Кора головного мозга
| Кора головного мозга | |
|---|---|
 | |
 Моторные и сенсорные области коры головного мозга | |
| Подробности | |
| Часть | Головность |
| Идентификаторы | |
| латинский | мозг коры |
| Сетка | D002540 |
| Нейроналы | 39 |
| Neurolex Id | Birnlex_1494 |
| TA98 | A14.1.09.003 A14.1.09.301 |
| TA2 | 5527 , 5528 |
| FMA | 61830 |
| Анатомические термины нейроанатомии | |
Кора головного мозга , также известная как мантия головного мозга , [ 1 ] внешним слоем ткани мозга мозга у . людей млекопитающих и других является нейронной Это крупнейший участок нейронной интеграции в центральной нервной системе , [ 2 ] и играет ключевую роль в внимании , восприятии , осознании , мысли , памяти , языке и сознании . Кора головного мозга является частью мозга, ответственной за познание .
Шестислойный неокортекс составляет приблизительно 90% коры , а AlloCortex составляет остаток. [ 3 ] Кора делится на левые и правые части продольной трещиной , которая разделяет два полушария головного мозга , которые соединяются под корой мозолистым корпусом . У большинства млекопитающих, помимо небольших млекопитающих, у которых есть маленькие мозги, сложена кора головного мозга, обеспечивая большую площадь поверхности в ограниченном объеме черепа . Помимо минимизации объема головного мозга и черепа, складывание коры имеет решающее значение для схемы мозга и ее функциональной организации. [ 4 ] У млекопитающих с маленьким мозгом нет складывания, а кора гладкая. [ 5 ] [ 6 ]
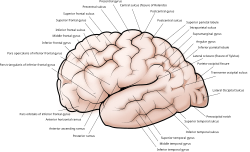
Склад или гребень в коре называется извилистой ( множественной гири), а канавка называется бороздкой (множественная бороздка). Эти поверхностные извилины появляются во время развития плода и продолжают созревать после рождения в процессе гирификации . В человеческом мозге большая часть коры головного мозга не видна снаружи, а похоронена на бороздке. [ 7 ] Основные разба и гири отмечают разногласия головного мозга на доли мозга . Четыре основных доли - фронтальные , теменные , затылочные и височные доли. Другие доли - лимбическая доля , а отдельная кора, часто называемая отдельной долей .
насчитывается от 14 до 16 миллиардов нейронов . В человеческой мозговой коре человека [ 2 ] Они организованы в горизонтальные слои коры и радиально в корковые колонны и миникологические добычи . Области коры имеют определенные функции, такие как движение в моторной коре , и зрение в зрительной коре . Моторная кора в основном расположена в презентальной извилине , а зрительная кора расположена в затылочной доле.
Структура
[ редактировать ]
Кора головного мозга представляет собой внешнее покрытие поверхностей полушарий головного мозга и складывается на пики, называемые гири , и канавки, называемые бороздками . В человеческом мозге он находится между 2 и 3-4 мм. толстый, [ 8 ] и составляет 40% массы мозга. [ 2 ] 90% коры головного мозга представляют собой шестислойный неокортекс, в то время как остальные 10% составляют трех/четырехслойных аллокортекса . [ 2 ] В коре от 14 до 16 миллиардов нейронов, [ 2 ] и они организованы радиально в корковых колоннах и миникологии , в горизонтально организованных слоях коры. [ 9 ] [ 10 ]
Некортекс разделяется на разные области коры, известные во множественном числе, как коры, и включают моторную кору и визуальную кору . Приблизительно две трети поверхности коры похоронены в бороздке, а острая кора полностью скрыта. Кора самая толстая по верхней части извилины и тонкая в нижней части бороздки. [ 11 ]
Складки
[ редактировать ]Кора головного мозга сложена таким образом, что позволяет большой площади поверхности нейронной ткани вписаться в ограничения нейрокрания . При развертывании в человеке каждая кора полушария имеет общую площадь поверхности около 0,12 квадратных метров (1,3 кв. Фута). [ 12 ] Складывание внутреннее от поверхности мозга, а также присутствует на медиальной поверхности каждого полушария в продольной трещине . У большинства млекопитающих есть кора головного мозга, которая запутана с пиками, известными как Гири, и впадины или канавки, известные как бороздки. Некоторые маленькие млекопитающие, в том числе некоторые мелкие грызуны, имеют гладкие мозговые поверхности без вращения . [ 6 ]
Доли
[ редактировать ]Большие бороздки и извилины отмечают подразделения коры головного мозга на доли мозга . [ 8 ] Есть четыре основные доли: лобная доля , теменная доля , височная доля и затылочная доля . часто Завязанная кора включается как островная доля. [ 13 ] Лимбическая доля представляет собой обод коры на медиальной стороне каждого полушария и также часто включается. [ 14 ] Существуют также три доля мозга, описанные: парацентральная доля , верхняя теменная доля и нижняя теменная долька .
Толщина
[ редактировать ]Для видов млекопитающих более крупные мозги (в абсолютном выражении, а не только по размеру тела), как правило, имеют более толстые кортики. [ 15 ] Самые маленькие млекопитающие, такие как зажженные , имеют неокортикальную толщину около 0,5 мм; Те, у кого есть самый большой мозг, такие как люди и пюры, имеют толщину 2–4 мм. [ 2 ] [ 8 ] Существует приблизительно логарифмическая связь между массой мозга и толщиной коры. [ 15 ] Магнитно -резонансная томография мозга (МРТ) позволяет получить меру толщины коры головного мозга человека и связывать ее с другими мерами. Толщина различных областей коры варьируется, но в целом сенсорная кора более тонкая, чем моторная кора. [ 16 ] Одно исследование показало некоторую положительную связь между толщиной коры и интеллектом . [ 17 ] Другое исследование показало, что соматосенсорная кора более толще у пациентов с мигрени , хотя неизвестно, является ли это результатом приступов мигрени, причины их или оба являются результатом общей причины. [ 18 ] [ 19 ] В более позднем исследовании с использованием более крупной популяции пациентов нет изменений в толщине коры у пациентов с мигренью. [ 20 ] Генетическое заболевание коры головного мозга, в результате которого уменьшение складывания в определенных областях приводит к микрогиру , где есть четыре слоя вместо шести, в некоторых случаях считается связанным с дислексией . [ 21 ]
Слои неокортекса
[ редактировать ]
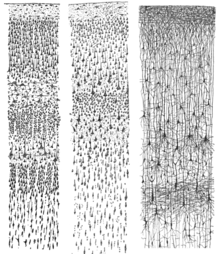


Neocortex белым образуется из шести слоев, пронумерованный I до VI, от самого внешнего слоя I - рядом с Pia Mater , до внутреннего слоя VI - рядом с основным веществом . Каждый кортикальный слой имеет характерное распределение различных нейронов и их связи с другими корковыми и подкорковыми областями. Существуют прямые связи между различными областями коры и косвенными соединениями через таламус.
Одним из самых ярких примеров слои коры является линия геннари в первичной зрительной коре . Это полоса белой ткани, которую можно наблюдать невооруженным глазом в кальцианской борозде затылочной доли. Линия геннари состоит из аксонов, которые приносят визуальную информацию из таламуса в слой IV зрительной коры .
Окрашивание поперечных сечений коры, чтобы выявить положение тел нейрональных клеток и внутрикорковые аксонные тракты позволили нейроанатомистам в начале 20-го века дать подробное описание ламинарной структуры коры у разных видов. Работа Корбинского Бродмана (1909) установила, что неокортекс млекопитающих последовательно разделяется на шесть слоев.
