Раковые стволовые клетки
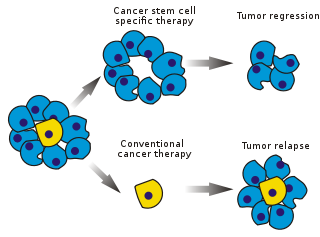
Раковые стволовые клетки ( РСК ) — это раковые клетки (обнаруженные в опухолях или гематологических раковых клетках ), которые обладают характеристиками, присущими нормальным стволовым клеткам , в частности способностью давать начало всем типам клеток, обнаруженным в конкретном раковом образце. Таким образом, РСК являются онкогенными (опухолеобразующими), возможно, в отличие от других неопухолевых раковых клеток. [1] РСК могут генерировать опухоли посредством процессов самообновления и дифференцировки стволовых клеток в несколько типов клеток. Предполагается, что такие клетки персистируют в опухолях как отдельная популяция и вызывают рецидив и метастазирование, порождая новые опухоли. Таким образом, разработка специфических методов лечения, нацеленных на РСК, дает надежду на улучшение выживаемости и качества жизни онкологических больных, особенно пациентов с метастатическим заболеванием .
Существующие методы лечения рака в основном были разработаны на основе животных моделей , где методы лечения, способные способствовать уменьшению опухоли, были признаны эффективными. Однако животные не обеспечивают полную модель заболеваний человека. В частности, у мышей, продолжительность жизни которых не превышает двух лет, рецидив опухоли трудно поддается изучению.
Эффективность лечения рака на начальных этапах тестирования часто измеряется фракцией абляции опухолевой массы ( фракционное уничтожение ). Поскольку РСК составляют небольшую часть опухоли, не обязательно выбирать препараты, действующие конкретно на стволовые клетки. Теория предполагает, что традиционная химиотерапия убивает дифференцированные или дифференцирующиеся клетки, которые составляют основную часть опухоли, но не производят новых клеток. Популяция РСК, породившая его, могла остаться нетронутой и вызвать рецидив.
Раковые стволовые клетки были впервые идентифицированы Джоном Диком при остром миелоидном лейкозе в конце 1990-х годов. С начала 2000-х годов они были объектом интенсивного исследования рака . [2] Сам термин был придуман в широко цитируемой статье в 2001 году биологами Танништой Рейей , Шоном Дж. Моррисоном , Майклом Ф. Кларком и Ирвингом Вайсманом . [3]
опухоли распространения Модели
При различных подтипах опухолей клетки опухолевой популяции проявляют функциональную гетерогенность , и опухоли образуются из клеток с различной пролиферативной и дифференцировочной способностью. [4] Эта функциональная гетерогенность раковых клеток привела к созданию множества моделей размножения, объясняющих гетерогенность и различия в регенеративной способности опухолей: раковые стволовые клетки (РСК) и стохастическая модель. Однако некоторые точки зрения утверждают, что это разграничение является искусственным, поскольку оба процесса действуют взаимодополняющим образом в отношении реальных популяций опухолей. [1] Важно отметить, что в то время как в эпителии пищевода здорового человека пролиферативную нагрузку встречает стохастически делящийся базальный эпителий. Однако при его переходе в эпителий предракового пищевода Барретта появляется небольшой выделенный компартмент стволовых клеток, который поддерживает пролиферацию эпителия, в то время как одновременно исчезают доказательства стохастически делящегося компартмента, способствующего поддержанию ткани. Следовательно, по крайней мере для некоторых неопластических тканей, выделенные компартменты стволовых клеток поддерживают и увеличивают размер трансформированного компартмента. [5]


Модель раковых клеток стволовых
Модель раковых стволовых клеток, также известная как иерархическая модель, предполагает, что опухоли организованы иерархически (РСК лежат на вершине [6] (Рис. 3).) В раковой популяции опухолей имеются раковые стволовые клетки (РСК), которые являются онкогенными клетками и биологически отличаются от других субпопуляций. [7] У них есть две определяющие особенности: их долговременная способность к самообновлению и их способность дифференцироваться в потомство, которое не является канцерогенным, но все же способствует росту опухоли. Эта модель предполагает, что только определенные субпопуляции раковых стволовых клеток обладают способностью стимулировать прогрессирование рака, а это означает, что существуют специфические (внутренние) характеристики, которые можно идентифицировать и затем нацелить на долгосрочное уничтожение опухоли без необходимости борьбы с целая опухоль. [8]
Стохастическая модель [ править ]
Чтобы клетка стала раковой, она должна претерпеть значительное количество изменений в последовательности ДНК. Эта клеточная модель предполагает, что эти мутации могут возникнуть в любой клетке организма, что приведет к раку. По сути, эта теория предполагает, что все клетки обладают способностью быть онкогенными, что делает все опухолевые клетки равносильными по способности к самообновлению или дифференцировке, что приводит к гетерогенности опухоли, в то время как другие могут дифференцироваться в не-РСК. [7] [9] На потенциал клетки могут влиять непредсказуемые генетические или эпигенетические факторы, что приводит к образованию фенотипически разнообразных клеток как в онкогенных, так и в неопухолевых клетках, составляющих опухоль. Согласно «стохастической модели» (или «модели клональной эволюции») каждая раковая клетка в опухоли может обрести способность самообновляться и дифференцироваться в многочисленные и гетерогенные линии раковых клеток, которые поражают опухоль. [10]
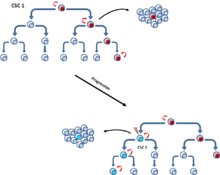
Эти мутации могут постепенно накапливаться и повышать устойчивость и приспособленность клеток, что позволит им вытеснять другие опухолевые клетки, более известную как модель соматической эволюции . [7] Модель клональной эволюции, которая встречается как в модели CSC, так и в стохастической модели, постулирует, что мутантные опухолевые клетки с преимуществом в росте превосходят другие. Клетки доминирующей популяции обладают сходным потенциалом инициации роста опухоли. [11] (рис. 4).

[12] Эти две модели не являются взаимоисключающими, поскольку сами РСК подвергаются клональной эволюции. Таким образом, могут возникнуть вторичные, более доминантные РСК, если мутация придает более агрессивные свойства. [13] (рис. 5).
CSC и стохастических вместе Связывание моделей
Исследование 2014 года утверждает, что разрыв между этими двумя противоречивыми моделями можно преодолеть, предоставив альтернативное объяснение гетерогенности опухолей. Они демонстрируют модель, которая включает в себя аспекты моделей «Dreamy» и BULL CSC. [9] Они исследовали пластичность раковых стволовых клеток , при которой раковые стволовые клетки могут переходить между нераковыми стволовыми клетками (Non-CSC) и CSC посредством in situ, поддерживая более стохастическую модель. [9] [14] Но существование биологически различных популяций, не относящихся к РСК, и РСК подтверждает модель, в большей степени РСК, предполагающую, что обе модели могут играть жизненно важную роль в гетерогенности опухолей. [9]
Модель иммунологии раковых клеток стволовых
Эта модель предполагает, что иммунологические свойства могут быть важны для понимания онкогенеза и гетерогенности. Таким образом, в некоторых опухолях ОСК могут встречаться очень редко. [15] но некоторые исследователи обнаружили, что большая часть опухолевых клеток может инициировать опухоли, если их трансплантировать мышам с тяжелым иммунодефицитом. [16] и, таким образом, поставил под сомнение актуальность редких CSC. Однако обе стволовые клетки [17] и CSC [18] обладают уникальными иммунологическими свойствами, которые делают их высокоустойчивыми к иммунонадзору. Таким образом, только РСК могут быть способны вызывать опухоли у пациентов с функциональным иммунонадзором, а иммунная привилегия может быть ключевым критерием для идентификации РСК. [19] Более того, модель предполагает, что ОСК могут изначально зависеть от ниш стволовых клеток и функционировать там как резервуар, в котором мутации могут накапливаться в течение десятилетий, не ограничиваясь иммунной системой. Клинически явные опухоли могут расти, если:А) РСК теряют зависимость от нишевых факторов (менее дифференцированные опухоли),Б) их потомки из высокопролиферативных, но изначально иммуногенных нормальных опухолевых клеток развивают средства, позволяющие избежать иммунонадзора или В) иммунная система может потерять способность подавлять опухоли, например, из-за старения. [19]
Дебаты [ править ]
Существование РСК является предметом дискуссий, поскольку во многих исследованиях не было обнаружено клеток с их специфическими характеристиками. [15] Раковые клетки должны быть способны к непрерывной пролиферации и самообновлению, чтобы сохранять множество мутаций, необходимых для канцерогенеза , и поддерживать рост опухоли, поскольку дифференцированные клетки (ограниченные пределом Хейфлика) [20] ) не может делиться бесконечно. С терапевтической точки зрения, если большинство опухолевых клеток наделены свойствами стволовых клеток, прямое воздействие на размер опухоли является действенной стратегией. Если CSC составляют небольшое меньшинство, борьба с ними может быть более эффективной. Другая дискуссия ведется по поводу происхождения ОСК — от нарушения регуляции нормальных стволовых клеток или от более специализированной популяции, которая приобрела способность к самообновлению (что связано с вопросом пластичности стволовых клеток ). Эту дискуссию усложняет открытие того, что многие раковые клетки демонстрируют фенотипическую пластичность под воздействием терапевтического воздействия, изменяя свои транскриптомы в более стеблеподобное состояние, чтобы избежать разрушения. [ нужна ссылка ]
Доказательства [ править ]
Первые убедительные доказательства существования РСК были получены в 1997 году. Боннет и Дик выделили субпопуляцию лейкозных клеток, которые экспрессировали поверхностный маркер CD34 , но не CD38 . [21] Авторы установили, что CD34 + /CD38 − субпопуляция способна инициировать опухоли у мышей NOD/ SCID , гистологически сходных с донором. Первые доказательства наличия стволоподобных клеток рака солидной опухоли последовали в 2002 году, когда была открыта клоногенная сферообразующая клетка, выделенная и охарактеризованная из глиомы головного мозга взрослого человека . опухоли человека Кортикальные глиальные содержат нервные стволоподобные клетки, экспрессирующие астроглиальные и нейрональные маркеры in vitro . [22] Было показано, что раковые стволовые клетки, выделенные из глиом взрослого человека, индуцируют опухоли, которые напоминали родительскую опухоль, при трансплантации на внутричерепные модели голых мышей. [23]
В экспериментах по исследованию рака опухолевые клетки иногда вводят подопытным животным, чтобы образовать опухоль. Затем во времени отслеживается прогрессирование заболевания, и новые лекарства могут быть проверены на предмет их эффективности. Для формирования опухоли требуется введение тысяч или десятков тысяч клеток. Классически это объяснялось плохой методологией (т.е. опухолевые клетки теряют жизнеспособность во время переноса) или критической важностью микроокружения, особого биохимического окружения инъецированных клеток. Сторонники парадигмы РСК утверждают, что только небольшая часть введенных клеток, РСК, потенциально способна вызвать опухоль. При остром миелолейкозе человека частота этих клеток составляет менее 1 на 10 000. [21]
Дальнейшие доказательства получены из гистологии . Многие опухоли гетерогенны и содержат несколько типов клеток, присущих органу-хозяину. Гетерогенность опухоли обычно сохраняется при метастазах опухоли . Это говорит о том, что клетка, которая их производила, обладала способностью генерировать несколько типов клеток, что является классической отличительной чертой стволовых клеток . [21]
Существование стволовых клеток лейкемии побудило к исследованию других видов рака. РСК недавно были идентифицированы в нескольких солидных опухолях, в том числе:
- Мозг [24]
- Грудь [25]
- Двоеточие [26]
- яичник [27] [28]
- поджелудочная железа [29]
- Простата [30] [31]
- Меланома [32] [33] [34] [35]
- Множественная миелома [36] [37]
- Немеланомный рак кожи (базальноклеточная карцинома [38] и плоскоклеточный рак [39] [40] )
Механистические и математические модели [ править ]
Как только выдвинута гипотеза о путях развития рака, можно разработать прогностические математические модели. [41] например, на основе метода клеточного компартмента . Например, рост аномальных клеток можно обозначить определенной вероятностью мутации. Такая модель предсказала, что повторное повреждение зрелых клеток увеличивает образование аномального потомства и риск рака. [42] Клиническая эффективность таких моделей [43] остается неустановленным.
