Ядро прилегание
| Ядро прилегание | |
|---|---|
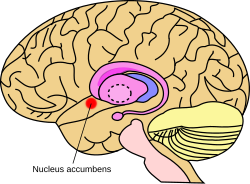 Приблизительное местоположение ядра прилегает в мозге | |
 Ядро прилегание мозга мыши | |
| Подробности | |
| Часть | Мезолимбический путь Базальные ганглии ( вентральный стриатум ) |
| Части | Ядро прикусы. Оболочка Ядро прилегает ядро |
| Идентификаторы | |
| латинский | Ядро теряет септи |
| Аббревиатура (ы) | NAC или NACC |
| Сетка | D009714 |
| Нейроналы | 277 |
| Neurolex Id | Birnlex_727 |
| TA98 | A14.1.09.440 |
| TA2 | 5558 |
| FMA | 61889 |
| Анатомические термины нейроанатомии | |
( Прилегающее ядро NAC или NACC ;, также известное как ядро accumbens , или ранее как ядро, прилегающее септи , латыни для « ядра , прилегающего к перегородке »), является областью в базальном переднем мозге ростала в область гипоталамуса преоптическую . [ 1 ] Ядро прилегает и обонятельный бугорок в совокупности образует вентральный стриатум . Вентральный стриатум и дорсальный стриатум в совокупности образуют стриатум , который является основным компонентом базальных ганглиев . [ 2 ] Дофаминергические нейроны мезолимбического пути проецируют на гамбаргическую среду колючих нейронов ядра, прилежащих и обонятельных бугорков. [ 3 ] [ 4 ] Каждое полушарие головного мозга имеет свое собственное ядро привыкание, которое можно разделить на две структуры: ядро прилежащее ядро и ядро прилегающее оболочку. Эти субструктуры имеют разные морфологии и функции.
Различные субрегионы NACC (ядра против оболочки) и субпопуляции нейронов в каждой области ( D1-типа против D2-типа колючих нейронов) отвечают за различные когнитивные функции . [ 5 ] [ 6 ] В целом, ядро accumbens играет значительную роль в когнитивной обработке мотивации , отвращения , вознаграждения (то есть, стимулирующей значимости , удовольствия и позитивного подкрепления ) и подкреплению обучения (например, павловско-инструментальный перенос ); [ 4 ] [ 7 ] [ 8 ] [ 9 ] [ 10 ] Следовательно, это играет значительную роль в зависимости . [ 4 ] [ 8 ] Кроме того, часть ядра Accumbens Accumbens является централизованно вовлеченной в индукцию медленного волнового сна . [ 11 ] [ 12 ] [ 13 ] [ 14 ] Accumbens ядро играет меньшую роль в обработке страха (форма отвращения), импульсивности и эффекта плацебо . [ 15 ] [ 16 ] [ 17 ] участвует в кодировании новых моторных программ . Он также [ 4 ]
Структура
[ редактировать ]Прилегающее ядро - это заполнитель нейронов, который описывается как имеющая внешнюю оболочку и внутреннее ядро. [ 4 ]
Вход
[ редактировать ]Основные глутаматергические входы в прилежащее ядро включают префронтальную кору (в частности, прелимбическая кора коры и инфралимбическая кора ), базолатеральная амигдала , вентральный гиппокамп , ядра талама (специфически ядерные ядерные и ядерные ядра, и ядра, и ядерные, и ядерные, и ядерные, и ядерные, именные ядры , и ядерные ядра , и ядерные, и ядра, и ядра Вентральная сегментарная область (VTA). [ 18 ] Прилегающее ядро получает дофаминергические входы из вентральной области тегментальной, которая соединяется через мезолимбический путь . Прилегающее ядро часто описывается как часть кортико-базальной ганглий-таламо-кортикальной петли . [ 19 ]
Дофаминергические входы из VTA модулируют активность ГАМКергических нейронов в прилегающем ядре. Эти нейроны прямо или косвенно активируются эйфориантными препаратами (например, амфетамином , опиатами и т. Д.) И участия в полезных опытах (например, пол, музыка, упражнения и т. Д.). [ 20 ] [ 21 ]
Другой основной источник ввода исходит от CA1 и вентрального до дорсомедиальной субикулума гиппокампа области прилегающего ядра. Небольшая деполяризация клеток в ядре прилегает коррелирует с позитивностью нейронов гиппокампа, что делает их более возбудимыми. Коррелированные клетки этих возбужденных состояний средних колючих нейронов в ядре, прилежащем, распределены в равной степени между субикулумом и CA1. Обнаружено, что нейроны субикулума гиперполяризуются (увеличивают негативность), в то время как нейроны CA1 «Ripple» (Fire> 50 Гц) для достижения этого праймирования. [ 22 ]
Прилегающее ядро является одной из немногих областей, которая получает высокую плотность гистаминергических проекций из туберомамиллярного ядра (единственный источник гистаминовых нейронов в мозге). [ 23 ]
Выход
[ редактировать ]Выходные нейроны ядра укубенки посылают аксональные проекции в базальные ганглии и вентральный аналог глобуса Pallidus , известного как вентральный паллидум (VP). Вице -президент, в свою очередь, проецирует в медиальное дорсальное ядро дорсального таламуса , который проецируется на префронтальную кору, а также обратно в вентральный и дорсальный стриатум . Другие эфферверы из прилегающего ядра включают соединения с хвостом вентральной сегментарной области , [ 24 ] Субъективная черная ретикулярная образование понков . и [ 1 ]
Оболочка
[ редактировать ]У Accumbens Shell ядра ( оболочка NACC ) представляет собой подструктуру прилегающего ядра. Оболочка и ядро вместе образуют все прилегающие ядра.
Расположение: оболочка является внешней областью прилегающего ядра, и - в отличие от ядра - считается частью расширенной миндалины , расположенной на его ростральном полюсе.
Типы клеток: нейроны в ядрах прилежащих в основном являются средними колючими нейронами (MSN), содержащими в основном D1-тип (IE, DRD1 и DRD5 ) или D2-типа (IE, DRD2 , DRD3 и DRD4 ) дофаминовых рецепторов . Субпопуляция MSN содержит рецепторы типа D1 и D2-типа, причем приблизительно 40% стриатальных MSN, экспрессирующих мРНК как DRD1 , так и DRD2 . [ 19 ] [ 25 ] [ 26 ] Эти MSN NACC смешанного типа с рецепторами типа D1 и D2-типа в основном ограничены оболочкой NACC. [ 19 ] Нейроны в оболочке, по сравнению с ядром, имеют более низкую плотность дендритных шипов , меньше терминальных сегментов и меньше сегментов ветвей, чем в ядре. Нейроны раковины проецируются в подкоммиссуральную часть вентрального паллидума , а также в вентральную сегментарную область, а также в обширные участки гипоталамуса и расширенной миндалины. [ 27 ] [ 28 ] [ 29 ]
Функция: оболочка прилегающего ядра участвует в когнитивной обработке вознаграждения , включая субъективные «симпатичные» реакции на определенные приятные стимулы , мотивационную значимость и позитивное подкрепление . [ 4 ] [ 5 ] [ 30 ] [ 31 ] Также было показано, что эта оболочка NACC опосредует специфический павловиан-инструментальный перенос , явление, при котором классически обусловленный стимул изменяет поведение операнта . [ 32 ] [ 9 ] [ 10 ] «Гедоническая горячая точка» или центр удовольствий, который отвечает за приятный или «симпатичный» компонент некоторых внутренних вознаграждений, также расположен в небольшом отделении в раковине Medial NACC. [ 30 ] [ 33 ] [ 34 ] Привыкающие препараты оказывают большее влияние на высвобождение дофамина в оболочке, чем в ядре. [ 4 ]
Основной
[ редактировать ]Ядро прилежащее ядро ( ядро NACC ) является внутренней субструктурой прилегающего ядра.