Слой i
[ редактировать ]Слой I представляет собой молекулярный слой коры головного мозга | Молекулярный слой и содержит несколько разбросанных нейронов, включая GABAergic Rosehip Neurons . [ 22 ] Слой I состоит в основном из расширений апикальных дендритных пучков пирамидных нейронов и горизонтально ориентированных аксонов, а также глиальных клеток . [ 4 ] Во время развития клетки Cajal - Retzius [ 23 ] и клетки субпиальных гранулированных слоев [ 24 ] присутствуют в этом слое. некоторые колючие звездчатые клетки Кроме того, здесь можно найти . Считается, что входные данные для апикальных пучков имеют решающее значение для взаимодействия обратной связи в коре головного мозга, участвующих в ассоциативном обучении и внимании. [ 25 ] Пока когда -то думали, что вход в слой, который я пришел из самой коры,, [ 26 ] Теперь понятно, что слой I через мантию коры головного мозга получает существенный вклад от матриц или клеток таламуса M-типа [ 27 ] (В отличие от ядра или C-типа, которые переходят на слой IV). [ 28 ]
Слой II
[ редактировать ]Слой II, внешний гранулированный слой , содержит небольшие пирамидные нейроны и многочисленные звездчатые нейроны.
Слой III
[ редактировать ]Слой III, внешний пирамидальный слой , содержит преимущественно малые и пирамидные нейроны среднего размера, а также не пирамидные нейроны с вертикально ориентированными внутрикортикатными аксонами; Слои I до III являются основной целью межполучанных кортикокортикальных афферентов , а слой III является основным источником кортикокортикальных эфферентов .
Слой IV
[ редактировать ]Слой IV, внутренний гранулированный слой , содержит различные типы звездчатых и пирамидальных клеток и является основной мишенью таламокортикальных афферентов из нейронов типа таламуса (ядро) [ 28 ] а также внутризлужие кортикокортикальные афференты. Слои над уровнем IV также называются надгранулярными слоями (слои I-III), тогда как приведенные ниже слои называются инфграгранулярными слоями (слои V и VI). Африканские слоны , китообразные и бегемоты не имеют слоя IV с аксонами, которые заканчиваются бы, вместо этого приходится на внутреннюю часть слоя III. [ 29 ]
Layer V
[ редактировать ]Слой V, внутренний пирамидальный слой , содержит большие пирамидальные нейроны. Аксоны из них покидают кору и соединяются с подкорковыми структурами, включая базальные ганглии . В первичной моторной коре лобной доли слой V содержит гигантские пирамидальные клетки, называемые клетками Betz , аксоны которых проходят через внутреннюю капсулу , ствол головного мозга и спинной мозг, образующий кортикоспинальный тракт , который является основным путем для добровольного контроля моторного движения Полем
Слой мы
[ редактировать ]Слой VI, полиморфный слой или многопрофильный слой , содержит несколько больших пирамидальных нейронов и множество небольших веретеноподобных пирамидальных и многопрофильных нейронов; Слой VI посылает эфферентные волокна в таламус, устанавливая очень точное взаимное взаимосвязь между корой и таламусом. [ 30 ] То есть, нейроны слоя VI из одной коры, соединяются с нейронами таламуса, которые обеспечивают вход в тот же коры. Эти связи являются как возбуждающими, так и ингибирующими. Нейроны посылают возбуждающие волокна в нейроны в таламусе, а также посылают коллатерали в таламическое ретикулярное ядро , которое ингибирует эти же нейроны таламуса или прилегающие к ним. [ 31 ] Одна теория заключается в том, что, поскольку ингибирующий выход уменьшается за счет холинергического ввода в кору головного мозга, это обеспечивает ствол мозга с регулируемым «контролем усиления для реле Lemniscal входов». [ 31 ]
Колонны
[ редактировать ]Корковые слои не просто сложены один над другим; Существуют характерные связи между различными слоями и типами нейронов, которые охватывают всю толщину коры. Эти корковые микросхемы сгруппированы в корковые колонны и миникологии . [ 32 ] Было предложено, чтобы миникологические добычи были основными функциональными единицами коры. [ 33 ] В 1957 году Вернон Маунткасл показал, что функциональные свойства резко изменяются между смежными точками коры между смежными точками; Однако они непрерывны в направлении перпендикулярно поверхности. Более поздние работы предоставили доказательства наличия функционально различных корковых колонн в зрительной коре (Hubel and Wiesel , 1959), [ 34 ] слуховая кора, и ассоциативная кора.
Корковые области, в которых отсутствует слой IV, называются агранными . Корковые области, которые имеют только рудиментарный слой IV, называются дисгранулярным. [ 35 ] Обработка информации в каждом слое определяется различной временной динамикой с той, что в слоях II/III, имеющих медленное 2 Гц колебание , в то время как в слое V есть быстрое колебание 10–15 Гц. [ 36 ]
Типы коры
[ редактировать ]Основываясь на различиях в ламинарной организации, кора головного мозга может быть классифицирована на два типа: большая область неокортекса , которая имеет шесть клеточных слоев, и гораздо меньшую площадь аллокордекса , которая имеет три или четыре слоя: [ 3 ]
- Neocortex также известен как изокортекс или неопаллий и является частью зрелой коры головного мозга с шестью различными слоями. Примеры неокортикальных областей включают гранулирующую первичную моторную кору и полосатую первичную зрительную кору . Neocortex имеет два подтипа, истинный изокортекс и проазокортекс , который представляет собой переходную область между изокортексом и областями периаллокортекса.
- AlloCortex является частью коры головного мозга с тремя или четырьмя слоями, и имеет три подтипа, палеокортекс с тремя пластинами кортикального коры, Archicortex , который имеет четыре или пять, и переходная область, прилегающая к Allocortex, периаллокортекс . Примерами AlloCortex являются обонятельная кора и гиппокамп .
Существует переходная площадь между неокортексом и аллокортексом, называемой паралимбической корой , где объединяются слои 2, 3 и 4. Эта область включает в себя просокортекс неокортекса и периаллокортекс Allocortex. Кроме того, кора головного мозга может быть классифицирована на четыре доли : лобная доля , височная доля , теменная доля и затылочная доля , названная из их вышележащих костей черепа.
Кровоснабжение и дренаж
[ редактировать ]
Кровяное снабжение коры головного мозга является частью мозговой циркуляции . Церебральные артерии поставляют кровь, которая перфузирует мозговой мозг. Эта артериальная кровь несет в кору кислорода, глюкозы и других питательных веществ. Церебральные вены истощают дезоксигенированную кровь, а метаболические отходы, включая углекислый газ, обратно в сердце.
Основными артериями, поставляющими кору, являются передняя мозговая артерия , средняя мозговая артерия и задняя мозговая артерия . Передняя церебральная артерия снабжает передние части мозга, включая большую часть лобной доли. Средняя церебральная артерия поставляет париетальные доли, височные доли и части затылочных дол. Средняя мозговая артерия разбивается на две ветви, чтобы поставить левое и правое полушарие, где они разветвляются дальше. Задняя мозговая артерия поставляет затылочные доли.
Круг Уиллиса является основной системой крови, которая занимается кровоснабжением в коре головного мозга и головного мозга.

