 26823998 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. Gebauer, Eva VI (2007). Филогения и эволюция горгонопсии со специальной ссылкой на череп и скелет GPIT/RE/7113 (PDF) (PhD). Эберхард-Карлс Университет Тюбингена. Архивировано из оригинала 2012-07-22.
- ^ Jump up to: а беременный Арауджо, Рикардо; Фернандес, Винсент; Полцин, Майкл Дж.; Fröbisch, Jörg; и Martins, Rui MS (2017). «Аспекты горгонопсийской палеобиологии и эволюции: понимание основного разрыва, затылка, костного лабиринта, сосудистой сети и нейроанатомии» . ПЕРЕЙ . 5 : E3119. doi : 10.7717/peerj.3119 . PMC 5390774 . PMID 28413721 .
- ^ Jump up to: а беременный Кермак, Дорис М. и Кермак, Кеннет А. (1984). «Минь, заменение зубами и артикуляция челюсти». Эволюция персонажей млекопитающих . С. 71–79. doi : 10.1007/978-1-4684-7817-4_5 . ISBN 978-1-4684-7817-4 .
- ^ Jump up to: а беременный Уитни, мистер; Лебланк, Арх; Рейнольдс, Ар; и Бринк, KS (2020). «Конвергентные зубные адаптации в заглушках гиперкарноядных синапсидов и динозавров» . Биологические письма . 16 (12). doi : 10.1098/rsbl.2020.0750 . PMC 7775981 . PMID 33321067 .
- ^ Jump up to: а беременный в Ван Валкенбург, Блэр и Дженкинс, Ян (2002). «Эволюционные закономерности в истории пермо-триассических и кайнозойских синапсид хищников». Документы палеонтологического общества . 8 : 273–275. doi : 10.1017/s10893326000011121 .
- ^ Jump up to: а беременный Кермак, Ка (1956). «Замена зубов у млекопитающих рептилий подчиненных горгонопсии и тероцефалии». Философские транзакции Королевского общества Лондона B: Биологические науки . 240 (670): 95–133. Bibcode : 1956rsptb.240 ... 95K . doi : 10.1098/rstb.1956.0013 .
- ^ Jump up to: а беременный Kümmell, SB; Фрей, Э. (2014). «Диапазон движения в лучах I от Manus и PES и предварительность автоподий в раннем пермском до позднего мела без аномодонта Synapsida» . Plos один . 9 (12): E113911. Bibcode : 2014ploso ... 9K3911K . doi : 10.1371/journal.pone.0113911 . PMC 4269487 . PMID 25517726 .
- ^ Jump up to: а беременный Оуэн, Ричард (1986). Описательный и иллюстрированный каталог ископаемых рептилии Южной Африки в коллекции Британского музея . Отпечатано по заказу попечителей. С. 27–29.
- ^ Jump up to: а беременный в дюймовый и Kammerer, Christian F. и Maseutin, Vladimir (2018). Молодой, Марк (ред.). «Gorgonopsian Therapsids ( Nochnitsa Gen. Nov. И Viatkogorgon ) из Permian Kotelnich Conitity в России» . ПЕРЕЙ . 6 : E4954. doi : 10.7717/peerj.4954 . PMC 5995105 . PMID 29900078 .
- ^ Янг, Чунг С. (1979). «Поздняя пермская фауна из Цзиюан, Хэнань». Повертеблат Паласиатика . 17 (2): 99–113.
- ^ Sigogneau-Russell, Дениз ; Sun, AI-Lin (1981). «Краткий обзор китайских синапсидов». Geobios . 14 (2): 276. Bibcode : 1981geobi..14..275s . doi : 10.1016/s0016-6995 (81) 80012-5 . S2CID 129556772 .
- ^ Лю, июнь; Ян, Ван (сентябрь 2022 г.). «Горгонопсиец из формации Вутонггу (Чангсин, Пермский) бассейна Турпана, Синьцзян, Китай» . Палеоворд . 31 (3): 383–388. doi : 10.1016/j.palwor.2022.04.004 .
- ^ Рэй, Сангхамитра; Bandyopadhyay, Saswati (март 2003 г.). «Поздние пермские позвоночные сообщества долины Пранхиты -Годавари, Индия» . Журнал азиатских наук о Земле . 21 (6): 643–654. Bibcode : 2003jaesc..21..643r . doi : 10.1016/s1367-9120 (02) 00050-0 . S2CID 140601673 . Получено 24 сентября 2022 года .
- ^ Jump up to: а беременный Смайли, Тара М.; Сидор, Кристиан А.; Мага, Абдулай; Ide, Oumarou (2008). «Фауна Верхнего Пермца Нигера . Журнал палеонтологии позвоночных . 28 (2): 543–547. Bibcode : 2008jvpal..28..543s . doi : 10.1671/0272-4634 (2008) 28 [543: tvfotu] 2.0.co; 2 . S2CID 131082914 .