Происхождение [ править ]

Происхождение РСК является активной областью исследований. Ответ может зависеть от типа и фенотипа опухоли . До сих пор гипотеза о том, что опухоли возникают из одной «клетки происхождения», не была продемонстрирована с использованием модели раковых стволовых клеток. Это связано с тем, что раковые стволовые клетки не присутствуют в опухолях терминальной стадии.
Гипотезы происхождения включают мутанты в развивающихся стволовых клетках или клетках-предшественниках, мутанты во взрослых стволовых клетках или взрослых клетках-предшественниках и мутантные дифференцированные клетки, которые приобретают свойства, подобные стволовым. Эти теории часто фокусируются на «клетке происхождения» опухоли.
Гипотезы [ править ]
клеток Мутация стволовых
Гипотеза «мутации в популяциях ниш стволовых клеток во время развития» утверждает, что эти развивающиеся популяции стволовых клеток мутируют, а затем размножаются, так что мутация передается многим потомкам. Эти дочерние клетки гораздо ближе к превращению в опухоли, и их количество увеличивает вероятность раковой мутации. [44]
стволовые Взрослые клетки
Другая теория связывает взрослый ствол (ASC) с образованием опухоли. Чаще всего это связано с тканями с высокой скоростью обновления клеток (например, с кожей или кишечником ). В этих тканях ASC являются кандидатами из-за их частых клеточных делений (по сравнению с большинством ASC) в сочетании с длительной продолжительностью жизни ASC. Эта комбинация создает идеальные условия для накопления мутаций: накопление мутаций является основным фактором, вызывающим возникновение рака . Данные показывают, что эта связь представляет собой реальное явление, хотя конкретные виды рака связаны с конкретной причиной. [45] [46]
Дедифференциация [ править ]
Дедифференцировка мутировавших клеток может создать характеристики, подобные стволовым клеткам, что позволяет предположить, что любая клетка может стать раковой стволовой клеткой. Другими словами, полностью дифференцированная клетка подвергается мутациям или внеклеточным сигналам, которые возвращают ее в состояние, подобное стволу. Эта концепция была совсем недавно продемонстрирована на рака простаты моделях , где клетки, подвергающиеся андрогенной депривационной терапии, по-видимому, временно изменяют свой транскриптом на транскриптом стволоподобных клеток нервного гребня с инвазивными и мультипотентными свойствами этого класса стволоподобных клеток. [ нужна ссылка ]
Иерархия [ править ]
Концепция иерархии опухолей утверждает, что опухоль представляет собой гетерогенную популяцию мутантных клеток, каждая из которых имеет некоторые общие мутации, но различается по специфическому фенотипу . Опухоль содержит несколько типов стволовых клеток, один из которых оптимален для конкретной среды, а другие менее успешные линии. Эти вторичные линии могут быть более успешными в других средах, позволяя опухоли адаптироваться, в том числе к терапевтическому вмешательству. Если это правда, эта концепция влияет на режимы лечения, специфичные для раковых стволовых клеток. [47] Такая иерархия усложнила бы попытки точно определить происхождение.
Идентификация [ править ]
ОСК, о которых сейчас сообщается в большинстве опухолей человека, обычно идентифицируются и обогащаются с использованием стратегий идентификации нормальных стволовых клеток, которые аналогичны во всех исследованиях. [48] Эти процедуры включают сортировку клеток, активируемую флуоресценцией (FACS), с использованием антител, направленных на маркеры клеточной поверхности, и функциональные подходы, включая анализ боковой популяции или анализ Aldefluor. [49] Результат, обогащенный РСК, затем в различных дозах имплантируют мышам с иммунодефицитом для оценки способности к развитию опухоли. Этот анализ in vivo называется анализом предельного разведения. Подмножества опухолевых клеток, которые могут инициировать развитие опухоли при небольшом количестве клеток, дополнительно проверяются на способность к самообновлению в серийных исследованиях опухолей. [50]
ОСК также можно идентифицировать по оттоку включенных красителей Hoechst через множественной лекарственной устойчивости (MDR) и АТФ-связывающей кассеты (ABC) транспортеры . [49]
Другой подход - сферообразующие анализы. Многие нормальные стволовые клетки, такие как гемопоэтические или стволовые клетки тканей , в особых условиях культивирования образуют трехмерные сферы, которые могут дифференцироваться. Как и нормальные стволовые клетки, ОСК, выделенные из головного мозга или опухолей предстательной железы , также обладают способностью образовывать независимые от якоря сферы. [51]
В последние годы появились генетические подходы к выявлению раковых стволовых клеток у экспериментальных грызунов. В таких исследованиях после индукции рака (обычно посредством применения мутагенов) активируется генетическая кассета, что приводит к экспрессии легко идентифицируемого маркера, например зеленого флуоресцентного белка (GFP). Это преодолевает ограничения традиционных подходов (например, классическая техника мечения бромдезоксиуридином (BrdU) использовалась для идентификации клеток с медленным циклом у животных), поскольку генетические подходы не зависят от клеточного цикла и могут использоваться для импульсного мечения in vivo для идентификации покоящихся клеток. /медленноциклирующие клетки. [52] Эта стратегия, например, сыграла важную роль в идентификации так называемого компартмента Lgr5+ как компартмента раковых стволовых клеток при раке печени и продемонстрировала его потенциал в качестве жизнеспособной терапевтической мишени. [53]
Гетерогенность (маркеры) [ править ]
Гетерогенность РСК представляет собой пул дифференцированных и недифференцированных опухолевых клеток, который пополняется клетками, обладающими как опухолевыми, так и стволовыми клетками свойствами и имеющими фенотипическую и метаболическую гетерогенность внутри единой опухолевой массы. Существуют две теории, объясняющие фенотипическую и метаболическую гетерогенность РСК; Клональная изменчивость и теория раковых стволовых клеток. В то время как прежняя теория диктует роль генетической, эпигенетической и микросреды, в которой находятся опухолевые клетки, для приобретения недифференцированных онкогенных признаков. Последняя теория больше фокусируется на признаках злокачественности, приобретаемых стволовыми клетками, когда эти недифференцированные и высококанцерогенные стволовые клетки повторно заселяют дифференцированную опухолевую массу. [54]
РСК были идентифицированы в различных солидных опухолях . Обычно для выделения РСК из солидных и гематологических опухолей используют маркеры, специфичные для нормальных стволовых клеток. Маркеры, наиболее часто используемые для выделения РСК, включают: CD133 (также известный как PROM1 ), CD44 , ALDH1A1 , [55] CD34 , CD24 и EpCAM ( молекула адгезии эпителиальных клеток , также известная как эпителиально-специфичный антиген, ESA ). [56] [55]
CD133 (проминин 1) представляет собой с пятью трансмембранными доменами, гликопротеин экспрессируемый на CD34. + стволовые клетки и клетки-предшественники , эндотелиальные предшественники и фетальные нейральные стволовые клетки . Он был обнаружен с использованием гликозилированного эпитопа , известного как AC133.
EpCAM (молекула адгезии эпителиальных клеток, ESA, TROP1) представляет собой гемофильный Ca. 2+ -независимая молекула клеточной адгезии, экспрессирующаяся на базолатеральной поверхности большинства эпителиальных клеток .
CD90 (THY1) представляет собой гликозилфосфатидилинозитолгликопротеин , закрепленный в плазматической мембране и участвующий в передаче сигнала . Он также может опосредовать адгезию между тимоцитами и стромой тимуса.
CD44 (PGP1) представляет собой молекулу адгезии, которая играет плейотропную роль в передаче сигналов клеток, миграции и возвращении домой. Он имеет несколько изоформ, включая CD44H, который проявляет высокое сродство к гиалуронату , и CD44V, обладающий метастатическими свойствами.
CD24 (HSA) представляет собой гликозилированную гликозилфосфатидилинозитол-заякоренную молекулу адгезии, которая играет костимулирующую роль в В- и Т-клетках .
CD200 1 типа (OX-2) представляет собой мембранный гликопротеин , который доставляет ингибирующий сигнал иммунным клеткам, включая Т-клетки, естественные клетки-киллеры и макрофаги .
Рецептор лептина ( LEPR ) представляет собой цитокиновый рецептор I типа для гормона, ассоциированного с жиром, лептина , который играет важную роль в содействии злокачественному прогрессированию. [40]
АЛДГ — это повсеместно распространенное семейство ферментов альдегиддегидрогеназы , которое катализирует окисление ароматических альдегидов до карбоновых кислот . Например, он играет роль в превращении ретинола в ретиноевую кислоту , которая необходима для выживания. [57]
Первым солидным злокачественным новообразованием, из которого были выделены и идентифицированы ОСК, был рак молочной железы , и он наиболее интенсивно изучается. ОСК молочной железы обогащены CD44 + CD24 −/низкий , [25] СП [58] и АЛДГ + субпопуляции. [59] [60] ОСК молочной железы, по-видимому, фенотипически разнообразны. Экспрессия маркера РСК в клетках рака молочной железы, очевидно, гетерогенна, а популяции РСК молочной железы различаются в зависимости от опухоли. [61] Оба CD44 + CD24 − и CD44 + CD24 + клеточные популяции представляют собой клетки, инициирующие опухоль; однако РСК наиболее обогащены при использовании маркерного профиля CD44. + CD49f привет CD133/2 привет . [62]
ОСК были обнаружены при многих опухолях головного мозга. Стволоподобные опухолевые клетки были идентифицированы с использованием маркеров клеточной поверхности, включая CD133, [63] SSEA-1 (стадиоспецифический эмбриональный антиген-1), [64] РЭФР [65] и CD44. [66] Использование CD133 для идентификации стволоподобных клеток опухолей головного мозга может быть проблематичным, поскольку туморогенные клетки обнаруживаются как в CD133, так и в + и CD133 − клетки в некоторых глиомах и в некоторых CD133 + Опухолевые клетки головного мозга могут не обладать способностью инициировать опухоль. [65]
ОСК были зарегистрированы при раке толстой кишки человека . [26] Для их идентификации используются маркеры клеточной поверхности, такие как CD133, [26] CD44 [67] и ABCB5 , [68] функциональный анализ, включая клональный анализ [69] и анализ Aldefluor. [70] Использование CD133 в качестве положительного маркера ОСК толстой кишки привело к противоречивым результатам. Эпитоп AC133, но не белок CD133, специфически экспрессируется в РСК толстой кишки, и его экспрессия теряется при дифференцировке. [71] Кроме того, CD44 + клетки рака толстой кишки и дополнительное субфракционирование CD44 + ЭпКАМ + популяция клеток с CD166 повышает успех приживления опухоли. [67]
Сообщалось о множественных РСК простаты , [72] легкие и многие другие органы, включая печень , поджелудочную железу , почки или яичники . [73] При раке простаты клетки, инициирующие опухоль, идентифицированы в CD44. + [74] подмножество клеток как CD44 + а2б1 + , [75] ТРА-1-60 + CD151 + CD166 + [76] или АЛДГ + [77] клеточные популяции. предполагаемых маркерах РСК легких , включая CD133. Сообщалось о + , [78] АЛДГ + , [79] CD44 + [80] и онкофетальный белок 5Т4 + . [81]
Метастазирование [ править ]
Метастазирование является основной причиной летальности опухолей. Однако не каждая опухолевая клетка может метастазировать. [82] Этот потенциал зависит от факторов, определяющих рост , ангиогенез , инвазию и другие основные процессы.
