К-комплекс
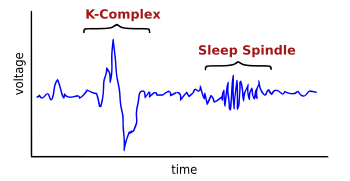
К -комплекс — это волна, которую можно увидеть на электроэнцефалограмме (ЭЭГ) . Это происходит во время 2-й стадии медленного сна . Это «крупнейшее событие в ЭЭГ здорового человека». [ 1 ] Они чаще встречаются в первых циклах сна.
К-комплексы имеют две предполагаемые функции: [ 1 ] во сне во-первых, подавление коркового возбуждения в ответ на стимулы, которые спящий мозг оценивает как не сигнализирующие об опасности, и, во-вторых, содействие консолидации памяти .
К-комплекс был открыт в 1937 году в частных лабораториях Альфреда Ли Лумиса . [ 2 ]
Нейрофизиология
[ редактировать ]К-комплекс состоит из короткого отрицательного пика высокого напряжения, обычно превышающего 100 мкВ, за которым следует более медленный положительный комплекс около 350 и 550 мс и последний отрицательный пик на 900 мс. К-комплексы возникают примерно каждые 1,0–1,7 минуты и часто сопровождаются вспышками веретена сна . Они возникают спонтанно [ 1 ] но также возникают в ответ на внешние раздражители, такие как звуки и прикосновения к коже. [ 3 ] и внутренние, такие как перерывы на вдохе. [ 4 ] Они генерируются в обширных корковых местах. [ 1 ] хотя они имеют тенденцию преобладать над лобными частями мозга. [ 5 ]
Активность как К-комплекса, так и дельта-волн во 2 стадии сна создает медленноволновые (0,8 Гц) и дельта-колебания (1,6–4,0 Гц). Однако их топографическое распределение различно, а дельта-мощность К-комплексов выше. [ 6 ]
Они создаются за счет возникновения в обширных областях коры наружных дендритных токов от среднего (III) к верхним (I) слоям коры головного мозга . Это сопровождается снижением мощности широкополосной ЭЭГ, в том числе активности гамма-волн . Это приводит к «неактивным состояниям» нейронного молчания, в которых активность нейронной сети снижается. [ 1 ] Активность К-комплексов передается в таламус , где синхронизирует таламокортикальную сеть во время сна, создавая колебания сна, такие как веретена и дельта-волны . [ 7 ] Было замечено, что они действительно идентичны по «ламинарному распределению трансмембранных токов» медленным волнам медленноволнового сна . [ 1 ]
Было предложено использовать К-комплексы как для защиты сна, так и для участия в обработке информации, поскольку они оба являются важной частью синхронизации медленного сна, а также реактивно реагируют как на внутренние, так и на внешние раздражители. [ 8 ] Это согласуется с функцией подавления коркового возбуждения в ответ на стимулы, которые мозгу необходимо первоначально обработать, чтобы определить, опасно это или нет. [ 1 ]
Другая предполагаемая функция — содействие активационному гомеостазу синапсов . [ 9 ] и консолидация памяти. Пороги активации корковых синапсов снижаются во время бодрствования, поскольку они обрабатывают информацию, что делает их более отзывчивыми, и поэтому их необходимо отрегулировать обратно, чтобы сохранить соотношение сигнал/шум. [ 9 ] Состояние подавленности, обеспечиваемое К-комплексами, достигается за счет уменьшения силы синаптических связей, которые возникают, когда человек бодрствует. [ 1 ] Кроме того, выход из состояния упадка, которое они вызывают, позволяет «перезапускаться» кортикальной активности в систематическом порядке», так что энграммы памяти, закодированные во время активации нейронов, можно «многократно практиковать и, таким образом, консолидировать». [ 1 ]
Разработка