Местоположение: ядро прилежащее ядро является частью вентрального стриатума , расположенного в базальных ганглиях. Типы клеток: ядро NACC состоит в основном из средних колючих нейронов, содержащих в основном рецепторы дофамина типа D1 или D2. Средние колючие нейроны среды D1 обеспечивают связанные с вознаграждением когнитивные процессы, [ 5 ] [ 35 ] [ 36 ] в то время как средние колючие нейроны среды D2-типа опосредуют связанное с отвращением познание. [ 6 ] Нейроны в ядре, по сравнению с нейронами в оболочке, имеют повышенную плотность дендритных шипов, сегментов ветвей и терминальных сегментов. Из ядра нейроны проецируют в другие субкотические области, такие как Globus pallidus и черная субстанция. ГАМК является одним из основных нейротрансмиттеров в NACC, и рецепторы ГАМК также в изобилии. [ 27 ] [ 29 ]
Функция: ядро прилегающее ядро участвует в когнитивной обработке двигательной функции , связанной с вознаграждением и подкреплением, а также регуляцией медленного сна . [ 4 ] [ 11 ] [ 12 ] [ 13 ] В частности, ядро кодирует новые моторные программы, которые облегчают приобретение данной награды в будущем. [ 4 ] Нейроны непрямого пути (т.е., D2-тип) в ядре NACC, которые совместно экспрессируют аденозин A 2A- рецепторы , зависит от активации, в зависимости от активации, способствуя медленной волне. [ 11 ] [ 12 ] [ 13 ] Также было показано, что ядро NACC опосредует общего павлова-инструментального переноса , явление, при котором классически обусловленный стимул изменяет поведение операнта. [ 32 ] [ 9 ] [ 10 ]
Типы клеток
[ редактировать ]Приблизительно 95% нейронов в NACC представляют собой GABAergic Spiny Neurons (MSN), которые в первую очередь экспрессируют рецепторы типа D1 или D2; [ 20 ] Около 1–2% оставшихся типов нейронов представляют собой большие брюшные холинергические интернейроны , а еще 1–2% - ГАМКергические интернейроны. [ 20 ] По сравнению с ГАМКергическими MSN в оболочке, в сердечнике есть повышенная плотность дендритных шипов, сегментов ветвей и терминальных сегментов. Из ядра нейроны проецируют в другие субкотические области, такие как Globus pallidus и черная субстанция. ГАМК является одним из основных нейротрансмиттеров в NACC, и рецепторы ГАМК также в изобилии. [ 27 ] [ 29 ] Эти нейроны также являются основными проекционными или выходными нейронами прилегания ядра.
Нейрохимия
[ редактировать ]Некоторые из нейротрансмиттеров, нейромодуляторов и гормонов, которые сигнализируют через рецепторы в прилежащем ядре, включают:
Дофамин : дофамин высвобождается в прилежащее ядро после воздействия вознаграждений стимулов , включая рекреационные препараты , такие как замещенные амфетамины , кокаин , никотин и морфин . [ 37 ] [ 38 ]
Фенетиламин и тирамин : фенотиламин и тирамин - это следовые амины , которые синтезируются в нейронах, которые экспрессируют ароматической аминокислот -гидроксилазы (AADC) фермент , который включает в себя все дофаминергические нейроны. [ 39 ] Оба соединения функционируют как дофаминергические нейромодуляторы , которые регулируют обратное захват и высвобождение дофамина в NACC посредством взаимодействия с VMAT2 и TAAR1 в терминале аксона мезолимбических дофаминовых нейронов.
Глюкокортикоиды и дофамин: глюкокортикоидные рецепторы являются единственными кортикостероидными рецепторами в раковине прикуса ядра. В настоящее время известно, что L-допа , стероиды и, в частности, глюкокортикоиды, которые являются единственными известными эндогенными соединениями, которые могут вызывать психотические проблемы, поэтому понимание гормонального контроля над дофаминергическими проекциями в отношении рецепторов глюкокортикоидов может привести к новым лечением психотических симптомов. Недавнее исследование продемонстрировало, что подавление глюкокортикоидных рецепторов привело к снижению высвобождения дофамина, что может привести к будущим исследованиям, включающим антиглукокортикоидные препараты, чтобы потенциально облегчить психотические симптомы. [ 40 ]
ГАМК: Недавнее исследование на крысах, в которых использовались агонисты ГАМК и антагонисты, показало, что в оболочке ГАМК -рецепторы NACC имеют ингибирующий контроль при поворотном поведении, на который влияет дофамин, а ГАМК B рецепторы имеют ингибирующий контроль над поворотом поведения, опосредованным ацетилхолином . [ 27 ] [ 41 ]
Глутамат : Исследования показали, что локальная блокада глутаматергических рецепторов NMDA в основном пространственном обучении NACC. [ 42 ] Другое исследование показало, что как NMDA, так и AMPA (оба глутаматные рецепторы ) играют важную роль в регуляции инструментального обучения. [ 43 ]
Серотонин (5-HT): в целом синапсы 5-HT более распространены и имеют большее количество синаптических контактов в оболочке NACC, чем в ядре. Они также больше и толще, и содержат более крупные плотные ядры, чем их аналоги в ядре.
Функция
[ редактировать ]Награда и подкрепление
[ редактировать ]Прилегающее ядро, являющееся одной из части системы вознаграждений, играет важную роль в обработке вознаграждения стимулов, укрепления стимулов (например, пищи и воды), и тех, которые являются как полезными, так и укрепляющими (наркотики, сексы и физические упражнения). [ 4 ] [ 44 ] Преобладающим ответом нейронов в ядре, приходящем на вознаграждение сахарозу , является ингибирование; Противоположность верна в ответ на введение отвратительного хинина . [ 45 ] Значительные данные из фармакологических манипуляций также свидетельствуют о том, что снижение возбудимости нейронов в прилежащем ядре является полезным, как, например, будет верно в случае стимуляции μ-опиоидного рецептора . [ 46 ] Сигнал зависимого от уровня кислорода в крови (жирный шрифт) в прилежащем ядре выборочно увеличивается во время восприятия приятных, эмоционально возбуждающих картинок и во время ментальных образов приятных, эмоциональных сцен. Однако, поскольку считается жирным шрифтом является косвенной мерой регионального чистого возбуждения с ингибированием, степень, в которой жирный жирный цвет измеряет валентность, зависимая от валентности обработки, неизвестна. [ 47 ] [ 48 ] Из -за обиливания входов NACC из лимбических областей и сильных выходов NACC в моторные области, ядро accumbens было описано Гордоном Могенсеном как раздела между лимбической и моторной системой. [ 49 ] [ 50 ]

Укусы ядра причинно связано с опытом удовольствия. Микроинъекции μ-опиоидных агонистов, агонистов Δ-опиоидов или κ-опиоидных агонистов в ростродорсальном квадранте медиальной оболочки усиливают «симпатичность», в то время как больше каудальных инъекций могут ингибировать реакции отвращения, симпатия реакций или обоих. [ 30 ] Области ядра приходится, которые могут быть приписаны причинной роли в производстве удовольствия, ограничены как анатомически, так и химически, поскольку помимо опиоидных агонистов только эндоканнабиноиды могут усилить вкусный. В ядре, прилежащем в целом, агонисты дофамина, рецептора GABA или антагонисты AMPA исключительно модифицируют мотивацию, в то время как то же самое относится и к опиоидным и эндоканнабиноидам за пределами горячей точки в медиальной оболочке. Существует ростро-каудальный градиент для улучшения аппетитных и страшных ответов, последний из которых традиционно требует только функции рецептора D1, а первая из которых требует функции D1 и D2. Одна интерпретация этого открытия, гипотеза о дезингибировании, утверждает, что ингибирование прилегающих MSN (которые являются ГАМКергическими) дезингибируют нисходящие структуры, что позволяет экспрессию аппетитивного или устойчивого поведения. [ 52 ] Мотивационные эффекты антагонистов AMPA и в меньшей степени агонисты ГАМК являются анатомически гибкими. Стрессовые условия могут расширить области, вызывающие страх, в то время как знакомая среда может уменьшить размер области, вызывающей страх. Кроме того, ввод кортикальной коры из орбитофронтальной коры (OFC) смещает реакцию на реакцию аппетитного поведения и инфралимбический вход, эквивалентный человеческой субгеновой коре, подавляет ответ независимо от валентности. [ 30 ]
