посыпать
| Посыпать | |||
|---|---|---|---|
 Серпин (белый) с «реактивной центральной петлей» (синий), связанной с протеазой (серый). Как только протеаза попытается катализировать процесс, он будет необратимо ингибирован . ( PDB : 1K9O ) | |||
| Идентификаторы | |||
| Символ | Серпин, ЗЕРПИН ( коренной символ семьи) | ||
| Пфам | PF00079 | ||
| ИнтерПро | IPR000215 | ||
| PROSITE | PDOC00256 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 1хле / ОБЛАСТЬ ПРИМЕНЕНИЯ / СУПФАМ | ||
| CDD | cd00172 | ||
| |||
Серпины представляют собой суперсемейство белков активности со схожими структурами, которые были впервые идентифицированы для их ингибирования протеазы и обнаружены во всех королевствах жизни . [ 1 ] [ 2 ] Серпин аббревиатура был первоначально придуман, потому что первые серпины, которые должны быть идентифицированы, действуют на химотрипсин-подобные протеазы ( Ser Ine Ploease сериновые у Hibitors). [ 3 ] [ 4 ] [ 5 ] Они примечательны своим необычным механизмом действия, при котором они необратимо ингибируют целевую протеазу , претерпевая значительные конформационные изменения мишени , разрушающие активный сайт . [ 6 ] [ 7 ] Это контрастирует с более распространенным конкурентным механизмом ингибиторов протеаз, которые связываются с активным центром протеазы и блокируют доступ к нему. [ 8 ] [ 9 ]
Ингибирование протеаз серпинами контролирует ряд биологических процессов, включая коагуляцию и воспаление , и, следовательно, эти белки являются целью медицинских исследований . [ 10 ] Их уникальные конформационные изменения также делают их интересными для структурной биологии и сворачивания белков . исследователей [ 7 ] [ 8 ] Механизм конформационных изменений дает определенные преимущества, но у него есть и недостатки: серпины уязвимы к мутациям , которые могут привести к серпинопатиям, таким как неправильное сворачивание белка и образование неактивных длинноцепочечных полимеров . [ 11 ] [ 12 ] серпина Полимеризация не только уменьшает количество активного ингибитора, но и приводит к накоплению полимеров, вызывая гибель клеток и недостаточность органов . [ 10 ]
Хотя большинство серпинов контролируют протеолитические каскады, некоторые белки со структурой серпинов не являются ингибиторами ферментов , а вместо этого выполняют разнообразные функции, такие как хранение (как в яичном белке — овальбумин ), транспортировка, как в белках-переносчиках гормонов ( тироксин-связывающий глобулин , кортизол-связывающий глобулин ) и молекулярный шаперон ( HSP47 ). [ 9 ] Термин серпин также используется для описания этих членов, несмотря на их неингибирующую функцию, поскольку они эволюционно связаны. [ 1 ]
История
[ редактировать ]Ингибирующая активность протеаз в плазме крови впервые была зарегистрирована в конце 1800-х годов. [ 13 ] но только в 1950-х годах были выделены серпины антитромбин и альфа-1-антитрипсин . [ 14 ] с последующим признанием их близкой семейной гомологии в 1979 году. [ 15 ] [ 16 ] Их принадлежность к новому семейству белков стало очевидным при их дальнейшем совмещении с неингибирующим белком яичного белка овальбумином , в результате чего образовалось то, что первоначально называлось альфа1-антитрипсин-антитромбин III-овальбумин . суперсемейством ингибиторов сериновых протеиназ [ 17 ] но впоследствии был лаконично переименован в Серпины. [ 18 ] Первоначальная характеристика нового семейства была сосредоточена на альфа1-антитрипсине , серпине, присутствующем в высокой концентрации в плазме крови, общее генетическое заболевание которого, как было показано, вызывает предрасположенность к эмфиземе легких. [ 19 ] и к циррозу печени . [ 20 ] Идентификация мутаций S и Z [ 21 ] [ 22 ] ответственный за генетический дефицит, и последующее выравнивание последовательностей альфа1-антитрипсина и антитромбина в 1982 году привело к признанию близкой гомологии активных центров двух белков. [ 23 ] [ 24 ] сосредоточен на метионине [ 25 ] в альфа1-антитрипсине как ингибиторе тканевой эластазы и в аргинине в антитромбине. [ 26 ] как ингибитор тромбина. [ 27 ]
Критическая роль остатка активного центра в определении специфичности ингибирования серпинов была однозначно подтверждена открытием, что естественная мутация метионина активного центра в альфа1-антитрипсине на аргинин, как и в антитромбине, приводит к тяжелому нарушению свертываемости крови. [ 28 ] Эта специфичность ингибирования активного центра также была очевидна во многих других семействах ингибиторов протеаз. [ 7 ] но серпины отличались от них тем, что представляли собой гораздо более крупные белки, а также обладали, как вскоре выяснилось, врожденной способностью изменять форму. Природа этого конформационного изменения была раскрыта после определения в 1984 году первой кристаллической структуры серпина - альфа1-антитрипсина после расщепления. [ 29 ] Это вместе с последующим решением структуры нативного (нерасщепленного) овальбумина. [ 30 ] показали, что механизм ингибирования серпинов включает значительный конформационный сдвиг с перемещением открытой пептидной петли, содержащей реактивный сайт, и ее включением в качестве средней цепи в основной бета-складчатый лист, который характеризует молекулу серпина. [ 31 ] [ 32 ] Ранние доказательства существенной роли этого движения петли в тормозном механизме были получены благодаря открытию того, что даже незначительные отклонения в аминокислотных остатках, которые образуют основу движения антитромбина, приводят к тромботическим заболеваниям. [ 31 ] [ 33 ] Окончательное подтверждение связанного смещения целевой протеазы при этом движении петли было предоставлено в 2000 г. структурой постингибиторного комплекса альфа1-антитрипсина с трипсином, [ 6 ] показывая, как смещение приводит к деформации и инактивации прикрепленной протеазы. Последующие структурные исследования выявили дополнительное преимущество конформационного механизма. [ 34 ] позволяя тонко модулировать ингибирующую активность, что особенно заметно на тканевом уровне. [ 35 ] с функционально разнообразными серпинами в плазме человека.
В настоящее время идентифицировано более 1000 серпинов, в том числе 36 белков человека, а также молекулы всех царств жизни — животных , растений , грибов , бактерий и архей — и некоторых вирусов . [ 36 ] [ 37 ] [ 38 ] Центральной особенностью всех них является строго консервативная структура, которая позволяет точно выравнивать их ключевые структурные и функциональные компоненты на основе шаблонной структуры альфа1-антитрипсина. [ 39 ] В 2000-х годах была введена систематическая номенклатура для классификации членов надсемейства серпинов на основе их эволюционных взаимоотношений. [ 1 ] Таким образом, серпины представляют собой самое большое и разнообразное суперсемейство ингибиторов протеаз. [ 40 ]
Активность
[ редактировать ]
Большинство серпинов являются ингибиторами протеаз внеклеточные химотрипсиноподобные , воздействуя на сериновые протеазы . Эти протеазы содержат нуклеофильный остаток серина в каталитической триаде в своем активном центре . Примеры включают тромбин , трипсин и эластазу нейтрофилов человека . [ 41 ] Серпины действуют как необратимые , ингибиторы самоубийства захватывая промежуточный продукт каталитического механизма протеазы. [ 6 ]
Некоторые серпины ингибируют другие классы протеаз, обычно цистеиновые протеазы , и называются «ингибиторами межклассовых протеаз». Эти ферменты отличаются от сериновых протеаз тем, что в их активном центре используется нуклеофильный остаток цистеина , а не серин . [ 42 ] Тем не менее, ферментативная химия аналогична, а механизм ингибирования серпинами одинаков для обоих классов протеаз. [ 43 ] Примеры ингибирующих серпинов межклассового типа включают серпин B4, антиген 1 плоскоклеточной карциномы (SCCA-1), а также птичий серпин, специфичный для стадии терминации ядра миелоидного и эритроидного ядра (MENT), которые оба ингибируют папаин-подобные цистеиновые протеазы . [ 44 ] [ 45 ] [ 46 ]
Биологическая функция и локализация
[ редактировать ]Ингибирование протеазы
[ редактировать ]Примерно две трети серпинов человека выполняют внеклеточную роль, ингибируя протеазы в кровотоке, чтобы модулировать их активность. Например, внеклеточные серпины регулируют протеолитические каскады, играющие центральную роль в свертывании крови (антитромбин), воспалительные и иммунные реакции (антитрипсин, антихимотрипсин и ингибитор C1 ) и ремоделирование тканей (PAI-1) . [ 9 ] Ингибируя протеазы сигнального каскада , они также могут влиять на развитие . [ 47 ] [ 48 ] В таблице серпинов человека (ниже) приведены примеры ряда функций, выполняемых серпинами человека, а также некоторых заболеваний, возникающих в результате дефицита серпинов.
Протеазные мишени внутриклеточных ингибирующих серпинов трудно идентифицировать, поскольку многие из этих молекул, по-видимому, выполняют перекрывающиеся роли. Кроме того, многим человеческим серпинам не хватает точных функциональных эквивалентов в модельных организмах, таких как мышь. Тем не менее важной функцией внутриклеточных серпинов может быть защита от нежелательной активности протеаз внутри клетки. [ 49 ] Например, одним из наиболее изученных внутриклеточных серпинов человека является серпин B9 , который ингибирует цитотоксическую гранулярную протеазу гранзим B. При этом серпин B9 может защищать от непреднамеренного высвобождения гранзима B и преждевременной или нежелательной активации путей гибели клеток . [ 50 ]
Некоторые вирусы используют серпины для нарушения функций протеаз в организме хозяина. Серпин коровьей оспы CrmA вируса (модификатор цитокинового ответа А) используется во избежание воспалительных и апоптотических реакций инфицированных клеток-хозяев. CrmA повышает инфекционность за счет подавления воспалительной реакции хозяина за счет ингибирования процессинга IL-1 и IL-18 цистеиновой протеазой каспазой -1. [ 51 ] У эукариот растительный серпин ингибирует обе метакаспазы. [ 52 ] и папаинподобную цистеиновую протеазу. [ 53 ]
Неингибирующие роли
[ редактировать ]Неингибирующие внеклеточные серпины также выполняют широкий спектр важных ролей. Тироксинсвязывающий глобулин и транскортин транспортируют гормоны тироксин и кортизол соответственно. [ 54 ] [ 55 ] Неингибирующий серпин овальбумин является наиболее распространенным белком в яичном белке . Его точная функция неизвестна, но считается, что он является запасным белком для развивающегося плода . [ 56 ] Серпин 47 теплового шока является шапероном , необходимым для сворачивания коллагена правильного . коллагена Он действует путем стабилизации тройной спирали во время его обработки в эндоплазматическом ретикулуме . [ 57 ]
Некоторые серпины одновременно являются ингибиторами протеаз и выполняют дополнительные функции. Например, ингибитор ядерной цистеиновой протеазы MENT у птиц также действует как молекула ремоделирования хроматина птиц в эритроцитах . [ 45 ] [ 58 ]
Структура
[ редактировать ]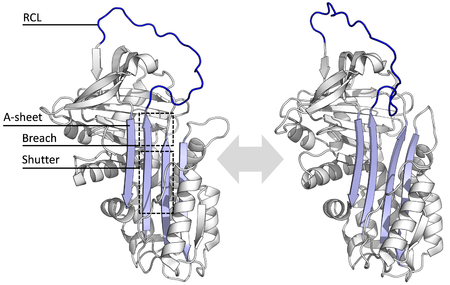
Все серпины имеют общую структуру (или складку), несмотря на различные функции. Все они обычно имеют три β-листа (названные A, B и C) и восемь или девять α-спиралей (называемых hA – hI). [ 29 ] [ 30 ] Наиболее важными областями для функции серпинов являются A-лист и петля реактивного центра (RCL). А-лист включает в себя две β-нити , которые расположены параллельно, с областью между ними, называемой «затвором», и верхней областью, называемой «разрывом». RCL образует начальное взаимодействие с целевой протеазой в ингибирующих молекулах. Были решены структуры, показывающие, что RCL либо полностью открыт, либо частично вставлен в A-лист, и считается, что серпины находятся в динамическом равновесии между этими двумя состояниями. [ 8 ] RCL также лишь временно взаимодействует с остальной структурой и поэтому очень гибок и подвержен воздействию растворителя. [ 8 ]
Установленные структуры серпинов охватывают несколько различных конформаций, что необходимо для понимания их многоступенчатого механизма действия. Таким образом, структурная биология сыграла центральную роль в понимании функции и биологии серпинов. [ 8 ]
Конформационные изменения и механизм торможения
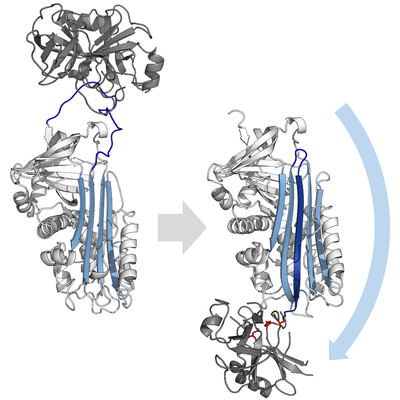
[ редактировать ]Ингибирующие серпины не ингибируют свои целевые протеазы с помощью типичного конкурентного механизма ( замок и ключ ), используемого большинством небольших ингибиторов протеаз (например, ингибиторов типа Кунитца ). Вместо этого серпины используют необычное конформационное изменение , которое нарушает структуру протеазы и не позволяет ей завершить катализ. Конформационное изменение включает в себя перемещение RCL к противоположному концу белка и вставку в β-лист А, образуя дополнительную антипараллельную β-цепь. Это переводит серпин из напряженного состояния в расслабленное состояние с более низкой энергией (переход S в R). [ 7 ] [ 8 ] [ 61 ]
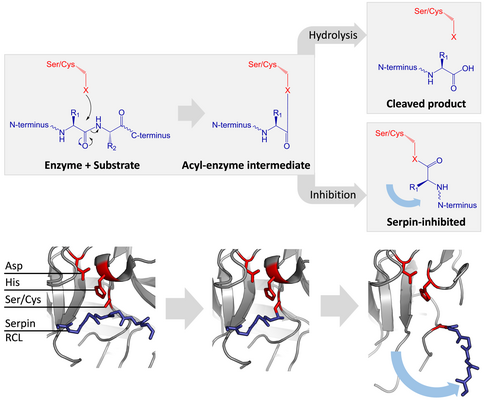
Сериновые и цистеиновые протеазы катализируют расщепление пептидной связи в два этапа. Первоначально каталитический остаток триады активного центра осуществляет нуклеофильную атаку на пептидную связь субстрата. При этом высвобождается новый N-конец и образуется ковалентная сложноэфирная связь между ферментом и субстратом. [ 7 ] Этот ковалентный комплекс между ферментом и субстратом называется промежуточным соединением ацил-фермента . В случае стандартных субстратов сложноэфирная связь гидролизуется и новый С-конец высвобождается для завершения катализа. Однако когда серпин расщепляется протеазой, он быстро претерпевает переход от S к R до того, как промежуточное соединение ацил-фермента гидролизуется. [ 7 ] Эффективность ингибирования зависит от того, что относительная кинетическая скорость конформационного изменения на несколько порядков превышает скорость гидролиза протеазой.
Поскольку RCL все еще ковалентно связан с протеазой через сложноэфирную связь, переход S к R тянет протеазу сверху вниз по серпину и искажает каталитическую триаду. Искаженная протеаза может гидролизовать ацильный промежуточный фермент очень медленно, поэтому протеаза остается ковалентно связанной в течение нескольких дней или недель. [ 6 ] Серпины классифицируются как необратимые ингибиторы и ингибиторы самоубийства , поскольку каждый белок серпина постоянно инактивирует одну протеазу и может функционировать только один раз. [ 7 ]
Аллостерическая активация
[ редактировать ]
Конформационная подвижность серпинов обеспечивает ключевое преимущество перед статическими ингибиторами протеаз «замок и ключ». [ 34 ] В частности, функция ингибирующих серпинов может регулироваться за счет аллостерических взаимодействий со специфическими кофакторами . Рентгеновские кристаллические структуры антитромбина , , кофактора гепарина II MENT и мышиного антихимотрипсина показывают, что эти серпины принимают конформацию, в которой первые две аминокислоты RCL вставлены в верхнюю часть A β-листа . Частично вставленная конформация важна, поскольку кофакторы способны конформационно переключать определенные частично вставленные серпины в полностью исключенную форму. [ 62 ] [ 63 ] Эта конформационная перестройка делает серпин более эффективным ингибитором.
