Непротеиногенные аминокислоты

В биохимии . некодированные или непротеиногенные аминокислоты отличаются от 22 протеиногенных аминокислот (21 у эукариот) [примечание 1] ), которые естественным образом закодированы в геноме организмов для сборки белков. Однако более 140 непротеиногенных аминокислот встречаются в белках в природе, а еще тысячи могут встречаться в природе или быть синтезированы в лаборатории. [1] Химически синтезированные аминокислоты можно назвать неприродными аминокислотами. Неприродные аминокислоты можно получить синтетически из их нативных аналогов посредством модификаций, таких как алкилирование амина, замена боковой цепи, циклизация удлинения структурных связей и изостерические замены в основной цепи аминокислоты. [2] Важны многие непротеиногенные аминокислоты:
- промежуточные продукты биосинтеза,
- в посттрансляционном образовании белков,
- в физиологической роли (например, компоненты клеточных стенок бактерий , нейротрансмиттеры и токсины ),
- природные или искусственные фармакологические соединения,
- присутствует в метеоритах или используется в пребиотических экспериментах (таких как эксперимент Миллера-Юри ),
- могут быть важными нейротрансмиттерами, такими как γ-аминомасляная кислота , [3] и
- может играть решающую роль в клеточной биоэнергетике, например, креатин . [4]
Определение отрицанием
[ редактировать ]
Технически любое органическое соединение с амина (–NH 2 ) и карбоновой кислоты (–COOH) функциональной группой является аминокислотой. Протеиногенные аминокислоты представляют собой небольшую подгруппу этой группы, которая имеет центральный атом углерода (α- или 2-), несущий аминогруппу, карбоксильную группу, боковую цепь и α-водородную левоконформацию , за исключением глицина , который ахиральный и пролин , аминогруппа которого является вторичным амином и, следовательно, по традиционным причинам часто упоминается как иминокислота , хотя и не имино.
Генетический код кодирует 20 стандартных аминокислот для включения в белки во время трансляции . Однако есть две дополнительные протеиногенные аминокислоты: селеноцистеин и пирролизин . Эти нестандартные аминокислоты не имеют специального кодона, но добавляются вместо стоп-кодона при наличии определенной последовательности, кодона UGA и элемента SECIS для селеноцистеина. [5] UAG Последующая последовательность PYLIS для пирролизина. [6] Все остальные аминокислоты называются «непротеиногенными».
 Селеноцистеин. Эта аминокислота содержит селенольную группу на своем β-углероде.
Селеноцистеин. Эта аминокислота содержит селенольную группу на своем β-углероде. Пирролизин. Эта аминокислота образуется путем присоединения к ε-аминогруппе лизина карбоксилированного пирролинового кольца.
Пирролизин. Эта аминокислота образуется путем присоединения к ε-аминогруппе лизина карбоксилированного пирролинового кольца.

Существуют различные группы аминокислот: [7]
- 20 стандартных аминокислот
- 22 протеиногенные аминокислоты
- более 80 аминокислот, созданных абиотически в высоких концентрациях
- около 900 производятся естественным путем
- более 118 инженерных аминокислот были помещены в белок
Эти группы пересекаются, но не идентичны. Все 22 протеиногенные аминокислоты биосинтезируются организмами, а некоторые, но не все, из них также являются абиотическими (обнаружены в пребиотических экспериментах и метеоритах). Некоторые природные аминокислоты, такие как норлейцин , неправильно включаются в белки трансляционно из-за неправильного процесса синтеза белка. Многие аминокислоты, такие как орнитин , являются промежуточными продуктами метаболизма, вырабатываемыми биосинтетически, но не включающимися трансляционно в белки. Посттрансляционная модификация аминокислотных остатков в белках приводит к образованию многих белковых, но непротеиногенных аминокислот. Другие аминокислоты встречаются исключительно в абиотических смесях (например, α-метилнорвалин). Более 30 неприродных аминокислот были вставлены трансляционно в белок в инженерных системах, но не являются биосинтетическими. [7]
Номенклатура
[ редактировать ]В дополнение к системе нумерации ИЮПАК, позволяющей различать различные атомы углерода в органической молекуле, путем последовательного присвоения номера каждому углероду, включая те, которые образуют карбоксильную группу, атомы углерода вдоль боковой цепи аминокислот также могут быть помечены греческими буквами. , где α-углерод представляет собой центральный хиральный углерод, обладающий карбоксильной группой, боковой цепью и, в α-аминокислотах, аминогруппой – углерод в карбоксильных группах не учитывается. [8] (Следовательно, названия многих непротеиногенных α-аминокислот в ИЮПАК начинаются с 2-амино- и заканчиваются на -ic кислоту .)