Эпителиально-мезенхимальный переход [ править ]
В эпителиальных опухолях эпителиально-мезенхимальный переход (ЕМТ). решающим событием считается [83] ЕМТ и обратный переход от мезенхимального к эпителиальному фенотипу ( МЕТ ) участвуют в эмбриональном развитии , которое включает нарушение гомеостаза эпителиальных клеток и приобретение мигрирующего мезенхимального фенотипа. [84] ЕМТ, по-видимому, контролируется каноническими путями, такими как WNT и трансформирующий фактор роста β . [85]
Важной особенностью ЕМТ является потеря мембранного E-кадгерина в слипчивых соединениях , где β-катенин может играть значительную роль. Транслокация β-катенина из слипчивых соединений в ядро может привести к потере E-кадгерина и, как следствие, к ЕМТ. Ядерный β-катенин, по-видимому, может напрямую, транскрипционно активировать гены- гена E-кадгерина мишени, связанные с ЕМТ, такие как репрессор SLUG (также известный как SNAI2 ). [86] Механические свойства микроокружения опухоли , такие как гипоксия , могут способствовать выживанию РСК и метастатическому потенциалу за счет стабилизации факторов, индуцируемых гипоксией, посредством взаимодействия с АФК ( активными формами кислорода ). [87] [88]
Опухолевые клетки, подвергшиеся ЕМТ, могут быть предшественниками метастатических раковых клеток или даже метастатических РСК. [89] [82] В инвазивном крае рака поджелудочной железы субпопуляция CD133 + CXCR4 + (рецептор для хемокина CXCL12, также известный как SDF1 лиганд ). Эти клетки продемонстрировали значительно более сильную миграционную активность, чем их аналог CD133. + CXCR4 − клетки, но оба показали одинаковую способность к развитию опухоли. [90] Более того, ингибирование рецептора CXCR4 снижало метастатический потенциал без изменения онкогенной способности. [91]
Эпителиально-мезенхимальный переход требует железа, которое поглощается клетками через CD44. Полученные в результате мезенхимальные клетки обладают свойствами раковых стволовых клеток. [92]
Двухфазный шаблон экспрессии [ править ]
При раке молочной железы CD44 + CD24 −/низкий клетки обнаруживаются в метастатических плевральных выпотах. [25] Напротив, повышенное количество CD24 + клетки были идентифицированы в отдаленных метастазах у больных раком молочной железы. [93] Возможно, CD44 + CD24 −/низкий клетки первоначально метастазируют и на новом участке меняют свой фенотип и подвергаются ограниченной дифференцировке. [94] Гипотеза о двухфазном характере экспрессии предполагает две формы раковых стволовых клеток — стационарную (SCS) и мобильную (MCS). СКС внедрены в ткани и сохраняются в различных областях на протяжении всего прогрессирования опухоли. MCS расположены на границе раздела опухоль-хозяин. Эти клетки, по-видимому, происходят из SCS путем приобретения временного ЕМТ (рис. 7). [95]

Последствия [ править ]
РСК имеют значение для терапии рака, в том числе для выявления заболеваний, выбора мишеней для лекарств, предотвращения метастазирования и стратегий вмешательства.
Лечение [ править ]
ОСК по своей природе более устойчивы к химиотерапевтическим агентам . Этому способствуют 5 основных факторов: [96]
- 1. Их ниша защищает их от контакта с большими концентрациями противораковых препаратов.
- 2. Они экспрессируют различные трансмембранные белки, такие как MDR1 и BCRP , которые выкачивают лекарства из цитоплазмы.
- 3. Они делятся медленно, как это обычно делают взрослые стволовые клетки , и поэтому не уничтожаются химиотерапевтическими агентами, которые нацелены на быстро реплицирующиеся клетки путем повреждения ДНК или ингибирования митоза.
- 4. Они активируют белки восстановления повреждений ДНК.
- 5. Для них характерна чрезмерная активация антиапоптотических сигнальных путей.
После химиотерапевтического лечения выжившие РСК способны повторно заселить опухоль и вызвать рецидив. Чтобы предотвратить это, необходимо использовать дополнительное лечение, направленное на удаление не только раковых соматических клеток, но и РСК.
Таргетинг [ править ]
Избирательное воздействие на РСК может позволить лечить агрессивные неоперабельные опухоли, а также предотвращать метастазирование и рецидивы. Гипотеза предполагает, что после элиминации РСК рак может регрессировать вследствие дифференцировки и/или гибели клеток. [ нужна ссылка ] Доля опухолевых клеток, которые являются РСК и, следовательно, должны быть удалены, неясна. [97]
Исследования искали конкретные маркеры [25] а также для протеомных и геномных признаков опухолей, которые отличают РСК от других. [98] В 2009 году ученые идентифицировали соединение салиномицин , которое избирательно снижает долю РСК молочной железы у мышей более чем в 100 раз по сравнению с паклитакселом , широко используемым химиотерапевтическим агентом. [99] Позже в 2017 году было показано, что салиномицин убивает раковые стволовые клетки, связывая железо в лизосомах и убивая клетки, используя их уязвимость к ферроптозу. [100] Некоторые типы раковых клеток могут выжить при лечении салиномицином посредством аутофагии . [101] при этом клетки используют кислые органеллы , такие как лизосомы, для разложения и переработки определенных типов белков. Использование ингибиторов аутофагии может убить раковые стволовые клетки, выживающие за счет аутофагии. [102]
Рецептор клеточной поверхности альфа-рецептора интерлейкина-3 (CD123) сверхэкспрессируется на CD34+CD38- лейкозных стволовых клетках (LSC) при остром миелогенном лейкозе (ОМЛ), но не на нормальных CD34+CD38- клетках костного мозга . [103] Обработка мышей NOD/SCID с трансплантированным ОМЛ моноклональным антителом , специфичным к CD123 , нарушала доставку ЛСК в костный мозг и уменьшала общую репопуляцию клеток ОМЛ, включая долю ЛСК у вторичных мышей-реципиентов. [104]
В исследовании 2015 года наночастицы с миР-34а и бикарбонатом аммония были упакованы и доставлены в РСК простаты на мышиной модели. Затем они облучали это место лазерным светом ближнего инфракрасного диапазона . Это привело к тому, что наночастицы увеличились в размерах в три или более раз, разорвав эндосомы и диспергировав РНК в клетке. миР-34а может снижать уровни CD44. [105] [106]
В 2017 году было продемонстрировано, что раковые стволовые клетки содержат больше железа и что это ахиллесова пята этих клеток. Нацеленное воздействие на железо, особенно путем его секвестрации в лизосомах, может избирательно убить эту популяцию клеток. [107]
Исследование 2018 года выявило ингибиторы семейства ферментов ALDH1A и показало, что они могут избирательно истощать предполагаемые раковые стволовые клетки в нескольких линиях клеток рака яичников. [108]
Новый низкомолекулярный ингибитор, названный соединением 974, специфически нацелен на раковые стволовые клетки и ингибирует пути и гены раковых стволовых клеток. Этот ингибитор также снизил частоту возникновения раковых стволовых клеток у мышей. Это исследование 2022 года также выявило новую связь между ALDH1A1 и старением. Блокирование старения соединением 974 подавляло стволовость рака в клетках рака яичников. [55]
Также было обнаружено, что РСК обладают способностью усугублять устойчивость к лекарствам за счет сверхэкспрессии белков-транспортеров ABC, которые могут перекачивать гидрофобные соединения. [109]
Пути [ править ]
Разработка новых препаратов для воздействия на РСК требует понимания клеточных механизмов, регулирующих пролиферацию клеток. Первые успехи в этой области были достигнуты с использованием гемопоэтических стволовых клеток (ГСК) и их трансформированных аналогов при лейкемии — заболевании, происхождение которого лучше всего изучено. Стволовые клетки многих органов имеют те же клеточные пути, что и ЗКП, полученные из лейкемии.
Нормальная стволовая клетка может быть трансформирована в РСК путем нарушения регуляции контролирующих ее путей пролиферации и дифференцировки или путем индукции активности онкопротеина .
ИМТ-1 [ править ]
был группы Polycomb транскрипции Репрессор Bmi-1 обнаружен как общий онкоген, активируемый при лимфоме. [110] и позже было показано, что он регулирует HSC. [111] Роль Bmi-1 была проиллюстрирована на нервных стволовых клетках. [112] Этот путь, по-видимому, активен в ОСК педиатрических опухолей головного мозга . [113]
Нотч [ править ]
Путь Notch играет роль в контроле пролиферации стволовых клеток нескольких типов клеток, включая гемопоэтические, нервные и молочные клетки. [114] СЦ. Было высказано предположение, что компоненты этого пути действуют как онкогены в молочной железе. [115] и другие опухоли.
Ветвь сигнального пути Notch, которая включает транскрипционный фактор Hes3, регулирует ряд культивируемых клеток с характеристиками РСК, полученных от пациентов с глиобластомой. [116]
Еж Соник и Wnt [ править ]
Эти пути развития являются регуляторами SC. [117] [118] Оба пути Sonic hedgehog (SHH) и Wnt обычно гиперактивируются в опухолях и необходимы для поддержания роста опухоли. Однако факторы транскрипции Gli, которые регулируются SHH, получили свое название от глиом , где они высоко экспрессируются. Между этими двумя путями существует определенная степень перекрестных помех , и они обычно активируются вместе. [119] Напротив, при раке толстой кишки передача сигналов hedgehog, по-видимому, противодействует Wnt. [120]
Доступны блокаторы Sonic Hedgehog, такие как циклопамин . Водорастворимый циклопамин может быть более эффективным при лечении рака. DMAPT, водорастворимое производное партенолида , вызывает окислительный стресс и ингибирует NF-κB. передачу сигналов [121] для ОМЛ (лейкоза) и, возможно, миеломы и рака простаты. Теломераза является предметом исследования физиологии РСК. [122] GRN163L ( Иметелстат ) недавно был начат в испытаниях по нацеливанию на стволовые клетки миеломы.