Прилегающее ядро не является ни необходимым, ни достаточным для инструментального обучения, хотя манипуляции могут повлиять на производительность инструментальных задач обучения. Одной из задач, при которой эффект поражений NACC очевиден, является павловско-инструментальный перенос (PIT), где сигнал в сочетании с конкретной или общей вознаграждением может усилить инструментальный ответ. Поражения до ядра NACC ухудшают производительность после девальвации и ингибируют эффект общей ямы. С другой стороны, поражения в оболочке только ухудшают эффект определенной ямы. Считается, что это различие отражает конфликтные и аппетитные кондиционированные ответы в оболочке NACC и в ядре NACC, соответственно. [ 53 ]
В дорсальном стриатум наблюдалась дихотомия между D1-MSN и D2-MSN, причем первое усиливает и усиливает локомоцию, а второй-это аверсивное и уменьшающее локомоцию. Такое различие традиционно предполагается, что применяется и к прилегающему ядра, но данные из фармакологических и оптогенетических исследований противоречивы. Кроме того, подмножество MSN NACC экспрессируют как D1, так и D2 MSN, а фармакологическая активация рецепторов D1 в сравнении с D2 не обязательно должна точно активировать нейронные популяции. В то время как большинство исследований не показывают влияния селективной оптогенетической стимуляции MSN D1 или D2 на локомоторную активность, в одном исследовании сообщалось о снижении базальной локомоции с стимуляцией D2-MSN. В то время как в двух исследованиях сообщалось о пониженных усиливающих эффектах кокаина с активацией D2-MSN, одно исследование не показало никакого эффекта. Сообщается также, что активация NACC D2-MSN усиливает мотивацию, как оценивается с помощью PIT, а активность рецептора D2 необходима для усиливающих эффектов стимуляции VTA. [ 54 ] В исследовании 2018 года сообщалось, что активация MSN D2 повышает мотивацию путем ингибирования вентрального паллидума, тем самым дезингибируя VTA. [ 55 ]
Материнское поведение
[ редактировать ]Исследование МРТ , проведенное в 2005 году, показало, что когда матери крысы находились в присутствии своих щенков, области мозга, участвующие в подкреплении, включая прилежащее ядро, были очень активными. [ 56 ] Уровни дофамина увеличиваются в ядре, укусающееся во время материнского поведения, в то время как поражения в этой области расстраивают материнское поведение. [ 57 ] Когда женщинам представлены картинки неродственных детей, FMRI показывают повышенную активность мозга в прилегающем ядре и прилегающем хвостовом ядре, пропорциональном в той степени, в которой женщины считают этих младенцев «милыми». [ 58 ]
Отвращение
[ редактировать ]Активация MSN-типа D1 в прилежащем ядре участвует в вознаграждении, тогда как активация MSN типа D2 в прицепении ядра способствует отвращению . [ 6 ]
Медленный сон
[ редактировать ]В конце 2017 года в исследованиях грызунов, в которых использовались оптогенетические и химиогенетические методы, показали, что средние колючие нейроны с космическим ядром в ядре в ядро, которые коэкспрессируют аденозин A 2A и проецируют в вентральный паллидум прилагаются . Регуляция медленного сна . [ 11 ] [ 12 ] [ 13 ] [ 14 ] В частности, оптогенетическая активация этих косвенных нейронов NACC NACC индуцирует медленную волновую сон, а химиогенетическая активация тех же нейронов увеличивает количество и продолжительность медленных эпизодов сна. [ 12 ] [ 13 ] [ 14 ] Хемогенетическое ингибирование этих нейронов NACC Core подавляет сон. [ 12 ] [ 13 ] Напротив, средние колючие нейроны среды D2 в оболочке NACC, которые экспрессируют аденозин A 2A- рецепторы, не играют роли в регуляции медленно волнистого сна. [ 12 ] [ 13 ]
Клиническое значение
[ редактировать ]Зависимость
[ редактировать ]Современные модели зависимости от хронического употребления лекарств включают изменения в экспрессию генов в мезокортиколимбической проекции . [ 20 ] [ 59 ] [ 60 ] Наиболее важными факторами транскрипции , которые вызывают эти изменения, являются ΔFOSB , циклический аденозин -монофосфатный ( CAM -мыский ) белок, связывающий белок ( CREB ) и ядерный фактор каппа B ( NFκB ). [ 20 ] ΔFOSB является наиболее значимым фактором транскрипции генов в зависимости, поскольку его или генетическая сверхэкспрессия в прилежащем ядре необходима и достаточна эффектов (например, зависимое для многих нейронных адаптаций и поведенческих вирусная наркомания. [ 20 ] [ 35 ] [ 61 ] Сверхэкспрессия ΔFOSB была вовлечена в зависимости от спирта (этанол) , каннабиноидов , кокаина , метилфенидата , никотина , опиоидов , фенциклидина , пропофола и замещенных амфетаминов , среди других. [ 20 ] [ 59 ] [ 61 ] [ 62 ] [ 63 ] Увеличение прилежащего ядра Экспрессии ΔJund может уменьшить или, с значительным увеличением, даже блокировать большую часть нервных изменений, наблюдаемых при хроническом злоупотреблении лекарственными средствами (то есть изменения, опосредованные ΔFOSB). [ 20 ]
ΔFOSB также играет важную роль в регулировании поведенческих реакций на естественные награды, такие как вкусная пища, пол и физические упражнения. [ 20 ] [ 21 ] Природные награды, такие как наркотики, индуцируют ΔFOSB в прилегающем ядре, и хроническое приобретение этих вознаграждений может привести к аналогичному патологическому привыкающему состоянию посредством сверхэкспрессии ΔFOSB. [ 20 ] [ 21 ] [ 44 ] Следовательно, ΔFOSB является ключевым фактором транскрипции, связанным с зависимостью от естественных вознаграждений; [ 20 ] [ 21 ] [ 44 ] В частности, ΔFOSB в ядре, прилежащем, имеет решающее значение для усиливающих эффектов сексуального вознаграждения. [ 21 ] Исследования взаимодействия между естественным и лекарственным вознаграждением позволяют предположить, что психостимуляторы и сексуальное поведение действуют на сходные биомолекулярные механизмы, чтобы индуцировать ΔFOSB в прилегающем ядре и обладает эффектами поперечной сенсибилизации, которые опосредованы через ΔFOSB. [ 44 ] [ 64 ]
Подобно вознаграждениям лекарств, не лекарственные вознаграждения также увеличивают уровень внеклеточного дофамина в оболочке NACC. Высвобождение дофамина, вызванное лекарством в раковине NACC и ядро NACC, обычно не подвержено привычению (то есть развитие терпимости к лекарственным средствам : снижение высвобождения дофамина от будущего воздействия лекарств в результате повторного воздействия лекарственного средства); Напротив, повторное воздействие препаратов, которые вызывают высвобождение дофамина в оболочке NACC и ядро, обычно приводит к сенсибилизации (то есть количество дофамина, которое выделяется в NACC от будущего воздействия лекарственного средства, увеличивается в результате повторного воздействия лекарственного средства). Сенсибилизация высвобождения дофамина в оболочке NACC после повторного воздействия лекарственного средства служит для укрепления ассоциаций стимула-лекарств (то есть классической кондиционирования , которая возникает, когда употребление лекарств неоднократно связано с экологическими стимулами), и эти ассоциации становятся менее подверженными вымиранию (т.е. «Неувлекательное» Эти классически обусловленные ассоциации между употреблением лекарств и стимулами окружающей среды становятся более сложными). После повторного спаривания эти классически обусловленные стимулы окружающей среды (например, контексты и объекты, которые часто в паре с употреблением лекарств) часто становятся Наркотические сигналы , которые функционируют как вторичные усилители употребления наркотиков (то есть после того, как эти ассоциации устанавливаются, воздействие парного экологического стимула запускает тягу или желание использовать препарат, с которым они становятся связанными ). [ 27 ] [ 38 ]
В отличие от лекарств, высвобождение дофамина в оболочке NACC многими типами полезных нелуряющих стимулов обычно подвергается привычке после повторного воздействия (т.е. количество дофамина, которое высвобождается из будущего воздействия вознаграждения, не связанного с лекарственным стимулом, обычно уменьшается в результате повторного воздействия этого стимула). [ 27 ] [ 38 ]
| Форма нейропластичности или поведенческая пластичность |
Тип подкрепления | Источники | |||||