Разработка
[ редактировать ]Пренатальное развитие коры головного мозга представляет собой сложный и тонко настроенный процесс, называемый кортикогенез , под влиянием взаимодействия между генами и окружающей средой. [ 37 ]
Нейронная трубка
[ редактировать ]Кора головного мозга развивается из самой передней части, области переднего мозга нейронной трубки . [ 38 ] [ 39 ] Нейронная пластина складывается и закрывается, чтобы сформировать нейронную трубку . Из полости внутри нейронной трубки развивается желудочковая система , и из нейроэпителиальных клеток ее стен нейроны и глия нервной системы. Самая передняя (передняя или черепная) часть нейронной плиты, Prosencephalon , который очевиден до начала невропалирования , приводит к полушарии головного мозга и более поздней коры. [ 40 ]
Развитие коркового нейрона
[ редактировать ]Корковые нейроны генерируются в желудочковой зоне , рядом с желудочками . Сначала эта зона содержит нервные стволовые клетки , которые переходят к радиальным глиальным клеткам - клетки -прогениторные клетки, которые делятся на продуцирование глиальных клеток и нейронов. [ 41 ]
Радиальная глия
[ редактировать ]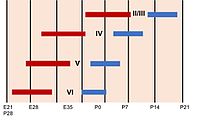
Кора головного мозга состоит из гетерогенной популяции клеток, которая приводит к различным типам клеток. Большинство этих клеток получены из радиальной миграции глиа , которые образуют различные типы клеток неокортекса, и это период, связанный с увеличением нейрогенеза . Точно так же процесс нейрогенеза регулирует ламинирование, образуя различные слои коры. Во время этого процесса наблюдается увеличение ограничения судьбы клеток, которая начинается с более ранних предшественников, которые вызывают любой тип клеток в коре, а затем предшественники, вызывающие только нейроны поверхностных слоев. Эта дифференциальная клеточная судьба создает топографию изнутри в коре с более молодыми нейронами в поверхностных слоях и более старых нейронах в более глубоких слоях. Кроме того, ламинарные нейроны останавливаются в фазе S или G2 , чтобы дать хорошее различие между различными слоями коры. Ламинарная дифференциация не полностью завершена до после рождения, поскольку во время развития ламинарные нейроны все еще чувствительны к внешним сигналам и сигналам окружающей среды. [ 42 ]
Хотя большинство клеток, составляющих кору, получены локально из радиальной глиа, существует подмножество нейронов, которые мигрируют из других областей. Радиальная глия приводит к нейронам, которые являются пирамидальной формой и используют глутамат в качестве нейротрансмиттера , однако эти мигрирующие клетки вносят нейроны, которые являются звездчатыми и используют ГАМК в качестве основного нейротрансмиттера. Эти ГАМКергические нейроны генерируются клетками -предшественниками в медиальной ганглионической возвышенности (MGE), которые мигрируют тангенциально в кору через субвентрикулярную зону . Эта миграция ГАМКергических нейронов особенно важна, поскольку рецепторы ГАМК возбуждают во время развития. Это возбуждение в первую очередь обусловлено потоком хлоридных ионов через рецептор ГАМК, однако у взрослых концентрации хлорида смещается, вызывая внутренний поток хлорида, который гиперполяризует постсинаптические нейроны . [ 43 ] Глиальные волокна, продуцируемые в первых подразделениях клеток -предшественников, радиально ориентированы, охватывая толщину коры от желудочковой зоны к внешней поверхности пиаль , и обеспечивают каркасы для миграции нейронов наружу из желудочковой зоны . [ 44 ] [ 45 ]
При рождении очень мало дендритов на клеточном теле кортикального нейрона, а аксон неразвит. В течение первого года жизни дендриты резко увеличиваются, так что они могут вместить до ста тысяч синаптических связей с другими нейронами. Аксон может развиваться, чтобы простираться далеко от тела ячейки. [ 46 ]
Асимметричное разделение
[ редактировать ]Первые подразделения клеток -предшественников симметричны, что дублирует общее количество клеток -предшественников в каждом митотическом цикле . Затем некоторые клетки -предшественники начинают делиться асимметрично, продуцируя одну постмитотическую клетку, которая мигрирует по радиальным глиальным волокнам, оставляя желудочковую зону , и одну клетку -предшественника, которая продолжает делиться до конца развития, когда он дифференцируется на глиальную клетку или или клетка Эпендимальная . Поскольку G1 фаза митоза удлиняется, в том , что рассматривается как селективное удлинение клеточного цикла, вновь рожденные нейроны мигрируют в более поверхностные слои коры. [ 47 ] Мигрирующие дочерние клетки становятся пирамидальными клетками коры головного мозга. [ 48 ] Процесс разработки упорядочен и регулируется сотнями генов и эпигенетическими регуляторными механизмами . [ 49 ]
Организация слоя
[ редактировать ]
Слоистая структура зрелой коры головного мозга образуется во время разработки. Первые пирамидные нейроны, генерируемые, мигрируют из желудочковой зоны и субвентрикулярной зоны , вместе с , нейронами, продуцирующими каджал -релис от подготовки . Затем, когорта нейронов, мигрирующих в середину подготовки, делит этот переходный слой на поверхностную маргинальную зону , которая станет слоем I зрелого неокортекса и подночика , [ 50 ] образуя средний слой, называемый кортикальной пластиной . Эти клетки будут образовывать глубокие слои зрелой коры, слои пять и шесть. Позже рожденные нейроны мигрируют радиально в корковую пластину мимо нейронов глубоких слоев и становятся верхними слоями (от двух до четырех). Таким образом, слои коры создаются в порядке изнутри. [ 51 ] Единственное исключение из этой внутренней последовательности нейрогенеза происходит в слое I приматов , в котором, в отличие от грызунов , нейрогенез продолжается на протяжении всего периода кортикогенеза . [ 52 ]
Корковое паттерн
[ редактировать ]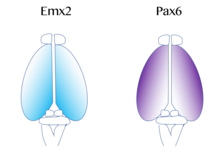
Карта функциональных областей коры, которые включают первичную моторную и визуальную кору, происходит от « протомапа », [ 53 ] который регулируется молекулярными сигналами, такими как фактор роста фибробластов FGF8 на ранних этапах эмбрионального развития. [ 54 ] [ 55 ] Эти сигналы регулируют размер, форму и положение областей коры на поверхности кортикального зачатка, частично, регулируя градиенты экспрессии транскрипционного фактора , посредством процесса, называемого паттерном кортикального . Примеры таких факторов транскрипции включают гены EMX2 и PAX6 . [ 56 ] Вместе оба фактора транскрипции образуют противоположный градиент выражения. Pax6 высоко экспрессируется на ростральном боковом полюсе, в то время как EMX2 высоко экспрессируется на каудомедиальном полюсе. Создание этого градиента важно для правильного развития. Например, мутации в PAX6 могут привести к расширению уровней EMX2 экспрессии EMX2, что в конечном итоге приведет к расширению областей, обычно полученных из хвостовой медиальной коры, таких как зрительная кора . Напротив, если возникают мутации в EMX2, это может привести к расширению домена, экспрессирующего PAX6, расширить и привести к увеличению лобных и моторных областей коры. Следовательно, исследователи считают, что подобные градиенты и сигнальные центры рядом с корой могут способствовать региональной экспрессии этих факторов транскрипции. [ 43 ] Два очень хорошо изученных сигнала паттерна для коры включают FGF и ретиноевую кислоту . Если FGFS MISEXEXPRED в различных областях развивающейся коры, паттерны кортикальной коры нарушаются. В частности, когда FGF8 увеличивается на переднем полюсе, EMX2 подавляется , и происходит хвостовое смещение в корковой области. Это в конечном итоге вызывает расширение ростральных областей. Следовательно, FGF8 и другие FGF играют роль в регуляции экспрессии EMX2 и PAX6 и представляют, как кора головного мозга может стать специализированной для различных функций. [ 43 ]
Быстрое расширение площади поверхности коры регулируется количеством самообновления радиальных глиальных клеток и частично регулируется генами FGF и Notch . [ 57 ] В период кортикального нейрогенеза и образования слоя многие более высокие млекопитающие начинают процесс гирификации , который генерирует характерные складки коры головного мозга. [ 58 ] [ 59 ] Гирификация регулируется ДНК-ассоциированным белком TRNP1 [ 60 ] FGF и SHH и передача сигналов [ 61 ] [ 62 ]