Передача сигналов Wnt может стать независимой от обычных стимулов за счет мутаций в нижестоящих онкогенах и генах-супрессорах опухолей, которые становятся постоянно активированными, даже если нормальный рецептор не получил сигнала. β-катенин связывается с факторами транскрипции, такими как белок TCF4 , и в сочетании молекулы активируют необходимые гены. LF3 сильно ингибирует это связывание in vitro, в клеточных линиях и снижает рост опухоли на мышиных моделях. Это предотвратило репликацию и снизило их способность к миграции, не затрагивая при этом здоровые клетки. После лечения раковых стволовых клеток не осталось. Это открытие стало результатом « рациональной разработки лекарств » с использованием AlphaScreens и ELISA . технологий [123]
Ссылки [ править ]
- ^ Jump up to: а б Шрипадманабх М., Толи Б.Дж. (2018). «Исследование ниши раковых стволовых клеток с использованием трехмерных моделей опухолей in vitro и микрофлюидики». Достижения биотехнологии . 36 (4): 1094–1110. doi : 10.1016/j.biotechadv.2018.03.009 . ПМИД 29559382 . S2CID 4039860 .
- ^ Мукерджи, Сиддхартха (29 октября 2010 г.). «Раковая спящая клетка» . Нью-Йорк Таймс . Проверено 15 июля 2014 г.
- ^ Рейя Т., Моррисон С.Дж., Кларк М.Ф., Вайсман И.Л. (ноябрь 2001 г.). «Стволовые клетки, рак и раковые стволовые клетки». Природа . 414 (6859): 105–11. Бибкод : 2001Natur.414..105R . дои : 10.1038/35102167 . hdl : 2027.42/62862 . ПМИД 11689955 . S2CID 4343326 .
- ^ Хеппнер Г.Х., Миллер Б.Е. (1983). «Гетерогенность опухоли: биологические последствия и терапевтические последствия». Обзоры рака и метастазов . 2 (1): 5–23. дои : 10.1007/BF00046903 . ПМИД 6616442 . S2CID 2769686 .
- ^ Пэн К., Николсон А.М., обр. Х., Харрисон Л.А., Уилсон Г.Д., Беркерт Дж., Джеффри Р., Элисон М.Р., Лойженга Л., Лин В.Р., Макдональд С.А., Райт Н.А., Харрисон Р., Пеппеленбош член парламента, Янковски Дж.А. (апрель 2013 г.). «Идентификация долгоживущих, сохраняющих метку клеток с незафиксированной линией в пищеводе и желудке здорового человека, а также в метапластическом пищеводе» (PDF) . Гастроэнтерология . 144 (4): 761–70. дои : 10.1053/j.gastro.2012.12.022 . ПМИД 23266557 .
- ^ Боннет Д., Дик Дж. Э. (июль 1997 г.). «Острый миелоидный лейкоз человека организован как иерархия, берущая начало от примитивной кроветворной клетки». Природная медицина . 3 (7): 730–7. дои : 10.1038/nm0797-730 . ПМИД 9212098 . S2CID 205381050 .
- ^ Jump up to: а б с Бек Б., Бланпейн С. (2013), «Раскрытие потенциала раковых стволовых клеток», Nat Rev Cancer , 13 (10): 727–38, doi : 10.1038/nrc3597 , PMID 24060864 , S2CID 11882727
- ^ Шеклтон М., Кинтана Э., Фирон Э.Р., Моррисон С.Дж. (сентябрь 2009 г.). «Гетерогенность рака: раковые стволовые клетки против клональной эволюции» . Клетка . 138 (5): 822–9. дои : 10.1016/j.cell.2009.08.017 . ПМИД 19737509 . S2CID 2615068 .
- ^ Jump up to: а б с д Ван В, Цюань Й, Фу Ц, Лю Й, Лян Й, Ву Дж, Ян Г, Луо С, Оуян Ц, Ван Й (2014): «Динамика между субпопуляциями раковых клеток раскрывает модель, координирующую как иерархические, так и стохастические концепции» , PLOS ONE , 9 (1): e84654, Bibcode : 2014PLoSO...984654W , doi : 10.1371/journal.pone.0084654 , PMC 3886990 , PMID 24416258
- ^ Кресо А., Дик Дж. Э. (2014), «Эволюция модели раковых стволовых клеток», Cell Stem Cell , 14 (3): 275–91, doi : 10.1016/j.stem.2014.02.006 , PMID 24607403
- ^ Барабе Ф., Кеннеди Дж.А., Хоуп К.Дж., Дик Дж.Э. (апрель 2007 г.). «Моделирование возникновения и прогрессирования острого лейкоза человека у мышей». Наука . 316 (5824): 600–4. Бибкод : 2007Sci...316..600B . дои : 10.1126/science.1139851 . ПМИД 17463288 . S2CID 34417642 .
- ^ Nowell PC (октябрь 1976 г.). «Клональная эволюция популяций опухолевых клеток». Наука . 194 (4260): 23–8. Бибкод : 1976Sci...194...23N . дои : 10.1126/science.959840 . ПМИД 959840 . S2CID 38445059 .
- ^ Кларк Э.А., Голуб Т.Р., Ландер Э.С., Хайнс Р.О. (август 2000 г.). «Геномный анализ метастазов показывает важную роль RhoC». Природа . 406 (6795): 532–5. Бибкод : 2000Natur.406..532C . дои : 10.1038/35020106 . ПМИД 10952316 . S2CID 4301092 .
- ^ Cabrera MC, Hollingsworth RE, Hurt EM (2015), «Пластичность раковых стволовых клеток и иерархия опухолей», World J Stem Cells , 7 (1): 27–36, doi : 10.4252/wjsc.v7.i1.27 , PMC 4300934 , PMID 25621103
- ^ Jump up to: а б Гупта П.Б., Чаффер К.Л., Вайнберг Р.А. (сентябрь 2009 г.). «Раковые стволовые клетки: мираж или реальность?». Природная медицина . 15 (9): 1010–2. дои : 10.1038/nm0909-1010 . ПМИД 19734877 . S2CID 14167044 . [ постоянная мертвая ссылка ] >
- ^ Кинтана Э., Шеклтон М., Сабель М.С., Фуллен Д.Р., Джонсон Т.М., Моррисон С.Дж. (декабрь 2008 г.). «Эффективное образование опухолей одиночными клетками меланомы человека» . Природа . 456 (7222): 593–8. Бибкод : 2008Natur.456..593Q . дои : 10.1038/nature07567 . ПМК 2597380 . ПМИД 19052619 .
- ^ Ичирю Н., Фэйрчайлд П.Дж. (2013). «Иммунная привилегия стволовых клеток». В Завазаве Н (ред.). Иммунобиология эмбриональных стволовых клеток . Методы молекулярной биологии. Том. 1029. стр. 1–16. дои : 10.1007/978-1-62703-478-4_1 . ISBN 978-1-62703-477-7 . ПМИД 23756938 .
- ^ Маккалли С, Волонте А, Чимминиелло С, Пармиани Дж (февраль 2014 г.). «Иммунология раковых стволовых клеток в солидных опухолях. Обзор». Европейский журнал рака . 50 (3): 649–55. дои : 10.1016/j.ejca.2013.11.014 . ПМИД 24333096 .
- ^ Jump up to: а б Бруттель В.С., Вишхузен Дж (2014). «Иммунология раковых стволовых клеток: ключ к пониманию онкогенеза и ускользания опухоли от иммунитета?» . Границы в иммунологии . 5 : 360. дои : 10.3389/fimmu.2014.00360 . ПМК 4114188 . ПМИД 25120546 .
- ^ Хейфлик Л. (март 1965 г.). «Ограниченное время жизни диплоидных клеточных штаммов человека in vitro». Экспериментальные исследования клеток . 37 (3): 614–36. дои : 10.1016/0014-4827(65)90211-9 . ПМИД 14315085 .
- ^ Jump up to: а б с Боннет Д., Дик Дж. Э. (июль 1997 г.). «Острый миелоидный лейкоз человека организован как иерархия, берущая начало от примитивной кроветворной клетки». Природная медицина . 3 (7): 730–7. дои : 10.1038/nm0797-730 . ПМИД 9212098 . S2CID 205381050 .
- ^ Игнатова Т.Н., Кукеков В.Г., Лейвелл Э.Д., Суслов О.Н., Врионис Ф.Д., Штейндлер Д.А. (сентябрь 2002 г.). «Кортикальные глиальные опухоли человека содержат нервные стволоподобные клетки, экспрессирующие астроглиальные и нейрональные маркеры in vitro». Глия . 39 (3): 193–206. дои : 10.1002/glia.10094 . ПМИД 12203386 . S2CID 37236481 .
- ^ Сундар С.Дж., Се Дж.К., Манджила С., Латия Дж.Д., Слоан А. (декабрь 2014 г.). «Роль раковых стволовых клеток при глиобластоме» . Нейрохирургический фокус . 37 (6): Е6. дои : 10.3171/2014.9.FOCUS14494 . ПМИД 25434391 . S2CID 2039638 .
- ^ Сингх С.К., Кларк И.Д., Терасаки М., Бонн В.Е., Хокинс С., Сквайр Дж., Диркс П.Б. (сентябрь 2003 г.). «Идентификация раковых стволовых клеток в опухолях головного мозга человека» . Исследования рака . 63 (18): 5821–8. ПМИД 14522905 .
- ^ Jump up to: а б с д Аль-Хадж М., Вича М.С., Бенито-Эрнандес А., Моррисон С.Дж., Кларк М.Ф. (апрель 2003 г.). «Проспективная идентификация туморогенных клеток рака молочной железы» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (7): 3983–8. Бибкод : 2003PNAS..100.3983A . дои : 10.1073/pnas.0530291100 . ПМК 153034 . ПМИД 12629218 .
- ^ Jump up to: а б с О'Брайен К.А., Поллетт А., Галлинджер С., Дик Дж.Э. (январь 2007 г.). «Клетка рака толстой кишки человека, способная инициировать рост опухоли у мышей с иммунодефицитом». Природа . 445 (7123): 106–10. Бибкод : 2007Natur.445..106O . дои : 10.1038/nature05372 . ПМИД 17122772 . S2CID 4419499 .
- ^ Чжан С., Балч С., Чан М.В., Лай Х.К., Матей Д., Шильдер Дж.М., Ян П.С., Хуан Т.Х., Племянник К.П. (июнь 2008 г.). «Идентификация и характеристика клеток, инициирующих рак яичников, из первичных опухолей человека» . Исследования рака . 68 (11): 4311–20. дои : 10.1158/0008-5472.CAN-08-0364 . ПМК 2553722 . ПМИД 18519691 .
- ^ Альверо А.Б., Чен Р., Фу Х.Х., Монтанья М., Шварц П.Е., Резерфорд Т., Силаси Д.А., Стеффенсен К.Д., Вальдстрем М., Висинтин И., Мор Г. (январь 2009 г.). «Молекулярное фенотипирование стволовых клеток рака яичников человека раскрывает механизмы восстановления и химиорезистентности» . Клеточный цикл . 8 (1): 158–66. дои : 10.4161/cc.8.1.7533 . ПМК 3041590 . ПМИД 19158483 .
- ^ Ли К., Хайдт Д.Г., Далерба П., Бурант К.Ф., Чжан Л., Адсай В., Вича М., Кларк М.Ф., Симеоне Д.М. (февраль 2007 г.). «Идентификация стволовых клеток рака поджелудочной железы» . Исследования рака . 67 (3): 1030–7. doi : 10.1158/0008-5472.CAN-06-2030 . ПМИД 17283135 .