|---|---|---|---|---|---|---|---|
| Опиаты | Психостимуляторы | С высоким содержанием жира или сахарной пищи | Половой акт | Физические упражнения (аэробный) |
Относящийся к окружающей среде обогащение | ||
| ΔFOSB Экспрессия в Ядро прилегание D1-типа MSNS MSNS |
↑ | ↑ | ↑ | ↑ | ↑ | ↑ | [ 44 ] |
| Поведенческая пластичность | |||||||
| Эскалация потребления | Да | Да | Да | [ 44 ] | |||
| Психостимулянт перекрестная сенсибилизация |
Да | Непригодный | Да | Да | Ослаблен | Ослаблен | [ 44 ] |
| Психостимулянт самостоятельное управление |
↑ | ↑ | ↓ | ↓ | ↓ | [ 44 ] | |
| Психостимулянт Условное место предпочтение |
↑ | ↑ | ↓ | ↑ | ↓ | ↑ | [ 44 ] |
| Восстановление поведения, связанного с наркотиками | ↑ | ↑ | ↓ | ↓ | [ 44 ] | ||
| Нейрохимическая пластичность | |||||||
| CREB фосфорилирование в прилегающем ядре |
↓ | ↓ | ↓ | ↓ | ↓ | [ 44 ] | |
| Сенсибилизированный дофаминовый ответ в прилегающем ядре |
Нет | Да | Нет | Да | [ 44 ] | ||
| Измененная стриатального дофамина сигнализация | ↓ DRD2 , ↑ DRD3 | ↑ DRD1 , ↓ DRD2 , ↑ DRD3 | ↑ DRD1 , ↓ DRD2 , ↑ DRD3 | ↑ DRD2 | ↑ DRD2 | [ 44 ] | |
| Измененная сигнализация опиоидов стриатальной опиоидов | Без изменений или ↑ μ-опиоидные рецепторы |
↑ μ-опиоидные рецепторы ↑ κ-опиоидные рецепторы |
↑ μ-опиоидные рецепторы | ↑ μ-опиоидные рецепторы | Нет изменений | Нет изменений | [ 44 ] |
| Изменения в полосатых опиоидных пептидах | ↑ dynorphin Нет изменений: энкефалин |
↑ dynorphin | ↓ enkephalin | ↑ dynorphin | ↑ dynorphin | [ 44 ] | |
| Мезокортиколимбическая синаптическая пластичность | |||||||
| Количество дендритов в прилегании ядра | ↓ | ↑ | ↑ | [ 44 ] | |||
| Дендритная плотность позвоночника в Ядро прилегает |
↓ | ↑ | ↑ | [ 44 ] | |||
Депрессия
[ редактировать ]В апреле 2007 года две исследовательские группы сообщили о том, чтобы вставить электроды в прилежащее ядро, чтобы использовать глубокую стимуляцию мозга для лечения тяжелой депрессии . [ 65 ] В 2010 году эксперименты сообщили, что глубокая стимуляция мозга прилежащего ядра была успешной в снижении симптомов депрессии у 50% пациентов, которые не реагировали на другие методы лечения, такие как электроконвульсивная терапия . [ 66 ] Accumbens также использовалось в качестве мишени для лечения небольших групп пациентов с обсессивно-компульсивным расстройством терапии. [ 67 ]
Абляция
[ редактировать ]Было выполнено радиочастотная абляция зависимости от зависимости и в попытке лечить радиочастотную абляцию ядра. Результаты неубедительны и противоречивы. [ 68 ] [ 69 ]
Эффект плацебо
[ редактировать ]Было показано, что активация NACC происходит в ожидании эффективности лекарственного средства, когда пользователю дают плацебо , что указывает на способность прилегающего ядра в эффекте плацебо . [ 16 ] [ 70 ]
Дополнительные изображения
[ редактировать ]-
 Дофамин и серотонин
Дофамин и серотонин -
 МРТ -корональный срез, показывающий ядро прилегание, изложенное в красном
МРТ -корональный срез, показывающий ядро прилегание, изложенное в красном -
 Стрельцо для сагиттального МРТ с выделением (красным), указывающим на прилежащее ядро
Стрельцо для сагиттального МРТ с выделением (красным), указывающим на прилежащее ядро -
 Прилегающее ядро выделено зеленым
Прилегающее ядро выделено зеленым -
 Прилегающее ядро, выделенное зеленым на сагиттальных изображениях МРТ T1
Прилегающее ядро, выделенное зеленым на сагиттальных изображениях МРТ T1 -
 Прилегающее ядро, выделенное зеленым
Прилегающее ядро, выделенное зеленым






Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный Карлсон Н.Р. (2013). Физиология поведения (11 -е изд.). Бостон: Пирсон. [ страница необходима ]
- ^ Ядро прилегание
- ^ Ikemoto S (ноябрь 2010). «Схема вознаграждения мозга за пределами мезолимбической дофаминовой системы: нейробиологическая теория» . Нейробиологии и биобиоэвиральные обзоры . 35 (2): 129–50. doi : 10.1016/j.neubiorev.2010.02.001 . PMC 2894302 . PMID 20149820 .
Недавние исследования внутричерепного самостоятельного введения нейрохимических веществ (препаратов) показали, что крысы учатся самостоятельно вводить различные препараты в мезолимбические дофаминовые структуры-задних вентральных тегментарных областей, ядро медиального раковины и медиальное обонятельное бугр. ... В 1970 -х годах было признано, что обонятельный бугорок содержит стриатальный компонент, который заполнен гуммигическими колючими нейронами, получающими глутаматергические входные данные образуют корковые области и дофаминергические входы от VTA и проецируя в вентральный палику
Рисунок 3: Вентральный стриатум и самостоятельное управление амфетамином - ^ Jump up to: а беременный в дюймовый и фон глин час я Дж Malenka RC, Nestler EJ, Hyman SE (2009). Sydor A, Brown Ry (Eds.). Молекулярная нейрофармакология: основание для клинической нейробиологии (2 -е изд.). Нью-Йорк: McGraw-Hill Medical. с. 147–148, 367, 376. ISBN 978-0-07-148127-4 Полем
Нейроны VTA DA играют решающую роль в мотивации, поведении, связанном с вознаграждением (глава 15), внимании и множественных формах памяти. Эта организация системы DA, широкая проекция от ограниченного числа клеточных тел, позволяет скоординировать ответы на мощные новые вознаграждения. Таким образом, действуя в разнообразных терминальных полях, дофамин придает мотивационную значимость («желание») на саму вознаграждение или связанные с ними Помогает консолидировать несколько форм памяти (миндалина и гиппокамп) и кодирует новые моторные программы, которые будут способствовать получению этого вознаграждения в Будущее (ядро accumbers Core Area и дорсальный стриатум). В этом примере дофамин модулирует обработку сенсомоторной информации в различных нейронных целях, чтобы максимизировать способность организма получать будущие вознаграждения. ...
Схема вознаграждения мозга, которая нацелена на наркотики, обычно опосредует удовольствие и укрепление поведения, связанных с естественными подкрепляющими, такими как пища, вода и сексуальный контакт. Дофаминовые нейроны в VTA активируются в пище и воде, а высвобождение дофамина в NAC стимулируется наличием натуральных артистов, таких как пища, вода или сексуальный партнер. ...
NAC и VTA являются центральными компонентами схемы, лежащих в основе вознаграждения и памяти вознаграждения. Как упоминалось ранее, активность дофаминергических нейронов в VTA, по -видимому, связана с прогнозом вознаграждения. NAC участвует в обучении, связанном с подкреплением и модуляцией автомобильных реакций на стимулы, которые удовлетворяют внутренним гомеостатическим потребностям. Оболочка NAC, по -видимому, особенно важна для первоначальных действий лекарств в схеме вознаграждения; Привыкающие препараты, по -видимому, оказывают большее влияние на высвобождение дофамина в оболочке, чем в ядре NAC. - ^ Jump up to: а беременный в MP Saddoris, Cacciapaglia F, Wightman RM, Carelli RM (август 2015 г.). «Дифференциальная динамика высвобождения дофамина в ядре Accumbens Core и Shell выявляет комплементарные сигналы для прогнозирования ошибок и стимулирующей мотивации» . Журнал нейробиологии . 35 (33): 11572–82. doi : 10.1523/jneurosci.2344-15.2015 . PMC 4540796 . PMID 26290234 .
Здесь мы обнаружили, что высвобождение дофамина в реальном времени в ядра Accumbens (первичная мишень дофаминовых нейронов среднего мозга), поразительно изменяется между субрегионами ядра и оболочки. По сути, динамика дофамина согласуется с теориями на основе обучения (таких как ошибка прогнозирования вознаграждений), тогда как в оболочке дофамин согласуется с теориями, основанными на мотивации (например, стимулирующей значимости).
- ^ Jump up to: а беременный в Калипари Э.С., Багот Р.К., Пурушотаман И., Дэвидсон Т.Дж., Йоргасон Дж.Т., Пенья С.Дж., Уокер Д.М., Пирпинис С.Т., Гиз К.Г., Рамакришнан С., Дейссерот К., Нестлер Э.Дж. (март 2016 г.). «Визуализация in vivo идентифицирует височную сигнатуру средних колючих нейронов D1 и D2 в награде кокаина» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (10): 2726–31. Bibcode : 2016pnas..113.2726c . doi : 10.1073/pnas.15212388113 . PMC 4791010 . PMID 26831103 .
