Атпаза водородного калия
| ATPase, H+/K+ обмен, альфа -полипептид | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | ATP4A | ||
| Ген NCBI | 495 | ||
| HGNC | 819 | ||
| Омим | 137216 | ||
| Refseq | NM_000704 | ||
| Uniprot | P20648 | ||
| Другие данные | |||
| ЕС номер | 7.2.2.19 | ||
| Локус | Хр. 19 Q13.1 | ||
| |||
| АТФаза, H+/K+ обмен, бета -полипептид | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | ATP4B | ||
| Ген NCBI | 496 | ||
| HGNC | 820 | ||
| Омим | 137217 | ||
| Refseq | NM_000705 | ||
| Uniprot | P51164 | ||
| Другие данные | |||
| ЕС номер | 7.2.2.19 | ||
| Локус | Chr | ||
| |||
Атпаза желудочного водорода калия , также известная как h + /K + АТФаза , является ферментом , который функционирует для подкисления желудка. [ 1 ] Он является членом АТФаз P -типа , также известных как E 1 -E 2 АТФазы из -за двух состояний. [ 2 ]
Биологическая функция и местоположение
[ редактировать ]желудочного водородного водорода Атпаза или h + /K + АТФаза является насосом желудка . протонным кишечника Он обменивается калием из просвета с цитоплазматическим гидрием [ 2 ] и является ферментом, главным образом, ответственным за подкисление содержания желудка и активацию пищеварительного фермента пепсин [ 3 ] (См. Желудочную кислоту ).
H. + /K + АТФаза обнаружена в теменных клетках , которые являются высокоспециализированными эпителиальными клетками, расположенными во внутренней клеточной слизистой оболочке желудка, называемой слизистой оболочкой желудка . Париетальные клетки обладают обширной секреторной мембранной системой и H + /K + АТФаза является основной составляющей белка этих мембран. Небольшое количество ч + /K + АТФаза также обнаружена в почечном мозговом мозге . [ 2 ]
Гены и структура белка
[ редактировать ]H. + /K + АТФаза является гетеродимерным белком , продуктом 2 генов. Ген ATP4A [ 4 ] кодирует h + /K + АТФаза α-субъединица и представляет собой белок ~ 1000-аминокислот, который содержит каталитические сайты фермента и образует пор через клеточную мембрану, которая позволяет транспортировать ионы. Ионы гидрония связываются с двумя активными сайтами, присутствующими в субъединице α. [ 5 ] Α -субъединица также имеет сайт фосфорилирования (ASP 385 ). [ 6 ] Ген ATP4B [ 7 ] кодирует β -субъединицу H + /K + АТФаза, которая представляет собой белок ~ 300-аминокислот с 3-аминокислотным N-концевым цитоплазматическим доменом, одним трансмембранным доменом и высокогликозилированным внеклеточным доменом.
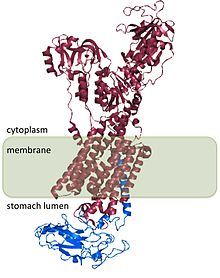
H. + /K + АТФаза β -субъединица стабилизирует H + /K + АТФаза α -субъединица и необходима для функции фермента. Β -субъединица предотвращает бег насос в обратном направлении, [ 8 ] и, по -видимому, также содержит сигналы, которые направляют гетеродимер к мембранным направлениям внутри клетки, хотя некоторые из этих сигналов подчинены сигналам, обнаруженным в H + /K + АТФаза α субъединица.