Типичным примером этой ситуации является антитромбин, который циркулирует в плазме в частично введенном относительно неактивном состоянии. Остаток, определяющий первичную специфичность (аргинин P1), направлен в сторону тела серпина и недоступен для протеазы. При связывании высокоаффинной пентасахаридной последовательности с длинноцепочечным гепарином антитромбин претерпевает конформационные изменения, вытеснение RCL и воздействие аргинина P1. Таким образом, форма антитромбина, связанная с пентасахаридом гепарина, является более эффективным ингибитором тромбина и фактора Ха . [ 64 ] [ 65 ] Кроме того, обе эти коагуляционные протеазы также содержат сайты связывания (называемые экзосайтами ) для гепарина. Таким образом, гепарин также действует как шаблон для связывания как протеазы, так и серпина, что еще больше ускоряет взаимодействие между двумя сторонами. После первоначального взаимодействия образуется конечный серпиновый комплекс и высвобождается гепариновая часть. Это взаимодействие физиологически важно. Например, после повреждения стенки кровеносного сосуда гепарин подвергается воздействию, и антитромбин активируется для контроля реакции свертывания крови. Понимание молекулярной основы этого взаимодействия позволило разработать Фондапаринукс , синтетическую форму пентасахарида гепарина, используемую в качестве препарата, препятствующего свертыванию крови . [ 66 ] [ 67 ]
Скрытая конформация
[ редактировать ]
Некоторые серпины спонтанно претерпевают переход от S к R без расщепления протеазой, образуя конформацию, называемую латентным состоянием. Латентные серпины не способны взаимодействовать с протеазами и поэтому больше не являются ингибиторами протеаз. Конформационное изменение латентного периода не совсем то же самое, что переход S в R расщепленного серпина. Поскольку RCL все еще не поврежден, первая нить C-листа должна отслоиться, чтобы обеспечить полное вставление RCL. [ 68 ]
Регуляция латентного перехода может действовать как механизм контроля у некоторых серпинов, таких как PAI-1 . Хотя PAI-1 вырабатывается в ингибирующей S-конформации, он «автоинактивируется», переходя в латентное состояние, если только он не связан с кофактором витронектином . [ 68 ] Аналогичным образом, антитромбин также может спонтанно переходить в латентное состояние в качестве дополнительного механизма модуляции его аллостерической активации гепарином. [ 69 ] Наконец, N-конец тенгпина, серпина из Thermoanaerobacter tengcongensis , необходим для фиксации молекулы в нативном ингибирующем состоянии. Нарушение взаимодействий N-концевой области приводит к спонтанному конформационному изменению этого серпина в латентную конформацию. [ 70 ] [ 71 ]
Конформационное изменение неингибирующих функций
[ редактировать ]Некоторые неингибирующие серпины также используют конформационные изменения серпинов как часть своей функции. Например, нативная (S) форма тироксинсвязывающего глобулина имеет высокое сродство к тироксину, тогда как расщепленная форма (R) имеет низкое сродство. Аналогично, транскортин имеет более высокое сродство к кортизолу в его нативном (S) состоянии, чем в расщепленном (R) состоянии. Т.о., в этих серпинах расщепление RCL и переход S к R были использованы для высвобождения лиганда, а не для ингибирования протеазы. [ 54 ] [ 55 ] [ 72 ]
В некоторых серпинах переход S на R может активировать сигнальные события клеток. В этих случаях серпин, образовавший комплекс с целевой протеазой, затем распознается рецептором. Событие связывания затем приводит к передаче сигналов рецептором ниже по течению. [ 73 ] Таким образом, переход S к R используется для предупреждения клеток о присутствии протеазной активности. [ 73 ] Это отличается от обычного механизма, при котором серпины влияют на передачу сигналов, просто ингибируя протеазы, участвующие в сигнальном каскаде. [ 47 ] [ 48 ]
Деградация
[ редактировать ]Когда серпин ингибирует целевую протеазу, он образует постоянный комплекс, от которого необходимо избавиться. Что касается внеклеточных серпинов, конечные комплексы серпин-фермент быстро выводятся из кровообращения. Одним из механизмов, посредством которого это происходит у млекопитающих, является белок, связанный с рецептором липопротеинов низкой плотности ( LRP ), который связывается с ингибирующими комплексами, образованными антитромбином, PA1-1 и нейросерпином, вызывая клеточное поглощение . [ 73 ] [ 74 ] Аналогично, некротический серпин дрозофилы разлагается в лизосомах после попадания в клетку липофоринового рецептора-1 (гомологичного семейству рецепторов ЛПНП млекопитающих ). [ 75 ]
Заболевания и серпинопатии
[ редактировать ]Серпины участвуют в широком спектре физиологических функций, поэтому мутации в кодирующих их генах могут вызывать ряд заболеваний. Мутации, которые изменяют активность, специфичность или агрегационные свойства серпинов, влияют на их функционирование. Большинство заболеваний, связанных с серпинами, являются результатом полимеризации серпинов в агрегаты, хотя также встречаются несколько других типов мутаций, связанных с болезнями. [ 8 ] [ 76 ] Нарушение дефицита альфа-1-антитрипсина является одним из наиболее распространенных наследственных заболеваний . [ 11 ] [ 77 ]
Бездействие или отсутствие
[ редактировать ]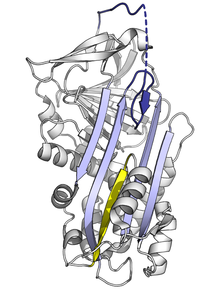
Поскольку напряженная серпиновая складка является высокоэнергетической, мутации могут привести к тому, что она неправильно перейдет в конформацию с более низкой энергией (например, расслабленную или латентную) до того, как она правильно выполнит свою ингибирующую роль. [ 10 ]
Мутации, которые влияют на скорость или степень вставки RCL в A-лист, могут привести к тому, что серпин подвергнется конформационному изменению S на R до того, как вступит в контакт с протеазой. Поскольку серпин может произвести это конформационное изменение только один раз, полученный серпин с ошибкой становится неактивным и не может должным образом контролировать свою целевую протеазу. [ 10 ] [ 78 ] Аналогичным образом, мутации, которые способствуют неадекватному переходу в латентное мономерное состояние, вызывают заболевание за счет уменьшения количества активного ингибирующего серпина. Например, связанные с заболеванием варианты антитромбина «виби» и «вобл» . [ 79 ] оба способствуют формированию латентного состояния .
В структуре связанного с болезнью мутанта антихимотрипсина (L55P) обнаружена еще одна, неактивная «δ-конформация». В δ-конформации четыре остатка RCL встраиваются в верхнюю часть β-листа А. Нижняя половина листа заполняется в результате частичного переключения одной из α-спиралей (F-спирали) на Конформация β-цепи, завершающая водородную связь β-листа. [ 80 ] Неясно, могут ли другие серпины принимать этот конформер и имеет ли эта конформация функциональную роль, но предполагается, что δ-конформация может быть принята тироксинсвязывающим глобулином во время высвобождения тироксина. [ 55 ] Неингибирующие белки, связанные с серпинами, также могут вызывать заболевания при мутации. Например, мутации в SERPINF1 вызывают у людей несовершенный остеогенез VI типа. [ 81 ]
В отсутствие необходимого серпина протеаза, которую он обычно регулирует, становится сверхактивной, что приводит к патологиям. [ 10 ] Следовательно, простой дефицит серпина (например, нулевая мутация ) может привести к заболеванию. [ 82 ] Нокауты генов , особенно у мышей , используются экспериментально для определения нормальных функций серпинов по эффекту их отсутствия. [ 83 ]
Изменение специфичности
[ редактировать ]В некоторых редких случаях изменение одной аминокислоты в RCL серпина меняет его специфичность, направленную на неправильную протеазу. Например, мутация антитрипсина-Питтсбурга (M358R) заставляет серпин α1-антитрипсина ингибировать тромбин, вызывая нарушение свертываемости крови . [ 28 ]
Полимеризация и агрегация
[ редактировать ]

Большинство заболеваний серпинов возникают из-за агрегации белков и называются «серпинопатиями». [ 12 ] [ 80 ] Серпины уязвимы к мутациям, вызывающим заболевания, которые способствуют образованию неправильно свернутых полимеров из-за их нестабильной структуры. [ 80 ] Хорошо изученные серпинопатии включают дефицит α1-антитрипсина (альфа-1), который может вызывать семейную эмфизему , а иногда и цирроз печени , некоторые семейные формы тромбозов , связанные с дефицитом антитромбина 1 и 2 типов , наследственный ангионевротический отек (НАО), связанный с дефицитом C1. -ингибиторная и семейная энцефалопатия с тельцами включения нейросерпина (ФЕНИБ; редкий тип деменция, вызванная полимеризацией нейросерпина). [ 11 ] [ 12 ] [ 84 ]
Каждый мономер агрегата серпина существует в неактивной, релаксированной конформации (с включением RCL в A-лист). Поэтому полимеры гиперстабильны к температуре и неспособны ингибировать протеазы. Таким образом, серпинопатии вызывают патологии, как и другие протеопатии (например, прионные заболевания), посредством двух основных механизмов. [ 11 ] [ 12 ] Во-первых, отсутствие активного серпина приводит к неконтролируемой активности протеаз и разрушению тканей. Во-вторых, гиперстабильные полимеры сами по себе закупоривают эндоплазматический ретикулум клеток, синтезирующих серпины, что в конечном итоге приводит к гибели клеток и повреждению тканей. В случае дефицита антитрипсина полимеры антитрипсина вызывают гибель клеток печени , что иногда приводит к повреждению печени и циррозу печени . Внутри клетки полимеры серпинов медленно удаляются путем деградации в эндоплазматической сети. [ 85 ] Однако детали того, как полимеры серпинов вызывают гибель клеток, еще предстоит полностью понять. [ 11 ]
Считается, что физиологические полимеры серпина образуются в результате событий замены доменов , когда сегмент одного белка серпина вставляется в другой. [ 86 ] Смена доменов происходит, когда мутации или факторы окружающей среды мешают заключительным стадиям сворачивания серпинов в нативное состояние, вызывая неправильное сворачивание высокоэнергетических промежуточных продуктов. [ 87 ] как димерные , так и тримерные Были решены структуры замены доменов. В димере (антитромбина) RCL и часть А-листа включаются в А-лист другой молекулы серпина. [ 86 ] Тример с замененным доменом (антитрипсина) образуется путем обмена совершенно другой области структуры, B-листа (при этом RCL каждой молекулы вставлен в свой собственный A-лист). [ 88 ] Также было высказано предположение, что серпины могут образовывать замены доменов путем вставки RCL одного белка в A-лист другого (полимеризация A-листа). [ 84 ] [ 89 ] Считается, что эти димерные и тримерные структуры с заменой доменов являются строительными блоками вызывающих заболевания полимерных агрегатов, но точный механизм до сих пор неясен. [ 86 ] [ 87 ] [ 88 ] [ 90 ]
Терапевтические стратегии
[ редактировать ]Несколько терапевтических подходов используются или исследуются для лечения наиболее распространенной серпинопатии: дефицита антитрипсина. [ 11 ] Аугментационная терапия антитрипсином одобрена при тяжелой эмфиземе, связанной с дефицитом антитрипсина. [ 91 ] При этой терапии антитрипсин очищается из плазмы доноров крови и вводится внутривенно (впервые продается как Проластин ). [ 11 ] [ 92 ] Для лечения тяжелых заболеваний, связанных с дефицитом антитрипсина, трансплантация легких и печени. доказала свою эффективность [ 11 ] [ 93 ] На животных моделях нацеливание на гены в индуцированных плюрипотентных стволовых клетках успешно использовалось для коррекции дефекта полимеризации антитрипсина и восстановления способности печени млекопитающих секретировать активный антитрипсин. [ 94 ] Также были разработаны небольшие молекулы, которые блокируют полимеризацию антитрипсина in vitro . [ 95 ] [ 96 ]
Эволюция
[ редактировать ]Серпины представляют собой наиболее широко распространенное и крупнейшее суперсемейство ингибиторов протеаз. [ 1 ] [ 40 ] Первоначально считалось, что они присущи только эукариотным организмам, но с тех пор были обнаружены у бактерий , архей и некоторых вирусов . [ 36 ] [ 37 ] [ 97 ] Остается неясным, являются ли гены прокариот потомками предкового прокариотического серпина или продуктом горизонтального переноса генов от эукариот. Большинство внутриклеточных серпинов принадлежат к одной филогенетической кладе, независимо от того, происходят ли они от растений или животных, что указывает на то, что внутриклеточные и внеклеточные серпины могли разойтись раньше растений и животных. [ 98 ] Исключения включают внутриклеточный серпин теплового шока HSP47, который является шапероном, необходимым для правильного сворачивания коллагена , и циклы между цис-Гольджи и эндоплазматическим ретикулумом . [ 57 ]
Считается, что ингибирование протеаз является наследственной функцией, а неингибирующие члены являются результатом эволюционной неофункционализации структуры. Конформационные изменения S на R также были адаптированы некоторыми связывающими серпинами для регулирования сродства к своим мишеням. [ 55 ]
Распределение
[ редактировать ]Животное
[ редактировать ]Человек
[ редактировать ]Геном человека кодирует 16 клад серпинов, называемых серпинами от серпинов до серпинов, включая 29 ингибирующих и 7 неингибирующих серпиновых белков. [ 9 ] [ 83 ] Система наименования серпинов человека основана на филогенетическом анализе примерно 500 серпинов, проведенных в 2001 году, с белками, названными serpinXY, где X - клада белка, а Y - номер белка в этой кладе. [ 1 ] [ 36 ] [ 83 ] Функции серпинов человека были определены с помощью сочетания биохимических исследований, генетических нарушений человека и моделей нокаутных мышей . [ 83 ]
Специализированные серпины млекопитающих
[ редактировать ]множество серпинов млекопитающих Было идентифицировано , которые не имеют очевидной ортологии с аналогом серпина человека. Примеры включают многочисленные серпины грызунов (особенно некоторые внутриклеточные серпины мышей ), а также маточные серпины . Термин маточный серпин относится к членам клады серпина А, которые кодируются геном SERPINA14. Маточные серпины производятся эндометрием ограниченной группы млекопитающих клады Laurasiatheria под влиянием прогестерона или эстрогена . [ 184 ] Они, вероятно, не являются функциональными ингибиторами протеиназ и могут функционировать во время беременности, подавляя материнский иммунный ответ против зачатия или участвуя в трансплацентарном транспорте. [ 185 ]
Насекомое
[ редактировать ]Геном Drosophila melanogaster содержит 29 генов, кодирующих серпины. Анализ аминокислотной последовательности позволил поместить 14 из этих серпинов в кладу серпинов Q и три в кладу серпинов K, а остальные двенадцать были классифицированы как осиротевшие серпины, не принадлежащие ни к одной кладе. [ 186 ] Систему классификации клад сложно использовать для серпинов дрозофилы , и вместо этого была принята номенклатурная система, основанная на положении генов серпинов в дрозофилы хромосомах . Тринадцать серпинов дрозофилы встречаются в геноме в виде изолированных генов (включая серпин-27A, см. ниже), а остальные 16 организованы в пять кластеров генов, которые встречаются в хромосомных положениях 28D (2 серпина), 42D (5 серпинов), 43A ( 4 змеевика), 77B (3 змеевика) и 88E (2 змеевика). [ 186 ] [ 187 ] [ 188 ]
Исследования серпинов дрозофилы показывают, что серпин-27A ингибирует протеазу Пасхи (последнюю протеазу в протеолитическом каскаде Нуделя, дефектной гаструляции, змеи и Пасхи) и, таким образом, контролирует формирование дорсовентрального паттерна . Функция East заключается в расщеплении Spätzle (лиганда хемокинового типа), что приводит к передаче сигналов, опосредованной toll . Помимо своей центральной роли в формировании эмбрионального паттерна, передача сигналов toll также важна для врожденного иммунного ответа у насекомых. Соответственно, серпин-27А также контролирует иммунный ответ насекомых. [ 48 ] [ 189 ] [ 190 ] У Tenebrio molitor (большого жука) белок (SPN93), содержащий два дискретных тандемных домена серпина, регулирует протеолитический каскад toll. [ 191 ]
Нематода
[ редактировать ]Геном нематодного червя C. elegans содержит 9 серпинов, все из которых лишены сигнальных последовательностей и поэтому, вероятно, являются внутриклеточными. [ 192 ] Однако только 5 из этих серпинов действуют как ингибиторы протеазы. [ 192 ] Один из них, SRP-6, выполняет защитную функцию и защищает от стресс-индуцированного кальпаин -ассоциированного разрушения лизосом. Кроме того, SRP-6 ингибирует лизосомальные цистеиновые протеазы, высвобождаемые после разрыва лизосом. Соответственно, черви, лишенные SRP-6, чувствительны к стрессу. В частности, черви, нокаутные по SRP-6, погибают при помещении в воду (летальный фенотип гипоосмотического стресса или Osl). Поэтому было высказано предположение, что лизосомы играют общую и контролируемую роль в определении судьбы клеток. [ 193 ]
Растение
[ редактировать ]Растительные змеи были одними из первых идентифицированных представителей надсемейства. [ 194 ] Белок серпина ячменя Z широко распространен в ячменном зерне и является одним из основных белковых компонентов пива. Геном модельного растения Arabidopsis thaliana содержит 18 серпиноподобных генов, хотя только 8 из них представляют собой полноразмерные последовательности серпинов.