Повышенная активность мезолимбической дофаминовой системы является центральным механизмом, лежащим в основе усиления и полезных действий лекарств от злоупотреблений, включая кокаин, а также компульсивное поиск лекарств, которые развиваются с течением времени и характеризуют зависимое состояние (10–12). Действие дофамина в NAC опосредовано преимущественно посредством активации рецепторов D1 или D2 дофамина, которые экспрессируются в значительной степени непересекающимися популяциями средних колючих нейронов (MSN) (13). Эти два подтипа MSN оказывают противоположное влияние на поведение, с оптогенетической активацией нейронов типа D1, способствующих положительному подкреплению и увеличению образования ассоциаций кокаина-вознаграждения и контекста и активации нейронов типа D2, которые являются отвратительными и уменьшающимися вознаграждением кокаина (14, 15). ; Связанные различия в поведенческих реакциях наблюдаются в ответ на агонисты D1 и D2 -рецептора или антагонисты (16). ... Предыдущая работа продемонстрировала, что оптогенетически стимулирующие D1 MSN способствуют вознаграждению, тогда как стимулирование D2 MSN вызывает отвращение.
- ^ Венцель Дж. М., Раушер Н.А., Черт Дж. Ф., Олесон Э.Б. (январь 2015 г.). «Роль фазического высвобождения дофамина в ядре, наполненном кодированием отвращения: обзор нейрохимической литературы» . ACS Химическая нейробиология . 6 (1): 16–26. doi : 10.1021/cn500255p . PMC 5820768 . PMID 25491156 .
Таким образом, вызванные страхом стимулы способны дифференциально изменять передачу фазовой дофамина в субрегионах NACC. Авторы предполагают, что наблюдаемое улучшение в дофамине NACC Shell, вероятно, отражает общую мотивационную значимость, возможно, из-за облегчения от CS-индуцированного состояния страха, когда США (шок) не доставляется. Это рассуждение поддерживается докладом Budygin и Colleags 112 Показывая, что у анестезированных крыс завершение закрепления хвоста приводит к увеличению высвобождения дофамина в оболочке.
- ^ Jump up to: а беременный Malenka RC, Nestler EJ, Hyman SE (2009). «Глава 10: Нейронный и нейроэндокринный контроль внутренней среды». В Сидоре А, Браун Р.Ю. (ред.). Молекулярная нейрофармакология: основание для клинической нейробиологии (2 -е изд.). Нью-Йорк: McGraw-Hill Medical. п. 266. ISBN 978-0-07-148127-4 Полем
Дофамин действует в ядре, прилежащем прикреплять мотивационное значение к стимулам, связанным с вознаграждением.
- ^ Jump up to: а беременный в Саламон Д.Д., Пардо М., Йон С.Е., Лопес-Круз Л., Санмигель Н., Корреа М. (2016). «Мезолимбический дофамин и регуляция мотивированного поведения». Современные темы в Behavoral Neurosciences . 27 : 231–57. Doi : 10.1007/7854_2015_383 . ISBN 978-3-319-26933-7 Полем PMID 26323245 .
Значительные доказательства указывают на то, что accumbens da важен для павловского подхода и павловского в инструментальный перенос [(PIT)] ... Яма-это поведенческий процесс, который отражает влияние стимулов, связанных с Павлованом (CS) на инструментальное реагирование. Например, представление павловского CS в сочетании с пищей может увеличить производство инструментального поведения, армированного пищевыми продуктами, такого как нажатие на рычага. Яма, специфичная для результатов, возникает, когда павловский безусловный стимул (США) и инструментальный усилитель являются одним и тем же стимулом, в то время как общая яма, как говорят, встречается, когда павловские США и подкрепление различаются. ... более поздние данные указывают на то, что прикусывание ядра и оболочка, по -видимому, опосредуют различные аспекты ямы; Повреждения и инактивацию в раковине уменьшили специфическую яму результата, в то время как ядро поражения и инактивация подавляют общую яму (Corbit and Balleine 2011). Эти различия в ядрах и оболочках, вероятно, связаны с различными анатомическими входами и паллидальными выходами, связанными с этими прикульными субрегионами (Root et al. 2015). Эти результаты привели к тому, что Corbit и Balleine (2011) предполагает, что Accupbences Core опосредует общее возбуждающее воздействие подсказок, связанных с вознаграждением. Яма обеспечивает фундаментальный поведенческий процесс, с помощью которого кондиционированные стимулы могут оказывать активирующие эффекты на инструментальный ответ
- ^ Jump up to: а беременный в Corbit LH, Balleine BW (2016). «Обучение и мотивационные процессы, способствующие павлова-инструментальному переносу и их нервным базам: дофамин и за его пределами». Современные темы в поведенческих нейронауках . 27 : 259–89. doi : 10.1007/7854_2015_388 . ISBN 978-3-319-26933-7 Полем PMID 26695169 .
Такие эффекты позволяют предположить, что конкретные мотивационные состояния направляют воздействие процессов стимулов павловских стимулов на инструментальные показатели ... Поведенческие результаты подтверждаются доказательствами того, что различные нейронные цепи, сосредоточенные на ядре NAC и оболочке, опосредуют общие и специфические формы переноса, соответственно и и Непрерывная работа начинает объяснять, как павловские и инструментальные процессы обучения, которые происходят независимо и в отдельные времена интегрируются в нейронные схемы, которые регулируют поведенческий контроль.
- ^ Jump up to: а беременный в дюймовый Cherasse Y, урада Y (ноябрь 2017 г.). «Диетический цинк действует как модулятор сна» . Международный журнал молекулярных наук . 18 (11): 2334. doi : 10.3390/ijms18112334 . PMC 5713303 . PMID 29113075 .
Совсем недавно лаборатория Фуллера также обнаружила, что сон может способствовать активации гамма-аминообутирической кислотной (GABAergic) популяции нейронов, расположенной в парафициальной зоне [11,12], в то время как роль GABAergic A2AR-экспрессирующих нейронов [11,12], в то время как роль GABAergic A2AR-экспрессирующих нейронов из ядра прилегающих [13] и стриатума только что выявлено [14,15].
- ^ Jump up to: а беременный в дюймовый и фон глин Валенсия Гарсия С., Форт П (февраль 2018 г.). «Ядро accumbens, новая область, регулирующая сна посредством интеграции мотивационных стимулов» . Acta Pharmacologica Sinica . 39 (2): 165–166. doi : 10.1038/aps.2017.168 . PMC 5800466 . PMID 29283174 .
Прилегающее ядро содержит контингент нейронов, специально экспрессирующих постсинаптический подтип A2A-рецептора (A2AR), что делает их возбудимыми с помощью аденозина, его естественный агонист, наделенный мощными способностями сна [4]. ... В обоих случаях большая активация A2AR-экспрессирующих нейронов в NAC способствует медленному волновому сону (SWS) за счет увеличения количества и продолжительности эпизодов. ... После оптогенетической активации ядра наблюдалось аналогичное продвижение SWS, тогда как никаких существенных эффектов не было индуцировано при активации A2AR-экспрессирующих нейронов в оболочке.
- ^ Jump up to: а беременный в дюймовый и фон глин Oishi Y, Xu Q, Wang L, Zhang BJ, Takahashi K, Takata Y, Luo YJ, Cherasse Y, Schiffmann SN, De Kerchove D'eSaerde A, Urade Y, QU WM, Huang ZL, Lazarus M (сентябрь 2017 г.). «Медленная волна сон контролируется подмножеством ядра, укупавших ядерных нейронов у мышей» . Природная связь . 8 (1): 734. Bibcode : 2017natco ... 8..734o . doi : 10.1038/s41467-017-00781-4 . PMC 5622037 . PMID 28963505 .
Здесь мы показываем, что химиогенетическая или оптогенетическая активация возбуждающих аденозиновых рецепторов A2A-экспрессирующих нейронов непрямого пути в основной области NAC сильно вызывает сон с медленной волной. Химиогенетическое ингибирование нейронов непрямого пути NAC предотвращает индукцию сна, но не влияет на гомеостатический отскок сна.
- ^ Jump up to: а беременный в Юань Х.С., Ван Л., Донг Х., Куо В.М., Ян С.Р., Черасе Ю., Лазарус М., Шиффманн С.Н., Д'Орде А.К., Ли Ркс, Хуан Зл (Octaber 2017). «2-й рецепторные нейроны контролируют сон с активным периодом посредством нейронов парвальбумина у внешнего глобуса Pallidus» . Элиф . 6 : E29055. Doi : 10.7554/elife.29055 . PMC 5655138 . PMID 29022877 .