Структура h + /K + АТФаза была определена для людей, собак, свиней, крыс и кроликов и составляет 98% гомологичен всем видам. [ 2 ]
Ферментный механизм и активность
[ редактировать ]ЧАС + /K + АТФаза представляет собой атпазу P 2 -типа, член эукариотического класса АТФаз P -типа . [ 9 ] Как CA 2+ и Na + /K + Атфазы, h + /K + АТФаза функционирует как протомер A, B. [ 10 ] В отличие от других эукариотических АТФаз, H + /K + АТФаза является электроунитральной, транспортируя один протон в просвет желудка на ион калия, извлеченного из просвета желудка. [ 9 ] Как ионный насос + /K + АТФаза способна транспортировать ионы против градиента концентрации с использованием энергии, полученной из гидролиза АТФ. Как и все АТФазы P-типа, фосфатная группа переносится из аденозинтрифосфата (АТФ) в H + /K + АТФаза во время транспортного цикла. Этот перенос фосфата способствует конформационному изменению в ферменте, которое помогает управлять транспортом ионов. [ Цитация необходима ]
Атпаза водорода калия активируется косвенно гастрином , который заставляет клетки ECL выделять гистамин . [ 11 ] Гистамин связывается с рецепторами H2 на теменной клетке, активируя цАМФ-зависимый путь , который заставляет фермент перемещаться от цитоплазматических канальских мембран к глубоко сложенным каналам стимулированных теменных клетки. [ 2 ] После локализации фермент чередуется между двумя конформациями, E1 и E2, чтобы транспортировать ионы через мембрану.

Конформация E1 связывает фосфат из АТФ и иона гидрония на цитоплазматической стороне. Затем фермент изменяется в конформации E2, позволяя высвобождать гидроний в просвете. Конформация E2 связывает калий и возвращается к конформации E1 для высвобождения фосфата и k + в цитоплазму, где другой АТФ может быть гидролизован, чтобы повторить цикл. [ 2 ] Субъединица β предотвращает возвращение конформации E2-P к конформации E1-P, что делает протонное качание однонаправленным. [ 8 ] Количество ионов, транспортируемых на ATP, варьируется от 2 часа + /2K + до 1H + /1K + в зависимости от рН желудка. [ 12 ]
Актуальность и ингибирование заболевания
[ редактировать ]Ингибирование насоса водорода калия для уменьшения кислотности желудка было наиболее распространенным методом лечения заболеваний, включая гастроэзофагеальное рефлюксное заболевание (GERD/GORD) и язвенную болезнь (PUD). [ 13 ] Снижение кислотности облегчает симптомы заболевания, но не лечит фактическую причину ГЭРБ (аномальная релаксация сфинктера пищевода) или PUD ( Helicobacter pylori и NSAID ). [ 14 ]
Три класса лекарств были использованы для ингибирования H + /K + -Atpases. H 2 -Рецептор -антагонисты , такие как циметидин (тагамет), ингибируют сигнальный путь, который приводит к активации АТФазы. Этот тип ингибитора эффективен при лечении язв, но не предотвращает их образование, и у пациентов развивается толерантность к ним примерно через неделю, что приводит к снижению эффективности на 50%. [ 15 ] Позже были разработаны ингибиторы протонного насоса (PPI), начиная с тимопразола в 1975 году. [ 15 ] PPI представляют собой кислотные пролекарства , которые ингибируют водородную атпазу, связывают ковалентно с активными насосами. [ 16 ] Текущие PPI, такие как омепразол, имеют короткий период полураспада примерно 90 минут. [ 17 ] Антагонисты кислотных насосов (APA) или калий-конкурентоспособные блокаторы кислот (PCAB) являются третьим типом ингибитора, который блокирует секрецию кислоты путем связывания с K + Активный сайт. [ 15 ] APA обеспечивают более быстрое ингибирование, чем PPI, поскольку они не требуют кислотной активации. Revaprazan был первым APA, используемым клинически в Восточной Азии, и другие APA разрабатываются, поскольку они, по -видимому, обеспечивают лучший контроль кислоты в клинических испытаниях. [ 17 ]