- ^ Мейтленд, штат Нью-Джерси, Коллинз, А.Т. (июнь 2008 г.). «Стволовые клетки рака простаты: новая мишень для терапии». Журнал клинической онкологии . 26 (17): 2862–70. дои : 10.1200/JCO.2007.15.1472 . ПМИД 18539965 .
- ^ Ланг С.Х., Frame FM, Коллинз А.Т. (январь 2009 г.). «Стволовые клетки рака простаты» . Журнал патологии . 217 (2): 299–306. дои : 10.1002/путь.2478 . ПМЦ 2673349 . ПМИД 19040209 .
- ^ Шаттон Т., Мерфи Г.Ф., Фрэнк Нью-Йорк, Ямаура К., Ваага-Гассер А.М., Гассер М., Жан К., Джордан С., Дункан Л.М., Вейсгаупт С., Фульбригге Р.К., Куппер Т.С., Сайег М.Х., Фрэнк М.Х. (январь 2008 г.). «Идентификация клеток, инициирующих меланомы человека» . Природа . 451 (7176): 345–9. Бибкод : 2008Natur.451..345S . дои : 10.1038/nature06489 . ПМК 3660705 . ПМИД 18202660 .
- ^ Бойко А.Д., Разоренова О.В., ван де Рейн М., Светтер С.М., Джонсон Д.Л., Ли Д.П., Батлер П.Д., Ян Г.П., Джошуа Б., Каплан М.Дж., Лонгакер М.Т., Вайсман И.Л. (июль 2010 г.). «Клетки, инициирующие меланому человека, экспрессируют рецептор фактора роста нервов нервного гребня CD271» . Природа . 466 (7302): 133–7. Бибкод : 2010Natur.466..133B . дои : 10.1038/nature09161 . ПМЦ 2898751 . ПМИД 20596026 .
- ^ Шмидт П., Копецкий С., Хомбах А., Зигрино П., Маух С., Абкен Х. (февраль 2011 г.). «Искоренение меланомы путем целенаправленного устранения незначительной субпопуляции опухолевых клеток» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (6): 2474–9. Бибкод : 2011PNAS..108.2474S . дои : 10.1073/pnas.1009069108 . ПМК 3038763 . ПМИД 21282657 .
- ^ Чивенни Дж., Уолтер А., Коберт Н., Михик-Пробст Д., Зипсер М., Беллони Б., Зайферт Б., Мох Х., Даммер Р., ван ден Брук М., Зоммер Л. (апрель 2011 г.). «Стволовые клетки меланомы человека, положительные по CD271, связанные с метастазами, устанавливают гетерогенность опухоли и долгосрочный рост» . Исследования рака . 71 (8): 3098–109. дои : 10.1158/0008-5472.CAN-10-3997 . ПМИД 21393506 .
- ^ Мацуи В., Хафф К.А., Ван К., Малехорн М.Т., Барбер Дж., Танхеко Ю., Смит Б.Д., Сивин С.И., Джонс Р.Дж. (март 2004 г.). «Характеристика клоногенных клеток множественной миеломы» . Кровь . 103 (6): 2332–6. дои : 10.1182/кровь-2003-09-3064 . ПМК 3311914 . ПМИД 14630803 .
- ^ Мацуи В., Ван Кью, Барбер Дж.П., Бреннан С., Смит Б.Д., Боррелло I, МакНис I, Лин Л., Амбиндер Р.Ф., Пикок С., Уоткинс Д.Н., Хафф К.А., Джонс Р.Дж. (январь 2008 г.). «Клоногенные предшественники множественной миеломы, свойства стволовых клеток и устойчивость к лекарствам» . Исследования рака . 68 (1): 190–7. дои : 10.1158/0008-5472.CAN-07-3096 . ПМК 2603142 . ПМИД 18172311 .
- ^ Колмонт К.С., Бенкетах А., Рид С.Х., Хоук Н.В., Телфорд В.Г., Охьяма М., Удей М.К., Йи К.Л., Фогель Дж.К., Патель Г.К. (январь 2013 г.). «Клетки базальноклеточной карциномы человека, экспрессирующие CD200, инициируют рост опухоли» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (4): 1434–9. Бибкод : 2013PNAS..110.1434C . дои : 10.1073/pnas.1211655110 . ПМК 3557049 . ПМИД 23292936 .
- ^ Патель Г.К., Йи К.Л., Терунума А., Телфорд В.Г., Вунг Н., Юспа С.Х., Фогель Дж.К. (февраль 2012 г.). «Идентификация и характеристика опухолевых клеток при первичном плоскоклеточном раке кожи человека» . Журнал исследовательской дерматологии . 132 (2): 401–9. дои : 10.1038/jid.2011.317 . ПМК 3258300 . ПМИД 22011906 .
- ^ Jump up to: а б Юань, Шаопэн; Стюарт, Кэтрин С.; Ян, Ихао; Абдуселамоглу, Мерве Дениз; Париджи, С. Мартина; Фейнберг, Тамар Ю.; Туманенг, Карен; Ян, Хансыль; Леворс, Джон М.; Полак, Лиза; Нг, Дэвид; Фукс, Элейн (30 ноября 2022 г.). «Ras вызывает злокачественные новообразования посредством взаимодействия стволовых клеток с микроокружением» . Природа . 612 (7940): 555–563. дои : 10.1038/s41586-022-05475-6 . ISSN 1476-4687 . ПМЦ 9750880 . ПМИД 36450983 . S2CID 254124217 .
- ^ Прециози, Луиджи (2003). Моделирование и симуляция рака . Бока-Ратон: CRC Press. ISBN 978-1-58488-361-6 .
- ^ Гангули Р., Пури И.К. (февраль 2006 г.). «Математическая модель гипотезы раковых стволовых клеток» . Пролиферация клеток . 39 (1): 3–14. дои : 10.1111/j.1365-2184.2006.00369.x . ПМК 6495990 . ПМИД 16426418 .
- ^ Гангули Р., Пури И.К. (июнь 2007 г.). «Математическая модель эффективности химиотерапевтических препаратов для остановки роста опухоли, основанная на гипотезе раковых стволовых клеток» . Пролиферация клеток . 40 (3): 338–54. дои : 10.1111/j.1365-2184.2007.00434.x . ПМК 6496895 . ПМИД 17531079 .
- ^ Ван Ю, Ян Дж, Чжэн Х, Томасек Г.Дж., Чжан П., Маккивер П.Е., Ли Э.Ю. , Чжу Ю. (июнь 2009 г.). «Экспрессия мутантных белков р53 указывает на родственные связи между нервными стволовыми клетками и злокачественной астроцитарной глиомой на мышиной модели» . Раковая клетка . 15 (6): 514–26. дои : 10.1016/j.ccr.2009.04.001 . ПМК 2721466 . ПМИД 19477430 .
- ^ Лопес-Ласаро М (01 января 2015 г.). «Миграционная способность стволовых клеток может объяснить существование рака неизвестной первичной локализации. Переосмысление метастазирования» . Онсознание . 2 (5): 467–75. doi : 10.18632/oncoscience.159 . ПМЦ 4468332 . ПМИД 26097879 .
- ^ Лопес-Ласаро М (18 августа 2015 г.). «Теория деления стволовых клеток рака» . Клеточный цикл . 14 (16): 2547–8. дои : 10.1080/15384101.2015.1062330 . ПМЦ 5242319 . ПМИД 26090957 .
- ^ Кларк М.Ф., Дик Дж.Э., Диркс П.Б., Ивс С.Дж., Джеймисон Ч., Джонс Д.Л., Висвадер Дж., Вайсман И.Л., Валь Г.М. (октябрь 2006 г.). «Раковые стволовые клетки - взгляды на текущее состояние и будущие направления: семинар AACR по раковым стволовым клеткам». Исследования рака . 66 (19): 9339–44. дои : 10.1158/0008-5472.CAN-06-3126 . ПМИД 16990346 .
- ^ Голебьевска А., Бронс Н.Х., Бьерквиг Р., Николу С.П. (февраль 2011 г.). «Критическая оценка анализа побочных популяций в исследованиях стволовых клеток и раковых стволовых клеток» . Клеточная стволовая клетка . 8 (2): 136–47. дои : 10.1016/j.stem.2011.01.007 . ПМИД 21295271 .
- ^ Jump up to: а б Шаренберг CW, Харки М.А., Торок-Сторб Б. (январь 2002 г.). «Транспортер ABCG2 является эффективным эффлюксным насосом Hoechst 33342 и преимущественно экспрессируется незрелыми человеческими гемопоэтическими предшественниками» . Кровь . 99 (2): 507–12. дои : 10.1182/blood.V99.2.507 . ПМИД 11781231 . S2CID 23736581 .
- ^ Пастрана Э., Сильва-Варгас В., Дётч Ф. (май 2011 г.). «С широко открытыми глазами: критический обзор образования сфер как анализа стволовых клеток» . Клеточная стволовая клетка . 8 (5): 486–98. дои : 10.1016/j.stem.2011.04.007 . ПМЦ 3633588 . ПМИД 21549325 .
- ^ Николис СК (февраль 2007 г.). «Раковые стволовые клетки и гены стволовости в нейроонкологии». Нейробиология болезней . 25 (2): 217–29. дои : 10.1016/j.nbd.2006.08.022 . ПМИД 17141509 . S2CID 25401054 .
- ^ Цао В, Чен К, Болкестейн М, Инь Ю, Верстеген ММ, Бийвелдс МДж, Ван В, Туйсуз Н, Тен Берге Д, Спренгерс Д, Метселаар Х.Дж., ван дер Лаан Л.Дж., Квеккебум Дж., Смитс Р., Пеппеленбош М.П., Пан К. (октябрь 2017 г.). «Динамика пролиферативных и покоящихся стволовых клеток при гомеостазе и повреждении печени». Гастроэнтерология . 153 (4): 1133–47. дои : 10.1053/j.gastro.2017.07.006 . ПМИД 28716722 .
- ^ Цао В, Ли М, Лю Дж, Чжан С, Ноордам Л, Верстеген ММ, Ван Л, Ма Б, Ли С, Ван В, Болкестейн М, Дукас М, Чен К, Ма З, Бруно М, Шпренгерс Д, Квеккебум Дж , ван дер Лаан Л.Дж., Смитс Р., Пеппеленбош, член парламента, Пан К. (апрель 2020 г.). «LGR5 маркирует целевые клетки, инициирующие опухоль, при раке печени у мышей» . Природные коммуникации . 11 (1): 1961. Бибкод : 2020NatCo..11.1961C . дои : 10.1038/s41467-020-15846-0 . ПМК 7181628 . ПМИД 32327656 .
- ^ Ядав У.П., Сингх Т., Кумар П., Шарма П., Каур Х., Шарма С., Сингх С., Кумар С. и Мехта К. (2020). «Метаболические адаптации раковых стволовых клеток» . Границы онкологии . 10 (Метаболическая пластичность рака): 1010. doi : 10.3389/fonc.2020.01010 . ПМК 7330710 . ПМИД 32670883 .