- ^ Schwienbacher I, Fendt M, Richardson R, Schnitzler Hu (ноябрь 2004 г.). «Временная инактивация ядра привыкает к приобретению и выражению награжденного страхом у крыс». Исследование мозга . 1027 (1–2): 87–93. doi : 10.1016/j.brainres.2004.08.037 . PMID 15494160 . S2CID 18338111 .
- ^ Jump up to: а беременный Zubieta JK, Stohler CS (март 2009 г.). «Нейробиологические механизмы ответов плацебо» . Анналы нью -йоркской академии наук . 1156 (1): 198–210. BIBCODE : 2009NASA1156..198Z . doi : 10.1111/j.1749-6632.2009.04424.x . PMC 3073412 . PMID 19338509 .
- ^ Басар К., Сезия Т., Гроеневеген Х., Стейнбуш Х.В., Виссер-Вандевелл В., Темель Ю (декабрь 2010 г.). «Ядро прилегает и импульсивность». Прогресс в нейробиологии . 92 (4): 533–57. Doi : 10.1016/j.pneurobio.2010.08.007 . PMID 20831892 . S2CID 16964212 .
- ^ Gipson CD, Kupchik YM, Kalivas PW (январь 2014 г.). «Быстрая, временная синаптическая пластичность в зависимости» . Нейрофармакология . 76 Pt B: 276–86. doi : 10.1016/j.neuropharm.2013.04.032 . PMC 3762905 . PMID 23639436 .
В пределах упрощенной схемы PFC-NAC-VTA NAC служит «шлюзом», через которую информация, касающаяся направления поведенческого вывода, обрабатывается из лимбической коры к моторным подкоркам. Считается, что переход к компульсивному поиску лекарств возникает из-за нарушения способности этого подкорса эффективно обрабатывать информацию о негативных условиях окружающей среды, что приводит к неспособности ингибировать препривоенные лекарственные реакции; Таким образом, наркоман станет подверженным рецидивам.
Рисунок 1: Глутаматергические афференты в ядро привыкают к привыканию к поведению - ^ Jump up to: а беременный в Ягер Л.М., Гарсия А.Ф., Вунш А.М., Фергюсон С.М. (август 2015 г.). «Входы и выходы стриатума: роль в наркомании» . Нейробиология . 301 : 529–541. doi : 10.1016/j.neuroscience.2015.06.033 . PMC 4523218 . PMID 26116518 .
[Стриатум] получает дофаминергические входные данные от вентральной сегментарной области (VTA) и субстанции NIGRA (SNR) и глутаматергических входных данных из нескольких областей, включая кору, гиппокамп, миндалину и таламус (Swanson, 1982; Phillipson and Griffiths, 1985; Финч, 1996; Al., 2012). Эти глутаматергические входы вступают в контакт на головках дендритных шипов в нейронах с колючими проекционными проекционными нейронами (MSN), тогда как синапс дофаминергических входов в модуляцию активности MSN ... Следует также отметить, что в NAC есть небольшая популяция нейронов, которая совместно сочетается с рецепторами D1 и D2, Хотя это в значительной степени ограничено оболочкой NAC (Bertran-Gonzalez et al., 2008). ... нейроны в подразделениях NAC Core и NAC Shell также различаются. Ядро NAC участвует в обработке кондиционированных стимулов, тогда как оболочка NAC более важна в обработке безусловных стимулов; Считается, что эти две полосатые популяции MSN оказывают противоположное воздействие на базальные ганглии. Активация DMSN вызывает чистое возбуждение таламуса, что приводит к положительной петле обратной связи коры; тем самым выступая в качестве сигнала «идти», чтобы инициировать поведение. Активация IMSN, однако, вызывает чистое ингибирование таламической активности, приводящее к негативной цикле коры обратной связи, и, следовательно, служит «тормозом», чтобы ингибировать поведение ... Существует также растущие доказательства того, что IMSN играют роль в мотивации и зависимости ( Lobo and Nestler, 2011; ... Вместе эти данные свидетельствуют о том, что IMSN обычно действуют для сдержания поведения наркотиков, и рекрутирование этих нейронов может фактически защищать от развития компульсивного употребления наркотиков.
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k Робисон AJ, Nestler EJ (октябрь 2011 г.). «Транскрипционные и эпигенетические механизмы зависимости» . Природные обзоры. Нейробиология . 12 (11): 623–37. doi : 10.1038/nrn3111 . PMC 3272277 . PMID 21989194 .
ΔFOSB был непосредственно связан с несколькими поведениями, связанными с зависимостью ... важно, генетическая или вирусная сверхэкспрессия ΔJund, доминирующее негативное мутант jund, который противодействует ΔFOSB- и другим AP-1-опосредованной транскрипционной активности, в NAC или OFC блокирует их. Ключевые эффекты воздействия лекарственного средства 14,22–24 Полем Это указывает на то, что ΔFOSB является одновременно необходимым и достаточным для многих изменений, внесенных в мозг при хроническом воздействии лекарственного средства. ΔFOSB также индуцируется в MSN NAC типа D1 путем хронического потребления нескольких природных вознаграждений, включая сахарозу, пищу с высоким содержанием жира, пол, бег колеса, где это способствует этому потреблению 14,26–30 Полем Это подразумевает ΔFOSB в регуляции естественных вознаграждений в нормальных условиях и, возможно, во время патологических привыкающих состояний. ... 95% нейронов NAC представляют собой GABAergic MSN (средние колючие нейроны), которые могут быть дополнительно дифференцированы в те MSN, которые экспрессируют рецептор дофамина D1 (D1-тип MSN) и экспрессируют динорфин и вещество P и те, которые экспрессируют D2 Dopamine Dopamine). рецептор (D2-тип MSNS) и экспресс-энкефалин 132 Полем Индукция лекарственного средства ΔFOSB 133,134 и влияние ΔFOSB и G9A на морфологию и поведение клеток различается между MSN типа D1 и D2 135 и нейрональная активность этих двух типов клеток вызывает противоположное влияние на полезные свойства кокаина 131 Полем ... Около 1–2% нейронов NAC - это аспипия большие холинергические интернейроны, которые, как было показано, играют важную роль в вознаграждении кокаина 130 и аналогичное число - ГАМКергические интернейроны, функция которых менее хорошо изучена.
- ^ Jump up to: а беременный в дюймовый и Блум К., Вернер Т., Карнс С., Карнс П., Боурат А., Джордано Дж., Оскар-Берман М., Голд М. (2012). «Секс, лекарства и рок -н -ролл: гипотеза общая мезолимбическая активация как функция полиморфизмов генов вознаграждения» . Журнал психоактивных лекарств . 44 (1): 38–55. doi : 10.1080/02791072.2012.662112 . PMC 4040958 . PMID 22641964 .
Было обнаружено, что ген Deltafosb в NAC имеет решающее значение для усиления последствий сексуального вознаграждения. Кувшины и коллеги (2010) сообщили, что было показано, что сексуальный опыт вызывает накопление DeltaFosB в нескольких лимбических областях мозга, включая NAC, медиальную пре-фронтальную кору, VTA, хвостат и путамен, но не в медиальном преоптическом ядре. Затем индукция C-Fos, нижестоящей (подавленной) цели DeltaFosB, была измерена у сексуально опытных и наивных животных. Количество клеток C-FOS-IR, индуцированных спариванием, было значительно снижено у животных, подвергшихся сексуально, по сравнению с сексуально наивными контролями. Наконец, уровни DeltaFoSB и его активность в NAC манипулировали с использованием вирусного переноса генов, чтобы изучить его потенциальную роль в опосредовании сексуального опыта и вызванного опытом облегчения сексуальных результатов. Животные с сверхэкспрессией DeltaFoSB демонстрировали улучшенное облегчение сексуальной эффективности с сексуальным опытом по сравнению с контролем. Напротив, экспрессия Deltajund, доминантно-негативного партнера по связыванию Deltafosb, ослабляющего сексуального опыта, вызванного облегчением сексуальных результатов и задерживало долгосрочное поддержание облегчения по сравнению с группой сверхэкспрессирующих Deltafosb. Вместе эти результаты подтверждают критическую роль для экспрессии DeltaFosB в NAC в усиливающих эффектах сексуального поведения и сексуального опыта, вызванного облегчением сексуальных результатов. ... Как наркомания, так и сексуальная зависимость представляют собой патологические формы нейропластичности наряду с появлением аберрантного поведения, включающего каскад нейрохимических изменений, главным образом в полезных схемах мозга.