Инактивация протонного насоса также может привести к проблемам со здоровьем. Исследование на мышах Krieg et al. [ 18 ] обнаружил, что мутация α-субъединицы насоса привела к ахлоргиджу , что привело к проблемам с поглощением железа, что приводит к дефициту железа и анемии . Использование PPI не было связано с повышенным риском анемии, поэтому H + /K + Считается, что АТФаза помогает поглощению железа, но не обязательно требуется. [ 18 ]
Текущая ассоциация деменции и ИПП была задокументирована в Германии и в исследовательских статьях, обозначающих то, как производные бензимидазола, астемизол (AST) и лансопразол (LNS) взаимодействуют с аномальными заполнителями тау белка ( нейрофибриллярные клубки ). [ 19 ] [ 20 ] [ 21 ] Текущие теории включают неселективную блокаду натриевых насосов в мозге, вызывающего осмотический дисбаланс или отеки в клетках. [Auth Mind] Взаимодействие PPI с другим препаратом, влияющим на насос натрия, например, дигоксин, варфарин и т. Д., Было хорошо задокументировано. [ 22 ] Память была связана с астроцитами, а субъединица Alpha3 рецептора аденозина, обнаруженная в насосах водорода/натрия, может быть фокусной точкой деменции. [ 23 ] [ 24 ] [ 25 ] Хроническое использование PPI может привести к снижению регуляции субъединицы Alpha3, увеличивающегося повреждения астроцитов. [ 26 ] Остеопетроз через ген TCIRG1 тесно связана с предсенильной деменцией. [ 27 ] [ 28 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Сакай Х., Фудзи Т., Таймгучи Н. (2016). «Глава 13. Proton-Potassium (H+/K+) АТФазы: свойства и роли в здоровье и заболеваниях». В Astrid S, Helmut S, Roland Ko S (Eds.). Ионы щелочных металлов: их роль в жизни . Металлические ионы в науках о жизни. Тол. 16. Springer. С. 459–483. doi : 10.1007/978-3-319-21756-7_13 . ISBN 978-3-319-21755-0 Полем PMID 26860309 .
- ^ Jump up to: а беременный в дюймовый и фон глин Shin, JM, Munson, K., Vagin, O., Sachs, G. (2009). «Желудочная HK-АТФаза: структура, функция и ингибирование» . Pflügers Archiv: Европейский журнал физиологии . 457 (3): 609–622. doi : 10.1007/s00424-008-0495-4 . PMC 3079481 . PMID 18536934 .
- ^ Berg, JM, Tymoczko, JL, Stryer, L. (2012). Биохимия (7 -е изд.). Нью -Йорк: WH Freeman and Company.
- ^ ATP4A ATPase H+/K+ Транспортировка альфа -субъединицы
- ^ Chourasia, M., Sastry, GM, Sastry. GN (2005). «Сайты связывания протонов и конформационный анализ H+K+-ATPase». Биохимическая и биофизическая исследовательская коммуникация . 336 (3): 961–966. doi : 10.1016/j.bbrc.2005.08.205 . PMID 16157306 .
- ^ Scheirlinckx, F., Raussens, V., Ruysschaert, J.-M., Goormaghtigh, E. (2004). «Конформационные изменения в желудочном H+/K+-АТФазах, контролируемых различием инфракрасной спектроскопии, трансформированной Фурье и обмена водорода/дейтерия» . Биохимический журнал . 382 (Pt 1) (Pt 1 Ed.): 121–129. doi : 10.1042/bj20040277 . PMC 1133922 . PMID 15096097 .
- ^ ATP4B ATPase H+/K+ Транспортировка бета -субъединицы
- ^ Jump up to: а беременный Abe, K., Tani, K., Nishizawa, T., Fujiyoshi, Y. (2009). "Межпуъединичное взаимодействие желудка h + К. + -Atpase предотвращает обратную реакцию транспортного цикла » . Embo Journal . 28 (11): 1637–1643. : 10.1038 /Emboj.2009.102 . PMC 2693145. . PMID 19387495 DOI
- ^ Jump up to: а беременный Shin, JM, Sachs, G. (2009). «Ионные мотивы АТФазы: АТФазы P-типа». эл . doi : 10.1002/9780470015902.a0001379.pub2 . ISBN 978-0-470-01617-6 .
- ^ Dach, I., Olesen, C., Signor, L., Nisse, P., The Mother, M., Møller, JV, Ebel, C. (2012). "Активные моющие средства-плавные h + К. + мономер . -это -Atpase »
- ^ Принц С., Каджимура М., Скотт Д., Хелландер Х., Шин Дж., Бесэнкон М., Бамберг К., Херси С., Сакс Г. (1992). «Кислотная секреция и H, K Атпаза желудка» . Йельский журнал биологии и медицины . 65 (6): 577–596. PMC 2589780 . PMID 1341065 .