[ редактировать ]Они присутствуют во сне 5-месячных детей и развиваются с возрастом. В возрасте от 3 до 5 лет негативный компонент проявляется быстрее и продолжает нарастать до подросткового возраста . Другое изменение происходит у взрослых: до 30 лет их частота и амплитуда выше, чем у пожилых людей, особенно старше 50 лет. [ 10 ] Это происходит параллельно с уменьшением других компонентов сна, таких как плотность веретена сна и дельта-мощность . [ 10 ]
Клинический
[ редактировать ]Эпилепсия
[ редактировать ]У людей с идиопатической генерализованной эпилепсией , вызванная К-комплексом, синхронизация может вызывать спайк-волновые разряды. Обычно это происходит в период между бодрствованием и медленным сном, а также между медленным и быстрым сном . [ 11 ] При аутосомно-доминантной ночной лобной эпилепсии К-комплексы почти всегда присутствуют в начале приступов. [ 12 ]
Синдром беспокойных ног
[ редактировать ]У людей с синдромом беспокойных ног увеличено количество К-комплексов, которые связаны с движениями ног (и часто предшествуют им). Препараты, повышающие выработку дофамина , такие как L-ДОФА , которые уменьшают движения ног, не уменьшают К-комплекс, что позволяет предположить, что они первичны, а движения ног вторичны по отношению к ним. Было высказано предположение, что неспособность таких препаратов уменьшить К-комплексы, несмотря на уменьшение движений ног, является причиной того, что пациенты после такого лечения продолжают жаловаться на невосстанавливающий сон. [ 13 ] Клоназепам — еще один метод лечения СБН; как и другие бензодиазепины , он подавляет быстрый сон за счет повышения уровня ГАМК . Это ингибирование быстрого сна значительно снижает количество К-комплексов, и в отличие от лечения L-ДОФА, исследования клоназепама сообщают об улучшении восстановления сна. [ 14 ] Следовательно, препараты, ингибирующие быстрый сон, также уменьшают количество К-комплексов.
Обструктивное апноэ во сне
[ редактировать ]Синдром обструктивного апноэ во сне связан с окклюзиями вдоха, вызывающими меньшее количество К-комплексов во время медленного сна, хотя К-комплексы обычно вызываются слуховыми стимулами, и такие люди нормально реагируют на остановки дыхания во время бодрствования. Это предполагает связь между таким апноэ во сне и притупленной реакцией коры головного мозга на респираторные проблемы. [ 15 ] [ 16 ] [ 17 ]
Примечания
[ редактировать ]- ^ Jump up to: а б с д и ж г час я Наличные СС; Халгрен Э.; Дегани Н.; и др. (2009). «Человеческий К-комплекс представляет собой изолированное корковое состояние» . Наука . 324 (5930): 1084–87. Бибкод : 2009Sci...324.1084C . дои : 10.1126/science.1169626 . ПМЦ 3715654 . ПМИД 19461004 .
- ^ Лумис А.Л.; Харви Э.Н.; Хобарт Джорджия (1937). «Мозговые состояния во время сна как исследование возможностей мозга человека». J Exp Psychol . 21 (2): 127–44. дои : 10.1037/h0057431 .
- ^ Рот М.; Шоу Дж.; Грин Дж. (1956). «Форма, распределение напряжения и физиологическое значение К-комплекса». Электроэнцефалическая клиника нейрофизиол . 8 (3): 385–402. дои : 10.1016/0013-4694(56)90004-9 . ПМИД 13330651 .
- ^ Вебстер К.Е.; Колрейн И.М. (1998). «Многоканальный ЭЭГ-анализ компонентов вызванного потенциала дыхания во время бодрствования и медленного сна». J Appl Physiol . 85 (5): 1727–35. дои : 10.1152/яп.1998.85.5.1727 . ПМИД 9804575 .
- ^ Маккормик Л., Нильсен Т., Николас А., Птито М., Монплезир Дж. (1997). «Топографическое распределение веретен и К-комплексов у нормальных людей» . Спать . 20 (11): 939–41. дои : 10.1093/сон/20.11.939 . ПМИД 9456457 .
- ^ Хаппе С.; Андерер П.; Грубер Г.; Клёш Г.; Салету Б.; Зейтлхофер Дж. (2002). «Топография скальпа спонтанного К-комплекса и дельта-волн во сне человека». Мозговой топогр . 15 (1): 43–9. дои : 10.1023/А:1019992523246 . ПМИД 12371676 . S2CID 21921834 .