- ^ Tectus в Чарльтоне Т. Льюис (1891) Элементарный латинский словарь , Harper & Brothers
- ^ Лидеккер, Ричард (1890). Каталог ископаемых рептилии и амфибий в Британском музее (естественная история), часть IV . Британский музей (естественная история). п. 111.
- ^ Сили, Гарри Г. (1895). «Исследования по структуре, организации и классификации ископаемых рептилии . Анналы и журнал естественной истории . 13 (6): 375. doi : 10.1080/002229394086777718 .
- ^ Осборн, Генри Ф. (1903). «Подклассы рептилий Diapsida и Synapsida и ранняя история Diaptosauria. Том I. Часть VIII» (PDF) . Мемуары Американского музея естественной истории . 1 : 451–466.
- ^ Метла, Роберт (1905). «При использовании термина аномодонтия» . Записи музея Олбани . 1 (4): 266–269.
- ^ Метла, Роберт (1913). «На Горгонопсии, подряд рептилий, похожих на млекопитающих» . Труды зоологического общества Лондона . 1913 (2): 225–230. doi : 10.1111/j.1469-7998.1913.tb07574.x .
- ^ Wyllie, Alistair (2003). «Обзор голотипов Robert Broom TherapsId: пережили ли они испытание временем?» (PDF) . Palaeontologia Africana . 39 (39): 2.
- ^ Kammerer, Christian F. (2014), «Theradentia: введение», в Kammerer, Christian F.; Ангелчик, Кеннет Д.; Jöbisch, история Synapsida , Ранняя эволюционная Fröbisch 165–169, doi : 10.1007/978-94-007-6841-3_10 , ISBN 978-94-007-6841-3
- ^ Jump up to: а беременный Рассел, Дениз (1989). Высоты, Питер (ред.). Theroidontia I: Phthinusch, Bearmosy, Eotitanusia, Corgonosia . Энциклопедия палеохерпетилогии. Тол. 17 б/я. Штутгарт: версия ISBN 978-3437304873 .
- ^ Рейс, Роберт Р.; Лаурин, Мишель (2004). «Переоценка загадочной пермской синапсидии Watongia и его стратиграфической значимости» . Канадский журнал наук о Земле . 41 (4): 377–386. Bibcode : 2004cajes..41..377r . doi : 10.1139/e04-016 . S2CID 129233127 .
- ^ Ивахненко, Михаил Ф. (2002). «Таксономия восточно -европейской горгонопии (Therapsida)». Палеонтологический журнал . 36 (3): 283–292. ISSN 0031-0301 .
- ^ Kammerer, Christian F.; Смит, Роджер М.Х.; День, Майкл О.; Рубидж, Брюс С. (2015). «Новая информация о морфологии и стратиграфической диапазоне средней пермианской горгонопсийской метлы эрифхостомы, 1911 год». Документы по палеонтологии . 1 (2): 201–221. Bibcode : 2015ppal .... 1..201k . doi : 10.1002/spp2.1012 . S2CID 128762256 .
- ^ Jump up to: а беременный в дюймовый Kammerer, Christian F.; Рубидж, Брюс С. (2022). «Самые ранние горгонопсийцы из бассейна Кару в Южной Африке». Журнал африканских наук о Земле . 194 : 104631. Bibcode : 2022jafes.19404631K . doi : 10.1016/j.jafrearsci.2022.104631 . S2CID 249977414 .
- ^ Jump up to: а беременный в Кемп, Том С. (2006). «Происхождение и раннее излучение рептилий, подобных млекопитающим,: палеобиологическая гипотеза» . Журнал эволюционной биологии . 19 (4): 1231–1247. doi : 10.1111/j.1420-9101.2005.01076.x . PMID 16780524 . S2CID 3184629 .
- ^ День, Майкл О.; Рамезани, Джахандар; Боуринг, Самуил А.; Садлер, Питер М.; Эрвин, Дуглас Х.; Абдала, Фернандо; и Рубидж, Брюс С. (2015). произошло земного массового вымирания в середине пермера? «Когда и как Труды Королевского общества б . 282 (1811): 20150834. DOI : 10.1098/rspb.2015.0834 . PMC 4528552 . PMID 26156768 .
- ^ Jump up to: а беременный в Антон, Маурисио (2013). Саблету Блумингтон, Индиана: Университет Индианы Пресс. Стр. 204–2 ISBN 978-0-253-01042-1 .