- ^ Гото, О'Доннелл П (февраль 2001 г.). «Синхронная активность в гиппокампе и ядре, прилежащем in vivo» . Журнал нейробиологии . 21 (4): RC131. doi : 10.1523/jneurosci.21-04-J0003.2001 . PMC 6762233 . PMID 11160416 .
- ^ Malenka RC, Nestler EJ, Hyman SE (2009). «Глава 6: широко выступающие системы: моноамины, ацетилхолин и орексин». В Сидоре А, Браун Р.Ю. (ред.). Молекулярная нейрофармакология: основание для клинической нейробиологии (2 -е изд.). Нью-Йорк: McGraw-Hill Medical. С. 175–176. ISBN 978-0-07-148127-4 Полем
Внутри мозга гистамин синтезируется исключительно нейронами с их клеточными телами в туберомамиллярном ядре (TMN), которые находятся в заднем гипоталамусе. У людей насчитывается приблизительно 64000 гистаминергических нейронов на сторону. Эти клетки проецируются на протяжении всего мозга и спинного мозга. Области, которые получают особенно плотные проекции, включают кору головного мозга, гиппокамп, неостриатум, прикусы ядра, миндалину и гипоталамус. ... В то время как наиболее охарактеризованной функцией гистаминовой системы в мозге является регуляция сна и возбуждения, гистамин также участвует в обучении и памяти ... также, похоже, гистамин участвует в регуляции кормления и энергетического баланса.
- ^ Барро М, Сесак С.Р., Жорж Ф., Пистис М., Хонг С., Джхоу Т.К. (октябрь 2012 г.). «Тормозные дофаминовые системы: новая главная структура ГАМК для мезолимбических и нигростриатальных функций» . Журнал нейробиологии . 32 (41): 14094–101. doi : 10.1523/jneurosci.3370-12.2012 . PMC 3513755 . PMID 23055478 .
- ^ Ferré S, Lluís C, Justinova Z, Quiroz C, Orru M, Navarro G, Canela EI, Franco R, Goldberg SR (июнь 2010 г.). «Аденозин-каннабиноидные рецепторные взаимодействия. Последствия для стриатальной функции» . Бренд J. Pharmacol . 160 (3): 443–453. doi : 10.1111/j.1476-5381.2010.00723.x . PMC 2931547 . PMID 20590556 .
Два класса MSN, которые гомогенно распределены в стриатуме, могут быть дифференцированы по их выходной связности и их экспрессии рецепторов дофамина и аденозина и нейропептидов. В дорсальном стриатуме (в основном представленным ядром каудат-путамен), энкефалинергические MSN соединяют стриатум с Globus pallidus (латеральный глобус Pallidus) и экспрессируют пептидный энкефалин и высокую плотность допамина D2 и аденозин A2 (они также экспрессируют аденозин. Рецепторы A1), в то время как динорфинергические MSN соединяют Стриатум с черной субстанции (PARS Compacta и Reticulata) и Entopeduncular ядра (Medial Globus pallidus) и экспрессируют пептиды динорфин и вещество P и дофамин D1 и аденозин A1, но не A2A -рецепторы ... эти два различных фенотипа MSN также присутствуют. в вентральном стриатуме (в основном представленным прилежащим ядром и обонятельным бугорку). Однако, хотя они фенотипически равны своим дорсальным аналогам, они имеют некоторые различия с точки зрения подключения. Во -первых, не только энкефалинергические, но и динорфинергические MSN Проецируют вентральный аналог латерального глобуса Pallidus, вентрального паллидума, который, на самом деле, имеет характеристики как бокового, так и медиального глобуса в его афферентной и эфферентной связности. В дополнение к вентральному паллидуму, медиальный глобус Pallidus и субстанция Nigra-VTA, вентральный стриатум посылает проекции в расширенную миндалину, латеральный гипоталамус и педункулопонтиновый тегментальный ядро. ... Также важно упомянуть, что небольшой процент MSN имеет смешанный фенотип и экспрессирует рецепторы D1 и D2 (Surmeier et al., 1996).
- ^ Nishi A, Kuroiwa M, Shoto T (июль 2011 г.). «Механизмы для модуляции передачи сигналов рецептора дофамина D (1) в стриатальных нейронах» . Передний нейроанат . 5 : 43. DOI : 10.3389/fnana.2011.00043 . PMC 3140648 . PMID 21811441 .
Дофамин играет критическую роль в регуляции психомоторных функций в мозге (Bromberg-Martin et al., 2010; Cools, 2011; Gerfen and Surmeier, 2011). Рецепторы дофамина представляют собой суперсемейство рецепторов, связанных с гептогилическим G-белком, и сгруппированы в две категории, D1-подобные (D1, D5) и D2-подобные (D2, D3, D4) рецепторы, основанные на функциональных свойствах, чтобы стимулировать аденилил циклаза (AC) через GS/OLF и ингибировать AC через GI/O, соответственно ... было продемонстрировано, что Рецепторы D1 образуют гетероолигомер с рецепторами D2 и что гетероолигомер рецептора D1-D2 предпочтительно сочетается с передачей сигналов GQ/PLC (Rashid et al., 2007a, b). Экспрессия рецепторов дофамина D1 и D2 в значительной степени разделена в нейронах прямого и косвенного пути в дорсальном стриатуме соответственно (Gerfen et al., 1990; Hersch et al., 1995; Heiman et al., 2008). Тем не менее, некоторые доли средних колючих нейронов, как известно, экспрессируют как D1, так и D2 -рецепторы (Hersch et al., 1995). Анализ экспрессии генов с использованием методики ОТ-ПЦР с одной клеток показал, что 40% средних колючих нейронов экспрессируют мРНК рецептора D1 и D2 (Surmeier et al., 1996).
- ^ Jump up to: а беременный в дюймовый и фон Shirayama Y, Chaki S (октябрь 2006 г.). «Нейрохимия ядра прилегает и его отношение к депрессии и антидепрессантным действиям у грызунов» . Текущая нейрофармакология . 4 (4): 277–91. doi : 10.2174/157015906778520773 . PMC 2475798 . PMID 18654637 .
- ^ Мередит Г.Е., Аголия Р., Артс М.П., Гроеневен Х.Дж., Захм Д.С. (сентябрь 1992 г.). «Морфологические различия между проекционными нейронами ядра и оболочкой в прилегании ядра крысы». Нейробиология . 50 (1): 149–62. doi : 10.1016/0306-4522 (92) 90389-J . PMID 1383869 . S2CID 11278705 .
- ^ Jump up to: а беременный в Мередит Г.Е., Пеннарц С.М. , Гроеневен Х.Дж. (1993). «Клеточная структура для химической передачи сигналов в прилегании ядра». Химическая передача сигналов в базальных ганглиях . Прогресс в исследовании мозга. Тол. 99. С. 3–24. doi : 10.1016/s0079-6123 (08) 61335-7 . ISBN 978-0-444-81562-0 Полем PMID 7906426 .
- ^ Jump up to: а беременный в дюймовый Берридж К.С., Крингельбах М.Л. (май 2015). «Системы удовольствия в мозге» . Нейрон . 86 (3): 646–64. doi : 10.1016/j.neuron.2015.02.018 . PMC 4425246 . PMID 25950633 .
- ^ Балики М.Н., Мансур А., Бария А.Т., Хуан Л., Бергер С.Е., Филдс Х.Л., Апкирян Ав (октябрь 2013 г.). «Размещение человека укупается в предполагаемое ядро и раковину диссоциирует кодирование ценностей для вознаграждения и боли» . Журнал нейробиологии . 33 (41): 16383–93. doi : 10.1523/jneurosci.1731-13.2013 . PMC 3792469 . PMID 24107968 .
Недавние данные указывают на то, что инактивация рецепторов D2 в непрямом пути стриатопаллидного пути у грызунов необходима как для приобретения, так и для экспрессии аверсивного поведения, а также прямого пути рецептора D1 контролирует обучение на основе вознаграждения (Hikida et al., 2010; Hikida et al. ., 2013). Похоже, мы можем сделать вывод, что прямые и косвенные пути NAC через рецепторы D1 и D2 сохраняют различные роли в ожидании и оценку в оболочке и ядро NAC, что согласуется с наблюдениями относительно пространственной сегрегации и разнообразия реакций дофаминергических нейронов среднего мозга. Для вознаграждения и неприятных условий, некоторая кодирующая мотивационная ценность, другие мотивационные значения, каждый из которых связан с отдельными сетями мозга и имеет различные Роли в мотивационном контроле (Bromberg-Martin et al., 2010; Cohen et al., 2012; Lammel et al., 2013). ... Таким образом, предыдущие результаты, в сочетании с текущими наблюдениями, подразумевают, что отклик NAC Pshell отражает прогнозирование/предвкушение или сигнал значимости, а реакция NAC PCORE - это оценка отклика (сигнал прогнозирования вознаграждения), который сигнализирует о отрицательном значении подкрепления о прекращении боли (то есть ожидаемой анальгезии).