Природные не- L -α-аминокислоты
[ редактировать ]Большинство природных аминокислот представляют собой α-аминокислоты в L- конфигурации, но существуют и некоторые исключения.
Неальфа
[ редактировать ]
Некоторые не-α-аминокислоты существуют в организмах. В этих структурах аминогруппа смещена дальше от карбонового конца молекулы аминокислоты. Таким образом, у β-аминокислоты аминогруппа связана со вторым атомом углерода, а у γ-аминокислоты она находится на третьем атоме углерода. Примеры включают β-аланин , ГАМК и δ- аминолевулиновую кислоту .
![β-аланин: аминокислота, продуцируемая аспартат-1-декарбоксилазой, предшественник кофермента А[9] и пептидов карнозина и ансерина.](//upload.wikimedia.org/wikipedia/commons/thumb/3/35/Beta-alanine_structure.svg/120px-Beta-alanine_structure.svg.png) β-аланин: аминокислота, вырабатываемая аспартат-1-декарбоксилазой и предшественник кофермента А. [9] и пептиды карнозин и ансерин .
β-аланин: аминокислота, вырабатываемая аспартат-1-декарбоксилазой и предшественник кофермента А. [9] и пептиды карнозин и ансерин . γ-аминомасляная кислота (ГАМК): нейромедиатор у животных.
γ-аминомасляная кислота (ГАМК): нейромедиатор у животных. δ-аминолевулиновая кислота: промежуточный продукт биосинтеза тетрапирролов ( гема , хлорофилла , кобаламина и т. д.).
δ-аминолевулиновая кислота: промежуточный продукт биосинтеза тетрапирролов ( гема , хлорофилла , кобаламина и т. д.). 4-аминобензойная кислота (ПАБА): промежуточный продукт фолиевой кислоты. биосинтеза
4-аминобензойная кислота (ПАБА): промежуточный продукт фолиевой кислоты. биосинтеза
![β-аланин: аминокислота, продуцируемая аспартат-1-декарбоксилазой, предшественник кофермента А[9] и пептидов карнозина и ансерина.](http://upload.wikimedia.org/wikipedia/commons/thumb/3/35/Beta-alanine_structure.svg/120px-Beta-alanine_structure.svg.png)


Причина, по которой α-аминокислоты используются в белках, связана с их частотой в метеоритах и экспериментах с пребиотиками. [10] Первоначальные предположения о вредных свойствах β-аминокислот с точки зрения вторичной структуры. [10] оказалось неправильным. [11]
D-аминокислоты
[ редактировать ]Некоторые аминокислоты содержат противоположную абсолютную хиральность, химические вещества, которые недоступны в обычном рибосомальном аппарате трансляции и транскрипции. Стенки большинства бактериальных клеток образованы пептидогликаном , полимером, состоящим из аминосахаров, сшитых короткими олигопептидами, соединенными между собой мостиками. Олигопептид синтезируется нерибосомально и содержит несколько особенностей, включая D-аминокислоты , обычно D -аланин и D -глутамат. Еще одной особенностью является то, что первый рацемизируется PLP -связывающими ферментами (кодируемыми alr или гомологом dadX ), тогда как последний рацемизируется независимым от кофактора ферментом ( murI ). Некоторые варианты присутствуют у Thermotoga spp. D Присутствует -лизин, а в некоторых устойчивых к ванкомицину бактериях D присутствует vanT ). -серин ( ген [12] [13]
Без водорода на α-углероде
[ редактировать ]Все протеиногенные аминокислоты имеют по крайней мере один водород на α-углероде. У глицина два атома водорода, а у всех остальных — один водород и одна боковая цепь. Замена оставшегося водорода более крупным заместителем, например метильной группой, искажает основную цепь белка. [10]
У некоторых грибов α-аминоизомасляная кислота вырабатывается как предшественник пептидов, некоторые из которых проявляют антибиотические свойства. [14] Это соединение похоже на аланин, но имеет дополнительную метильную группу на α-углероде вместо водорода. Следовательно, оно ахирально. Другое соединение, похожее на аланин без α-водорода, — это дегидроаланин , имеющий метиленовую боковую цепь. Это одна из нескольких встречающихся в природе дегидроаминокислот .