- ^ Jump up to: а б с Мураликришнан, Вайшнави; Клык, Клык; Учитывая, Тайлер С.; Подичети, Рам; Щербинин Михаил; Меткалф, Тара X.; Шрирамкумар, Шрути; О'Хаган, Хизер М.; Херли, Томас Д.; Племянник Кеннет П. (15 июля 2022 г.). «Новый ингибитор ALDH1A1 блокирует индуцированное платиной старение и стволовидность при раке яичников» . Раки . 14 (14): 3437. doi : 10.3390/cancers14143437 . ISSN 2072-6694 . ПМЦ 9318275 . ПМИД 35884498 .
- ^ Ким Ю.С., Кайдина А.М., Чан Дж.Х., Ярыгин К.Н., Лупатов А.Ю. (2017). «Молекулярные маркеры раковых стволовых клеток, проверенные in vivo». Биохим. Московское доп. Сер. Б. 11 (1): 43–54. дои : 10.1134/S1990750817010036 . S2CID 90912166 .
- ^ Висвадер Дж. Э., Линдеман Г. Дж. (октябрь 2008 г.). «Раковые стволовые клетки в солидных опухолях: накопление данных и нерешенные вопросы». Обзоры природы. Рак . 8 (10): 755–68. дои : 10.1038/nrc2499 . ПМИД 18784658 . S2CID 40382183 .
- ^ Хиршманн-Джакс С., Фостер А.Е., Вульф Г.Г., Нухтерн Дж.Г., Джакс Т.В., Гобель У., Гуделл М.А., Бреннер М.К. (сентябрь 2004 г.). «Отдельная «побочная популяция» клеток с высокой способностью оттока лекарств в опухолевых клетках человека» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (39): 14228–33. Бибкод : 2004PNAS..10114228H . дои : 10.1073/pnas.0400067101 . ПМК 521140 . ПМИД 15381773 .
- ^ Жинестье С., Хур М.Х., Шараф-Жофре Э., Монвиль Ф., Датчер Дж., Браун М., Жакемье Дж., Вьенс П., Клеер К.Г., Лю С., Шотт А., Хейс Д., Бирнбаум Д., Вича М.С., Донту Дж. (ноябрь 2007 г.) . «ALDH1 является маркером нормальных и злокачественных стволовых клеток молочной железы человека и предиктором плохого клинического исхода» . Клеточная стволовая клетка . 1 (5): 555–67. дои : 10.1016/j.stem.2007.08.014 . ПМЦ 2423808 . ПМИД 18371393 .
- ^ Пече С., Тосони Д., Конфалоньери С., Маццарол Г., Векки М., Ронзони С., Бернар Л., Виале Дж., Пеличчи П.Г., Ди Фиоре П.П. (январь 2010 г.). «Биологическая и молекулярная гетерогенность рака молочной железы коррелирует с содержанием в нем раковых стволовых клеток» . Клетка . 140 (1): 62–73. дои : 10.1016/j.cell.2009.12.007 . ПМИД 20074520 . S2CID 5547104 .
- ^ Дэн С., Ян X, Лассус Х, Лян С., Каур С., Йе Q, Ли С., Ван Л.П., Роби К.Ф., Орсулик С., Коннолли Д.С., Чжан Й., Монтоне К., Бютцов Р., Кукош Г., Чжан Л. (апрель 2010 г.) ). Цао Ю (ред.). «Различные уровни экспрессии и характер маркера стволовых клеток, изоформы 1 альдегиддегидрогеназы (ALDH1), при эпителиальном раке человека» . ПЛОС ОДИН . 5 (4): е10277. Бибкод : 2010PLoSO...510277D . дои : 10.1371/journal.pone.0010277 . ПМЦ 2858084 . ПМИД 20422001 .
- ^ Мейер М.Дж., Флеминг Дж.М., Лин А.Ф., Хусснэйн С.А., Гинзбург Э., Вондерхаар Б.К. (июнь 2010 г.). «CD44posCD49fhiCD133/2hi определяет клетки, инициирующие ксенотрансплантат, при раке молочной железы, отрицательном по рецепторам эстрогена» . Исследования рака . 70 (11): 4624–33. дои : 10.1158/0008-5472.CAN-09-3619 . ПМЦ 4129519 . ПМИД 20484027 .
- ^ Сингх С.К., Хокинс С., Кларк И.Д., Сквайр Дж.А., Баяни Дж., Хиде Т., Хенкельман Р.М., Кузимано М.Д., Диркс П.Б. (ноябрь 2004 г.). «Идентификация клеток, инициирующих опухоль головного мозга человека». Природа . 432 (7015): 396–401. Бибкод : 2004Natur.432..396S . дои : 10.1038/nature03128 . ПМИД 15549107 . S2CID 4430962 .
- ^ Сон М.Дж., Вулард К., Нам Д.Х., Ли Дж., Файн Х.А. (май 2009 г.). «SSEA-1 является маркером накопления клеток, инициирующих опухоль, в глиобластоме человека» . Клеточная стволовая клетка . 4 (5): 440–52. дои : 10.1016/j.stem.2009.03.003 . ПМЦ 7227614 . ПМИД 19427293 .
- ^ Jump up to: а б Маццолени С., Полити Л.С., Пала М., Коминелли М., Франзин А., Серджи Серджи Л., Фалини А., Де Пальма М., Бульфоне А., Полиани П.Л., Галли Р. (октябрь 2010 г.). «Экспрессия рецептора эпидермального фактора роста идентифицирует функционально и молекулярно различные клетки, инициирующие опухоль, в мультиформной глиобластоме человека и необходима для глиомагенеза» . Исследования рака . 70 (19): 7500–13. дои : 10.1158/0008-5472.CAN-10-2353 . ПМИД 20858720 .
- ^ Анидо Х, Саес-Бордериас А, Гонсалес-Хунка А, Родон Л, Фолч Г, Кармона М.А., Прието-Санчес Р.М., Барба И, Мартинес-Саес Э, Прудкин Л, Куартас И, Равентос К, Мартинес-Рикарте Ф, Пока М.А., Гарсия-Дорадо Д., Лан М.М., Йинглинг Х.М., Родон Дж., Саукильо Дж., Базельга Дж., Сеоан Дж. (декабрь 2010 г.). «Ингибиторы рецептора TGF-β нацелены на популяцию клеток, инициирующих глиому, CD44 (высокий)/Id1 (высокий) при глиобластоме человека» . Раковая клетка . 18 (6): 655–68. дои : 10.1016/j.ccr.2010.10.023 . ПМИД 21156287 .
- ^ Jump up to: а б Далерба П., Дилла С.Дж., Парк И.К., Лю Р., Ван X, Чо Р.В., Хоуи Т., Герни А., Хуанг Э.Х., Симеоне Д.М., Шелтон А.А., Пармиани Г., Кастелли С., Кларк М.Ф. (июнь 2007 г.). «Фенотипическая характеристика стволовых клеток колоректального рака человека» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (24): 10158–63. Бибкод : 2007PNAS..10410158D . дои : 10.1073/pnas.0703478104 . ПМК 1891215 . ПМИД 17548814 .
- ^ Уилсон Б.Дж., Шаттон Т., Жан К., Гассер М., Ма Дж., Сааб КР, Шанче Р., Ваага-Гассер А.М., Голд Дж.С., Хуанг К., Мерфи Г.Ф., Фрэнк М.Х., Фрэнк Нью-Йорк (август 2011 г.). «ABCB5 идентифицирует резистентную к терапии популяцию опухолевых клеток у пациентов с колоректальным раком» . Исследования рака . 71 (15): 5307–16. дои : 10.1158/0008-5472.CAN-11-0221 . ПМК 3395026 . ПМИД 21652540 .
- ^ Оду С., Форер Х., Хоппо Т., Гузик Л., Штольц Д.Б., Льюис Д.В., Голлин С.М., Гамблин Т.К., Геллер Д.А., Лагасс Э. (сентябрь 2008 г.). «Стохастическая модель происхождения раковых стволовых клеток при метастатическом раке толстой кишки» . Исследования рака . 68 (17): 6932–41. дои : 10.1158/0008-5472.CAN-07-5779 . ПМЦ 2562348 . ПМИД 18757407 .
- ^ Хуанг Э.Х., Хайнс М.Дж., Чжан Т., Жинестье С., Донту Дж., Аппельман Х., Филдс Дж.З., Вича М.С., Боман Б.М. (апрель 2009 г.). «Альдегиддегидрогеназа 1 является маркером нормальных и злокачественных стволовых клеток толстой кишки (SC) человека и отслеживает перенаселение SC во время онкогенеза толстой кишки» . Исследования рака . 69 (8): 3382–9. дои : 10.1158/0008-5472.CAN-08-4418 . ПМК 2789401 . ПМИД 19336570 .
- ^ Кемпер К., Сприк М.Р., де Бри М., Скопеллити А., Вермюлен Л., Хук М., Зейлстра Дж., Палс С.Т., Мехмет Х., Стасси Г., Медема Дж.П. (январь 2010 г.). «Эпитоп AC133, но не белок CD133, теряется при дифференцировке раковых стволовых клеток» . Исследования рака . 70 (2): 719–29. doi : 10.1158/0008-5472.CAN-09-1820 . ПМИД 20068153 .
- ^ Лю С, Келнар К, Лю Б, Чен X, Кэлхун-Дэвис Т, Ли Х, Патравала Л, Ян Х, Джетер С, Онорио С, Виггинс Дж. Ф., Бадер А. Г., Феджин Р., Браун Д., Тан Д. Г. (февраль 2011 г.) . «МикроРНК миР-34а ингибирует стволовые клетки и метастазы рака простаты путем прямой репрессии CD44» . Природная медицина . 17 (2): 211–5. дои : 10.1038/нм.2284 . ПМК 3076220 . ПМИД 21240262 .
- ^ Хо ММ, Нг А.В., Лам С., Хунг Дж.Ю. (май 2007 г.). «Боковая популяция клеточных линий и опухолей рака легких человека обогащена стволоподобными раковыми клетками». Исследования рака . 67 (10): 4827–33. дои : 10.1158/0008-5472.CAN-06-3557 . ПМИД 17510412 .
- ^ Патравала Л., Калхун Т., Шнайдер-Бруссар Р., Ли Х., Бхатия Б., Тан С., Рейли Дж.Г., Чандра Д., Чжоу Дж., Клейпул К., Коглан Л., Тан Д.Г. (март 2006 г.). «Высокоочищенные CD44+ раковые клетки простаты из ксенотрансплантатов опухолей человека обогащены онкогенными и метастатическими клетками-предшественниками» . Онкоген . 25 (12): 1696–708. дои : 10.1038/sj.onc.1209327 . ПМИД 16449977 . S2CID 28427525 .
- ^ Дубровска А., Ким С., Саламоне Р.Дж., Уокер Дж.Р., Майра С.М., Гарсия-Эчеверрия С., Шульц П.Г., Редди В.А. (январь 2009 г.). «Роль передачи сигналов PTEN/Akt/PI3K в поддержании и жизнеспособности популяций стволоподобных клеток рака простаты» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (1): 268–73. Бибкод : 2009PNAS..106..268D . дои : 10.1073/pnas.0810956106 . ПМК 2629188 . ПМИД 19116269 .
- ^ Раджасекхар В.К., Студер Л., Джеральд В., Соччи Н.Д., Шер Х.И. (январь 2011 г.). «Стволово-подобные клетки, инициирующие опухоль, при раке предстательной железы человека демонстрируют повышенную передачу сигналов NF-κB» . Природные коммуникации . 2 (1): 162. Бибкод : 2011NatCo...2..162R . дои : 10.1038/ncomms1159 . ПМК 3105310 . ПМИД 21245843 .