Эволюция
[ редактировать ]Из всех различных областей мозга кора головного мозга демонстрирует наибольшую эволюционную вариацию и развивалась в последнее время. [ 6 ] в отличие от высоко консервативной схемы мозгового мозга Например, , которая выполняет критические функции, такие как регуляция сердца и частоты дыхания, многие области коры головного мозга не являются строго необходимыми для выживания. Таким образом, в эволюции коры головного мозга появилась появление и модификация новых функциональных областей, особенно областей ассоциации, которые непосредственно не получают вход снаружи коры. [ 6 ]
Ключевая теория эволюции коры воплощена в гипотезе радиальной единицы и связанной с родственной гипотезой протомапа , впервые предложенной Rakic. [ 63 ] Эта теория гласит, что новые области коры формируются путем добавления новых радиальных единиц, которые достигаются на уровне стволовых клеток . Гипотеза Protomap утверждает, что клеточная и молекулярная идентичность и характеристики нейронов в каждой области коры определяются корковыми стволовыми клетками , известными как радиальные глиальные клетки , на первичной карте. Эта карта контролируется секретируемыми сигнальными белками и нижестоящими факторами транскрипции . [ 64 ] [ 65 ] [ 66 ]
Функция
[ редактировать ]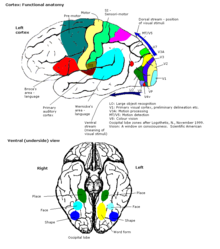
Соединения
[ редактировать ]Кора головного мозга связана с различными подкорковыми структурами, такими как таламус и базальные ганглии , отправляя им информацию вдоль эфферентных соединений и получение от них информации через афферентные соединения . Большая часть сенсорной информации направляется в кору головного мозга через таламус. Обонятельная информация, однако, проходит через обонятельную лампу в обонятельную кору ( пироформная кора ). Большинство соединений взяты из одной области коры к другой, а не от подкорковых областей; Braitenberg и Schüz (1998) утверждают, что в первичных сенсорных областях на уровне коры, где заканчиваются входные волокна, до 20% синапсов поставляются внекортикальными афферентами, но что в других областях и других слоях, вероятно, будет много ниже. [ 67 ]
Корковые области
[ редактировать ]Вся кора головного мозга была разделена на 52 различных областях в ранней презентации Корбинского Бродмана . Эти области, известные как области Бродмана , основаны на их цитоархитектуре , но также относятся к различным функциям. Примером является Brodmann Area 17, которая является основной визуальной корой .
В более общих терминах кора, как правило, описывается как три части: сенсорные, моторные и ассоциативные области.
Сенсорные области
[ редактировать ]
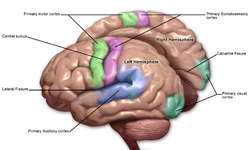
Сенсорные области - это корковые области, которые получают и обрабатывают информацию от чувств . Части коры, которые получают сенсорные входы от таламуса , называются первичными сенсорными областями. Чувства зрения, слуха и прикосновения обслуживаются первичной зрительной корой, первичной слуховой корой и первичной соматосенсорной корой соответственно. В целом, два полушария получают информацию с противоположной (контралатеральной) стороны тела . Например, правая первичная соматосенсорная кора получает информацию от левых конечностей, а правая визуальная кора получает информацию из левого поля зрения .
Организация сенсорных карт в коре отражает организацию соответствующего чувствительного органа в том, что известно как топографическая карта . соседние точки в первичной визуальной коре Например, соответствуют соседним точкам в сетчатке . Эта топографическая карта называется ретинотопической картой . Точно так же существует тонотопическая карта в первичной слуховой коре и соматотопическая карта в первичной сенсорной коре. Эта последняя топографическая карта тела на задней центральной извилине была иллюстрирована как деформированное человеческое представление, соматосенсорное гомункулус , где размер различных частей тела отражает относительную плотность их иннервации. Области с большой сенсорной иннервацией, такие как кончики пальцев и губы, требуют большей области кортикальной области для обработки более тонких ощущений.
Моторные зоны
[ редактировать ]Моторные зоны расположены в обоих полушариях коры. Моторные области очень тесно связаны с управлением добровольными движениями, особенно тонкими фрагментированными движениями, выполняемыми рукой. Правая половина области двигателя контролирует левую сторону тела, и наоборот.
Две области коры обычно называют мотором:
- Первичная моторная кора , которая выполняет добровольные движения [ 68 ]
- Дополнительные моторные области и премоторная кора , которая выбирает добровольные движения. [ 69 ]
Кроме того, моторные функции были описаны для:
- Задняя теменная кора , которая направляет добровольные движения в пространстве
- Дорсолатеральная префронтальная кора , которая решает, какие добровольные движения делать в соответствии с инструкциями высшего порядка, правилами и самогригнутыми мыслями.
Прямо под корой головного мозга находятся взаимосвязанные подкорковые массы серого вещества, называемых базальными ганглиями (или ядрами). Базальные ганглии получают вклад от субстанционной сутки среднего мозга и моторных областей коры головного мозга и отправляют сигналы обратно в обоих этих мест. Они участвуют в управлении двигателем. Они найдены латеральными для таламуса. Основными компонентами базальных ганглиев являются хвостовое ядро , путамен , Globus pallidus , субстанция нигра , прилежащее ядро и субталамическое ядро . Путамен и Globus pallidus также коллективно известны как ядро лентиформ , потому что вместе они образуют корпус в форме линзы. Путамен и хвостовое ядро также коллективно называют стриатумом корпуса после их полосатого внешнего вида. [ 70 ] [ 71 ]
Ассоциация областей
[ редактировать ]
Области ассоциации представляют собой части коры головного мозга, которые не принадлежат основным регионам. Они функционируют для создания значимого опыта восприятия мира, позволяют нам эффективно взаимодействовать и поддерживать абстрактное мышление и язык. Париетальные временные , - все , и затылочные доли расположенные в задней части коры - интегрируют сенсорную информацию и информацию, хранящуюся в памяти. Фронтальная доля или комплекс префронтальной ассоциации участвует в планировании действий и движения, а также абстрактной мысли. Во всем мире районы ассоциации организованы как распределенные сети. [ 72 ] Каждая сеть соединяет области, распределенные по широко расположенным областям коры. Отдельные сети расположены рядом друг с другом, что дает сложную серию переплетенных сетей. Конкретная организация ассоциационных сетей обсуждается с доказательствами взаимодействия, иерархических отношений и конкуренции между сетями.
У людей сети ассоциаций особенно важны для языковой функции. В прошлом было теоретизировано, что языковые способности локализованы в районе Брока в районах левой нижней лобной извилины , BA44 и BA45 , для выражения языка и в области Вернике BA22 , для приема языка. Тем не менее, было показано, что процессы выражения языка и приема происходят в областях, отличных от только этих структур вокруг боковой борозды , включая лобную долю, базальные ганглии , мозжечок и поны . [ 73 ]
Клиническое значение
[ редактировать ]
Нейродегенеративные заболевания , такие как болезнь Альцгеймера , демонстрируют как маркер, атрофию серой материи коры головного мозга. [ 75 ]
Другие заболевания центральной нервной системы включают неврологические расстройства, такие как эпилепсия , нарушения движения и различные типы афазии (трудности в выражении или понимании речи).
Повреждение головного мозга от болезней или травмы может включать повреждение определенной доли, например, при расстройстве лобной доли , и связанные с ними функции будут затронуты. Барьер -кровь -мрачный барьер , который служит для защиты мозга от инфекции, может стать скомпрометированным, что позволяет проникнуть в патогенные микроорганизмы .