 аланин
аланин аминоизомасляная кислота
аминоизомасляная кислота дегидроаланин
дегидроаланин



Двойные аминокислотные стереоцентры
[ редактировать ]Подмножество L -α-аминокислот неоднозначно относительно того, какой из двух концов является α-углеродом. В белках остаток цистеина может образовывать дисульфидную связь с другим остатком цистеина, тем самым сшивая белок. Два сшитых цистеина образуют молекулу цистина . Цистеин и метионин обычно производятся путем прямого сульфурилирования, но у некоторых видов они могут быть получены путем транссульфурации , когда активированный гомосерин или серин сплавляется с цистеином или гомоцистеином, образуя цистатионин . Аналогичным соединением является лантионин , который можно рассматривать как две молекулы аланина, соединенные тиоэфирной связью, и он встречается в различных организмах. Аналогично, джэнколевая кислота , растительный токсин из бобов дженгколь , состоит из двух цистеинов, соединенных метиленовой группой. Диаминопимелиновая кислота используется как мостик в пептидогликане, так и в качестве предшественника лизина (посредством его декарбоксилирования).
 цистин
цистин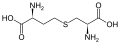 цистатионин
цистатионин лантионин
лантионин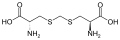 дженколевая кислота
дженколевая кислота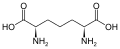 диаминопимелиновая кислота
диаминопимелиновая кислота
Пребиотические аминокислоты и альтернативная биохимия
[ редактировать ]В метеоритах и в экспериментах с пребиотиками (например, эксперимент Миллера-Юри ) обнаруживается гораздо больше аминокислот, чем двадцать стандартных аминокислот, некоторые из которых находятся в более высоких концентрациях, чем стандартные. Было высказано предположение, что если бы жизнь, основанная на аминокислотах, возникла где-то во Вселенной, не более 75% аминокислот были бы общими. [10] Наиболее заметной аномалией является отсутствие аминомасляной кислоты.