Растительные серпины являются мощными ингибиторами химотрипсиноподобных сериновых протеаз млекопитающих in vitro , наиболее изученным примером является серпин Zx ячменя (BSZx), который способен ингибировать трипсин и химотрипсин, а также несколько факторов свертывания крови. [ 195 ] Однако близкие родственники химотрипсиноподобных сериновых протеаз у растений отсутствуют. RCL нескольких серпинов из зерна пшеницы и ржи содержат последовательности повторов поли-Q, аналогичные тем, которые присутствуют в белках хранения проламина эндосперма. [ 196 ] [ 197 ] Поэтому было высказано предположение, что растительные серпины могут ингибировать протеазы насекомых или микробов, которые в противном случае переваривали бы запасные белки зерна. В подтверждение этой гипотезы во флоэмном соке тыквы были идентифицированы специфические растительные серпины (CmPS-1). [ 198 ] и огуречные растения. [ 199 ] [ 200 ] Хотя наблюдалась обратная корреляция между усилением экспрессии CmPS-1 и выживаемостью тлей, эксперименты по кормлению in vitro показали, что рекомбинантный CmPS-1, по-видимому, не влияет на выживаемость насекомых. [ 198 ]
Были предложены альтернативные роли и мишени протеаз для растительных серпинов. Серпин Arabidopsis , AtSerpin1 (At1g47710; 3LE2 ), обеспечивает заданный контроль над запрограммированной гибелью клеток путем воздействия на папаин-подобную цистеиновую протеазу, реагирующую на высыхание-21 (RD21). [ 53 ] [ 201 ] AtSerpin1 также ингибирует метакаспазоподобные протеазы in vitro . [ 52 ] Два других серпина Arabidopsis , AtSRP2 (At2g14540) и AtSRP3 (At1g64030), по-видимому, участвуют в реакциях на повреждение ДНК. [ 202 ]
Грибковый
[ редактировать ]единственный грибной На сегодняшний день охарактеризован серпин: сельпин из Piromyces spp. штамм Е2. Piromyces — это род анаэробных грибов, обитающих в кишечнике жвачных животных и играющих важную роль в переваривании растительного материала. Предполагается, что Celpin обладает ингибирующим действием и содержит два N-концевых домена докерина в дополнение к домену серпина. Докерины обычно обнаруживаются в белках, которые локализуются в грибковой целлюлосоме — большом внеклеточном мультибелковом комплексе, расщепляющем целлюлозу. [ 38 ] Поэтому предполагается, что сельпин может защищать целлюлосому от растительных протеаз. Некоторые бактериальные серпины аналогичным образом локализуются в целлюлосоме. [ 203 ]
Прокариотический
[ редактировать ]Предсказанные гены серпинов спорадически распространены у прокариот . Исследования некоторых из этих молекул in vitro показали, что они способны ингибировать протеазы, и предполагается, что они действуют как ингибиторы in vivo . обнаружено несколько прокариотических серпинов У экстремофилов . Соответственно, в отличие от серпинов млекопитающих, эти молекулы обладают повышенной устойчивостью к тепловой денатурации. [ 204 ] [ 205 ] Точная роль большинства бактериальных серпинов остается неясной, хотя серпин Clostridium thermocellum локализуется в целлюлосоме . Предполагается, что роль серпинов, связанных с целлюлосомой, может заключаться в предотвращении нежелательной активности протеазы против целлюлосомы. [ 203 ]
Популярный
[ редактировать ]Серпины также экспрессируются вирусами как способ обойти иммунную защиту хозяина. [ 206 ] В частности, серпины, экспрессируемые вирусами оспы , в том числе коровьей оспой (вакцинией) и кроличьей оспой (миксомой), представляют интерес из-за их потенциального использования в качестве новых терапевтических средств при иммунных и воспалительных заболеваниях, а также в трансплантационной терапии. [ 207 ] [ 208 ] Serp1 подавляет TLR-опосредованный врожденный иммунный ответ и обеспечивает неопределенное выживание сердечного аллотрансплантата у крыс. [ 207 ] [ 209 ] Crma и Serp2 являются межклассовыми ингибиторами и нацелены как на сериновые (гранзим B; хотя и слабо), так и на цистеиновые протеазы (каспаза 1 и каспаза 8). [ 210 ] [ 211 ] По сравнению со своими аналогами млекопитающих вирусные серпины содержат значительные делеции элементов вторичной структуры. В частности, у crmA отсутствует D-спираль, а также значительные части A- и E-спиралей. [ 212 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и Сильверман Г.А., Бёрд П.И., Каррелл Р.В., Черч ФК, Кофлин П.Б., Геттинс П.Г., Ирвинг Дж.А., Ломас Д.А., Люк С.Дж., Мойер Р.В., Пембертон П.А., Ремолд-О'Доннелл Э., Салвесен Г.С., Трэвис Дж., Уиссток Дж.К. (сентябрь) 2001). «Серпины представляют собой расширяющееся суперсемейство структурно сходных, но функционально разнообразных белков. Эволюция, механизм ингибирования, новые функции и пересмотренная номенклатура» . Журнал биологической химии . 276 (36): 33293–33296. дои : 10.1074/jbc.R100016200 . ПМИД 11435447 .
- ^ Спенс М.А., Мортимер М.Д., Бакл А.М., Минь Б.К., Джексон С.Дж. (июнь 2021 г.). Эчаве Дж. (ред.). «Комплексный филогенетический анализ надсемейства серпинов» . Молекулярная биология и эволюция . 38 (7): 2915–2929. дои : 10.1093/molbev/msab081 . ПМЦ 8233489 . ПМИД 33744972 .
- ^ Каррелл Р.В., Босуэлл Д.Р. (1986). «Серпины: суперсемейство ингибиторов сериновых протеиназ плазмы». В Барретт А.Дж., Салвесен Дж. (ред.). Ингибиторы протеиназ . Научные монографии по физиологии клеток и тканей. Том. 12. Амстердам: Elsevier Science Publishers BV . стр. 403–420. ISBN 0-444-80763-2 .
- ^ Сильверман Г.А., Уиссток Дж.К., Боттомли С.П., Хантингтон Дж.А., Кайзерман Д., Люк С.Дж., Пак С.К., Райххарт Дж.М., Берд ПИ (август 2010 г.). «Змеи тренируют мышцы: I. Остановка протеолиза в различных биологических системах» . Журнал биологической химии . 285 (32): 24299–24305. дои : 10.1074/jbc.R110.112771 . ПМЦ 2915665 . ПМИД 20498369 .
- ^ Уиссток Дж.К., Сильверман Г.А., Берд П.И., Боттомли С.П., Кайзерман Д., Люк С.Дж., Пак С.К., Райххарт Дж.М., Хантингтон Дж.А. (август 2010 г.). «Змеи тренируют мышцы: II. Структурные данные о распознавании, полимеризации и транспортировке целевой пептидазы» . Журнал биологической химии . 285 (32): 24307–24312. дои : 10.1074/jbc.R110.141408 . ПМЦ 2915666 . ПМИД 20498368 .
- ^ Перейти обратно: а б с д Хантингтон Дж. А., Рид Р. Дж., Каррел Р. В. (октябрь 2000 г.). «Структура комплекса серпин-протеаза демонстрирует ингибирование за счет деформации». Природа . 407 (6806): 923–926. Бибкод : 2000Natur.407..923H . дои : 10.1038/35038119 . ПМИД 11057674 . S2CID 205009937 .
- ^ Перейти обратно: а б с д и ж г Геттинс П.Г. (декабрь 2002 г.). «Структура, механизм и функции серпина». Химические обзоры . 102 (12): 4751–4804. дои : 10.1021/cr010170 . ПМИД 12475206 .
- ^ Перейти обратно: а б с д и ж г Уиссток Дж.К., Боттомли СП (декабрь 2006 г.). «Молекулярная гимнастика: змеевидная структура, сворачивание и неправильное сворачивание». Современное мнение в области структурной биологии . 16 (6): 761–768. дои : 10.1016/j.sbi.2006.10.005 . ПМИД 17079131 .
- ^ Перейти обратно: а б с д и ж Лоу Р.Х., Чжан К., Макгоуэн С., Бакл А.М., Сильверман Г.А., Вонг В., Росадо С.Дж., Лангендорф К.Г., Пайк Р.Н., Берд П.И., Уиссток Дж.К. (2006). «Обзор надсемейства серпинов» . Геномная биология . 7 (5): 216. doi : 10.1186/gb-2006-7-5-216 . ПМЦ 1779521 . ПМИД 16737556 .
- ^ Перейти обратно: а б с д и Стейн П.Е., Каррел Р.В. (февраль 1995 г.). «Что дисфункциональные серпины говорят нам о молекулярной подвижности и заболеваниях?». Структурная биология природы . 2 (2): 96–113. дои : 10.1038/nsb0295-96 . ПМИД 7749926 . S2CID 21223825 .
- ^ Перейти обратно: а б с д и ж г час я Янчаускене С.М., Балс Р., Кочулла Р., Фогельмайер С., Кёнляйн Т., Вельте Т. (август 2011 г.). «Открытие α1-антитрипсина и его роль в здоровье и болезнях» . Респираторная медицина . 105 (8): 1129–1139. дои : 10.1016/j.rmed.2011.02.002 . ПМИД 21367592 .
- ^ Перейти обратно: а б с д Каррелл Р.В., Ломас Д.А. (июль 1997 г.). «Конформационная болезнь». Ланцет . 350 (9071): 134–138. дои : 10.1016/S0140-6736(97)02073-4 . ПМИД 9228977 . S2CID 39124185 .
- ^ Ферми С., Перносси Л. (декабрь 1894 г.). «Исследование фермента, сравнительное исследование» ( PDF) . Журнал гигиены и инфекционных заболеваний (на немецком языке) (18): 83–89. дои : 10.1007/BF02216836 . S2CID 24373770 .
- ^ Шульце Х.У., Гёлльнер И., Хайде К., Шёненбергер М., Швик Г. (август 1955 г.). «За знание альфа-глобулина нормальных сывороток человека» . Журнал Nature Research B (на немецком языке). 10 (8): 463. doi : 10.1515/znb-1955-0810 . S2CID 95960716 .
- ^ Петерсен Т.Е., Дудек-Войцеховска Г., Соттруп-Йенсен Л., Магнуссон С. (1979). «Первичная структура антитромбина III (кофактора гепарина): частичная гомология между альфа-1-антитрипсином и антитромбином III». В Коллен Д., Виман Б., Верстраете М. (ред.). Физиологические ингибиторы коагуляции и фибринолиза . Амстердам: Эльзевир . стр. 43–54.
- ^ Каррелл Р., Оуэн М., Бреннан С., Воган Л. (декабрь 1979 г.). «Карбокси-концевой фрагмент человеческого альфа-1-антитрипсина после расщепления гидроксиламина: гомология с антитромбином III». Связь с биохимическими и биофизическими исследованиями . 91 (3): 1032–1037. дои : 10.1016/0006-291X(79)91983-1 . ПМИД 316698 .
- ^ Хант LT, Дайхофф, Миссури (июль 1980 г.). «Удивительное новое суперсемейство белков, содержащее овальбумин, антитромбин-III и ингибитор альфа-1-протеиназы». Связь с биохимическими и биофизическими исследованиями . 95 (2): 864–871. дои : 10.1016/0006-291X(80)90867-0 . ПМИД 6968211 .
- ^ Каррелл Р.В., Джеппссон Дж.О., Лорелл С.Б., Бреннан С.О., Оуэн М.К., Воган Л., Босуэлл Д.Р. (июль 1982 г.). «Структура и вариация альфа-1-антитрипсина человека» . Природа . 298 (5872): 329–334. дои : 10.1016/0968-0004(85)90011-8 . ПМК 7172600 . ПМИД 7045697 .
- ^ Лорел CB, Эрикссон С (март 2013 г.). «Электрофоретический характер α1-глобулина сыворотки при дефиците α1-антитрипсина. 1963» . ХОБЛ . 10 (Приложение 1): 3–8. дои : 10.3109/15412555.2013.771956 . ПМИД 23527532 . S2CID 36366089 .
- ^ Шарп Х.Л., Бриджес Р.А., Кривит В., Фрейер Э.Ф. (июнь 1969 г.). «Цирроз печени, связанный с дефицитом альфа-1-антитрипсина: ранее невыявленное наследственное заболевание» . Журнал лабораторной и клинической медицины . 73 (6): 934–939. ПМИД 4182334 .
- ^ «Альфа-1-антитрипсин: молекулярная аномалия S-варианта» . Британский медицинский журнал . 1 (6002): 130–131. Январь 1976 г. doi : 10.1136/bmj.1.6002.130-a . ПМЦ 1638590 . ПМИД 1082356 .
- ^ Джеппссон Д.О. (июнь 1976 г.). «Аминокислотная замена Glu приводит к образованию Lys-альфа1-антитрипсина PiZ» . Письма ФЭБС . 65 (2): 195–197. дои : 10.1016/0014-5793(76)80478-4 . ПМИД 1084290 . S2CID 84576569 .
- ^ Каррелл Р.В., Джеппссон Дж.О., Лорелл С.Б., Бреннан С.О., Оуэн М.К., Воган Л., Босуэлл Д.Р. (июль 1982 г.). «Структура и вариация альфа-1-антитрипсина человека». Природа . 298 (5872): 329–334. Бибкод : 1982Natur.298..329C . дои : 10.1038/298329a0 . ПМИД 7045697 . S2CID 11904305 .
- ^ Каррелл Р.В., Босуэлл Д.Р., Бреннан С.О., Оуэн М.С. (март 1980 г.). «Активный сайт альфа-1-антитрипсина: гомологичный сайт в антитромбине-III». Связь с биохимическими и биофизическими исследованиями . 93 (2): 399–402. дои : 10.1016/0006-291X(80)91090-6 . ПМИД 6966929 .
- ^ Джонсон Д., Трэвис Дж. (октябрь 1978 г.). «Структурные доказательства наличия метионина в реактивном сайте ингибитора альфа-1-протеиназы человека» . Журнал биологической химии . 253 (20): 7142–7144. дои : 10.1016/S0021-9258(17)34475-7 . ПМИД 701239 .
- ^ Йорнвалль Х., Фиш WW, Бьорк I (октябрь 1979 г.). «Сайт расщепления тромбина в бычьем антитромбине» . Письма ФЭБС . 106 (2): 358–362. Бибкод : 1979FEBSL.106..358J . дои : 10.1016/0014-5793(79)80532-3 . ПМИД 499520 . S2CID 32540550 .