подвержен Развивающийся плод целому ряду факторов окружающей среды, которые могут вызвать врожденные дефекты и проблемы в более позднем развитии. Например, потребление алкоголя матери может вызвать расстройство спектра алкоголя плода . [ 76 ] Другими факторами, которые могут вызвать расстройства развития нейродеоразвития, являются токсиканты , такие как лекарства , и воздействие радиации как от рентгеновских лучей . Инфекции также могут повлиять на развитие коры. Вирусная инфекция является одной из причин лиссенсфалии , которая приводит к гладкой коре без вращения .
Тип электрокортикографии, называемой картированием коры стимуляции , является инвазивной процедурой, которая включает в себя размещение электродов непосредственно на открытый мозг, чтобы локализовать функции определенных областей коры. Он используется в клинических и терапевтических применениях, включая предварительное картирование. [ 77 ]
Гены, связанные с нарушениями коры
[ редактировать ]Существует ряд генетических мутаций, которые могут вызвать широкий спектр генетических нарушений коры головного мозга, включая микроцефалию , шизенсифалию и типы лиссенсфалии . [ 78 ] Аномалии хромосомы также могут привести к тому, что у ряд нарушений нейродийного развития, таких как синдром хрупкого X и синдром Ретта .
Коды MCPH1 для микроцефалина , и расстройства в этом и в ASPM связаны с микроцефалией. [ 78 ] Мутации в гене NBS1 , который кодирует нибрин, могут вызвать синдром поломки Nijmegen , характеризующийся микроцефалией. [ 78 ]
Мутации в EMX2 , [ 79 ] и col4a1 связаны с шизенсефалией , [ 80 ] Состояние, отмеченное отсутствием больших частей полушарий головного мозга.
История
[ редактировать ]В 1909 году Корбинский Бродманн отличил 52 различных областях коры головного мозга на основе их цитоархитектуры. Они известны как области Бродмана . [ 81 ]
Рафаэль Лоренте -де -Ни , ученик Сантьяго Рамона и Каджал , определил более 40 различных типов корковых нейронов на основе распределения их дендритов и аксонов. [ 81 ]
Другие животные
[ редактировать ]Кора головного мозга получена из паллия , слоистую структуру, обнаруженную в переднем мозге всех позвоночных . Основной формой паллия является цилиндрический слой, охватывающий желудочки, заполненные жидкостью. Вокруг окружности цилиндра находится четыре зоны: дорсальный паллиум, медиальный паллий, вентральный паллий и латеральный паллий, которые, как считается, являются гомологичными для неокортекса , гиппокампа , миндалины и обонятельной коры соответственно.
В мозгах птичьего мозга данные свидетельствуют о том, что нейроархитектура птичьего паллия напоминает кору головного мозга млекопитающих. [ 82 ] Птиевой паллий также был предположил, что является эквивалентной нейронной основой для сознания . [ 83 ] [ 84 ]
До недавнего времени ни один аналог коры головного мозга не был признан в беспозвоночных. Тем не менее, исследование, опубликованное в «Журнальной ячейке сообщалось о сильном сродстве между корой головного мозга и грибными телами раггвормы » в 2010 году, основанное на профилях экспрессии генов , Platynereis dumerilii . [ 85 ] Грибные тела - это структуры в мозге многих типов червей и членистоногих, которые, как известно, играют важную роль в обучении и памяти; Генетические данные указывают на общее эволюционное происхождение и, следовательно, указывают на то, что происхождение самых ранних предшественников коры головного мозга датируется докамбрийской эрой.
Дополнительные изображения
[ редактировать ]-
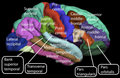 Боковая поверхность коры головного мозга человека
Боковая поверхность коры головного мозга человека -
 Медиальная поверхность коры головного мозга человека
Медиальная поверхность коры головного мозга человека -
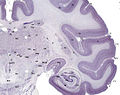 Ткани срез из мозга взрослой обезьяны макаки . Кора головного мозга изображена в темной фиолетовой.
Ткани срез из мозга взрослой обезьяны макаки . Кора головного мозга изображена в темной фиолетовой.



Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ "Церебральная мантия" . FreeDictionary.com . Получено 9 мая 2024 года .
- ^ Jump up to: а беременный в дюймовый и фон Саладин, Кеннет (2011). Анатомия человека (3 -е изд.). МакГроу-Хилл. С. 416–422. ISBN 9780071222075 .
- ^ Jump up to: а беременный Strominger, Norman L.; Demarest, Robert J.; Laemle, Lois B. (2012). "Церебральная кора". Человеческая нервная система Нобака, седьмое издание . Humana Press. С. 429–451. doi : 10.1007/978-1-61779-779-8_25 . ISBN 978-1-61779-778-1 .
- ^ Jump up to: а беременный Шипп, Стюарт (17 июня 2007 г.). «Структура и функция коры головного мозга» . Текущая биология . 17 (12): R443–9. Bibcode : 2007cbio ... 17.r443s . doi : 10.1016/j.cub.2007.03.044 . PMC 1870400 . PMID 17580069 .
- ^ Фернандес, V; Llinares-Benadero, c; Боррелл, V (17 мая 2016 г.). "Расширение и складывание коры головного мозга: что мы узнали?" Полем Embo Journal . 35 (10): 1021–44. doi : 10.15252/embj.201593701 . PMC 4868950 . PMID 27056680 .
- ^ Jump up to: а беременный в дюймовый Rakic, P (октябрь 2009 г.). «Эволюция неокортекса: перспектива биологии развития» . Nature Reports Neuroscience . 10 (10): 724–35. doi : 10.1038/nrn2719 . PMC 2913577 . PMID 19763105 .
- ^ Принципы нейронной науки (4 -е изд.). МакГроу-Хилл, Отдел медицинских профессий. 5 января 2000 г. ISBN 978-0838577011 .
- ^ Jump up to: а беременный в Робертс П. (1992). Нейроанатомия (3 -е изд.). Springer-Verlag. С. 86–92. ISBN 9780387977775 .
- ^ Лодато, Симона; Арлотта, Паола (13 ноября 2015 г.). «Генерирование разнообразия нейронов в коре головного мозга млекопитающих» . Ежегодный обзор биологии клеток и развития . 31 (1): 699–720. doi : 10.1146/annurev-cellbio-100814-125353 . PMC 4778709 . PMID 26359774 .
Функциональные столбцы были сначала определены в коре Маунтканом (1957), которые предложили столбчатую гипотезу, которая утверждает, что кора состоит из дискретных модульных столбцов нейронов, характеризующихся последовательным профилем связности.
- ^ Ansen-Wilson, LJ; Липински, RJ (январь 2017 г.). «Взаимодействие генов с окружающей средой в развитии и дисфункции кортикального интернейрона: обзор доклинических исследований» . Нейротоксикология . 58 : 120–129. Bibcode : 2017neutx..58..120a . doi : 10.1016/j.neuro.2016.12.002 . PMC 5328258 . PMID 27932026 .
- ^ Карпентер (1985). Основной текст нейроанатомии (3 -е изд.). Уильямс и Уилкинс. С. 348–358. ISBN 978-0683014556 .
- ^ Торо, Роберто; Перрон, Мишель; Пайк, Брюс; Богатнее, Луи; Вейлет, Сюзанна; Пасова, Здена; Паус, Томаш (1 октября 2008 г.). «Размер мозга и складывание коры головного мозга человека» . Кора головного мозга . 18 (10): 2352–2357. doi : 10.1093/cercor/bhm261 . ISSN 1047-3211 . PMID 18267953 .
- ^ Nieuwenhuys, R (2012). «Островная кора». Эволюция мозга примата . Прогресс в исследовании мозга. Тол. 195. С. 123–63. doi : 10.1016/b978-0-444-53860-4.00007-6 . ISBN 978-0-444-53860-4 Полем PMID 22230626 .