| Молекула | Электрический разряд | Мерчинсонский метеорит |
|---|---|---|
| глицин | 100 | 100 |
| аланин | 180 | 36 |
| α-амино- н -масляная кислота | 61 | 19 |
| норвалин | 14 | 14 |
| валин | 4.4 | |
| норлейцин | 1.4 | |
| лейцин | 2.6 | |
| изолейцин | 1.1 | |
| аллоизолейцин | 1.2 | |
| т-лейцин | < 0,005 | |
| α-амино- н -гептановая кислота | 0.3 | |
| пролин | 0.3 | 22 |
| пипеколиновая кислота | 0.01 | 11 |
| α,β-диаминопропионовая кислота | 1.5 | |
| α,γ-диаминомасляная кислота | 7.6 | |
| орнитин | < 0,01 | |
| лизин | < 0,01 | |
| аспарагиновая кислота | 7.7 | 13 |
| глутаминовая кислота | 1.7 | 20 |
| серин | 1.1 | |
| треонин | 0.2 | |
| аллотреонин | 0.2 | |
| метионин | 0.1 | |
| гомоцистеин | 0.5 | |
| гомосерин | 0.5 | |
| β-аланин | 4.3 | 10 |
| β-амино- н -масляная кислота | 0.1 | 5 |
| β-аминоизомасляная кислота | 0.5 | 7 |
| γ-аминомасляная кислота | 0.5 | 7 |
| α-аминоизомасляная кислота | 7 | 33 |
| изовалин | 1 | 11 |
| саркозин | 12.5 | 7 |
| N -этилглицин | 6.8 | 6 |
| N- пропилглицин | 0.5 | |
| N -изопропилглицин | 0.5 | |
| N -метилаланин | 3.4 | 3 |
| N -этилаланин | < 0,05 | |
| N -метил-β-аланин | 1.0 | |
| N -этил-β-аланин | < 0,05 | |
| изосерин | 1.2 | |
| α-гидрокси-γ-аминомасляная кислота | 17 |
Прямая боковая цепь
[ редактировать ]Генетический код был описан как замороженная случайность, и причиной того, что существует только одна стандартная аминокислота с прямой цепью, аланином, может быть просто избыточность валина, лейцина и изолейцина. [10] Однако сообщается, что аминокислоты с прямой цепью образуют гораздо более стабильные альфа-спирали. [15]
 глицин (водородная боковая цепь)
глицин (водородная боковая цепь)- аланин (метильная боковая цепь)
 гомоаланин или α-аминомасляная кислота (этильная боковая цепь)
гомоаланин или α-аминомасляная кислота (этильная боковая цепь) норвалин ( н -пропильная боковая цепь)
норвалин ( н -пропильная боковая цепь) норлейцин ( н- бутильная боковая цепь)
норлейцин ( н- бутильная боковая цепь) гомонорлейцин ( н -пентила, гептановая кислота) показана боковая цепь
гомонорлейцин ( н -пентила, гептановая кислота) показана боковая цепь




халькоген
[ редактировать ]Серин, гомосерин , О -метилгомосерин и О -этилгомосерин обладают гидроксиметильной, гидроксиэтильной, О -метилгидроксиметильной и О- метилгидроксиэтильной боковой цепью; тогда как цистеин, гомоцистеин , метионин и этионин обладают тиоловыми эквивалентами. Эквивалентами селенола являются селеноцистеин, селеногомоцистеин, селенометионин и селенэтионин. Аминокислоты со следующим халькогеном встречаются и в природе: некоторые виды, такие как Aspergillus fumigatus , Aspergillus terreus и Penicillium chrysogenum, в отсутствие серы способны продуцировать и включаться в белок теллуроцистеин и теллурометионин. [16]
Расширенный генетический код
[ редактировать ]Роли
[ редактировать ]В клетках, особенно автотрофных, в качестве промежуточных продуктов метаболизма обнаруживаются некоторые непротеиногенные аминокислоты. Однако, несмотря на каталитическую гибкость PLP-связывающих ферментов, многие аминокислоты синтезируются в виде кетокислот (например, 4-метил-2-оксопентаноат в лейцин) и аминируются на последней стадии, таким образом сохраняя количество непротеиногенных аминокислот. промежуточные продукты довольно низкие.
Орнитин и цитруллин участвуют в цикле мочевины аминокислот , являясь частью катаболизма (см. ниже). [17]
Помимо первичного метаболизма , некоторые непротеиногенные аминокислоты являются предшественниками или конечными продуктами вторичного метаболизма для образования небольших соединений или нерибосомальных пептидов (например, некоторых токсинов ).
Посттрансляционно включается в белок
[ редактировать ]Несмотря на то, что некоторые нестандартные аминокислоты не кодируются генетическим кодом как протеиногенные аминокислоты, тем не менее они обнаруживаются в белках. Они образуются путем посттрансляционной модификации боковых цепей стандартных аминокислот, присутствующих в целевом белке. Эти модификации часто необходимы для функционирования или регуляции белка; например, в γ-карбоксиглутамате карбоксилирование связывать глутамата кальция позволяет лучше катионы , [18] в гидроксипролине гидроксилирование пролина а тканей имеет решающее значение для поддержания соединительных . [19] Другим примером является образование гипузина в факторе инициации трансляции EIF5A посредством модификации остатка лизина. [20] Такие модификации могут также определять локализацию белка, например, добавление длинных гидрофобных групп может привести к связыванию белка с фосфолипидной мембраной. [21]
 Карбоксиглутаминовая кислота. В то время как глутаминовая кислота имеет одну γ-карбоксильную группу, карбоксиглутаминовая кислота имеет две.