- ^ Амзика Ф.; Стериаде М. (1998). «Клеточные субстраты и ламинарный профиль К-комплекса сна». Нейронаука . 82 (3): 671–86. дои : 10.1016/s0306-4522(97)00319-9 . ПМИД 9483527 . S2CID 26975545 .
- ^ Халас П. (2005). «К-комплекс, реактивный ЭЭГ-графоэлемент медленного сна: старик в новой одежде». Сон Мед. Преподобный . 9 (5): 391–412. дои : 10.1016/j.smrv.2005.04.003 . ПМИД 16122950 .
- ^ Jump up to: а б Тонони Г.; Сирелли К. (2006). «Функция сна и синаптический гомеостаз». Сон Мед. Преподобный . 10 (1): 49–62. дои : 10.1016/j.smrv.2005.05.002 . ПМИД 16376591 .
- ^ Jump up to: а б Вокье А. (октябрь 1993 г.). «Старение и изменения фазовых явлений во время сна». Физиол. Поведение . 54 (4): 803–6. дои : 10.1016/0031-9384(93)90095-w . PMID 8248360 . S2CID 7736698 .
- ^ Стериаде М.; Амзика Ф. (1998). «Медленные колебания сна, ритмические К-комплексы и их пароксизмальное развитие» . J Sleep Res . 7 (С1): 30–5. дои : 10.1046/j.1365-2869.7.s1.4.x . ПМИД 9682191 .
- ^ Эль Хелу Дж.; Наварро В.; Депьен К.; Федирко Е.; ЛеГерн Э.; Баулак М.; Ан-Гурфинкель И.; Адам С. (2008). «Припадки, индуцированные K-комплексом, при аутосомно-доминантной ночной эпилепсии лобных долей». Клин Нейрофизиол . 119 (10): 2201–4. дои : 10.1016/j.clinph.2008.07.212 . ПМИД 18762450 . S2CID 26640365 .
- ^ Монплезир Ж.; Баучер С.; Госселин А.; Пуарье Г.; Лавин Г. (1996). «Постоянство повторяющихся возбуждений ЭЭГ (комплексов К-альфа) у пациентов с СБН, получавших L-ДОФА» . Спать . 19 (3): 196–9. дои : 10.1093/sleep/19.3.196 . ПМИД 8723375 .
- ^ Салету М (2001). «Синдром беспокойных ног (RLS) и расстройство периодических движений конечностей (PLMD) острые плацебо-контролируемые лабораторные исследования сна с клоназепамом». Европейская нейропсихофармакология . 11 (2): 153–161. дои : 10.1016/s0924-977x(01)00080-3 . ПМИД 11313161 . S2CID 24658410 .
- ^ Хуан Дж.; Колрен ИМ; Мелендрес МС; Карамессинис Л.Р.; Пепе МЭ; Сэмюэл Дж.М.; Аби-Раад РФ; Трешер WH; Маркус КЛ (2008). «Корковая обработка дыхательных афферентных стимулов во время сна у детей с синдромом обструктивного апноэ во сне» . Спать . 31 (3): 403–10. дои : 10.1093/sleep/31.3.403 . ПМК 2276751 . ПМИД 18363317 .
- ^ Гора Дж., Триндер Дж., Пирс Р., Колрейн И.М. (ноябрь 2002 г.). «Доказательства притупленной реакции коры головного мозга на окклюзии вдоха при легком синдроме обструктивного апноэ во сне» . Являюсь. Дж. Респир. Крит. Уход Мед . 166 (9): 1225–34. дои : 10.1164/rccm.2106005 . ПМИД 12403692 .
- ^ Афифи Л.; Гиймино К.; Колрейн И.М. (2003). «Специфическое ослабление корковой реакции при СОАС во время сна и респираторных стимулов». Дыхание Физиол Нейробиол . 136 (2–3): 221–34. дои : 10.1016/s1569-9048(03)00084-3 . ПМИД 12853013 . S2CID 34199792 .