- ^ Tortora, G; Дерриксон, Б. (2011). Принципы анатомии и физиологии (13 -е изд.). Уайли. п. 549. ISBN 9780470646083 .
- ^ Jump up to: а беременный Newenhuys R, Denken HJ, Nicholson C (1998). Центральная нервная нервная система или диммеры, том 1 . Спрингер. стр. 2011–2012. ISBN 978-3-540-56013-5 .
- ^ Фритжоф Круггел; Мартина К. Брюкнер; Томас Арендт; Кристофер Дж. Виггинс; Д. Ив фон Крамон (2003). «Анализ неокортикальной тонкой структуры» . Анализ медицинского изображения . 7 (3): 251–264. doi : 10.1016/s1361-8415 (03) 00006-9 . HDL : 11858/00-001M-0000-0010-9C60-3 . PMID 12946467 .
- ^ Кэтрин Л. Нарр; Роджер П. Вудс; Пол М. Томпсон; Филипп Шеско; Дилберт Робинсон; Теодора Димтчева; Мала Гурбани; Артур В. Тога; Роберт М. Билдер (2007). «Отношения между IQ и региональной толщиной серого вещества у здоровых взрослых» . Кора головного мозга . 17 (9): 2163–2171. doi : 10.1093/cercor/bhl125 . PMID 17118969 .
- ^ Александр Ф.М. Дасильва; Кристина Гранцьера; Джош Снайдер; Наначена Хаджихани (2007). «Утолщение в соматосенсорной коре пациентов с мигренью» . Неврология . 69 (21): 1990–1995. doi : 10.1212/01.wnl.0000291618.32247.2d . PMC 3757544 . PMID 18025393 .
- ^ Катарина Паддок (20 ноября 2007 г.). «У страдающих мигрени есть более толстая кора головного мозга» . Медицинские новости сегодня . Архивировано из оригинала 11 мая 2008 года.
- ^ Datte R, Detre JA, et al. (Октябрь 2011). «Отсутствие изменений в толщине коры у пациентов с мигрени» . Цефалалгия . 31 (14): 1452–8. doi : 10.1177/0333102411421025 . PMC 3512201 . PMID 21911412 .
- ^ Хабиб М. (2000). «Неврологическая основа дислексии развития: обзор и рабочая гипотеза» . Мозг . 123 (12): 2373–99. doi : 10.1093/мозг/123.12.2373 . PMID 11099442 .
- ^ «Ученые идентифицируют новый вид клеток мозга человека» . Аллен институт . 27 августа 2018 года.
- ^ Мейер, Гундела; Гоффинет, Андре М.; Fairén, Alfonso (1999). «Статья о функциях: что такое ячейка Cajal -Rretzius? Переоценка классического типа клеток на основе недавних наблюдений в развивающихся неокортекс». Кора головного мозга . 9 (8): 765–775. doi : 10.1093/cercor/9.8.765 . PMID 10600995 .
- ^ Иудаш, Милош; Pletikos, Mihovil (2010). «Открытие подпиального гранулированного слоя в коре головного мозга человека» . Трансляционная нейробиология . 1 (3): 255–260. doi : 10.2478/v10134-010-0037-4 . S2CID 143409890 .
- ^ Гилберт CD, Sigman M (2007). «Состояния мозга: нисходящие влияния в сенсорной обработке» . Нейрон . 54 (5): 677–96. doi : 10.1016/j.neuron.2007.05.019 . HDL : 11336/67502 . PMID 17553419 .
- ^ Cauller L (1995). «Слой I первичного сенсорного неокортекса: где сверху вниз сходится нанизу вверх». Поведение мозг res . 71 (1–2): 163–70. doi : 10.1016/0166-4328 (95) 00032-1 . PMID 8747184 . S2CID 4015532 .
- ^ Рубио-Гарридо П., Перес-де-Манзо Ф., Порреро С., Галазо М.Дж., Класка Ф. (2009). «Таламический вход в дистальные апикальные дендриты в неокортикальном слое 1 является массивным и очень конвергентным» . Мозговая кора . 19 (10): 2380–95. doi : 10.1093/cercor/bhn259 . PMID 19188274 .
- ^ Jump up to: а беременный Джонс, EG (1998). «Точка зрения: ядро и матрица таламической организации». Нейробиология . 85 (2): 331–45. doi : 10.1016/s0306-4522 (97) 00581-2 . PMID 9622234 . S2CID 17846130 .
- ^ Bhawwadin, Adil; Моллнар, Золтан; Bertellen, Mads F.; Карлссон, Карл Эр.; Agaili, Abdulaziz n.; Беннетт, Нигель С.; Хоф, Патрик Р.; Kapwalk -Comamy, Assoled; Знак, Эммануэль; Джайкар, Джайкшан; Мангер, Пол Р. (2024). Полем Журнал 532 (7). doi : 10.1002/cne . ISSN 0021-9
- ^ Creutzfeldt, O. 1995. Cortex cerebri . Спрингер работает.
- ^ Jump up to: а беременный Лам Ю.В., Шерман С.М. (2010). «Функциональная организация соматосенсорного кортикального слоя 6 обратной связи с таламусом» . Мозговая кора . 20 (1): 13–24. doi : 10.1093/cercor/bhp077 . PMC 2792186 . PMID 19447861 .
- ^ Suzuki, ik; Хирата, Т (январь 2013 г.). «Некортикальный нейрогенез на самом деле не является« нео »: новая эволюционная модель, полученная в результате сравнительного исследования паллиального развития цыпочек» (PDF) . Развитие, рост и дифференциация . 55 (1): 173–87. doi : 10.1111/dgd.12020 . PMID 23230908 . S2CID 36706690 .
- ^ Mountcastle V (1997). «Сторонная организация неокортекс» . Мозг . 120 (4): 701–722. doi : 10.1093/мозг/120.4.701 . PMID 9153131 .
- ^ Hubel DH, Wiesel TN (октябрь 1959 г.). «Восприимчивые поля отдельных нейронов в полосатой коре кошки» . Журнал физиологии . 148 (3): 574–91. doi : 10.1113/jphysiol.1959.sp006308 . PMC 1363130 . PMID 14403679 .
- ^ SM Dombrowski, CC Hilgetag и H. Barbas. Количественная архитектура различает префронтальные системы кортикальной коры в архивировании обезьян-резуса 2008-08-29 на машине Wayback .cereb. Кора 11: 975–988. «... им либо не хватает (агранного), либо имеют только рудиментарный гранулированный слой IV (дисгранулярный)».
- ^ Sun W, Dan Y (2009). «Специфичные для слоя сетевые колебания и пространственно-временное восприимчивое поле в зрительной коре» . Proc Natl Acad Sci USA . 106 (42): 17986–17991. Bibcode : 2009pnas..10617986S . doi : 10.1073/pnas.0903962106 . PMC 2764922 . PMID 19805197 .
- ^ Плетикос, Миховил; Соуза, Андре М.М.; и др. (22 января 2014 г.). «Временная спецификация и двусторонность неокортикальной топографической экспрессии генов человека» . Нейрон . 81 (2): 321–332. doi : 10.1016/j.neuron.2013.11.018 . PMC 3931000 . PMID 24373884 .
- ^ Wolpert, Lewis (2015). Принципы развития (пятое изд.). Великобритания: издательство Оксфордского университета. п. 533. ISBN 9780199678143 .
- ^ Уоррен Н., Карич Д., Пратт Т., Клаузен Дж.А., Асаваритикрай П., Мейсон Дж.О., Хилл Р.Е., Прайс Д.Джея (1999). «Фактор транскрипции, PAX6, необходим для пролиферации и дифференцировки клеток в развивающейся коре головного мозга» . Кора головного мозга . 9 (6): 627–35. doi : 10.1093/cercor/9.6.627 . PMID 10498281 .