Карбоксиглутаминовая кислота. В то время как глутаминовая кислота имеет одну γ-карбоксильную группу, карбоксиглутаминовая кислота имеет две. Гидроксипролин. Эта иминокислота отличается от пролина наличием гидроксильной группы у углерода 4.
Гидроксипролин. Эта иминокислота отличается от пролина наличием гидроксильной группы у углерода 4.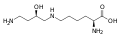 Гипузин . Эту аминокислоту получают добавлением к ε-аминогруппе лизина 4-аминобутильного фрагмента (полученного из спермидина ).
Гипузин . Эту аминокислоту получают добавлением к ε-аминогруппе лизина 4-аминобутильного фрагмента (полученного из спермидина ).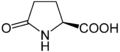



Имеются некоторые предварительные доказательства того, что аминомалоновая кислота может присутствовать в белке, возможно, в результате неправильного включения. [22] [23]
Токсичные аналоги
[ редактировать ]Некоторые непротеиногенные аминокислоты токсичны из-за их способности имитировать определенные свойства протеиногенных аминокислот, таких как тиализин . Некоторые непротеиногенные аминокислоты нейротоксичны, имитируя аминокислоты, используемые в качестве нейротрансмиттеров (то есть не участвующих в биосинтезе белка), включая кискваловую кислоту , канаванин и азетидин-2-карбоновую кислоту . [24] Цефалоспорин C имеет основу α-аминоадипиновой кислоты (гомоглутамата), амидированную цефалоспориновым фрагментом. [25] Пеницилламин — терапевтическая аминокислота, механизм действия которой неизвестен.
 Тиализин
Тиализин любая кислота
любая кислота канаванин
канаванин азетидин-2-карбоновая кислота
азетидин-2-карбоновая кислота цефалоспорин С
цефалоспорин С пеницилламин
пеницилламин



Природные цианотоксины также могут включать непротеиногенные аминокислоты. микроцистин и нодуларин Например, являются производными ADDA , β-аминокислоты.
Не аминокислоты
[ редактировать ]Таурин представляет собой аминосульфоновую кислоту , а не аминокарбоновую кислоту, однако иногда его считают таковым, поскольку количества, необходимые для подавления ауксотрофа у некоторых организмов (например, у кошек), ближе к количествам «незаменимых аминокислот» (аминокислотная ауксотрофия). ), чем витаминов (кофакторная ауксотрофия).
Осмолиты, саркозин и глицин-бетаин происходят из аминокислот, но содержат вторичный и четвертичный амин соответственно.
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ плюс формилметионин у эукариот с прокариотными органеллами, такими как митохондрии
Ссылки
[ редактировать ]- ^ Амброгелли, А.; Палиура, С.; Зёлль, Д. (2007). «Естественное расширение генетического кода». Химическая биология природы . 3 (1): 29–35. дои : 10.1038/nchembio847 . ПМИД 17173027 .
- ^ Аван, Илькер; Холл, К. Деннис; Катрицки, Алан Р. (22 апреля 2014 г.). «Пептидомиметика путем модификации аминокислот и пептидных связей» . Обзоры химического общества . 43 (10): 3575–3594. дои : 10.1039/C3CS60384A . ПМИД 24626261 .
- ^ Сараса, Сабна Б.; Махендран, Рамасами; Мутхусами, Гаятри; Сакаппан, Бенси; Сельта, Дэниел Раджа Фемил; Ангаярканни, Джаяраман (2020). «Краткий обзор небелковой аминокислоты гамма-аминомасляной кислоты (ГАМК): ее производство и роль в микробах». Современная микробиология . 77 (4): 534–544. дои : 10.1007/s00284-019-01839-w . ПМИД 31844936 .