- ^ Abe, K., Tani, K., Friedrich, T., Fujiyoshi, Y. (2012). «Крио-эм структура желудка H+, K+-ATPase с одним занятым катионо-связывающим сайтом» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (45): 18401–18406. Bibcode : 2012pnas..10918401a . doi : 10.1073/pnas.1212294109 . PMC 3494912 . PMID 23091039 .
- ^ Shin, JM, Vagin, O., Munson, K., Kidd, M., Modlin, IM, Sachs, G. (2008). «Молекулярные механизмы при терапии заболеваний, связанных с кислотами» . Клеточные и молекулярные науки о жизни . 65 (2): 264–281. doi : 10.1007/s00018-007-7249-x . PMC 3081136 . PMID 17928953 .
- ^ Yeomans, ND (2011). «Язвенные сыли: поиск причины язвенных язв» . Журнал гастроэнтерологии и гепатологии . 26 : 35–41. doi : 10.1111/j.1440-1746.2010.06537.x . PMID 21199512 . S2CID 42592868 .
- ^ Jump up to: а беременный в Sachs, G., Shin, JM, Vagin, O., Lambrecht, N., Yakubov, I., Munson, K. (2007). «Желудочный H, K ATPase как цель лекарства: прошлое, настоящее и будущее» . Журнал клинической гастроэнтерологии . 41 (Suppl 2): S226 - S242. doi : 10.1097/mcg.0b013e31803233b7 . PMC 2860960 . PMID 17575528 .
- ^ Shin JM, Sachs G. (2008). «Фармакология ингибиторов протонного насоса» . Текущие гастроэнтерологические отчеты . 10 (6): 528–534. doi : 10.1007/s11894-008-0098-4 . PMC 2855237 . PMID 19006606 .
- ^ Jump up to: а беременный Shin, JM, Sachs, G. (2009). «Длительные ингибиторы желудка H, K-ATPase» . Экспертный обзор клинической фармакологии . 2 (5): 461–468. doi : 10.1586/ecp.09.33 . PMC 2995460 . PMID 21132072 .
- ^ Jump up to: а беременный Криг Л., Милштейн О., Кребс П., Ся Ю., Бетлер Б., Дю, Х. (2011). «Мутация желудочно-водородной атпазы альфа-субъединицы вызывает анемию дефицита железа у мышей» . Кровь . 118 (24): 6418–6425. doi : 10.1182/blood-2011-04-350082 . PMC 3236123 . PMID 21976678 .
- ^ Gomm W, Von Holt K, Thomé F, Broich K, Maier W, Fink A, Doblhammer G, Haenisch B (2016). «Ассоциация ингибиторов протонных насосов с риском деменции: анализ данных фармакоэпидемиологических претензий» . Джама Нейрол . 73 (4): 410–416. doi : 10.1001/jamaneurol.2015.4791 . PMID 26882076 .
- ^ Rojo Le, Alzate-Morales J, Saavedra In, Davies P, Maccioni RB (2010). «Селективное взаимодействие лансопразола и ашемизола с полимерами тау: потенциальное новое клиническое использование при диагностике болезни Альцгеймера» . J Alzheimers Dis . 19 (2): 573–89. doi : 10.3233/jad-2010-1262 . PMC 2951486 . PMID 20110603 .
- ^ Фаваз М.В., Брукс А.Ф., Родник М.Е., Карпентер Г.М., Шао Х, Десмонд Т.Дж., Шерман П., Кесада К.А., Хокли Б.Г., Килбурн М.Р., Альбин Р.Л., Фрей К.А., Скотт П.Дж. (2014). «Радиофармацевтические препараты с высокой аффинностью, основанные на лансопразоле для визуализации домашних животных агрегированного тау при болезни Альцгеймера и прогрессирующего наддерного паралича: синтез, доклиническая оценка и выбор свинца» . ACS Chem Neurosci . 5 (8): 718–30. doi : 10.1021/cn500103u . PMC 4140593 . PMID 24896980 . Epub 2014 16 июня.