- ^ Рандау, Марсела; Карбон, Крис; и Turvey, Samuel T. (2013). Эванс, Алистер Роберт (ред.). "Эволюция собак в саблезубых плотоядных животных: естественный отбор или сексуальный отбор?" Полем Plos один . 8 (8): E72868. BIBCODE : 2013PLOSO ... 872868R . doi : 10.1371/journal.pone.0072868 . PMC 3738559 . PMID 23951334 .
- ^ Круикшанк, Ари (1973). «Режим жизни горгонопсинцев» (PDF) . Palaeontologica Africana . 15 (1–2): 65–67.
- ^ Эстефа, Джорди; Таффоро, Пол; Клемент, Алиса М.; Клембара, Йозеф; Niedźwiedzki, Grzegorz; Берруйер, Камилла; и Санчес, Софи (2021). Tautz, Diethard (ed.). «Новый свет проливается на раннюю эволюцию ростовой плиты конечностей и костного мозга» . элиф . 10 : 51581. DOI : 10.7554/elife.51581 . PMC 7924947 . PMID 33648627 .
- ^ Kriloff, A.; Жермен, Д.; Canoville, A.; Винсент, П.; Sache, M. & Laurin, M. (2008). «Эволюция костной микроанатомии тетраподной голени и ее использование при палеобиологическом выводе» (PDF) . Журнал эволюционной биологии . 21 (3): 807–826. doi : 10.1111/j.1420-9101.2008.01512.x . PMID 18312321 .
- ^ Маркетти, Лоренцо; Кляйн, Хендрик; Бухвиц, Майкл; Рончи, Асонио; Смит, Роджер М.Х.; Де Клерк, Уильям Дж.; Sciscio, Lara; Groenewald, Gideon H. (2019). «Пермские триасовые следы позвоночных из Южной Африки: Ichnotaxonomy, продюсеры и биостратиграфия через два основных кризиса фауны» . Gondwana Research . 72 : 159. Bibcode : 2019gondr..72..139M . doi : 10.1016/j.gr.2019.03.009 . S2CID 133781923 .
- ^ Бенуа, Жюльен; Абдала, Фернандо; Мангер, Пол Р.; и Рубидж, Брюс С. (2016). «Шестой смысл у предшественников млекопитающих: изменчивость теменного отверстия и эволюция пинецкового глаза у южноафриканской пермо-триасасической эвриодонтской терапиды» . Acta Palaeontologica Polonica . 61 (4): 777–789. doi : 10.4202/app.00219.2015 . S2CID 59143925 .
- ^ Angielczyk, KD и Schmitz, L. (2014). «Ночная продукция в синапсидах предшествует происхождению млекопитающих более чем на 100 миллионов лет» . Труды Королевского общества б . 281 (1793). doi : 10.1098/rspb.2014.1642 . PMC 4173690 . PMID 25186003 .
- ^ Jump up to: а беременный Хиллуениус, Виллем Дж. (1992). Носовые турбинаты и эволюция эндотермии млекопитающих (PhD). Орегонский государственный университет. С. 74–76. HDL : 1957/35986 .
- ^ Хиллуениус, Виллем Дж. (2000). «Септомаксилла не меммерских синапсидов: корреляты мягких тканей и новая функциональная интерпретация». Журнал морфологии . 245 (1): 29–50. doi : 10.1002/1097-4687 (200007) 245: 1 <29 :: Aid-Jmor3> 3.0.co; 2-B . PMID 10861830 . S2CID 22452028 .
- ^ Кемп, Т.С. (2006). «Происхождение эндотермии млекопитающих: парадигма эволюции сложной биологической структуры» (PDF) . Зоологический журнал Линневого общества . 147 (4): 473–488. doi : 10.1111/j.1096-3642.2006.00226.x .
- ^ Тернер, Дж. Скотт и Трейси, С. Ричард (1986). «Размер тела, гомеотермия и контроль тепла тела у млекопитающих рептилий». В Хоттоне, Николас III; Маклин, Пол Д.; Рот, январь Дж.; и Рот, Э. Кэрол (ред.). Экология и биология рептилий, подобных млекопитающим (PDF) . Смитсоновская институциональная пресса. Архивировано из оригинала (PDF) 2021-01-20 . Получено 2020-09-26 .
- ^ Ван Велан, Ли (сентябрь 1960 г.). «Терапсиды как млекопитающие». Эволюция 14 (3): 304–313. doi : 10.2307/2405973 . JSTOR 2405973 .
{{cite journal}}: Cs1 Maint: дата и год ( ссылка ) - ^ Като, Кайл М.; Рега, Элизабет А.; Сидор, Кристиан А.; и Huttenlocker, Adam K. (2020). «Исследование поражения костей у горгонопсийского (синапсида) из Пермия Замбии и периостаальных реакций у ископаемых не млекопитающих тетрапод» . Философские транзакции Королевского общества б . 375 (1793). doi : 10.1098/rstb.2019.0144 . PMC 7017433 . PMID 31928188 .