- ^ Остойич, Сергей М. (01 августа 2021 г.). «Креатин как пищевая добавка для населения» . Журнал функциональных продуктов питания . 83 : 104568. doi : 10.1016/j.jff.2021.104568 . ISSN 1756-4646 .
- ^ Бёк, А.; Форчхаммер, К.; Хайдер, Дж.; Барон, К. (1991). «Синтез селенопротеина: расширение генетического кода». Тенденции биохимических наук . 16 (12): 463–467. дои : 10.1016/0968-0004(91)90180-4 . ПМИД 1838215 .
- ^ Теобальд-Дитрих, А.; Жье, Р.; Рюдингер-Тирион, Дж.Л. (2005). «Доказательства существования в мРНК шпильечного элемента, ответственного за рибосомно-зависимую вставку пирролизина в белки». Биохимия . 87 (9–10): 813–817. дои : 10.1016/j.biochi.2005.03.006 . ПМИД 16164991 .
- ^ Jump up to: а б Лу, Ю.; Фриланд, С. (2006). «Об эволюции стандартного аминокислотного алфавита» . Геномная биология . 7 (1): 102. doi : 10.1186/gb-2006-7-1-102 . ПМК 1431706 . ПМИД 16515719 .
- ^ Фут, Д.; Фут, Дж. Г. (2004). Биохимия (3-е изд.). Джон Уайли и сыновья. ISBN 978-0471193500 .
- ^ Чакауя, Э.; Коксон, КМ; Оттенхоф, ХХ; Уитни, HM; Бланделл, ТЛ; Абель, К.; Смит, АГ (2005). «Биосинтез пантотената у высших растений». Труды Биохимического общества . 33 (4): 743–746. дои : 10.1042/BST0330743 . ПМИД 16042590 .
- ^ Jump up to: а б с д и Вебер, Алабама; Миллер, С.Л. (1981). «Причины появления двадцати кодируемых в белке аминокислот». Журнал молекулярной эволюции . 17 (5): 273–284. Бибкод : 1981JMolE..17..273W . дои : 10.1007/BF01795749 . ПМИД 7277510 . S2CID 27957755 .
- ^ Койак, MJ; Ченг, Р.П. (2006). «Дизайн и синтез β-пептидов с биологической активностью». Белковый дизайн . Методы молекулярной биологии. Том. 340. стр. 95–109. дои : 10.1385/1-59745-116-9:95 . ISBN 978-1-59745-116-1 . ПМИД 16957334 .
- ^ Бонифаций, А.; Паркет, К.; Артур, М.; Менжен-Лекрель, Д.; Бланот, Д. (2009). «Выяснение структуры пептидогликана Thermotoga maritima выявило два новых типа поперечных связей» . Журнал биологической химии . 284 (33): 21856–21862. дои : 10.1074/jbc.M109.034363 . ПМЦ 2755910 . ПМИД 19542229 .
- ^ Ариас, Калифорния; Мартин-Мартинес, М.; Бланделл, ТЛ; Артур, М.; Курвален, П.; Рейнольдс, ЧП (1999). «Характеристика и моделирование vanT: новая мембраносвязанная сериновая рацемаза из устойчивого к ванкомицину Enterococcus Gallinarum BM4174» . Молекулярная микробиология . 31 (6): 1653–1664. дои : 10.1046/j.1365-2958.1999.01294.x . ПМИД 10209740 . S2CID 25796469 .
- ^ Гао, X.; Чуй, Ю.Х.; Эймс, Б.Д.; Ван, П.; Уолш, Коннектикут; Тан, Ю. (2011). «Биосинтез грибковых индольных алкалоидов: генетическое и биохимическое исследование триптоквиаланинового пути у Penicillium aethiopicum » . Журнал Американского химического общества . 133 (8): 2729–2741. дои : 10.1021/ja1101085 . ПМК 3045477 . ПМИД 21299212 .