- ^ Эгеберг О. (июнь 1965 г.). «Наследственный дефицит антитромбина, вызывающий тромбофилию». Тромбоз и геморрагический диатез . 13 (2): 516–530. дои : 10.1055/s-0038-1656297 . ПМИД 14347873 . S2CID 42594050 .
- ^ Перейти обратно: а б Оуэн MC, Бреннан С.О., Льюис Дж.Х., Каррелл Р.В. (сентябрь 1983 г.). «Мутация антитрипсина в антитромбин. Альфа-1-антитрипсин Питтсбург (358 Met приводит к Arg), смертельное нарушение свертываемости крови». Медицинский журнал Новой Англии . 309 (12): 694–698. дои : 10.1056/NEJM198309223091203 . ПМИД 6604220 .
- ^ Перейти обратно: а б Леберманн Х., Токуока Р., Дайзенхофер Дж., Хубер Р. (август 1984 г.). «Ингибитор альфа-1-протеиназы человека. Анализ кристаллической структуры двух кристаллических модификаций, молекулярная модель и предварительный анализ влияния на функцию». Журнал молекулярной биологии . 177 (3): 531–557. дои : 10.1016/0022-2836(84)90298-5 . ПМИД 6332197 .
- ^ Перейти обратно: а б Стейн П.Е., Лесли А.Г., Финч Дж.Т., Тернелл В.Г., Маклафлин П.Дж., Каррелл Р.В. (сентябрь 1990 г.). «Кристаллическая структура овальбумина как модель реактивного центра серпинов». Природа . 347 (6288): 99–102. Бибкод : 1990Natur.347...99S . дои : 10.1038/347099a0 . ПМИД 2395463 . S2CID 4342263 .
- ^ Перейти обратно: а б Каррелл Р.В., Эванс Д.Л., Стейн П.Е. (октябрь 1991 г.). «Мобильный реактивный центр серпинов и контроль тромбообразования» . Природа . 353 (6344): 576–578. Бибкод : 1991Natur.353..576C . дои : 10.1038/353576a0 . ПМИД 1922367 . S2CID 4361304 .
- ^ Моттонен Дж., Стрэнд А., Симерски Дж., Свит Р.М., Дэнли Д.Е., Геохеган К.Ф., Джерард Р.Д., Голдсмит Э.Дж. (январь 1992 г.). «Структурные основы латентного периода ингибитора активатора плазминогена-1». Природа . 355 (6357): 270–273. Бибкод : 1992Natur.355..270M . дои : 10.1038/355270a0 . ПМИД 1731226 . S2CID 4365370 .
- ^ Остин Р.К., Рачубински Р.А., Офосу Ф.А., Блайхман М.А. (май 1991 г.). «Антитромбин-III-Гамильтон, от Ala 382 до Thr: вариант антитромбина-III, который действует как субстрат, но не ингибитор альфа-тромбина и фактора Ха» . Кровь . 77 (10): 2185–2189. дои : 10.1182/blood.V77.10.2185.2185 . ПМИД 2029579 .
- ^ Перейти обратно: а б с Хантингтон Дж. А. (август 2006 г.). «Змеи, меняющие форму - преимущества мобильного механизма». Тенденции биохимических наук . 31 (8): 427–435. дои : 10.1016/j.tibs.2006.06.005 . ПМИД 16820297 .
- ^ Каррелл Р.В., Рид Р.Дж. (февраль 2017 г.). «Как серпины транспортируют гормоны и регулируют их высвобождение» . Семинары по клеточной биологии и биологии развития . 62 : 133–141. дои : 10.1016/j.semcdb.2016.12.007 . ПМИД 28027946 .
- ^ Перейти обратно: а б с Ирвинг Дж.А., Пайк Р.Н., Леск А.М., Уиссток Дж.К. (декабрь 2000 г.). «Филогения суперсемейства серпинов: последствия закономерностей консервации аминокислот для структуры и функции» . Геномные исследования . 10 (12): 1845–1864. дои : 10.1101/gr.GR-1478R . ПМИД 11116082 .
- ^ Перейти обратно: а б Ирвинг Дж.А., Стинбакерс П.Дж., Леск А.М., Оп ден Кэмп Х.Дж., Пайк Р.Н., Уиссток Дж.К. (ноябрь 2002 г.). «Зерпины у прокариот» . Молекулярная биология и эволюция . 19 (11): 1881–1890. doi : 10.1093/oxfordjournals.molbev.a004012 . ПМИД 12411597 .
- ^ Перейти обратно: а б Стинбакерс П.Дж., Ирвинг Дж.А., Харханги Х.Р., Свинкелс В.Дж., Ахманова А., Дийкерман Р., Джеттен М.С., ван дер Дрифт С., Уиссток Дж.К., Оп ден Кэмп Х.Дж. (август 2008 г.). «Серпин в целлюлозоме анаэробного гриба Piromyces sp., штамм E2» . Микологические исследования . 112 (Часть 8): 999–1006. дои : 10.1016/j.mycres.2008.01.021 . hdl : 2066/72679 . ПМИД 18539447 .
- ^ Хубер Р., Каррел Р.В. (ноябрь 1989 г.). «Влияние трехмерной структуры альфа-1-антитрипсина на структуру и функцию серпинов». Биохимия . 28 (23): 8951–8966. дои : 10.1021/bi00449a001 . ПМИД 2690952 .
- ^ Перейти обратно: а б Роулингс Н.Д., Толле Д.П., Барретт А.Дж. (март 2004 г.). «Эволюционные семейства ингибиторов пептидаз» . Биохимический журнал . 378 (Часть 3): 705–716. дои : 10.1042/BJ20031825 . ПМК 1224039 . ПМИД 14705960 .
- ^ Барретт А.Дж., Роулингс, Северная Дакота (апрель 1995 г.). «Семейства и кланы сериновых пептидаз». Архив биохимии и биофизики . 318 (2): 247–250. дои : 10.1006/abbi.1995.1227 . ПМИД 7733651 .
- ^ Барретт А.Дж., Роулингс, Северная Дакота (май 2001 г.). «Эволюционные линии цистеинпептидаз» . Биологическая химия . 382 (5): 727–733. дои : 10.1515/BC.2001.088 . ПМИД 11517925 . S2CID 37306786 .
- ^ Ирвинг Дж.А., Пайк Р.Н., Дай В., Бромме Д., Уорролл Д.М., Сильверман Г.А., Кетцер Т.Х., Деннисон С., Боттомли С.П., Уиссток Дж.К. (апрель 2002 г.). «Доказательства того, что архитектура серпинов по своей сути поддерживает ингибирование папаин-подобной цистеиновой протеазы: разработка альфа (1)-антитрипсина для ингибирования катепсиновых протеаз». Биохимия . 41 (15): 4998–5004. дои : 10.1021/bi0159985 . ПМИД 11939796 .
- ^ Перейти обратно: а б Шик С., Бромме Д., Бартуски А.Дж., Уэмура Ю., Шехтер Н.М., Сильверман Г.А. (ноябрь 1998 г.). «Петля реактивного центра серпина SCCA1 необходима для ингибирования цистеиновой протеиназы» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (23): 13465–13470. Бибкод : 1998PNAS...9513465S . дои : 10.1073/pnas.95.23.13465 . ПМК 24842 . ПМИД 9811823 .
- ^ Перейти обратно: а б Макгоуэн С., Бакл А.М., Ирвинг Дж.А., Онг ПК, Баштанник-Пухалович Т.А., Кан В.Т., Хендерсон К.Н., Булынко Ю.А., Попова Е.Ю., Смит А.И., Боттомли С.П., Россджон Дж., Григорьев С.А., Пайк Р.Н., Уиссток Дж.К. (июль 2006 г.) . «Рентгеновская кристаллическая структура МЕНТ: доказательства существования функциональных петлевых полимеров при конденсации хроматина» . Журнал ЭМБО . 25 (13): 3144–3155. дои : 10.1038/sj.emboj.7601201 . ПМК 1500978 . ПМИД 16810322 .
- ^ Онг ПК, Макгоуэн С., Пирс М.С., Ирвинг Дж.А., Кан В.Т., Григорьев С.А., Терк Б., Сильверман Г.А., Брикс К., Боттомли С.П., Уиссток Дж.К., Пайк Р.Н. (декабрь 2007 г.). «ДНК ускоряет ингибирование человеческого катепсина V серпинами» . Журнал биологической химии . 282 (51): 36980–36986. дои : 10.1074/jbc.M706991200 . ПМИД 17923478 .
- ^ Перейти обратно: а б Акоста Х., Илиев Д., Гран Т.Х., Гуиньяр Н., Маккарана М., Грисбах Дж., Херцманн С., Сага М., Климент М., Пера Э.М. (март 2015 г.). «Серпин PN1 является регулятором обратной связи передачи сигналов FGF в формировании зародышевого слоя и первичной оси» . Разработка . 142 (6): 1146–1158. дои : 10.1242/dev.113886 . ПМИД 25758225 .
- ^ Перейти обратно: а б с Хасимото С., Ким Д.Р., Вайс Л.А., Миллер Дж.В., Морисато Д. (декабрь 2003 г.). «Пространственная регуляция передачи сигналов развития серпином» . Развивающая клетка . 5 (6): 945–950. дои : 10.1016/S1534-5807(03)00338-1 . ПМИД 14667416 .
- ^ Берд ПИ (февраль 1999 г.). «Регуляция проапоптотических сериновых протеиназ лейкоцитарных гранул с помощью внутриклеточных серпинов». Иммунология и клеточная биология . 77 (1): 47–57. дои : 10.1046/j.1440-1711.1999.00787.x . ПМИД 10101686 . S2CID 44268106 .
- ^ Берд CH, Саттон В.Р., Сан Дж., Херст CE, Новак А., Кумар С., Трапани Дж.А., Берд ПИ (ноябрь 1998 г.). «Селективная регуляция апоптоза: цитотоксический ингибитор серпинпротеиназы 9 лимфоцитов защищает от апоптоза, опосредованного гранзимом B, не нарушая путь гибели Fas-клеток» . Молекулярная и клеточная биология . 18 (11): 6387–6398. дои : 10.1128/mcb.18.11.6387 . ПМК 109224 . ПМИД 9774654 .
- ^ Рэй К.А., Блэк Р.А., Кронхейм С.Р., Гринстрит Т.А., Слит PR, Салвесен Г.С., Pickup DJ (май 1992 г.). «Вирусное ингибирование воспаления: вирус коровьей оспы кодирует ингибитор бета-превращающего фермента интерлейкина-1». Клетка . 69 (4): 597–604. дои : 10.1016/0092-8674(92)90223-Y . ПМИД 1339309 . S2CID 7398844 .
- ^ Перейти обратно: а б Веркаммен Д., Беленги Б., ван де Котт Б., Бёненс Т., Гавиган Дж.А., Де Райк Р., Бракеньер А., Инзе Д., Харрис Дж.Л., Ван Бреузегем Ф. (декабрь 2006 г.). «Серпин1 Arabidopsis thaliana является самоубийственным ингибитором метакаспазы 9». Журнал молекулярной биологии . 364 (4): 625–636. дои : 10.1016/j.jmb.2006.09.010 . ПМИД 17028019 .
- ^ Перейти обратно: а б Лампл Н., Будай-Адриан О., Давыдов О., Джосс Т.В., Харроп С.Дж., Курми П.М., Робертс Т.Х., Флюр Р. (апрель 2010 г.). «Arabidopsis AtSerpin1, кристаллическая структура и взаимодействие in vivo с целевой протеазой, РЕАГИРУЮЩЕЙ НА ВЫСУШЕНИЕ-21 (RD21)» . Журнал биологической химии . 285 (18): 13550–13560. дои : 10.1074/jbc.M109.095075 . ПМЦ 2859516 . ПМИД 20181955 .
- ^ Перейти обратно: а б с Клибер М.А., Андерхилл С., Хаммонд Г.Л., Мюллер Ю.А. (октябрь 2007 г.). «Кортикостероидсвязывающий глобулин, структурная основа транспорта стероидов и высвобождения, запускаемого протеиназами» . Журнал биологической химии . 282 (40): 29594–29603. дои : 10.1074/jbc.M705014200 . ПМИД 17644521 .
- ^ Перейти обратно: а б с д и Чжоу А., Вэй З., Рид Р.Дж., Каррелл Р.В. (сентябрь 2006 г.). «Структурный механизм переноса и высвобождения тироксина в кровь» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (36): 13321–13326. Бибкод : 2006PNAS..10313321Z . дои : 10.1073/pnas.0604080103 . ПМЦ 1557382 . ПМИД 16938877 .
- ^ Хантингтон Дж. А., Штейн П. Е. (май 2001 г.). «Структура и свойства овальбумина». Журнал хроматографии. Б. Биомедицинские науки и их применения . 756 (1–2): 189–198. дои : 10.1016/S0378-4347(01)00108-6 . ПМИД 11419711 .
- ^ Перейти обратно: а б с Мала Дж.Г., Роуз С. (ноябрь 2010 г.). «Взаимодействие белка теплового шока 47 с коллагеном и реакция на стресс: нетрадиционная модель шаперона?». Науки о жизни . 87 (19–22): 579–586. дои : 10.1016/j.lfs.2010.09.024 . ПМИД 20888348 .
- ^ Григорьев С.А., Беднар Дж., Вудкок С.Л. (февраль 1999 г.). «MENT, белок гетерохроматина, который обеспечивает сворачивание хроматина более высокого порядка, является новым членом семейства серпинов» . Журнал биологической химии . 274 (9): 5626–5636. дои : 10.1074/jbc.274.9.5626 . ПМИД 10026180 .
- ^ Эллиотт П.Р., Ломас Д.А., Каррелл Р.В., Абрахамс Дж.П. (август 1996 г.). «Ингибирующая конформация реактивной петли альфа-1-антитрипсина». Структурная биология природы . 3 (8): 676–681. дои : 10.1038/nsb0896-676 . PMID 8756325 . S2CID 22976014 .
- ^ Хорват А.Дж., Ирвинг Дж.А., Россджон Дж., Лоу Р.Х., Боттомли С.П., Куинси Н.С., Пайк Р.Н., Кофлин П.Б., Уиссток Дж.К. (декабрь 2005 г.). «Мышиный ортолог человеческого антихимотрипсина: структурная парадигма серпинов клады А3» . Журнал биологической химии . 280 (52): 43168–43178. дои : 10.1074/jbc.M505598200 . ПМИД 16141197 .
- ^ Уиссток Дж.К., Скиннер Р., Каррелл Р.В., Леск А.М. (февраль 2000 г.). «Конформационные изменения серпинов: I. Нативная и расщепленная конформации альфа (1)-антитрипсина». Журнал молекулярной биологии . 296 (2): 685–699. дои : 10.1006/jmbi.1999.3520 . ПМИД 10669617 .
- ^ Джин Л., Абрахамс Дж.П., Скиннер Р., Петиту М., Пайк Р.Н., Каррелл Р.В. (декабрь 1997 г.). «Антикоагулянтная активация антитромбина гепарином» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (26): 14683–14688. Бибкод : 1997PNAS...9414683J . дои : 10.1073/pnas.94.26.14683 . ПМК 25092 . ПМИД 9405673 .
- ^ Уиссток Дж.К., Пайк Р.Н., Джин Л., Скиннер Р., Пей XY, Каррелл Р.В., Леск А.М. (сентябрь 2000 г.). «Конформационные изменения серпинов: II. Механизм активации антитромбина гепарином». Журнал молекулярной биологии . 301 (5): 1287–1305. дои : 10.1006/jmbi.2000.3982 . ПМИД 10966821 .
- ^ Ли В., Джонсон DJ, Эсмон CT, Хантингтон JA (сентябрь 2004 г.). «Структура тройного комплекса антитромбин-тромбин-гепарин раскрывает антитромботический механизм гепарина». Структурная и молекулярная биология природы . 11 (9): 857–862. дои : 10.1038/nsmb811 . ПМИД 15311269 . S2CID 28790576 .