- ^ Ларсен, W J. Эмбриология человека 3-е издание 2001 г. с. 421-422 ISBN 0-443-06583-7
- ^ Стивен С. Ноктор; Александр С. Флинт; Тамилия А. Вайсман ; Райан С. Даммерман и Арнольд Р. Кригштейн (2001). «Нейроны, полученные из радиальных глиальных клеток, устанавливают радиальные единицы в неокортексе». Природа . 409 (6821): 714–720. Bibcode : 2001natur.409..714n . doi : 10.1038/350555553 . PMID 11217860 . S2CID 3041502 .
- ^ Sur, Mriganka; Лими, Кэтрин А. (2001). «Разработка и пластичность корковых областей и сетей». Nature Reports Neuroscience . 2 (4): 251–262. doi : 10.1038/35067562 . PMID 11283748 . S2CID 893478 .
- ^ Jump up to: а беременный в Sanes, Dan H.; Рех, Томас А.; Харрис, Уильям А. (2012). Разработка нервной системы . Elsevier Inc. ISBN 978-0-12-374539-2 .
- ^ Rakic, P (октябрь 2009 г.). «Эволюция неокортекса: перспектива биологии развития» . Nature Reports Neuroscience . 10 (10): 724–35. doi : 10.1038/nrn2719 . PMC 2913577 . PMID 19763105 .
- ^ Rakic, P (ноябрь 1972). «Внешние цитологические детерминанты дендритной картины корзины и звездчатых клеток в молекулярном слое мозжечка». Журнал сравнительной неврологии . 146 (3): 335–54. doi : 10.1002/cne.901460304 . PMID 4628749 . S2CID 31900267 .
- ^ Гилберт, Скотт (2006). Биология развития (8 -е изд.). Sinauer Associates Publishers. С. 394–395. ISBN 9780878932504 .
- ^ Калегари, F; Haubensack w; Хаффнер С; Хаттнер WB (2005). «Селективное удлинение клеточного цикла в нейрогенной субпопуляции нейронных клеток -предшественников во время развития мозга мыши» . Журнал нейробиологии . 25 (28): 6533–8. doi : 10.1523/jneurosci.0778-05.2005 . PMC 6725437 . PMID 16014714 .
- ^ П. Ракич (1988). «Спецификация областей коры головного мозга». Наука . 241 (4862): 170–176. Bibcode : 1988sci ... 241..170r . doi : 10.1126/science.3291116 . PMID 3291116 .
- ^ Ху, XL; Wang, y.; Шен, Q. (2012). «Эпигенетический контроль по выбору клеточной судьбы в нейронных стволовых клетках» . Белок и клетка . 3 (4): 278–290. doi : 10.1007/s13238-012-2916-6 . PMC 4729703 . PMID 22549586 .
- ^ Костович, Авика (1990). «История развития зоны переходной подтчики в визуальной и соматосенсорной коре макак -обезьяны и человеческого мозга». Журнал сравнительной неврологии . 297 (3): 441–470. doi : 10.1002/cne.902970309 . PMID 2398142 . S2CID 21371568 .
- ^ Rakic, P (1 февраля 1974 г.). «Нейроны в визуальной коре обезьян -резуса: систематическая связь между временем происхождения и возможным расположением». Наука . 183 (4123): 425–7. Bibcode : 1974sci ... 183..425r . doi : 10.1126/science.183.4123.425 . PMID 4203022 . S2CID 10881759 .
- ^ Zecevic N, Rakic P (2001). «Развитие нейронов слоя I в коре головного мозга приматов» . Журнал нейробиологии . 21 (15): 5607–19. doi : 10.1523/jneurosci.21-15-05607.2001 . PMC 6762645 . PMID 11466432 .
- ^ Rakic, P (8 июля 1988 г.). «Спецификация областей коры головного мозга». Наука . 241 (4862): 170–6. Bibcode : 1988sci ... 241..170r . doi : 10.1126/science.3291116 . PMID 3291116 .
- ^ Фукучи-симогори, т; Гроув, EA (2 ноября 2001 г.). «Неокортекс -паттерн секретируемой сигнальной молекулой FGF8» . Наука . 294 (5544): 1071–4. Bibcode : 2001sci ... 294.1071f . doi : 10.1126/science.1064252 . PMID 11567107 . S2CID 14807054 .
- ^ Garel, S; Хаффман, KJ; Рубенштейн, JL (май 2003 г.). «Молекулярная регионализация неокортекса нарушается у гипоморфных мутантов FGF8». Разработка . 130 (9): 1903–14. doi : 10.1242/dev.00416 . PMID 12642494 . S2CID 6533589 .
- ^ Епископ, Км; Гудро, G; О'Лири, DD (14 апреля 2000 г.). «Регуляция идентичности площади в неокортексе млекопитающих от EMX2 и PAX6». Наука . 288 (5464): 344–9. Bibcode : 2000sci ... 288..344b . doi : 10.1126/science.288.5464.344 . PMID 10764649 .
- ^ Rash, Bg; Lim, HD; Брейниг, JJ; Vaccarino, FM (26 октября 2011 г.). «Передача сигналов FGF расширяет площадь эмбриональной кортикальной поверхности, регулируя Notch-зависимый нейрогенез» . Журнал нейробиологии . 31 (43): 15604–17. doi : 10.1523/jneurosci.4439-11.2011 . PMC 3235689 . PMID 22031906 .
- ^ Раджагопалан, V; Скотт, J; Хабас, Пенсильвания; Ким, К; Corbett-Detig, J; Руссо, F; Баркович, AJ; Гленн, ОА; Studholme, C (23 февраля 2011 г.). «Местные паттерны роста тканей, лежащие в основе нормального гирификации мозга человека плода, определяли количественно в утробере» . Журнал нейробиологии . 31 (8): 2878–87. doi : 10.1523/jneurosci.5458-10.2011 . PMC 3093305 . PMID 21414909 .
- ^ Луи, январь Х.; Хансен, Дэвид В.; Кригштейн, Арнольд Р. (8 июля 2011 г.). «Развитие и эволюция человеческого неокортекса» . Клетка . 146 (1): 18–36. doi : 10.1016/j.cell.2011.06.030 . ISSN 1097-4172 . PMC 3610574 . PMID 21729779 .
- ^ Шталь, Ронни; Уолчер, Тесса; Де Хуан Ромеро, Камино; Пилз, Грегор Александр; Каппелло, Сильвия; Ирмлер, Мартин; Санц-Акела, Хосе Мигель; Бекерс, Йоханнес; Блум, Роберт (25 апреля 2013 г.). «TRNP1 регулирует расширение и складывание коры головного мозга млекопитающих путем контроля радиальной глиальной судьбы» . Клетка . 153 (3): 535–549. doi : 10.1016/j.cell.2013.03.027 . HDL : 10261/338716 . ISSN 1097-4172 . PMID 23622239 .
- ^ Ван, Лей; Хоу, Шируи; Хан, Янг-Гу (23 мая 2016 г.). «Передача сигналов ежа способствует расширению базального предшественника и росте и складывании неокортекса» . Nature Neuroscience . 19 (7): 888–96. doi : 10.1038/nn.4307 . ISSN 1546-1726 . PMC 4925239 . PMID 27214567 .
- ^ Rash, Брайан Дж.; Томаси, Симона; Лим Х. Дэвид; Су, Кэрол Ю.; Vaccarino, Flora M. (26 июня 2013 г.). «Корковое вращение, вызванное фактором роста фибробластов 2 в мозге мыши» . Журнал нейробиологии . 33 (26): 10802–10814. doi : 10.1523/jneurosci.3621-12.2013 . ISSN 1529-2401 . PMC 3693057 . PMID 23804101 .