- ^ Уитни, Меган Р.; Мосей, Ларри; и Сидор, Кристиан А. (2017). «Одонтома у 255-миллионного добычи млекопитающих» . Джама Онкология . 3 (7): 998–1000. doi : 10.1001/Jamaoncol.2016.5417 . PMC 5824274 . PMID 27930769 .
- ^ Бенуа, Жюльен; Браунинг, Клэр; Нортон, Люк А. (2021). «Первая исцеленная отметка укуса и встроенный зуб в морде среднего пермского горгонопса (Synapsida: Therapsida)» . Границы в экологии и эволюции . 6 : 699298. DOI : 10.3389/fevo.2021.699298 .
- ^ Kammerer, Christian F. и Maseutin, Vladimir (2018). Суды, Ганс-Дитер (ред.). «Новый тероцефайец ( Gorynychus Masyutinae Gen. Et sp. Nov.) Из Permian Kotelnich Conitity, регион Киров, Россия» . ПЕРЕЙ . 6 : E4933. doi : 10.7717/peerj.4933 . PMC 5995100 . PMID 29900076 .
- ^ Уилсон, Эндрю; Флинт, Стивен; Пейенберг, Тобиас; Товер, Эрик; Ланчи, Лука (2014). «Архитектурные стили и седиментология речной группы Нижнего Бофорта, бассейн Кару, Южная Африка». Журнал осадочных исследований . 84 (4): 326–348. Bibcode : 2014JSEDR..84..326W . doi : 10.2110/jsr.2014.28 . S2CID 129643680 .
- ^ Jump up to: а беременный Смит, Роджер М.Х. (1987). «Морфология и история осаждения эксгумированных пермских батончиков на юго -западе Кару, Южная Африка». Журнал осадочных исследований . 57 (1): 19–20. doi : 10.1306/212F8A8F-2B24-11D7-8648000102C1865D . S2CID 130393112 .
- ^ Якмант, Елена; Inozemtsev, swatlav a.; Сергени В. (2004). "Opper Permian платформы . de Ceensias Revista Mexicana 21 (1): 110–1
- ^ Looy, Cindy v.; Ряды, Стефани Л.; Чейни, Дэн С.; Санчес, Софи; Стейер, Жан-Себастьен; Смит, Роджер М.Х.; Сидор, Кристиан А.; Майерс, Тимоти С.; Ide, Oumarou; Табор, Нил Дж. (Июнь 2016 г.). «Биологические и вещественные доказательства экстремальной сезонности в центральной пермской Пангее» . Палеогеография, палеоклиматология, палеоэкология . 451 : 210–226. Bibcode : 2016ppp ... 451..210L . doi : 10.1016/j.palaeo.2016.02.016 . S2CID 131336926 .
- ^ Бентон, MJ (2018). «Гипертермальное массовое вымирание: модели убийства во время исчезновения массы перм-триаса» . Философские транзакции Королевского общества а . 376 (2130). BIBCODE : 2018RSPTA.37670076B . doi : 10.1098/rsta.2017.0076 . PMC 6127390 . PMID 30177561 .
- ^ Бенуа, Жюльен; Kammerer, Christian F.; Кукла, Кэтлин; Groenewald, David P.; Смит, Роджер М.Х. (2024). «Выживали ли горгонопсии энмианскую« Великую умирающую »? Повторно одобряет три горгонопского образца (Therapsida, Theriodontia) из Триаса Листрозаврской зоны деклавра, бассейн Кару, Южная Африка». Палеогеография, палеоклиматология, палеоэкология . 638 : 112044. Bibcode : 2024ppp ... 63812044b . doi : 10.1016/j.palaeo.2024.112044 . S2CID 267080711 .
- ^ Сукиас, Роланд Б.; Батлер, Ричард Дж.; и Бенсон, Роджер Б.Дж. (2012). «Рост динозавров показывает, что основные переходы размера тела обусловлены пассивными процессами эволюции признаков» . Труды Королевского общества б . 279 (1736): 2180–2187. doi : 10.1098/rspb.2011.2441 . PMC 3321709 . PMID 22298850 . S2CID 1231897 .
Внешние ссылки
[ редактировать ] СМИ, связанные с Горгонопсией в Wikimedia Commons
СМИ, связанные с Горгонопсией в Wikimedia Commons  Данные, связанные с Gorgonopsia в Wikispecies
Данные, связанные с Gorgonopsia в Wikispecies







| Базы данных управления авторитетом : национальный |
|---|