- ^ Джонсон DJ, Ли В., Адамс Т.Е., Хантингтон Дж.А. (май 2006 г.). «Структура антитромбина-S195A фактора Ха-гепарина раскрывает аллостерический механизм активации антитромбина» . Журнал ЭМБО . 25 (9): 2029–2037. дои : 10.1038/sj.emboj.7601089 . ПМЦ 1456925 . ПМИД 16619025 .
- ^ Валенга Дж. М., Джеске В. П., Самама М. М., Frapaise FX, Бик Р. Л., Фарид Дж. (март 2002 г.). «Фондапаринукс: синтетический пентасахарид гепарина как новый антитромботический агент». Экспертное заключение об исследуемых препаратах . 11 (3): 397–407. дои : 10.1517/13543784.11.3.397 . ПМИД 11866668 . S2CID 24796086 .
- ^ Петиту М., ван Бекель, Калифорния (июнь 2004 г.). «Синтетический пентасахарид, связывающий антитромбин III, теперь является лекарством! Что будет дальше?». Ангеванде Хеми . 43 (24): 3118–3133. дои : 10.1002/anie.200300640 . ПМИД 15199558 .
- ^ Перейти обратно: а б Линдал Т.Л., Сигурдардоттир О., Виман Б. (сентябрь 1989 г.). «Стабильность ингибитора активатора плазминогена 1 (PAI-1)». Тромбоз и гемостаз . 62 (2): 748–751. дои : 10.1055/s-0038-1646895 . ПМИД 2479113 . S2CID 19433778 .
- ^ Мушуньдже А., Эванс Дж., Бреннан С.О., Каррелл Р.В., Чжоу А. (декабрь 2004 г.). «Латентный антитромбин и его обнаружение, образование и оборот в кровообращении» . Журнал тромбозов и гемостаза . 2 (12): 2170–2177. дои : 10.1111/j.1538-7836.2004.01047.x . ПМИД 15613023 . S2CID 43029244 .
- ^ Чжан К., Бакл А.М., Лоу Р.Х., Пирс М.К., Кабрита Л.Д., Ллойд Г.Дж., Ирвинг Дж.А., Смит А.И., Рузыла К., Россджон Дж., Боттомли С.П., Уиссток Дж.К. (июль 2007 г.). «N-конец серпина, тенгпин, удерживает метастабильное нативное состояние» . Отчеты ЭМБО . 8 (7): 658–663. дои : 10.1038/sj.embor.7400986 . ПМК 1905895 . ПМИД 17557112 .
- ^ Чжан Кью, Лоу Р.Х., Боттомли С.П., Уиссток Дж.К., Бакл А.М. (март 2008 г.). «Структурная основа полимеризации петлевых C-листов в серпинах». Журнал молекулярной биологии . 376 (5): 1348–1359. дои : 10.1016/j.jmb.2007.12.050 . ПМИД 18234218 .
- ^ Пембертон П.А., Стейн П.Е., Пепис М.Б., Поттер Дж.М., Каррелл Р.В. (ноябрь 1988 г.). «Гормонсвязывающие глобулины претерпевают конформационные изменения серпина при воспалении». Природа . 336 (6196): 257–258. Бибкод : 1988Natur.336..257P . дои : 10.1038/336257a0 . ПМИД 3143075 . S2CID 4326356 .
- ^ Перейти обратно: а б с Цао С., Лоуренс Д.А., Ли Ю., Фон Арним К.А., Герц Дж., Су Э.Дж., Макарова А., Хайман Б.Т., Стрикленд Д.К., Чжан Л. (май 2006 г.). «Эндоцитарный рецептор LRP вместе с tPA и PAI-1 координирует Mac-1-зависимую миграцию макрофагов» . Журнал ЭМБО . 25 (9): 1860–1870. дои : 10.1038/sj.emboj.7601082 . ПМК 1456942 . ПМИД 16601674 .
- ^ Дженсен Дж. К., Долмер К., Геттинс П. Г. (июль 2009 г.). «Специфичность связывания белка, родственного рецептору липопротеина низкой плотности, с различными конформационными состояниями ингибитора-1 активатора плазминогена клады E серпинов и протеиназы нексина-1» . Журнал биологической химии . 284 (27): 17989–17997. дои : 10.1074/jbc.M109.009530 . ПМК 2709341 . ПМИД 19439404 .
- ^ Соукуп С.Ф., Кули Дж., Габб Д. (июнь 2009 г.). Рулифсон Э (ред.). «Поглощение некротического серпина у Drosophila melanogaster через рецептор липофорина-1» . ПЛОС Генетика . 5 (6): e1000532. дои : 10.1371/journal.pgen.1000532 . ПМЦ 2694266 . ПМИД 19557185 .
- ^ Кайзерман Д., Уиссток Дж.К., Берд П.И. (декабрь 2006 г.). «Механизмы дисфункции серпинов при заболеваниях». Обзоры экспертов в области молекулярной медицины . 8 (31): 1–19. дои : 10.1017/S1462399406000184 . ПМИД 17156576 . S2CID 20760165 .
- ^ де Серр FJ (ноябрь 2002 г.). «Расовое и этническое распределение дефицита альфа1-антитрипсина во всем мире: резюме анализа опубликованных генетических эпидемиологических исследований». Грудь . 122 (5): 1818–1829. дои : 10.1378/сундук.122.5.1818 . ПМИД 12426287 .
- ^ Хопкинс ПК, Каррелл Р.В., Стоун С.Р. (август 1993 г.). «Эффекты мутаций в шарнирной области серпинов». Биохимия . 32 (30): 7650–7657. дои : 10.1021/bi00081a008 . ПМИД 8347575 .
- ^ Бошан, Нью-Джерси, Пайк Р.Н., Дейли М., Батлер Л., Макрис М., Даффорн Т.Р., Чжоу А., Фиттон Х.Л., Престон Ф.Е., Пик И.Р., Каррелл Р.В. (октябрь 1998 г.). «Антитромбиновое колебание и колебание (T85M/K): архетипические конформационные заболевания с латентным переходом in vivo, тромбозом и активацией гепарина». Кровь . 92 (8): 2696–2706. дои : 10.1182/blood.V92.8.2696 . ПМИД 9763552 .
- ^ Перейти обратно: а б с Гупту Б., Хейзес Б., Чанг В.С., Даффорн Т.Р., Каррелл Р.В., Рид Р.Дж., Ломас Д.А. (январь 2000 г.). «Неактивная конформация серпина альфа(1)-антихимотрипсина указывает на двухэтапное включение реактивной петли: последствия для тормозной функции и конформационного заболевания» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (1): 67–72. Бибкод : 2000ПНАС...97...67Г . дои : 10.1073/pnas.97.1.67 . ПМЦ 26617 . ПМИД 10618372 .
- ^ Перейти обратно: а б Хоман Э.П., Раух Ф., Граф И., Литман С., Долл Дж.А., Доусон Б., Бертин Т., Напиерала Д., Морелло Р., Гиббс Р., Уайт Л., Мики Р., Кон Д.Х., Кроуфорд С., Трэверс Р., Глориё Ф.Х., Ли Б. (декабрь 2011 г.). «Мутации в SERPINF1 вызывают несовершенный остеогенез VI типа» . Журнал исследований костей и минералов . 26 (12): 2798–2803. дои : 10.1002/jbmr.487 . ПМК 3214246 . ПМИД 21826736 .
- ^ Фэй В.П., Паркер А.С., Кондри Л.Р., Шапиро А.Д. (июль 1997 г.). «Дефицит человеческого ингибитора активатора плазминогена-1 (PAI-1): характеристика крупного родственника с нулевой мутацией в гене PAI-1» . Кровь . 90 (1): 204–208. дои : 10.1182/blood.V90.1.204 . ПМИД 9207454 .
- ^ Перейти обратно: а б с д и ж Хейт С., Джексон BC, МакЭндрюс М., Райт М.В., Томпсон Д.С., Сильверман Г.А., Неберт Д.В., Василиу В. (октябрь 2013 г.). «Обновление суперсемейства генов SERPIN человека и мыши» . Геномика человека . 7 (1): 22. дои : 10.1186/1479-7364-7-22 . ПМЦ 3880077 . ПМИД 24172014 .
- ^ Перейти обратно: а б Ломас Д.А., Эванс Д.Л., Финч Дж.Т., Каррелл Р.В. (июнь 1992 г.). «Механизм накопления Z альфа 1-антитрипсина в печени». Природа . 357 (6379): 605–607. Бибкод : 1992Natur.357..605L . дои : 10.1038/357605a0 . ПМИД 1608473 . S2CID 4359543 .
- ^ Крегер Х., Миранда Э., МакЛауд И., Перес Дж., Кроутер Д.К., Марчиниак С.Дж., Ломас Д.А. (август 2009 г.). «Деградация, связанная с эндоплазматическим ретикулумом (ERAD), и аутофагия взаимодействуют, разрушая полимерогенные мутантные серпины» . Журнал биологической химии . 284 (34): 22793–22802. дои : 10.1074/jbc.M109.027102 . ПМЦ 2755687 . ПМИД 19549782 .
- ^ Перейти обратно: а б с Ямасаки М., Ли В., Джонсон DJ, Хантингтон Дж.А. (октябрь 2008 г.). «Кристаллическая структура стабильного димера раскрывает молекулярную основу полимеризации серпинов». Природа . 455 (7217): 1255–1258. Бибкод : 2008Natur.455.1255Y . дои : 10.1038/nature07394 . ПМИД 18923394 . S2CID 205215121 .
- ^ Перейти обратно: а б Боттомли SP (сентябрь 2011 г.). «Структурное разнообразие неправильного сворачивания α1-антитрипсина» . Отчеты ЭМБО . 12 (10): 983–984. дои : 10.1038/embor.2011.187 . ПМК 3185355 . ПМИД 21921939 .
- ^ Перейти обратно: а б Ямасаки М., Сендалл Т.Дж., Пирс М.К., Уиссток Дж.К., Хантингтон Дж.А. (сентябрь 2011 г.). «Молекулярные основы дефицита α1-антитрипсина, выявленные структурой тримера с замененным доменом» . Отчеты ЭМБО . 12 (10): 1011–1017. дои : 10.1038/embor.2011.171 . ПМЦ 3185345 . ПМИД 21909074 .
- ^ Чанг В.С., Уиссток Дж., Хопкинс ПК, Леск А.М., Каррелл Р.В., Уорделл М.Р. (январь 1997 г.). «Важность высвобождения цепи 1C для механизма полимеризации ингибирующих серпинов» . Белковая наука . 6 (1): 89–98. дои : 10.1002/pro.5560060110 . ПМК 2143506 . ПМИД 9007980 .
- ^ Миранда Э., Перес Дж., Экеова У.И., Хаджич Н., Калшекер Н., Гоопту Б., Портманн Б., Белорджи Д., Хилл М., Чемберс С., Текман Дж., Александр Г.Дж., Марчиняк С.Дж., Ломас Д.А. (сентябрь 2010 г.). «Новое моноклональное антитело для характеристики патогенных полимеров при заболеваниях печени, связанных с дефицитом альфа1-антитрипсина». Гепатология . 52 (3): 1078–1088. дои : 10.1002/hep.23760 . ПМИД 20583215 . S2CID 8188156 .
- ^ Сандхаус РА (октябрь 2004 г.). «Дефицит альфа1-антитрипсина. 6: новые и новые методы лечения дефицита альфа1-антитрипсина» . Торакс . 59 (10): 904–909. дои : 10.1136/thx.2003.006551 . ПМЦ 1746849 . ПМИД 15454659 .
- ^ Льюис ЕС (сентябрь 2012 г.). «Расширение клинических показаний к терапии α(1)-антитрипсином» . Молекулярная медицина . 18 (6): 957–970. дои : 10.2119/molmed.2011.00196 . ПМЦ 3459478 . ПМИД 22634722 .
- ^ Фрегонезе Л., Столк Дж. (июнь 2008 г.). «Наследственный дефицит альфа-1-антитрипсина и его клинические последствия» . Сиротский журнал редких заболеваний . 3:16 . дои : 10.1186/1750-1172-3-16 . ПМК 2441617 . ПМИД 18565211 .
- ^ Юса К., Рашид С.Т., Стрик-Маршан Х., Варела И., Лю П.К., Пашон Д.Э., Миранда Э., Ордоньес А., Ханнан Н.Р., Роухани Ф.Дж., Дарче С., Александр Дж., Марчиняк С.Дж., Фусаки Н., Хасегава М., Холмс М.К., Ди Санто Дж.П., Ломас Д.А., Брэдли А., Валье Л. (октябрь 2011 г.). «Направленная генная коррекция дефицита α1-антитрипсина в индуцированных плюрипотентных стволовых клетках» . Природа . 478 (7369): 391–394. Бибкод : 2011Природа.478..391Y . дои : 10.1038/nature10424 . ПМК 3198846 . ПМИД 21993621 .
- ^ Малля М, Филлипс Р.Л., Салданха С.А., Гоопту Б, Браун С.С., Термине DJ, Ширвани А.М., Ву Ю, Сиферс Р.Н., Абагян Р., Ломас Д.А. (ноябрь 2007 г.). «Маленькие молекулы блокируют полимеризацию Z-альфа1-антитрипсина и увеличивают клиренс внутриклеточных агрегатов» . Журнал медицинской химии . 50 (22): 5357–5363. дои : 10.1021/jm070687z . ПМЦ 2631427 . ПМИД 17918823 .
- ^ Госай С.Дж., Квак Дж.Х., Люк С.Дж., Лонг О.С., Кинг Д.Е., Ковач К.Дж., Джонстон П.А., Шун Т.Я., Лазо Дж.С., Перлмуттер Д.Х., Сильверман Г.А., Пак С.С. (ноябрь 2010 г.). «Автоматизированный скрининг препаратов с высоким содержанием живых животных с использованием C. elegans, экспрессирующих склонный к агрегации серпин α1-антитрипсин Z» . ПЛОС ОДИН . 5 (11): е15460. Бибкод : 2010PLoSO...515460G . дои : 10.1371/journal.pone.0015460 . ПМЦ 2980495 . ПМИД 21103396 .
- ^ Кабрита Л.Д., Ирвинг Дж.А., Пирс М.К., Уиссток Дж.К., Боттомли С.П. (сентябрь 2007 г.). «Аэропин из экстремофила Pyrobaculum aerophilum обходит ловушку неправильного сворачивания серпина» . Журнал биологической химии . 282 (37): 26802–26809. дои : 10.1074/jbc.M705020200 . ПМИД 17635906 .
- ^ Флюр Р., Лампл Н., Робертс Т.Х. (май 2012 г.). «Ингибиторы серпин-протеазы в биологии растений». Физиология Плантарум . 145 (1): 95–102. дои : 10.1111/j.1399-3054.2011.01540.x . ПМИД 22085334 .
- ^ Столлер Дж.К., Абуссуан Л.С. (2005). «Дефицит альфа1-антитрипсина». Ланцет . 365 (9478): 2225–2236. дои : 10.1016/S0140-6736(05)66781-5 . ПМИД 15978931 . S2CID 54415934 .
- ^ Мюнх Дж, Стендкер Л, Адерманн К, Шульц А, Шиндлер М, Чиннадурай Р, Полманн С, Чайпан С, Бит Т, Петерс Т, Мейер Б, Вильгельм Д, Лу Х, Цзин В, Цзян С, Форссманн В.Г., Кирхгоф Ф. (апрель 2007 г.). «Открытие и оптимизация природного ингибитора проникновения ВИЧ-1, нацеленного на слитый пептид gp41» . Клетка . 129 (2): 263–275. дои : 10.1016/j.cell.2007.02.042 . ПМИД 17448989 .
- ^ Гоопту Б., Диккенс Дж.А., Ломас Д.А. (февраль 2014 г.). «Молекулярная и клеточная патология дефицита α₁-антитрипсина». Тенденции молекулярной медицины . 20 (2): 116–127. doi : 10.1016/j.molmed.2013.10.007 . ПМИД 24374162 .
- ^ Сейшас С., Суриано Г., Карвальо Ф., Серука Р., Роча Дж., Ди Риенцо А. (февраль 2007 г.). «Разнообразие последовательностей в проксимальном субкластере SERPIN 14q32.1: свидетельства естественного отбора, способствующего псевдогенизации SERPINA2» . Молекулярная биология и эволюция . 24 (2): 587–598. дои : 10.1093/molbev/msl187 . ПМИД 17135331 .