- ^ Трифиро Г., Коррао С., Алаква М., Моретти С., Тари М., Капути А.П., Аркорачи В. (2006). «Риск взаимодействия с ингибиторами протонных насосов в общей практике: значительное несогласие между различными источниками информации, связанных с наркотиками» . Br J Clin Pharmacol . 62 (5): 582–90. doi : 10.1111/j.1365-2125.2006.02687.x . PMC 1885178 . PMID 16822281 .
- ^ Björklund O, Shang M, Tonazzini I, Daré E, Fredholm BB (2008). «Аденозин A1 и A3 -рецепторы защищают астроциты от гипоксического повреждения». Eur J Pharmacol . 596 (1–3): 6–13. doi : 10.1016/j.ejphar.2008.08.002 . PMID 18727925 . EPUB 2008 13 августа.
- ^ Carmona MA, Murai KK, Wang L, Roberts AJ, Pasquale EB (2009). «Glial Ephrin-A3 регулирует морфологию дендритного позвоночника гиппокампа и транспорт глутамата» . Proc Natl Acad Sci USA . 106 (30): 12524–12529. BIBCODE : 2009PNAS..10612524C . doi : 10.1073/pnas.0903328106 . PMC 2718351 . PMID 19592509 .
- ^ Бен Хаим Л., Каррильо-де Соваге М.А., Цейзериат К., Эскартин С (2015). «Неуловимые роли реактивных астроцитов при нейродегенеративных заболеваниях» . Передние клеточные нейроски . 9 : 278. doi : 10.3389/fncel.2015.00278 . PMC 4522610 . PMID 26283915 . Ecollection 2015
- ^ Gessi S, Merighi S, Stefanelli A, Fazzi D, Varani K, Borea PA (2013). «A (1) и A (3) рецепторы аденозина ингибируют LPS-индуцированное накопление гипоксии, индуцируемое фактором-1 в мышиных астроцитах». Pharmacol Res . 76 : 157–70. doi : 10.1016/j.phrs.2013.08.002 . PMID 23969284 . Epub 2013 19 августа.
- ^ "Tcirg1" . Генетика дома ссылка .
- ^ Kafut T, Nagana J, Inui M, Mishima K, Momiyama T, Kaji M, Sugahara, Koito H, Ujike-Asai A, Nakura, Kanazawa K, Tan-Takeuchi K, Yokoayama WM, Kudo, Fijiwaria M, Dogoh Т (2003). «Остеопетроз и таламический гиперелиноз с синаптической дегенерацией у мышей с дефицитом DAP12 » » J Clin Invest 111 (3): 323–3 Doi : 10.1172/ jci1 PMC 151867 PMID 12569157
Внешние ссылки
[ редактировать ]- Yao X, Forte JG (2003). «Клеточная биология кислотной секреции париетальной клеткой» . Анну. Преподобный физиол . 65 : 103–31. doi : 10.1146/annurev.physiol.65.072302.114200 . PMID 12500969 .
- Kühlbrandt W (апрель 2004 г.). «Биология, структура и механизм АТФаз P-типа». НАТ Преподобный Мол. Клеточная биол . 5 (4): 282–95. doi : 10.1038/nrm1354 . PMID 15071553 . S2CID 24927167 .
- Данбар Л.А., Каплан М.Дж. (август 2001 г.). "Ионные насосы в поляризованных клетках: сортировка и регуляция NA + К. + - и ч + К. + -Atpases " . J. Biol. Chem . 276 (32): 29617–20. DOI : 10.1074/jbc.r100023200 . PMID 11404365 .
- Sachs G, Shin JM, Briving C, Wallmark B, Hersey S (1995). "Фармакология желудочного кислотного насоса: h + К. + ATPase " . Annu. Rev. Pharmacol. Toxicol . 35 : 277–305. DOI : 10.1146/annurev.pa.35.040195.001425 . PMID 7598495 .
- Калий+водород+АТФаза в Национальной библиотеке медицинской библиотеки США. Медицинские заголовки (Mesh)