- ^ Падманабхан, С.; Болдуин, Р.Л. (1991). «Неполярные аминокислоты с прямой цепью являются хорошими спиралеобразователями в воде» . Журнал молекулярной биологии . 219 (2): 135–137. дои : 10.1016/0022-2836(91)90553-I . ПМИД 2038048 .
- ^ Рамадан, ЮВ; Разак, А.А.; Рагаб, AM; Эль-Мелиджи, М. (1989). «Включение теллура в аминокислоты и белки у устойчивых к теллуру грибов». Исследование биологических микроэлементов . 20 (3): 225–232. дои : 10.1007/BF02917437 . ПМИД 2484755 . S2CID 9439946 .
- ^ Курис, Э.; Николис, И.; Муанар, К.; Осовска, С.; Зеррук, Н.; Бенасет, С.; Кинобер, Л. (2005). «Почти все о цитруллине у млекопитающих». Аминокислоты . 29 (3): 177–205. дои : 10.1007/s00726-005-0235-4 . ПМИД 16082501 . S2CID 23877884 .
- ^ Вермеер, К. (1990). «Гамма-карбоксиглутаматсодержащие белки и витамин К-зависимая карбоксилаза» . Биохимический журнал . 266 (3): 625–636. дои : 10.1042/bj2660625 . ПМЦ 1131186 . ПМИД 2183788 .
- ^ Бхаттачарджи, А; Бансал, М. (2005). «Структура коллагена: тройная спираль Мадраса и текущий сценарий» . ИУБМБ Жизнь . 57 (3): 161–172. дои : 10.1080/15216540500090710 . ПМИД 16036578 . S2CID 7211864 .
- ^ Парк, Миннесота (2006). «Посттрансляционный синтез аминокислоты, полученной из полиамина, гипузина, в эукариотическом факторе инициации трансляции 5А (eIF5A)» . Журнал биохимии . 139 (2): 161–169. дои : 10.1093/jb/mvj034 . ПМК 2494880 . ПМИД 16452303 .
- ^ Бленис, Дж; Реш, доктор медицины (1993). «Субклеточная локализация, определяемая ацилированием и фосфорилированием белков». Современное мнение в области клеточной биологии . 5 (6): 984–989. дои : 10.1016/0955-0674(93)90081-з . ПМИД 8129952 .
- ^ Копли, С.Д.; Франк, Э.; Кирш, В.М.; Кох, TH (1992). «Обнаружение и возможное происхождение аминомалоновой кислоты в белковых гидролизатах». Аналитическая биохимия . 201 (1): 152–157. дои : 10.1016/0003-2697(92)90188-D . ПМИД 1621954 .
- ^ Ван Бускирк, Джей-Джей; Кирш, В.М.; Клейер, Д.Л.; Баркли, РМ; Кох, TH (1984). «Аминомалоновая кислота: Идентификация в Escherichia coli и атеросклеротических бляшках» . Труды Национальной академии наук . 81 (3): 722–725. Бибкод : 1984PNAS...81..722V . дои : 10.1073/pnas.81.3.722 . ПМК 344907 . ПМИД 6366787 .
- ^ Дасури, К.; Эбенезер, П.Дж.; Уранга, РМ; Гавилан, Э.; Чжан, Л.; Фернандес-Ким, СОК; Брюс-Келлер, AJ; Келлер, Дж. Н. (2011). «Токсичность аналогов аминокислот в первичных культурах нейронов и астроцитов крыс: последствия для неправильного сворачивания белка и регуляции TDP-43» . Журнал нейробиологических исследований . 89 (9): 1471–1477. дои : 10.1002/jnr.22677 . ПМК 3175609 . ПМИД 21608013 .
- ^ Траун, П.В.; Смит, Б.; Авраам, EP (1963). «Биосинтез цефалоспорина С из аминокислот» . Биохимический журнал . 86 (2): 284–291. дои : 10.1042/bj0860284 . ПМК 1201751 . ПМИД 13994319 .