- ^ Калшекер Н.А. (сентябрь 1996 г.). «Альфа 1-антихимотрипсин». Международный журнал биохимии и клеточной биологии . 28 (9): 961–964. дои : 10.1016/1357-2725(96)00032-5 . ПМИД 8930118 . S2CID 11230631 .
- ^ Сантамария М., Пардо-Саганта А., Альварес-Асиаин Л., Ди Скала М., Цянь С., Прието Дж., Авила М.А. (апрель 2013 г.). «Ядерный α1-антхимотрипсин способствует конденсации хроматина и ингибирует пролиферацию клеток гепатоцеллюлярной карциномы человека» . Гастроэнтерология . 144 (4): 818–828.e4. дои : 10.1053/j.gastro.2012.12.029 . ПМИД 23295442 .
- ^ Чжан С., Янчаускене С. (апрель 2002 г.). «Многофункциональные возможности белков: альфа1-антихимотрипсин и корреляция с болезнью Альцгеймера». Журнал болезни Альцгеймера . 4 (2): 115–122. дои : 10.3233/JAD-2002-4206 . ПМИД 12214135 .
- ^ Чао Дж., Сталлоне Дж. Н., Лян Ю. М., Чен Л. М., Ван Д. З., Чао Л. (июль 1997 г.). «Каллистатин — новый мощный сосудорасширяющий препарат» . Журнал клинических исследований . 100 (1): 11–17. дои : 10.1172/JCI119502 . ПМК 508159 . ПМИД 9202051 .
- ^ Мяо RQ, Агата Дж, Чао Л, Чао Дж (ноябрь 2002 г.). «Каллистатин — новый ингибитор ангиогенеза и роста опухолей» . Кровь . 100 (9): 3245–3252. дои : 10.1182/кровь-2002-01-0185 . ПМИД 12384424 .
- ^ Лю Ю, Бледсо Г, Хагивара М, Шен Б, Чао Л, Чао Дж (октябрь 2012 г.). «Истощение эндогенного каллистатина усугубляет почечный и сердечно-сосудистый окислительный стресс, воспаление и ремоделирование органов» . Американский журнал физиологии. Почечная физиология . 303 (8): Ф1230–Ф1238. дои : 10.1152/ajprenal.00257.2012 . ПМЦ 3469672 . ПМИД 22811485 .
- ^ Гейгер М. (март 2007 г.). «Ингибитор белка С, серпин с функциями внутри и вне сосудистой биологии». Тромбоз и гемостаз . 97 (3): 343–347. дои : 10.1160/ч06-09-0488 . ПМИД 17334499 . S2CID 4828322 .
- ^ Баумгертнер П., Гейгер М., Цизенисс С., Маллейер Дж., Хантингтон Дж.А., Хохрайнер К., Билек Э., Стокельхубер М., Лаубер К., Шерфельд Д., Швилле П., Вельделе К., Бейер К., Энгельманн Б. (ноябрь 2007 г.). «Фосфатидилэтаноламин критически поддерживает интернализацию ингибитора проникающего в клетку протеина С» . Журнал клеточной биологии . 179 (4): 793–804. дои : 10.1083/jcb.200707165 . ПМК 2080921 . ПМИД 18025309 .
- ^ Ухрин П., Деверчин М., Хильперт М., Хренек П., Шефер С., Цехмайстер-Маххарт М., Крёнке Г., Валес А., Кармелиет П., Биндер Б.Р., Гейгер М. (декабрь 2000 г.). «Нарушение гена ингибитора протеина С приводит к нарушению сперматогенеза и мужскому бесплодию» . Журнал клинических исследований . 106 (12): 1531–1539. дои : 10.1172/JCI10768 . ПМЦ 381472 . ПМИД 11120760 .
- ^ Хан М.Х., Хван С.И., Рой Д.Б., Лундгрен Д.Х., Прайс Дж.В., Усман С.С., Фернальд Г.Х., Герлиц Б., Робинсон В.Х., Баранзини С.Е., Гриннелл Б.В., Рейн К.С., Собел Р.А., Хан Д.К., Стейнман Л. (февраль 2008 г.). «Протеомный анализ активных поражений рассеянного склероза выявляет терапевтические цели». Природа . 451 (7182): 1076–1081. Бибкод : 2008Natur.451.1076H . дои : 10.1038/nature06559 . ПМИД 18278032 . S2CID 4421395 .
- ^ Torpy DJ, Ho JT (август 2007 г.). «Полиморфизм гена кортикостероид-связывающего глобулина: клинические последствия и связи с идиопатическими расстройствами хронической усталости» . Клиническая эндокринология . 67 (2): 161–167. дои : 10.1111/j.1365-2265.2007.02890.x . ПМИД 17547679 . S2CID 43352358 .
- ^ Барталена Л., Роббинс Дж. (1992). «Вариации белков-переносчиков гормонов щитовидной железы и их клиническое значение». Щитовидная железа . 2 (3): 237–245. дои : 10.1089/thy.1992.2.237 . ПМИД 1422238 .
- ^ Персани Л. (сентябрь 2012 г.). «Клинический обзор: Центральный гипотиреоз: патогенетические, диагностические и терапевтические проблемы» . Журнал клинической эндокринологии и метаболизма . 97 (9): 3068–3078. дои : 10.1210/jc.2012-1616 . ПМИД 22851492 .
- ^ Кумар Р., Вице-президент Сингх, Бейкер К.М. (июль 2007 г.). «Внутриклеточная ренин-ангиотензиновая система: новая парадигма». Тенденции в эндокринологии и обмене веществ . 18 (5): 208–214. дои : 10.1016/j.tem.2007.05.001 . ПМИД 17509892 . S2CID 24041932 .
- ^ Танимото К., Сугияма Ф., Гото Ю., Исида Дж., Такимото Э., Ягами К., Фукамидзу А., Мураками К. (декабрь 1994 г.). «Мыши с дефицитом ангиотензиногена и гипотонией» . Журнал биологической химии . 269 (50): 31334–31337. дои : 10.1016/S0021-9258(18) 31697-1 ПМИД 7989296 .
- ^ Женемайтер Икс, Хименес-Рокепло А.П., Сельерье Ж, Корвол П (1999). «Варианты ангиотензиногена и гипертония человека». Текущие отчеты о гипертонии . 1 (1): 31–41. дои : 10.1007/s11906-999-0071-0 . ПМИД 10981040 . S2CID 42614761 .
- ^ Сети А.А., Нордестгаард Б.Г., Тыбьерг-Хансен А. (июль 2003 г.). «Полиморфизм гена ангиотензиногена, ангиотензиноген плазмы и риск гипертонии и ишемической болезни сердца: метаанализ» . Атеросклероз, тромбоз и сосудистая биология . 23 (7): 1269–1275. дои : 10.1161/01.ATV.0000079007.40884.5C . ПМИД 12805070 .
- ^ Диксон М.Э., Зигмунд CD (июль 2006 г.). «Генетическая основа гипертонии: новый взгляд на ангиотензиноген» . Гипертония . 48 (1): 14–20. дои : 10.1161/01.HYP.0000227932.13687.60 . ПМИД 16754793 .
- ^ Фрейзер Дж.К., Джексон Д.Г., Гайяр Дж.П., Люттер М., Лю Ю.Дж., Баншеро Дж. , Капра Дж.Д., Паскуаль В. (октябрь 2000 г.). «Идентификация центрина: новый серпин, ограниченный В-клетками зародышевого центра человека» . Европейский журнал иммунологии . 30 (10): 3039–3048. doi : 10.1002/1521-4141(200010)30:10<3039::AID-IMMU3039>3.0.CO;2-H . ПМИД 11069088 .
- ^ Патерсон М.А., Хорват А.Дж., Пайк Р.Н., Кофлин П.Б. (август 2007 г.). «Молекулярная характеристика центрина, серпина зародышевой клетки» . Биохимический журнал . 405 (3): 489–494. дои : 10.1042/BJ20070174 . ПМК 2267310 . ПМИД 17447896 .
- ^ Патерсон М.А., Хоскинг П.С., Кофлин П.Б. (июль 2008 г.). «Экспрессия серпинцентрина определяет фенотип зародышевого центра при В-клеточных лимфомах» . Американский журнал клинической патологии . 130 (1): 117–126. дои : 10.1309/9QKE68QU7B825A3U . ПМИД 18550480 .
- ^ Эштон-Рикардт PG (апрель 2013 г.). «Новая роль ингибиторов сериновых протеаз в иммунитете Т-лимфоцитов и за его пределами». Письма по иммунологии . 152 (1): 65–76. дои : 10.1016/j.imlet.2013.04.004 . ПМИД 23624075 .
- ^ Хан X, Филер Р., Брозе Г.Дж. (ноябрь 2000 г.). «Характеристика белка Z-зависимого ингибитора протеазы». Кровь . 96 (9): 3049–3055. дои : 10.1182/blood.V96.9.3049 . ПМИД 11049983 .
- ^ Хида К , Вада Дж , Эгучи Дж , Чжан Х , Баба М , Сейда А , Хасимото И , Окада Т , Ясухара А , Накацука А , Шиката К , Хорай С , Футами Дж , Ватанабэ Е , Мацуки Ю , Хирамацу Р , Акаги С , Макино Х., Канвар Ю.С. (июль 2005 г.). «Ингибитор сериновой протеазы висцеральной жировой ткани: уникальный инсулин-сенсибилизирующий адипоцитокин при ожирении» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (30): 10610–10615. Бибкод : 2005PNAS..10210610H . дои : 10.1073/pnas.0504703102 . ПМЦ 1180799 . ПМИД 16030142 .
- ^ Фэн Р., Ли Ю, Ван С, Луо С, Лю Л, Чуо Ф, Ли Ц, Сунь С (октябрь 2014 г.). «Более высокие уровни васпина у субъектов с ожирением и сахарным диабетом 2 типа: метаанализ». Исследования диабета и клиническая практика . 106 (1): 88–94. дои : 10.1016/j.diabres.2014.07.026 . ПМИД 25151227 .
- ^ Ремолд-О'Доннелл Э., Чин Дж., Альбертс М. (июнь 1992 г.). «Последовательность и молекулярная характеристика ингибитора эластазы моноцитов/нейтрофилов человека» . Труды Национальной академии наук Соединенных Штатов Америки . 89 (12): 5635–5639. Бибкод : 1992PNAS...89.5635R . дои : 10.1073/pnas.89.12.5635 . ПМК 49347 . ПМИД 1376927 .
- ^ Бенарафа С., Прибе Г.П., Ремолд-О'Доннелл Э. (август 2007 г.). «Ингибитор сериновой протеазы нейтрофилов serpinb1 сохраняет защитные функции легких при инфекции Pseudomonas aeruginosa» . Журнал экспериментальной медицины . 204 (8): 1901–1909. дои : 10.1084/jem.20070494 . ПМК 2118684 . ПМИД 17664292 .
- ^ Анталис Т.М., Ла Линн М., Доннан К., Матео Л., Гарднер Дж., Дикинсон Дж.Л., Буттиджиг К., Зурбье А. (июнь 1998 г.). «Ингибитор сериновой протеиназы (серпин) и ингибитор активации плазминогена типа 2 защищает от вирусных цитопатических эффектов посредством конститутивного праймирования интерферона альфа/бета» . Журнал экспериментальной медицины . 187 (11): 1799–1811. дои : 10.1084/jem.187.11.1799 . ПМК 2212304 . ПМИД 9607921 .
- ^ Чжао А., Ян З., Сунь Р., Гринчук В., Нетцель-Арнетт С., Энглин И.Е., Дрисбо К.Х., Нотари Л., Бол Дж.А., Мэдден К.Б., Урбан Дж.Ф., Анталис Т.М., Ши-Донохью Т. (июнь 2013 г.). «SerpinB2 имеет решающее значение для иммунитета Th2 против инфекции кишечных нематод» . Журнал иммунологии . 190 (11): 5779–5787. doi : 10.4049/jimmunol.1200293 . ПМК 4068334 . ПМИД 23630350 .
- ^ Догерти К.М., Пирсон Дж.М., Ян А.Ю., Вестрик Р.Дж., Бейкер М.С., Гинзбург Д. (январь 1999 г.). «Ген ингибитора активатора плазминогена-2 не требуется для нормального развития или выживания мышей» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (2): 686–691. Бибкод : 1999PNAS...96..686D . дои : 10.1073/pnas.96.2.686 . ПМК 15197 . ПМИД 9892694 .
- ^ Такеда А., Ямамото Т., Накамура Ю., Такахаши Т., Хибино Т. (февраль 1995 г.). «Антиген плоскоклеточной карциномы является мощным ингибитором цистеиновой протеиназы катепсина L» . Письма ФЭБС . 359 (1): 78–80. Бибкод : 1995FEBSL.359...78T . дои : 10.1016/0014-5793(94)01456-б . ПМИД 7851535 . S2CID 35146299 .
- ^ Перейти обратно: а б Турато С, Понтиссо П (март 2015 г.). «SERPINB3 (ингибитор серпинпептидазы, клада B (овальбумин), член 3)» . Атлас генетики и цитогенетики в онкологии и гематологии . 19 (3): 202–209. doi : 10.4267/2042/56413 (неактивен 26 июля 2024 г.). ПМЦ 4430857 . ПМИД 25984243 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на июль 2024 г. ( ссылка ) - ^ Перейти обратно: а б Сивапрасад Ю, Аскью DJ, Эриксен М.Б., Гибсон А.М., Стир М.Т., Брандт Э.Б., Басс С.А., Дэйнс М.О., Чакир Дж., Стрингер К.Ф., Верт С.Е., Уитсетт Дж.А., Ле Крас Т.Д., Уиллс-Карп М., Сильверман Г.А., Хурана Херши ГК (январь 2011 г.). «Неизбыточная роль мышиного Serpinb3a в индукции выработки слизи при астме» . Журнал аллергии и клинической иммунологии . 127 (1): 254–61, 261.e1–6. дои : 10.1016/j.jaci.2010.10.009 . ПМК 3058372 . ПМИД 21126757 .
- ^ Шик С., Камачи Ю., Бартуски А.Дж., Каталтепе С., Шехтер Н.М., Пембертон П.А., Сильверман Г.А. (январь 1997 г.). «Антиген 2 плоскоклеточной карциномы представляет собой новый серпин, который ингибирует химотрипсиноподобные протеиназы катепсин G и химазу тучных клеток» . Журнал биологической химии . 272 (3): 1849–1855. дои : 10.1074/jbc.272.3.1849 . ПМИД 8999871 .
- ^ Тео С.С., Уиссток Дж.К., Бёрд П.И. (апрель 2010 г.). «Маспин (SERPINB5) представляет собой облигатный внутриклеточный серпин» . Журнал биологической химии . 285 (14): 10862–10869. дои : 10.1074/jbc.M109.073171 . ПМЦ 2856292 . ПМИД 20123984 .
- ^ Цзоу З, Анисович А, Хендрикс МДж, Тор А, Неве М, Шэн С, Рафиди К, Сектор Э, Сагер Р (январь 1994 г.). «Маспин, серпин, обладающий опухолесупрессирующей активностью в эпителиальных клетках молочной железы человека». Наука . 263 (5146): 526–529. Бибкод : 1994Sci...263..526Z . дои : 10.1126/science.8290962 . ПМИД 8290962 .
- ^ Перейти обратно: а б с Тео С.С., Вьессё Дж., Пракаш М., Беркович С., Луу Дж., Бёрд Ч., Лоу Р.Х., Росадо С., Прайс Дж.Т., Уиссток Дж.К., Берд ПИ (2014). «Маспин не требуется для эмбрионального развития или подавления опухоли» . Природные коммуникации . 5 : 3164. Бибкод : 2014NatCo...5.3164T . дои : 10.1038/ncomms4164 . ПМЦ 3905777 . ПМИД 24445777 .
- ^ Гао Ф., Ши ХИ, Дати С., Селла Н., Чжан М. (апрель 2004 г.). «Маспин играет важную роль в раннем эмбриональном развитии» . Разработка . 131 (7): 1479–1489. дои : 10.1242/dev.01048 . ПМИД 14985257 .