- ^ Ли Т, Су Ю, Мэй Ю, Ленг К, Ленг Б, Лю З, Стасс С.А., Цзян Ф (февраль 2010 г.). «ALDH1A1 является маркером злокачественных стволовых клеток простаты и предиктором исхода заболевания у пациентов с раком простаты» . Лабораторные исследования; Журнал технических методов и патологии . 90 (2): 234–44. дои : 10.1038/labinvest.2009.127 . ПМЦ 3552330 . ПМИД 20010854 .
- ^ Эрамо А., Лотти Ф., Сетте Г., Пилоцци Э., Биффони М., Ди Вирджилио А., Контичелло С., Руко Л., Пешле С., Де Мария Р. (март 2008 г.). «Идентификация и расширение популяции туморогенных стволовых клеток рака легких» . Смерть клеток и дифференцировка . 15 (3): 504–14. дои : 10.1038/sj.cdd.4402283 . ПМИД 18049477 . S2CID 2484961 .
- ^ Салливан Дж.П., Спинола М., Додж М., Расо М.Г., Беренс С., Гао Б., Шустер К., Шао С., Ларсен Дж.Э., Салливан Л.А., Онорио С., Се Ю, Скальони П.П., ДиМайо Дж.М., Газдар А.Ф., Шей Дж.В., Вистуба II , Минна Джей Ди (декабрь 2010 г.). «Активность альдегиддегидрогеназы выбирает стволовые клетки аденокарциномы легких в зависимости от передачи сигналов notch» . Исследования рака . 70 (23): 9937–48. дои : 10.1158/0008-5472.CAN-10-0881 . ПМК 3058307 . ПМИД 21118965 .
- ^ Люнг Э.Л., Фискус Р.Р., Тунг Дж.В., Тин В.П., Ченг Л.К., Сихое А.Д., Финк Л.М., Ма Ю, Вонг М.П. (ноябрь 2010 г.). Джин Д.И. (ред.). «Клетки немелкоклеточного рака легких, экспрессирующие CD44, обладают свойствами, подобными стволовым клеткам» . ПЛОС ОДИН . 5 (11): e14062. Бибкод : 2010PLoSO...514062L . дои : 10.1371/journal.pone.0014062 . ПМЦ 2988826 . ПМИД 21124918 .
- ^ Дамелин М, Гелес К.Г., Фоллетти М.Т., Юань П., Бакстер М., Голас Дж., ДиДжозеф Дж.Ф., Карнуб М., Хуанг С., Дизль В., Беренс С., Чой С.Э., Риос К., Грузас Дж., Шридхаран Л., Догер М., Кунц А. , Хаманн П.Р., Эванс Д., Армеллино Д., Хандке К., Маркетт К., Чистякова Л., Богхарт Э.Р., Авраам Р.Т., Вистуба II, Чжоу Б.Б. (июнь 2011 г.). «Определение клеточной иерархии при раке легких выявило онкофетальный антиген, экспрессируемый на клетках, инициирующих опухоль» . Исследования рака . 71 (12): 4236–46. дои : 10.1158/0008-5472.CAN-10-3919 . ПМИД 21540235 .
- ^ Jump up to: а б Гонсалес-Вильярреал Калифорния, Кирос-Рейес АГ, Ислас Х.Ф., Гарса-Тревиньо EN (20 августа 2020 г.). «Стволовые клетки колоректального рака в стадии метастазирования в печень» . Границы онкологии . 10 : 1511. doi : 10.3389/fonc.2020.01511 . ПМЦ 7468493 . ПМИД 32974184 .
- ^ Тьери Дж.П. (июнь 2002 г.). «Эпителиально-мезенхимальные переходы при опухолевой прогрессии». Обзоры природы. Рак . 2 (6): 442–54. дои : 10.1038/nrc822 . ПМИД 12189386 . S2CID 5236443 .
- ^ Ангерер Л.М., Ангерер Р.К. (июнь 1999 г.). «Регулятивное развитие эмбриона морского ежа: сигнальные каскады и градиенты морфогена» . Семинары по клеточной биологии и биологии развития . 10 (3): 327–34. дои : 10.1006/scdb.1999.0292 . ПМИД 10441547 .
- ^ Мани С.А., Ян Дж., Брукс М., Шванингер Г., Чжоу А., Миура Н., Куток Дж.Л., Хартвелл К., Ричардсон А.Л., Вайнберг Р.А. (июнь 2007 г.). «Мезенхима Forkhead 1 (FOXC2) играет ключевую роль в метастазировании и связана с агрессивным базальноподобным раком молочной железы» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (24): 10069–74. Бибкод : 2007PNAS..10410069M . дои : 10.1073/pnas.0703900104 . ЧВК 1891217 . ПМИД 17537911 .
- ^ Коначчи-Соррелл М., Симха И., Бен-Йедидиа Т., Блехман Дж., Савагнер П., Бен-Зеев А. (ноябрь 2003 г.). «Ауторегуляция экспрессии E-кадгерина посредством кадгерин-кадгериновых взаимодействий: роль передачи сигналов бета-катенина, Slug и MAPK» . Журнал клеточной биологии . 163 (4): 847–57. дои : 10.1083/jcb.200308162 . ПМК 2173691 . ПМИД 14623871 .
- ^ Комито Дж., Кальвани М., Джаннони Э., Бьянкини Ф., Калорини Л., Торре Э., Мильоре С., Джордано С., Кьяруджи П. (август 2011 г.). «Стабилизация HIF-1α с помощью митохондриальных АФК способствует Met-зависимому инвазивному росту и васкулогенной мимикрии в клетках меланомы». Свободно-радикальная биология и медицина . 51 (4): 893–904. doi : 10.1016/j.freeradbiomed.2011.05.042 . hdl : 2158/496457 . ПМИД 21703345 .
- ^ Спилл Ф, Рейнольдс Д.С., Камм Р.Д., Заман М.Х. (август 2016 г.). «Влияние физического микроокружения на прогрессирование и метастазирование опухоли» . Современное мнение в области биотехнологии . 40 : 41–48. дои : 10.1016/j.copbio.2016.02.007 . ПМЦ 4975620 . ПМИД 26938687 .
- ^ Каплан Р.Н., Риба Р.Д., Захарулис С., Брэмли А.Х., Винсент Л., Коста С. и др. (декабрь 2005 г.). «VEGFR1-положительные гематопоэтические предшественники костного мозга инициируют преметастатическую нишу» . Природа . 438 (7069): 820–7. Бибкод : 2005Natur.438..820K . дои : 10.1038/nature04186 . ПМЦ 2945882 . ПМИД 16341007 .
- ^ Герман П.С., Хубер С.Л., Херрлер Т., Айхер А., Эллварт Дж.В., Губа М. и др. (сентябрь 2007 г.). «Различные популяции раковых стволовых клеток определяют рост опухоли и метастатическую активность при раке поджелудочной железы человека» . Клеточная стволовая клетка . 1 (3): 313–23. дои : 10.1016/j.stem.2007.06.002 . ПМИД 18371365 .
- ^ Ян З.Ф., Хо Д.В., Нг М.Н., Лау К.К., Ю В.К., Нгай П. и др. (февраль 2008 г.). «Значение раковых стволовых клеток CD90+ при раке печени человека» . Раковая клетка . 13 (2): 153–66. дои : 10.1016/j.ccr.2008.01.013 . ПМИД 18242515 .
- ^ Мюллер, Себастьян; Синдикубвабо, Фабьен; Каньеке, Татьяна; Лафон, Энн; Версини, Антуан; Ломбард, Беранжер; Лоу, Дамарис; Ву, Тинг-Ди; Жинестье, Кристоф; Шараф-Жофре, Эммануэль; Дюран, Аделина; Валло, Селин; Бауланд, Сильвен; Слуга Николас; Родригес, Рафаэль (3 августа 2020 г.). «CD44 регулирует эпигенетическую пластичность, опосредуя эндоцитоз железа» . Природная химия . 12 (10): 929–938. дои : 10.1038/s41557-020-0513-5 . ПМЦ 7612580 . ПМИД 32747755 .
- ^ Шипицин М, Кэмпбелл ЛЛ, Аргани П, Веремович С, Блуштайн-Кимрон Н, Яо Дж, Никольская Т, Серебрянская Т, Бероухим Р, Ху М, Халушка М.К., Сукумар С., Паркер Л.М., Андерсон К.С., Харрис Л.Н., Гарбер Дж.Е., Ричардсон А.Л., Шнитт С.Дж., Никольский Ю., Гельман Р.С., Поляк К. (март 2007 г.). «Молекулярное определение гетерогенности опухоли молочной железы» . Раковая клетка . 11 (3): 259–73. дои : 10.1016/j.ccr.2007.01.013 . ПМИД 17349583 .
- ^ Шмельков С.В., Батлер Дж.М., Хупер А.Т., Хормиго А., Кушнер Дж., Милд Т., Сент-Клер Р., Бальевич М., Уайт И., Джин Д.К., Чедберн А., Мерфи А.Дж., Валенсуэла Д.М., Гейл Н.В., Терстон Г., Янкопулос Г.Д., Д. 'Анжелика М., Кемени Н., Лайден Д., Рафии С. (июнь 2008 г.). «Экспрессия CD133 не ограничивается стволовыми клетками, и как CD133+, так и CD133- метастатические клетки рака толстой кишки инициируют опухоли» . Журнал клинических исследований . 118 (6): 2111–20. дои : 10.1172/JCI34401 . ПМК 2391278 . ПМИД 18497886 .
- ^ Браблец Т., Юнг А., Спадерна С., Хлубек Ф., Киршнер Т. (сентябрь 2005 г.). «Мнение: миграция раковых стволовых клеток - комплексная концепция прогрессирования злокачественных опухолей». Обзоры природы. Рак . 5 (9): 744–9. дои : 10.1038/nrc1694 . PMID 16148886 . S2CID 20197701 .
- ^ Чжао Дж (2016). «Раковые стволовые клетки и химиорезистентность: в набеге выживает самый умный» . Фармакол Тер . 160 : 145–58. doi : 10.1016/j.pharmthera.2016.02.008 . ПМЦ 4808328 . ПМИД 26899500 .
- ^ Диркс П. (июль 2010 г.). «Раковые стволовые клетки: приглашение на второй раунд» . Природа . 466 (7302): 40–1. Бибкод : 2010Natur.466...40D . дои : 10.1038/466040a . ПМИД 20596007 . S2CID 205056717 .
- ^ Хаскинс В.Е., Идала С., Джадхав Ю.Л., Лабхан М.С., Перичерла В.К., Перлман Э.Дж. (май 2012 г.). «Понимание неопластических стволовых клеток на основе гелевой протеомики детских опухолей зародышевых клеток» . Детская кровь и рак . 58 (5): 722–8. дои : 10.1002/pbc.23282 . ПМК 3204330 . ПМИД 21793190 .
- ^ Гупта П.Б., Ондер Т.Т., Цзян Г., Тао К., Купервассер С., Вайнберг Р.А., Ландер Э.С. (август 2009 г.). «Идентификация селективных ингибиторов стволовых раковых клеток методом высокопроизводительного скрининга» . Клетка . 138 (4): 645–659. дои : 10.1016/j.cell.2009.06.034 . ПМЦ 4892125 . ПМИД 19682730 .