- ^ Rakic, P (8 июля 1988 г.). «Спецификация областей коры головного мозга». Наука . 241 (4862): 170–6. Bibcode : 1988sci ... 241..170r . doi : 10.1126/science.3291116 . PMID 3291116 .
- ^ Фукучи-симогори, т; Гроув, EA (2 ноября 2001 г.). «Неокортекс -паттерн секретируемой сигнальной молекулой FGF8» . Наука . 294 (5544): 1071–4. Bibcode : 2001sci ... 294.1071f . doi : 10.1126/science.1064252 . PMID 11567107 . S2CID 14807054 .
- ^ Епископ, Км; Гудро, G; О'Лири, DD (14 апреля 2000 г.). «Регуляция идентичности площади в неокортексе млекопитающих от EMX2 и PAX6». Наука . 288 (5464): 344–9. Bibcode : 2000sci ... 288..344b . doi : 10.1126/science.288.5464.344 . PMID 10764649 .
- ^ Гроув, EA; Фукучи-Симогори, Т (2003). «Создание карты кортикальной области головного мозга». Ежегодный обзор нейробиологии . 26 : 355–80. doi : 10.1146/annurev.neuro.26.041002.131137 . PMID 14527269 . S2CID 12282525 .
- ^ Braitenberg, V и Schüz, 1998. "Кора: статистика и геометрия связности нейронов. Второе тщательно пересмотренное издание" Нью-Йорк: Springer-Verlag
- ^ "Церебральная кора - наука о науке" .
- ^ «Поведенческое планирование: нейрофизиологический подход функции лобной доли у приматов - ScienceDirect» .
- ^ Саладин, Кеннет. Анатомия и физиология: единство формы и функции, 5 -е изд. Нью-Йорк: McGraw-Hill Companies Inc., 2010. Печать.
- ^ Медицинский словарь Дорланда для потребителей здравоохранения, 2008.
- ^ Yeo BT, Krienen FM, Sepulcre J, Sabuncu MR, Lashkari D, Hollinshead M, Roffman JL, Smoller JW, Zöllei L, Polimeni JR, Fischl B, Liu H, Buckner RL (2011). «Организация коры головного мозга человека, оцениваемой по внутренней функциональной связности» . Журнал нейрофизиологии . 106 (3): 1125–1165. doi : 10.1152/jn.00338.2011 . PMC 3174820 . PMID 21653723 .
- ^ Кэти Дж. Прайс (2000). «Анатомия языка: вклад от функционального нейровизуализации» . Журнал анатомии . 197 (3): 335–359. doi : 10.1046/j.1469-7580.2000.19730335.x . PMC 1468137 . PMID 11117622 .
- ^ Кентар, Модар; Манн, Мартина; Сам, Феликс; Оливарес-Ривера, Артуро; Санчес-Пуррас, Ренан; Zerelles, Роланд; Саковиц, Оливер В.; Unterberg, Andreas W.; Сантос, Эдгар (15 января 2020 года). «Обнаружение распространения деполяризации в модели окклюзии средней мозговой артерии у свиней». Acta Neurochirurgica . 162 (3): 581–592. doi : 10.1007/s00701-019-04132-8 . ISSN 0942-0940 . PMID 31940093 . S2CID 210196036 .
- ^ Наказава Т., Охара Т., Хирабаяши Н., Фурута Ю., Хата Дж., Шибата М., Хонда Т., Китазоно Т., Накао Т., Ниномия Т (март 2022 г.). «Атрофия с несколькими регионами серого вещества как предиктора развития деменции в сообществе: исследование Хисаямы» . J Neurol Neurosurg Psychiatry . 93 (3): 263–271. doi : 10.1136/jnnp-2021-3266111 . PMC 8862082 . PMID 34670843 .
- ^ Мукерджи, Раджа как; Холлинс, Шейла (2006). «Спектр спектра алкоголя плода: обзор» . Журнал Королевского общества медицины . 99 (6): 298–302. doi : 10.1177/014107680609900616 . PMC 1472723 . PMID 16738372 .
- ^ Тарапор, PE; и др. (Август 2012 г.). «Предоперационное мультимодальное отображение двигателя: сравнение магнитоэнцефалографической визуализации, навигационной транскраниальной магнитной стимуляции и прямой стимуляции коры» . Журнал нейрохирургии . 117 (2): 354–62. doi : 10.3171/2012.5.jns112124 . PMC 4060619 . PMID 22702484 .
- ^ Jump up to: а беременный в Уолш, Кристофер А.; Mochida, Ganeshwaran H. (1 мая 2004 г.). «Генетическая основа развития развития коры головного мозга». Архив неврологии . 61 (5): 637–640. doi : 10.1001/archneur.61.5.637 . PMID 15148137 .
- ^ «EMX2 пустые воздушные спиралы Homeobox 2 [Homo sapiens (человек)] - ген - NCBI» . www.ncbi.nlm.nih.gov .
- ^ Смигиэль, R; Кабала, м; Якубиак, а; Kodera, h; Sasiadek, MJ; Matsumoto, n; Sasiadek, MM; Saitsu, H (апрель 2016 г.). «Новая мутация col4a1 у младенца с тяжелым дисморфическим синдромом с шизенсефалией, перивентрикулярными кальцификациями и катаракта, напоминающей врожденную инфекцию». Врожденные дефекты исследования. Часть A, клиническая и молекулярная тератология . 106 (4): 304–7. doi : 10.1002/bdra.23488 . PMID 26879631 .
- ^ Jump up to: а беременный Принципы нейронной науки . Кандель, Эрик Р. (5 -е изд.). Нью-Йорк. 2013. С. 347–348. ISBN 9780071390118 Полем OCLC 795553723 .
{{cite book}}: CS1 Maint: местоположение отсутствует издатель ( ссылка ) CS1 Maint: Другие ( ссылка ) - ^ Стахо, Мартин; Херольд, Кристина; Ладья, Ноэми; Вагнер, Германн; Аксер, Маркус; Амунты, Катрин; Гюнюркюн, Онур (25 сентября 2020 г.). «Каноническая цепь, подобная коре, в переднем мозге птиц» . Наука . 369 (6511). doi : 10.1126/science.abc5534 . ISSN 0036-8075 . PMID 32973004 .
- ^ Нидер, Андреас; Вагенер, Лисанн; Риннерт, Пол (25 сентября 2020 г.). «Нейронная корреляция сенсорного сознания в птице Корвида» . Наука . 369 (6511): 1626–1629. Bibcode : 2020sci ... 369.1626N . doi : 10.1126/science.abb1447 . ISSN 0036-8075 . PMID 32973028 . S2CID 221881862 .
- ^ Геркулано-Хоузель, Сюзана (25 сентября 2020 г.). «У птиц есть кора мозга - и думают» . Наука . 369 (6511): 1567–1568. Bibcode : 2020sci ... 369.1567H . doi : 10.1126/science.abe0536 . ISSN 0036-8075 . PMID 32973020 . S2CID 221882004 .
- ^ Томер, R; DENES, AS; Tessmar-rabible, k; Арендт, D; Томер R; DENS AS; Tessmar-rabible k; Арендт Д. (2010). «Профилирование по регистрации изображений показывает общее происхождение грибных тел и паллия с позвоночными» . Клетка . 142 (5): 800–809. doi : 10.1016/j.cell.2010.07.043 . PMID 20813265 . S2CID 917306 .
Внешние ссылки
[ редактировать ]- Здесь-20 в нейронаме
- Окрашенные изображения срезов мозга, которые включают «кору головного мозга» в проекте Brainmaps
- «Основная визуальная кора» , Webvision: Комплексная статья о структуре и функции первичной зрительной коры.
- «Основные типы ячеек» , WebVision: изображение основных типов ячеек коры головного мозга обезьяны.
- Кора головного мозга - база данных с центром ячейки