- ^ Скотт Флорида, Херст CE, Сан Дж, Берд CH, Боттомли SP, Берд PI (март 1999 г.). «Внутриклеточный ингибитор серпинпротеиназы 6 экспрессируется в моноцитах и гранулоцитах и является мощным ингибитором азурофильной гранулярной протеазы, катепсина G». Кровь . 93 (6): 2089–2097. дои : 10.1182/blood.V93.6.2089.406k10_2089_2097 . ПМИД 10068683 .
- ^ Тан Дж., Пракаш, доктор медицинских наук, Кайзерман Д., Берд П.И. (июль 2013 г.). «Отсутствие SERPINB6A вызывает нейросенсорную тугоухость с множественными гистопатологиями во внутреннем ухе мыши». Американский журнал патологии . 183 (1): 49–59. дои : 10.1016/j.ajpath.2013.03.009 . ПМИД 23669344 .
- ^ Скарфф К.Л., Унг К.С., Нандуркар Х., Крэк П.Дж., Берд CH, Берд ПИ (май 2004 г.). «Целевое нарушение SPI3/Serpinb6 не приводит к дефектам развития или роста, дисфункции лейкоцитов или предрасположенности к инсульту» . Молекулярная и клеточная биология . 24 (9): 4075–4082. дои : 10.1128/MCB.24.9.4075-4082.2004 . ПМЦ 387772 . ПМИД 15082799 .
- ^ Сирмаджи А, Эрбек С, Прайс Дж, Хуанг М, Думан Д, Ченгиз ФБ, Бадемчи Г, Токгёз-Йилмаз С, Хишми Б, Оздаг Х, Озтюрк Б, Кулаксизоглу С, Йылдирим Е, Кокотас Х, Григориаду М, Петерсен МБ, Шахин Х, Канаан М, Кинг МС, Чен Зи, Блэнтон С.Х., Лю XZ, Цухнер С., Акар Н., Текин М. (май 2010 г.). «Усекательная мутация в SERPINB6 связана с аутосомно-рецессивной несиндромальной сенсоневральной тугоухостью» . Американский журнал генетики человека . 86 (5): 797–804. дои : 10.1016/j.ajhg.2010.04.004 . ПМК 2869020 . ПМИД 20451170 .
- ^ Мията Т., Инаги Р., Нангаку М., Имасава Т., Сато М., Изухара Ю., Судзуки Д., Ёсино А., Оноги Х., Кимура М., Сугияма С., Курокава К. (март 2002 г.). «Сверхэкспрессия серпин-мегсина индуцирует прогрессирующую пролиферацию и расширение мезангиальных клеток» . Журнал клинических исследований . 109 (5): 585–593. дои : 10.1172/JCI14336 . ПМК 150894 . ПМИД 11877466 .
- ^ Перейти обратно: а б Miyata T, Li M, Yu X, Hirayama N (май 2007). «Ген мегсина: его геномный анализ, патобиологические функции и терапевтические перспективы» . Текущая геномика . 8 (3): 203–208. doi : 10.2174/138920207780833856 . PMC 2435355 . PMID 18645605 .
- ^ Кубо А (август 2014 г.). «Пальмоплантар-кератоз типа Нагасима: распространенный азиатский тип, вызванный дефицитом ингибитора протеазы Serpinb7» . Журнал следственной дерматологии . 134 (8): 2076–2079. doi : 10.1038/jid.2014.156 . PMID 25029323 .
- ^ Дален-младший, Жан Ф., Томас Дж., Фостер Д.С., Кисиэль В. (январь 1998 г.). «Ингибирование растворимого рекомбинантного фурина ингибитором протеиназы человека 8» . Журнал биологической химии . 273 (4): 1851–1854. дои : 10.1074/jbc.273.4.1851 . ПМИД 9442015 .
- ^ Сан Дж., Берд CH, Саттон В., Макдональд Л., Кофлин П.Б., Де Йонг Т.А., Трапани Дж.А., Берд ПИ (ноябрь 1996 г.). «Ингибитор цитозольного гранзима B, родственный вирусному регулятору апоптоза, модификатору цитокинового ответа A, присутствует в цитотоксических лимфоцитах» . Журнал биологической химии . 271 (44): 27802–27809. дои : 10.1074/jbc.271.44.27802 . ПМИД 8910377 .
- ^ Чжан М., Пак С.М., Ван Ю, Шах Р., Лю Н., Мурманн А.Е., Ван Ч.Р., Питер М.Э., Эштон-Рикардт П.Г. (апрель 2006 г.). «Ингибитор сериновой протеазы 6 защищает цитотоксические Т-клетки от самостоятельного повреждения, обеспечивая целостность цитотоксических гранул» . Иммунитет . 24 (4): 451–461. doi : 10.1016/j.immuni.2006.02.002 . ПМИД 16618603 .
- ^ Риццителли А., Меутер С., Вега Рамос Дж., Берд CH, Минтерн Дж.Д., Манган М.С., Вилладангос Дж., Берд ПИ (октябрь 2012 г.). «У мышей с дефицитом Serpinb9 (Spi6) нарушена перекрестная презентация антигена, опосредованная дендритными клетками». Иммунология и клеточная биология . 90 (9): 841–851. дои : 10.1038/icb.2012.29 . ПМИД 22801574 . S2CID 39276036 .
- ^ Ривальд М., Чуанг Т., Нойбауэр А., Рисс Х., Шлиф Р.Р. (февраль 1998 г.). «Экспрессия бомапина, нового человеческого серпина, в нормальном/злокачественном кроветворении и в моноцитарных клеточных линиях THP-1 и AML-193» . Кровь . 91 (4): 1256–1262. дои : 10.1182/blood.V91.4.1256 . ПМИД 9454755 .
- ^ Перейти обратно: а б Аскью Д.Д., Каталтепе С., Кумар В., Эдвардс К., Пейс С.М., Ховарт Р.Н., Пак СК, Аскью Ю.С., Бромме Д., Люк С.Дж., Уиссток Дж.К., Сильверман Г.А. (август 2007 г.). «SERPINB11 — это новый неингибирующий внутриклеточный серпин. Обычные однонуклеотидные полиморфизмы в каркасе нарушают конформационные изменения» . Журнал биологической химии . 282 (34): 24948–24960. дои : 10.1074/jbc.M703182200 . ПМИД 17562709 .
- ^ Финно С.Дж., Стивенс С., Янг А., Аффолтер В., Джоши Н.А., Рамзи С., Баннаш Д.Л. (апрель 2015 г.). «Вариант сдвига рамки SERPINB11, связанный с новым специфическим фенотипом копыт у пони Коннемара» . ПЛОС Генетика . 11 (4): e1005122. дои : 10.1371/journal.pgen.1005122 . ПМЦ 4395385 . ПМИД 25875171 .
- ^ Аскью Ю.С., Пак С.С., Люк С.Дж., Аскью Д.Д., Каталтепе С., Миллс Д.Р., Като Х., Лехочки Дж., Дьюар К., Биррен Б., Сильверман Г.А. (декабрь 2001 г.). «SERPINB12 — новый член человеческого семейства ов-серпинов, который широко экспрессируется и ингибирует трипсиноподобные сериновые протеиназы» . Журнал биологической химии . 276 (52): 49320–49330. дои : 10.1074/jbc.M108879200 . ПМИД 11604408 .
- ^ Уэлсс Т., Сан Дж., Ирвинг Дж.А., Блюм Р., Смит А.И., Уиссток Дж.К., Пайк Р.Н., фон Микеч А., Ружичка Т., Берд П.И., Абтс Х.Ф. (июнь 2003 г.). «Гурпин является селективным ингибитором лизосомального катепсина L и защищает кератиноциты от апоптоза, индуцированного ультрафиолетом». Биохимия . 42 (24): 7381–7389. дои : 10.1021/bi027307q . ПМИД 12809493 .
- ^ Исигуро К., Кодзима Т., Кадомацу К., Накаяма Ю., Такаги А., Сузуки М., Такеда Н., Ито М., Ямамото К., Мацусита Т., Кусугами К., Мурамацу Т., Сайто Х. (октябрь 2000 г.). «Полный дефицит антитромбина у мышей приводит к эмбриональной смертности» . Журнал клинических исследований . 106 (7): 873–878. дои : 10.1172/JCI10489 . ПМК 517819 . ПМИД 11018075 .
- ^ Патнаик М.М., Молл С. (ноябрь 2008 г.). «Наследственный дефицит антитромбина: обзор» . Гемофилия . 14 (6): 1229–1239. дои : 10.1111/j.1365-2516.2008.01830.x . ПМИД 19141163 . S2CID 20768425 .
- ^ Хантингтон Дж. А. (июль 2011 г.). «Структура, функции и дисфункция серпинов» . Журнал тромбозов и гемостаза . 9 (Приложение 1): 26–34. дои : 10.1111/j.1538-7836.2011.04360.x . ПМИД 21781239 . S2CID 1020630 .
- ^ Висенте К.П., Хе Л., Павао М.С., Толлефсен Д.М. (декабрь 2004 г.). «Антитромботическая активность дерматансульфата у мышей с дефицитом кофактора гепарина II» . Кровь . 104 (13): 3965–3970. дои : 10.1182/кровь-2004-02-0598 . ПМИД 15315969 .
- ^ Аихара К , Адзума Х , Акаике М , Икеда Й , Сата М , Такамори Н , Яги С , Ивасе Т , Сумитомо Й , Кавано Х , Ямада Т , Фукуда Т , Мацумото Т , Сэкине К , Сато Т , Накамичи Й , Ямамото Й , Ёсимура К , Ватанабэ Т , Накамура Т , Оомидзу А , Цукада М , Хаяши Х , Судо Т., Като С., Мацумото Т. (июнь 2007 г.). «Зависимая от штамма эмбриональная летальность и чрезмерное ремоделирование сосудов у мышей с дефицитом кофактора гепарина II» . Журнал клинических исследований . 117 (6): 1514–1526. дои : 10.1172/JCI27095 . ПМК 1878511 . ПМИД 17549254 .
- ^ Кейл Дж. М., Лоуренс Д. А. (сентябрь 2007 г.). «Структурно-функциональные взаимосвязи ингибитора активатора плазминогена-1 и его потенциал как терапевтического агента». Текущие цели по борьбе с наркотиками . 8 (9): 971–981. дои : 10.2174/138945007781662337 . ПМИД 17896949 .
- ^ Лино М.М., Атанасоски С., Квайо М., Файард Б., Морено Э., Бреннер Х.Р., Сутер У., Монард Д. (апрель 2007 г.). «У мышей, у которых отсутствует протеаза нексин-1, наблюдается задержка структурного и функционального восстановления после повреждения седалищного нерва» . Журнал неврологии . 27 (14): 3677–3685. doi : 10.1523/JNEUROSCI.0277-07.2007 . ПМК 6672422 . ПМИД 17409231 .
- ^ Мурер В., Спец Дж. Ф., Хенгст У., Альтрогге Л. М., де Агостини А., Монар Д. (март 2001 г.). «Дефекты мужской фертильности у мышей, у которых отсутствует ингибитор сериновой протеазы протеаза нексин-1» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (6): 3029–3033. Бибкод : 2001PNAS...98.3029M . дои : 10.1073/pnas.051630698 . ПМК 30601 . ПМИД 11248026 .
- ^ Люти А., Ван дер Путтен Х., Боттери Ф.М., Мансуи И.М., Майнс М., Фрей У., Сансиг Г., Портет С., Шмутц М., Шредер М., Нитч С., Лоран Дж. П., Монар Д. (июнь 1997 г.). «Эндогенный ингибитор сериновой протеазы модулирует эпилептическую активность и долговременное потенцирование гиппокампа» . Журнал неврологии . 17 (12): 4688–4699. doi : 10.1523/JNEUROSCI.17-12-04688.1997 . ПМК 6573330 . ПМИД 9169529 .
- ^ Перейти обратно: а б Долл Дж.А., Штельмах В.М., Бук Н.П., Берг А.Р., Ли С., Абрамсон Л.П., Корнуэлл М.Л., Пинс М.Р., Боренцтайн Дж., Кроуфорд С.Е. (июнь 2003 г.). «Фактор пигментного эпителия регулирует сосудистую сеть и массу предстательной железы и поджелудочной железы». Природная медицина . 9 (6): 774–780. дои : 10.1038/nm870 . ПМИД 12740569 . S2CID 5967666 .
- ^ Бесерра С.П., Перес-Медиавилла Л.А., Уэлдон Дж.Э., Локателли-Хупс С., Сенанаяке П., Нотари Л., Нотарио В., Холлифилд Дж.Г. (ноябрь 2008 г.). «Фактор пигментного эпителия связывается с гиалуронаном. Картирование сайта связывания гиалуронана» . Журнал биологической химии . 283 (48): 33310–33320. дои : 10.1074/jbc.M801287200 . ПМК 2586245 . ПМИД 18805795 .
- ^ Андреу-Агулло К., Моранте-Редолат Х.М., Дельгадо АС, Фариньяс I (декабрь 2009 г.). «Фактор сосудистой ниши PEDF модулирует Notch-зависимую стволовость в субэпендимальной зоне взрослых». Природная неврология . 12 (12): 1514–1523. дои : 10.1038/nn.2437 . ПМИД 19898467 . S2CID 5332822 .
- ^ Виман Б., Коллен Д. (сентябрь 1979 г.). «О механизме реакции альфа-2-антиплазмина человека с плазмином» . Журнал биологической химии . 254 (18): 9291–9297. дои : 10.1016/S0021-9258(19)86843-6 . ПМИД 158022 .
- ^ Лийнен Х.Р., Окада К., Мацуо О., Коллен Д., Деверчин М. (апрель 1999 г.). «Дефицит гена альфа2-антиплазмина у мышей связан с повышенным фибринолитическим потенциалом без явных кровотечений». Кровь . 93 (7): 2274–2281. дои : 10.1182/blood.V93.7.2274 . ПМИД 10090937 .
- ^ Карпентер С.Л., Мэтью П. (ноябрь 2008 г.). «Альфа2-антиплазмин и его дефицит: нарушение баланса фибринолиза» . Гемофилия . 14 (6): 1250–1254. дои : 10.1111/j.1365-2516.2008.01766.x . ПМИД 19141165 . S2CID 205295156 .
- ^ Фавье Р., Аоки Н., де Мурлуз П. (июль 2001 г.). «Врожденный дефицит ингибиторов альфа(2)-плазмина: обзор» . Британский журнал гематологии . 114 (1): 4–10. дои : 10.1046/j.1365-2141.2001.02845.x . ПМИД 11472338 . S2CID 71010865 .
- ^ Бейнрор Л., Хармат В., Добо Дж., Лоринц З., Галь П., Заводски П. (июль 2007 г.). «Структура домена серпина ингибитора C1 раскрывает вероятный механизм потенцирования гепарина и конформационного заболевания» . Журнал биологической химии . 282 (29): 21100–21109. дои : 10.1074/jbc.M700841200 . hdl : 10831/92835 . ПМИД 17488724 .
- ^ Моллнес Т.Э., Джокиранта Т.С., Трудссон Л., Нильссон Б., Родригес де Кордоба С., Киршфинк М. (сентябрь 2007 г.). «Анализ комплемента в 21 веке». Молекулярная иммунология . 44 (16): 3838–3849. дои : 10.1016/j.molimm.2007.06.150 . hdl : 10261/61732 . ПМИД 17768101 .
- ^ Триджианезе П., Чименти М.С., Туби Э., Балланти Э., Гуарино М.Д., Перриконе С., Перриконе Р. (август 2015 г.). «Аутоиммунная сторона наследственного ангионевротического отека: взгляд на патогенез». Обзоры аутоиммунитета . 14 (8): 665–669. дои : 10.1016/j.autrev.2015.03.006 . ПМИД 25827463 .
- ^ Нагай Н., Хосокава М., Итохара С., Адачи Э., Мацусита Т., Хосокава Н., Нагата К. (сентябрь 2000 г.). «Эмбриональная летальность мышей с нокаутом молекулярного шаперона hsp47 связана с дефектами биосинтеза коллагена» . Журнал клеточной биологии . 150 (6): 1499–1506. дои : 10.1083/jcb.150.6.1499 . ПМК 2150697 . ПМИД 10995453 .