- ^ Май, Транг Тхи; Хамаи, Ахмед; Хинцш, Антье; Каньеке, Татьяна; Мюллер, Себастьян; Вичински, Жюльен; Кабо, Оливье; Лерой, Кристина; Дэвид, Амандина; Асеведо, Вероника; Ре, Акихидэ; Джинестье, Кристоф; Бирнбаум, Дэниел; Шараф-Жофре, Эммануэль; Кодоньо, Патрис; Мехпур, Марьям; xРодригес, Рафаэль Родригес (октябрь 2017 г.). «Салиномицин убивает раковые стволовые клетки, связывая железо в лизосомах» . Природная химия . 9 (10): 1025–1033. дои : 10.1038/nchem.2778 . ПМК 5890907 . ПМИД 28937680 .
- ^ Джангамредди-младший, Гавами С., Грабарек Дж., Крац Г., Вечек Е., Фредрикссон Б.А., Рао Парити Р.К., Цеслар-Побуда А., Паниграхи С., Лос М.Дж. (сентябрь 2013 г.). «Салиномицин индуцирует активацию аутофагии, митофагии и влияет на полярность митохондрий: различия между первичными и раковыми клетками» . Biochimica et Biophysical Acta (BBA) - Исследования молекулярных клеток . 1833 (9): 2057–69. дои : 10.1016/j.bbamcr.2013.04.011 . ПМИД 23639289 . S2CID 9298118 .
- ^ Влахопулос С., Крицелис Е., Вутсас И.Ф., Перес С.А., Мошови М., Баксеванис К.Н., Хрусос Г.П. (2014). «Новое применение старых лекарств? Перспективные цели использования хлорохинов в терапии рака». Текущие цели по борьбе с наркотиками . 15 (9): 843–51. дои : 10.2174/1389450115666140714121514 . ПМИД 25023646 .
- ^ Джордан Коннектикут, Апчерч Д., Сильвасси С.Дж., Гусман М.Л., Ховард Д.С., Петтигрю А.Л., Мейерроуз Т., Росси Р., Граймс Б., Рицциери Д.А., Люгер С.М., Филлипс Г.Л. (октябрь 2000 г.). «Альфа-цепь рецептора интерлейкина-3 является уникальным маркером стволовых клеток острого миелогенного лейкоза человека». Лейкемия . 14 (10): 1777–84. дои : 10.1038/sj.leu.2401903 . ПМИД 11021753 . S2CID 9467715 .
- ^ Джин Л., Ли Э.М., Рамшоу Х.С., Басфилд С.Дж., Пеоппл А.Г., Уилкинсон Л., Гатридж М.А., Томас Д., Барри Э.Ф., Бойд А., Гиринг Д.П., Вайро Г., Лопес А.Ф., Дик Дж.Е., Лок Р.Б. (июль 2009 г.). «Опосредованное моноклональными антителами воздействие на CD123, альфа-цепь рецептора IL-3, уничтожает стволовые клетки острого миелолейкоза человека» . Клеточная стволовая клетка . 5 (1): 31–42. дои : 10.1016/j.stem.2009.04.018 . hdl : 10536/DRO/DU:30132380 . ПМИД 19570512 .
- ^ « Нанобомбы, взрывающие раковые клетки | KurzweilAI» . www.kurzweilai.net . 07.12.2015 . Проверено 20 февраля 2016 г.
- ^ Ван Х, Агарвал П., Чжао С., Ю Дж, Лу Х, Хэ Х (январь 2016 г.). «Нанобомба, активируемая лазером ближнего инфракрасного диапазона, для преодоления барьеров на пути доставки микроРНК». Продвинутые материалы . 28 (2): 347–55. дои : 10.1002/adma.201504263 . ПМИД 26567892 . S2CID 205264763 .
- ^ Май, Транг Тхи; Хамаи, Ахмед; Хинцш, Антье; Каньеке, Татьяна; Мюллер, Себастьян; Вичински, Жюльен; Кабо, Оливье; Лерой, Кристина; Дэвид, Амандина; Асеведо, Вероника; Ре, Акихидэ; Джинестье, Кристоф; Бирнбаум, Дэниел; Шараф-Жофре, Эммануэль; Кодоньо, Патрис; Мехпур, Марьям; xРодригес, Рафаэль Родригес (октябрь 2017 г.). «Салиномицин убивает раковые стволовые клетки, связывая железо в лизосомах» . Природная химия . 9 (10): 1025–1033. дои : 10.1038/nchem.2778 . ПМК 5890907 . ПМИД 28937680 .
- ^ Хаддл BC, Гримли Э., Бухман С.Д., Ччербинин М., Дебнат Б., Мехта П., Ян К., Морган К.А., Ли С., Фелтон Дж.А., Сан Д., Мета Г., Неамати Н., Буканович Р.Дж., Херли Т.Д., Ларсен С.Д. (2018) . «Структурная оптимизация нового класса селективных ингибиторов подсемейства альдегиддегидрогеназы 1А (ALDH1A) как потенциальное дополнение к химиотерапии рака яичников» . J Med Chem . 61 (19): 8754–8773. doi : 10.1021/acs.jmedchem.8b00930 . ПМК 6477540 . ПМИД 30221940 .
- ^ Света, Бишт. «Раковые стволовые клетки: от понимания основ до последних достижений и терапевтического таргетинга» . Хидави .
- ^ Хаупт Ю., Бат М.Л., Харрис А.В., Адамс Дж.М. (ноябрь 1993 г.). «Трансген bmi-1 индуцирует лимфомы и сотрудничает с myc в онкогенезе». Онкоген . 8 (11): 3161–4. ПМИД 8414519 .
- ^ Пак И.К., Цянь Д., Киль М., Беккер М.В., Пихаля М., Вайсман И.Л., Моррисон С.Дж., Кларк М.Ф. (май 2003 г.). «ИМТ-1 необходим для поддержания самообновляющихся гемопоэтических стволовых клеток взрослых» (PDF) . Природа . 423 (6937): 302–5. Бибкод : 2003Natur.423..302P . дои : 10.1038/nature01587 . hdl : 2027.42/62508 . ПМИД 12714971 . S2CID 4403711 .
- ^ Молофски А.В. , Пардал Р., Ивашита Т., Парк И.К., Кларк М.Ф., Моррисон С.Дж. (октябрь 2003 г.). «Зависимость от Bmi-1 отличает самообновление нервных стволовых клеток от пролиферации предшественников» . Природа . 425 (6961): 962–7. Бибкод : 2003Natur.425..962M . дои : 10.1038/nature02060 . ПМК 2614897 . ПМИД 14574365 .
- ^ Хеммати Х.Д., Накано И., Лазарефф Дж.А., Мастерман-Смит М., Гешвинд Д.Х., Броннер-Фрейзер М., Корнблюм Х.И. (декабрь 2003 г.). «Раковые стволовые клетки могут возникать из педиатрических опухолей головного мозга» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (25): 15178–83. Бибкод : 2003PNAS..10015178H . дои : 10.1073/pnas.2036535100 . ПМК 299944 . ПМИД 14645703 .
- ^ Донту Дж., Джексон К.В., МакНиколас Э., Кавамура М.Дж., Абдалла В.М., Вича М.С. (2004). «Роль передачи сигналов Notch в определении судьбы клеток стволовых клеток/клеток-предшественников молочной железы человека» . Исследование рака молочной железы . 6 (6): Р605–15. дои : 10.1186/bcr920 . ПМЦ 1064073 . ПМИД 15535842 .
- ^ Дьевар А., Болье Н., Жоликер П. (октябрь 1999 г.). «Участие Notch1 в развитии опухолей молочной железы у мышей». Онкоген . 18 (44): 5973–81. дои : 10.1038/sj.onc.1202991 . hdl : 1807.1/567 . ПМИД 10557086 . S2CID 2533675 .
- ^ Пак Д.М., Юнг Дж., Масйкур Дж., Макрогкикас С., Эберманн Д., Саха С., Рольяно Р., Паолилло Н., Пачиони С., Маккей Р.Д., Позер С., Андруцеллис-Теотокис А. (2013). «Hes3 регулирует количество клеток в культурах мультиформной глиобластомы с характеристиками стволовых клеток» . Научные отчеты . 3 : 1095. Бибкод : 2013NatSR...3E1095P . дои : 10.1038/srep01095 . ПМК 3566603 . ПМИД 23393614 .
- ^ Милошевич, В. и др. Аутокринные схемы Wnt/IL-1β/IL-8 контролируют хеморезистентность клеток, инициирующих мезотелиому, путем индукции ABCB5.Int. Дж. Рак, https://doi.org/10.1002/ijc.32419
- ^ Бичи, Пенсильвания, Кархадкар СС, Берман DM (ноябрь 2004 г.). «Восстановление тканей и обновление стволовых клеток при канцерогенезе». Природа . 432 (7015): 324–31. Бибкод : 2004Natur.432..324B . дои : 10.1038/nature03100 . ПМИД 15549094 . S2CID 4428056 .
- ^ Чжоу Б.П., Хунг MC (июнь 2005 г.). «Wnt, еж и улитка: сестринские пути, которые контролируют регуляцию метастазирования с помощью GSK-3beta и бета-Trcp» . Клеточный цикл . 4 (6): 772–6. дои : 10.4161/cc.4.6.1744 . ПМИД 15917668 . S2CID 31467958 .
- ^ Акиёси Т., Накамура М., Кога К., Накашима Х., Яо Т., Цунэёси М., Танака М., Катано М. (июль 2006 г.). «Gli1, уровень экспрессии которого снижается при колоректальном раке, ингибирует пролиферацию клеток рака толстой кишки, включая активацию передачи сигналов Wnt» . Гут . 55 (7): 991–9. дои : 10.1136/gut.2005.080333 . ПМК 1856354 . ПМИД 16299030 .
- ^ Ше М, Чен X (январь 2009 г.). «Нацеливание на сигнальные пути, активные в раковых стволовых клетках, для преодоления лекарственной устойчивости». Чжунго Фэй АИ За Чжи = Китайский журнал рака легких . 12 (1): 3–7. doi : 10.3779/j.issn.1009-3419.2009.01.001 . ПМИД 20712949 .
- ^ Боллманн FM (август 2008 г.). «Многоликость теломеразы: новые экстрателомерные эффекты». Биоэссе . 30 (8): 728–32. doi : 10.1002/bies.20793 . ПМИД 18623070 . S2CID 205478338 .
- ^ Ходж Р. (25 января 2016 г.). «Взлом программ раковых стволовых клеток» . www.medicalxpress.com . Медицинский экспресс . Проверено 12 февраля 2016 г.
Дальнейшее чтение [ править ]
- Раджасекхар В.К., изд. (2014). Раковые стволовые клетки . Хобокен, Нью-Джерси: Уайли. ISBN 9781118356166 .
- Милошевич В. и др. (январь 2020 г.). «Аутокринные схемы Wnt/IL-1β/IL-8 контролируют химиорезистентность в клетках, инициирующих мезотелиому, путем индукции ABCB5» . Межд. Дж. Рак . 146 (1): 192–207. дои : 10.1002/ijc.32419 . hdl : 2318/1711962 . ПМИД 31107974 . S2CID 160014053 .
| Источники/типы | |
|---|---|
| Эффективность клеток |
|
| Похожие статьи | |