- ^ Марини Дж. К., Райх А., Смит С. М. (август 2014 г.). «Несовершенный остеогенез вследствие мутаций неколлагеновых генов: уроки биологии костеобразования» . Современное мнение в педиатрии . 26 (4): 500–507. дои : 10.1097/MOP.0000000000000117 . ПМЦ 4183132 . ПМИД 25007323 .
- ^ Байерс П.Х., Пайотт С.М. (1 января 2012 г.). «Рецессивно наследуемые формы несовершенного остеогенеза». Ежегодный обзор генетики . 46 : 475–497. doi : 10.1146/annurev-genet-110711-155608 . ПМИД 23145505 .
- ^ Остервальдер Т., Чинелли П., Байчи А., Пеннелла А., Крюгер С.Р., Шримпф С.П., Майнс М., Зондереггер П. (январь 1998 г.). «Ингибитор сериновых протеиназ, секретируемый аксонами, нейросерпин, ингибирует активаторы плазминогена и плазмин, но не тромбин» . Журнал биологической химии . 273 (4): 2312–2321. дои : 10.1074/jbc.273.4.2312 . ПМИД 9442076 .
- ^ Кроутер, округ Колумбия (июль 2002 г.). «Семейные конформационные заболевания и деменции» . Человеческая мутация . 20 (1): 1–14. дои : 10.1002/humu.10100 . ПМИД 12112652 . S2CID 22326349 .
- ^ Белорджей Д., Хэгглоф П., Карлссон-Ли С., Ломас Д.А. (1 марта 2007 г.). «Неправильное сворачивание белков и серпинопатии» . Прион . 1 (1): 15–20. дои : 10.4161/при.1.1.3974 . ПМЦ 2633702 . ПМИД 19164889 .
- ^ Одзаки К., Нагата М., Сузуки М., Фудзивара Т., Миёси Ю., Исикава О., Охигаси Х., Имаока С., Такахаши Э., Накамура Ю. (июль 1998 г.). «Выделение и характеристика нового гена, специфичного для поджелудочной железы человека, панпина, активность которого снижается в клетках рака поджелудочной железы». Гены, хромосомы и рак . 22 (3): 179–185. doi : 10.1002/(SICI)1098-2264(199807)22:3<179::AID-GCC3>3.0.CO;2-T . ПМИД 9624529 . S2CID 9525421 .
- ^ Лофтус С.К., Кэннонс Дж.Л., Инкао А., Пак Е., Чен А., Зерфас П.М., Брайант М.А., Бизекер Л.Г., Шварцберг П.Л., Паван У.Дж. (сентябрь 2005 г.). «Апоптоз ацинарных клеток у мышей с дефицитом Serpini2 моделирует недостаточность поджелудочной железы» . ПЛОС Генетика . 1 (3): е38. дои : 10.1371/journal.pgen.0010038 . ПМЦ 1231717 . ПМИД 16184191 .
- ^ Падуя М.Б., Ковальски А.А., Каньяс М.Ю., Хансен П.Дж. (февраль 2010 г.). «Молекулярная филогения маточных серпинов и ее связь с эволюцией плаценты» . Журнал ФАСЭБ . 24 (2): 526–537. дои : 10.1096/fj.09-138453 . ПМИД 19825977 . S2CID 9248169 .
- ^ Падуя МБ, Хансен П.Дж. (октябрь 2010 г.). «Эволюция и функция серпинов матки (SERPINA14)» . Американский журнал репродуктивной иммунологии . 64 (4): 265–274. дои : 10.1111/j.1600-0897.2010.00901.x . ПМИД 20678169 .
- ^ Перейти обратно: а б Райххарт Дж. М. (декабрь 2005 г.). «Вершина еще одного айсберга: змеи дрозофилы». Тенденции в клеточной биологии . 15 (12): 659–665. дои : 10.1016/j.tcb.2005.10.001 . ПМИД 16260136 .
- ^ Тан Х, Камбрис З, Леметр Б, Хашимото С (октябрь 2008 г.). «Серпин, регулирующий иммунную меланизацию в дыхательной системе дрозофилы» . Развивающая клетка . 15 (4): 617–626. дои : 10.1016/j.devcel.2008.08.017 . ПМЦ 2671232 . ПМИД 18854145 .
- ^ Шерфер С., Тан Х., Камбрис З., Лхосин Н., Хашимото С., Леметр Б. (ноябрь 2008 г.). «Дрозофила Serpin-28D регулирует активность фенолоксидазы гемолимфы и пигментацию взрослых» . Биология развития . 323 (2): 189–196. дои : 10.1016/j.ydbio.2008.08.030 . ПМИД 18801354 .
- ^ Рашлоу С. (январь 2004 г.). «Дорсовентральный паттерн: наконец-то прижатая змея» . Современная биология . 14 (1): С16–С18. Бибкод : 2004CBio...14..R16R . дои : 10.1016/j.cub.2003.12.015 . ПМИД 14711428 .
- ^ Лигоксигакис П., Рот С., Райххарт Дж.М. (декабрь 2003 г.). «Серпин регулирует формирование дорсально-вентральной оси у эмбриона дрозофилы» . Современная биология . 13 (23): 2097–2102. Бибкод : 2003CBio...13.2097L . дои : 10.1016/j.cub.2003.10.062 . ПМИД 14654000 .
- ^ Цзян Р., Чжан Б., Курокава К., Со Йи, Ким Э.Х., Хван Хо, Ли Дж.Х., Ширацучи А., Чжан Дж., Наканиси Ю., Ли Х.С., Ли Б.Л. (октябрь 2011 г.). «Двухдоменный ингибитор сериновой протеазы 93 кДа (серпин) выполняет регуляторную функцию в протеолитическом сигнальном каскаде Toll жуков» . Журнал биологической химии . 286 (40): 35087–35095. дои : 10.1074/jbc.M111.277343 . ПМК 3186399 . ПМИД 21862574 .
- ^ Перейти обратно: а б Пак С.К., Кумар В., Цу С., Люк СиДжей, Аскью Ю.С., Аскью Д.Д., Миллс Д.Р., Бромме Д., Сильверман Г.А. (апрель 2004 г.). «SRP-2 представляет собой межклассовый ингибитор, который участвует в постэмбриональном развитии нематоды Caenorhabditis elegans: первоначальная характеристика клады L serpins» . Журнал биологической химии . 279 (15): 15448–15459. дои : 10.1074/jbc.M400261200 . ПМИД 14739286 .
- ^ Люк С.Дж., Пак С.С., Аскью Ю.С., Навилья Т.Л., Аскью DJ, Нобар С.М., Ветика AC, Лонг ОС, Уоткинс С.С., Штольц Д.Б., Барстед Р.Дж., Молдер Г.Л., Бремме Д., Сильверман Г.А. (сентябрь 2007 г.). «Внутриклеточный серпин регулирует некроз, ингибируя индукцию и последствия лизосомального повреждения» . Клетка . 130 (6): 1108–1119. дои : 10.1016/j.cell.2007.07.013 . ПМК 2128786 . ПМИД 17889653 .
- ^ Хейгаард Дж., Расмуссен С.К., Брандт А., Свендсени (1985). «Гомология последовательностей между белком Z эндосперма ячменя и ингибиторами протеазы семейства альфа-1-антитрипсина» . ФЭБС Летт . 180 (1): 89–94. Бибкод : 1985FEBSL.180...89H . дои : 10.1016/0014-5793(85)80238-6 . S2CID 84790014 .
- ^ Даль С.В., Расмуссен С.К., Петерсен Л.К., Хейгаард Дж. (сентябрь 1996 г.). «Ингибирование факторов свертывания крови рекомбинантным серпином ячменя BSZx» . Письма ФЭБС . 394 (2): 165–168. Бибкод : 1996FEBSL.394..165D . дои : 10.1016/0014-5793(96)00940-4 . ПМИД 8843156 .
- ^ Хейгаард Дж. (январь 2001 г.). «Ингибирующие серпины из зерна ржи с глютамином в виде остатков Р1 и Р2 в реактивном центре» . Письма ФЭБС . 488 (3): 149–153. Бибкод : 2001FEBSL.488..149H . дои : 10.1016/S0014-5793(00)02425-X . ПМИД 11163762 . S2CID 27933086 .
- ^ Остергаард Х., Расмуссен С.К., Робертс Т.Х., Хейгаард Дж. (октябрь 2000 г.). «Ингибирующие серпины из зерна пшеницы с реактивными центрами, напоминающими богатые глутамином повторы белков-хранилищ проламина. Клонирование и характеристика пяти основных молекулярных форм» . Журнал биологической химии . 275 (43): 33272–33279. дои : 10.1074/jbc.M004633200 . ПМИД 10874043 .
- ^ Перейти обратно: а б Ю Б.К., Аоки К., Сян Ю., Кэмпбелл Л.Р., Халл Р.Дж., Ксоконостл-Казарес Б., Монцер Дж., Ли Дж.Ю., Ульман Д.Э., Лукас У.Дж. (ноябрь 2000 г.). «Характеристика серпина-1 флоэмы тыквы maxima (CmPS-1). Ингибитор эластазы, регулируемый развитием» . Журнал биологической химии . 275 (45): 35122–35128. дои : 10.1074/jbc.M006060200 . ПМИД 10960478 .
- ^ ла Кур Петерсен М., Хейгаард Дж., Томпсон Г.А., Шульц А. (декабрь 2005 г.). «Зерпины флоэмы тыквы передаются прививкой и, по-видимому, устойчивы к обновлению в комплексе ситовидный элемент-клетка-компаньон» . Журнал экспериментальной ботаники . 56 (422): 3111–3120. дои : 10.1093/jxb/eri308 . ПМИД 16246856 .
- ^ Робертс Т.Х., Хейгаард Дж. (февраль 2008 г.). «Зерпины у растений и зеленых водорослей». Функциональная и интегративная геномика . 8 (1): 1–27. дои : 10.1007/s10142-007-0059-2 . ПМИД 18060440 . S2CID 22960858 .
- ^ Лампл Н., Алкан Н., Давыдов О., Флюр Р. (май 2013 г.). «Контроль заданного значения активности протеазы RD21 с помощью AtSerpin1 контролирует гибель клеток арабидопсиса» . Заводской журнал . 74 (3): 498–510. дои : 10.1111/tpj.12141 . ПМИД 23398119 .
- ^ Ан Дж.В., Этвелл Б.Дж., Робертс Т.Х. (май 2009 г.). «Гены серпина AtSRP2 и AtSRP3 необходимы для нормальной чувствительности роста к агенту, алкилирующему ДНК, у арабидопсиса» . Биология растений BMC . 9:52 . дои : 10.1186/1471-2229-9-52 . ПМЦ 2689219 . ПМИД 19426562 .
- ^ Перейти обратно: а б Канг С., Барак Ю., Ламед Р., Байер Э.А., Моррисон М. (июнь 2006 г.). «Функциональный репертуар прокариотных целлюлосом включает суперсемейство серпинов ингибиторов сериновых протеиназ» . Молекулярная микробиология . 60 (6): 1344–1354. дои : 10.1111/j.1365-2958.2006.05182.x . ПМИД 16796673 . S2CID 23738804 .
- ^ Ирвинг Дж.А., Кабрита Л.Д., Россджон Дж., Пайк Р.Н., Боттомли С.П., Уиссток Дж.К. (апрель 2003 г.). «Кристаллическая структура 1,5 А серпина прокариота: контроль конформационных изменений в нагретой среде» . Структура . 11 (4): 387–397. дои : 10.1016/S0969-2126(03)00057-1 . ПМИД 12679017 .
- ^ Фултон К.Ф., Бакл А.М., Кабрита Л.Д., Ирвинг Дж.А., Батчер Р.Э., Смит И., Рив С., Леск А.М., Боттомли С.П., Россджон Дж., Уиссток Дж.К. (март 2005 г.). «Кристаллическая структура природного термостабильного серпина с высоким разрешением раскрывает сложный механизм, лежащий в основе перехода от напряжения к расслаблению» . Журнал биологической химии . 280 (9): 8435–8442. дои : 10.1074/jbc.M410206200 . ПМИД 15590653 .
- ^ Тернер ПК, Мойер Р.В. (сентябрь 2002 г.). «Имуномодуляторы поксвируса: функциональные данные на моделях животных». Вирусные исследования . 88 (1–2): 35–53. дои : 10.1016/S0168-1702(02)00119-3 . ПМИД 12297326 .
- ^ Перейти обратно: а б Ричардсон Дж., Вишванатан К., Лукас А. (январь 2006 г.). «Серпины, сосудистая сеть и вирусная терапия» . Границы бионауки . 11 : 1042–1056. дои : 10.2741/1862 . ПМИД 16146796 .
- ^ Цзян Дж., Арп Дж., Кубелик Д., Зассоко Р., Лю В., Уайз Ю., Маколей С., Гарсия Б., Макфадден Г., Лукас А.Р., Ван Х. (ноябрь 2007 г.). «Индукция неопределенной выживаемости сердечного аллотрансплантата коррелирует со снижением регуляции toll-подобных рецепторов 2 и 4 после лечения ингибитором сериновой протеазы-1 (Serp-1)» . Трансплантация . 84 (9): 1158–1167. дои : 10.1097/01.tp.0000286099.50532.b0 . ПМИД 17998872 . S2CID 20168458 .
- ^ Дай Э., Гуань Х., Лю Л., Литтл С., Макфадден Г., Вазири С., Цао Х., Иванова И.А., Бокш Л., Лукас А. (май 2003 г.). «Serp-1, вирусный противовоспалительный серпин, регулирует клеточную сериновую протеиназу и реакцию серпина на повреждение сосудов» . Журнал биологической химии . 278 (20): 18563–18572. дои : 10.1074/jbc.M209683200 . ПМИД 12637546 .
- ^ Тернер ПК, Санчо MC, Тоеннес С.Р., Капуто А., Бликли Р.К., Мойер Р.В. (август 1999 г.). «Вирус миксомы Serp2 является слабым ингибитором гранзима B и фермента, превращающего интерлейкин-1бета, in vitro и, в отличие от CrmA, не может блокировать апоптоз в клетках, инфицированных вирусом коровьей оспы» . Журнал вирусологии . 73 (8): 6394–6404. doi : 10.1128/JVI.73.8.6394-6404.1999 . ПМЦ 112719 . ПМИД 10400732 .
- ^ Мунусвами-Рамануджам Г., Хан К.А., Лукас А.Р. (декабрь 2006 г.). «Вирусные противовоспалительные реагенты: возможности лечения артритов и васкулитов». Целевые препараты для лечения эндокринных, метаболических и иммунных расстройств . 6 (4): 331–343. дои : 10.2174/187153006779025720 . ПМИД 17214579 .
- ^ Ренатус М., Чжоу К., Стеннике Х.Р., Снипас С.Дж., Терк Д., Бэнкстон Л.А., Лиддингтон Р.К., Салвесен Г.С. (июль 2000 г.). «Кристаллическая структура супрессора апоптоза CrmA в расщепленной форме» . Структура . 8 (7): 789–797. дои : 10.1016/S0969-2126(00)00165-9 . ПМИД 10903953 .
Внешние ссылки
[ редактировать ]- PDB Молекула месяца Серпин
- Хромота ингибитора протеазы Merops (семейство I4). Архивировано 8 декабря 2016 г. в Wayback Machine.
- Серпинс в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Лаборатория Джеймса Уисстока в Университете Монаша
- Лаборатория Джима Хантингтона. Архивировано 30 октября 2016 года в Wayback Machine в Кембриджском университете.
- Лаборатория Фрэнка Черча в Университете Северной Каролины в Чапел-Хилл
- Лаборатория Пола Деклера в Католическом университете Левена
- Лаборатория Тома Робертса в Сиднейском университете
- Лаборатория Роберта Флюра в Институте науки Вейцмана
- Лаборатория Питера Геттинса в Университете Иллинойса в Чикаго
- Обзор всей структурной информации, доступной в PDB для UniProt : P01009 (Человеческий альфа-1-антитрипсин) в PDBe-KB .