- ^ Jump up to: а беременный Картони Э., Пуглиси-Аллегра С., Бальдассарр Г. (ноябрь 2013). «Три принципа действий: гипотеза павлова-инструментального переноса» . Границы в поведенческой нейробиологии . 7 : 153. doi : 10.3389/fnbeh.2013.00153 . PMC 3832805 . PMID 24312025 .
- ^ Ричард Дж. М., Кастро, округ Колумбия, Difeliceantonio AG, Robinson MJ, Berridge KC (ноябрь 2013 г.). «Сопоставление мозговых цепей вознаграждения и мотивации: по стопам Энн Келли» . Нейробиологии и биобиоэвиральные обзоры . 37 (9 пт а): 1919–31. doi : 10.1016/j.neubiorev.2012.12.008 . PMC 3706488 . PMID 23261404 .
- ^ Берридж К.С., Робинсон Т.Е., Олдридж Дж.В. (февраль 2009 г.). «Расширение компонентов вознаграждения:« Любие »,« Желание »и обучение» . Современное мнение о фармакологии . 9 (1): 65–73. doi : 10.1016/j.coph.2008.12.014 . PMC 2756052 . PMID 19162544 .
- ^ Jump up to: а беременный Nestler EJ (декабрь 2013 г.). «Сотовая основа памяти для зависимости» . Диалоги в клинической нейробиологии . 15 (4): 431–43. doi : 10.31887/dcns.2013.15.4/enestler . PMC 3898681 . PMID 24459410 .
Несмотря на важность многочисленных психосоциальных факторов, в своей основе наркомания включает в себя биологический процесс: способность повторного воздействия лекарства от злоупотреблений вызывать изменения в уязвимом мозге, который стимулирует навязчивое поиск и принятие лекарств, и потеря контроля За употреблением наркотиков это определяет состояние зависимости. ... Большая литература продемонстрировала, что такая индукция ΔFOSB в нейронах NAC типа D1 повышает чувствительность животного к лекарственным средствам, а также естественные вознаграждения и способствует самостоятельному применению лекарств, предположительно благодаря процессу положительного подкрепления ... например , оболочка и основные субрегионы NAC демонстрируют различия в синаптической пластичности, вызванной лекарственным средством, как D1- против D2-типа колючих нейронов внутри каждого субрегион. 60,63,64,67
- ^ Dumitriu D, Laplant Q, Grossman YS, Dias C, Janssen WG, Russo SJ, Morrison JH, Nestler EJ (май 2012 г.). «Субрегиональный, дендритный компартмент и специфичность подтипа позвоночника в регуляции кокаина дендритных шипов в прилегании ядра» . Журнал нейробиологии . 32 (20): 6957–66. doi : 10.1523/jneurosci.5718-11.2012 . PMC 3360066 . PMID 22593064 .
Непреходящее изменение плотности позвоночника в сердечнике, но не оболочке хорошо соответствует установленной идее о том, что оболочка преимущественно участвует в развитии зависимости, в то время как ядро опосредует долгосрочное выполнение обученного поведения, связанного с зависимостью (Ito et al., 2004 Di Chiara, 2002; В соответствии с идеей NAC Core является локусом длительной лекарственной индуцированной нейропластичности, несколько исследований показали, что электрофизиологические изменения в сердечнике сохраняются дольше, чем их аналоги Shell. ... Кроме того, данные, представленные здесь, подтверждают идею о том, что NAC Shell преимущественно участвует в немедленном вознаграждении лекарств, в то время как ядро может сыграть более явную роль в долгосрочных аспектах зависимости.
- ^ Pontieri FE, Tanda G, Di Chiara G (декабрь 1995 г.). «Внутривенный кокаин, морфин и амфетамин преимущественно увеличивают внеклеточный дофамин в« оболочке »по сравнению с« ядро »прилегающего ядра крыс» . Труды Национальной академии наук Соединенных Штатов Америки . 92 (26): 12304–8. Bibcode : 1995pnas ... 9212304p . doi : 10.1073/pnas.92.26.12304 . JSTOR 2369093 . PMC 40345 . PMID 8618890 .
- ^ Jump up to: а беременный в Ди Чиара Г (декабрь 2002 г.). «Ядро прилегание оболочки и ядро дофамин: дифференциальная роль в поведении и зависимости». Поведенческое исследование мозга . 137 (1–2): 75–114. doi : 10.1016/s0166-4328 (02) 00286-3 . PMID 12445717 . S2CID 54410118 .
- ^ Эйден Ле, Вейхе Е (январь 2011 г.). «VMAT2: динамический регулятор моноаминергической нейрональной функции мозга, взаимодействующих с наркотиками, злоупотребляющими» . Анналы нью -йоркской академии наук . 1216 (1): 86–98. BIBCODE : 2011NASA1216 ... 86E . doi : 10.1111/j.1749-6632.2010.05906.x . PMC 4183197 . PMID 21272013 .
VMAT2 является везикулярным транспортером ЦНС не только для биогенных аминов DA, NE, EPI, 5-HT и HIS, но, вероятно, также для следовых аминов Tyr, PEA и тиронамин (THYR) ... [следа аминергических] нейронов в нейронах в ЦНС млекопитающих будет идентифицируется как нейроны, экспрессирующие VMAT2 для хранения, и биосинтетический фермент ароматическая аминокислота декарбоксилаза (AADC).
- ^ Барро М, Маринелли М, Аббукс Д.Н., Руге-Пон Ф., Ле Моал М., Пьяцца П.В. (март 2000 г.). «Дофаминергическая гипер-чувствительность оболочки ядра прилегает гормон-зависимость». Европейский журнал нейробиологии . 12 (3): 973–9. doi : 10.1046/j.1460-9568.2000.00996.x . PMID 10762327 . S2CID 6111417 .
- ^ Akiyama G, Ikeda H, Matsuzaki S, Sato M, Moribe S, Koshikawa N, Cools AR (июнь 2004 г.). «Рецепторы GABAA и GABAB в раковине Accumbens Deculas Дифференциально модулируют дофамин и опосредованное рецептором-рецепторным поведением рецептора». Нейрофармакология . 46 (8): 1082–8. doi : 10.1016/j.neuropharm.2004.02.007 . PMID 15111014 . S2CID 25631633 .
- ^ Смит-Рол С.Л., Садегян К., Келли А.Е. (август 1999 г.). «Пространственное обучение и производительность в лабиринте радиального рук нарушаются после блокады рецептора N-метил-D-аспартата (NMDA) в стриатальных субрегионах». Поведенческая нейробиология . 113 (4): 703–17. doi : 10.1037/0735-7044.113.4.703 . PMID 10495079 .
- ^ Giertler C, Bohn I, Hauber W (март 2005 г.). «Участие NMDA и AMPA/KA-рецепторов в ядре прилегает ядро в инструментальном обучении, руководствуясь вознагражденными сигналами». Европейский журнал нейробиологии . 21 (6): 1689–702. doi : 10.1111/j.1460-9568.2005.03983.x . PMID 15845096 . S2CID 14206373 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. Olsen CM (декабрь 2011 г.). «Естественные награды, нейропластичность и не лекарства» . Нейрофармакология . 61 (7): 1109–22. doi : 10.1016/j.neuropharm.2011.03.010 . PMC 3139704 . PMID 21459101 .
Поперечная сенсибилизация также является двунаправленной, так как история введения амфетамина способствует сексуальному поведению и усиливает связанное увеличение NAC DA ... Как описано для вознаграждения за пищу, сексуальный опыт также может привести к активации сигнальных каскадов, связанных с пластичностью. Дельта транскрипционного фактора FOSB увеличивается в NAC, PFC, дорсальном стриатуме и VTA после повторного сексуального поведения (Wallace et al., 2008; Pithers et al., 2010b). Это естественное увеличение дельта -FOSB или избыточной экспрессии Delta FOSB в рамках NAC модулирует сексуальную эффективность, а блокада NAC Delta FOSB ослабляет это поведение (Hedges et al., 2009; Piters et al., 2010b). Кроме того, вирусная сверхэкспрессия Delta FOSB усиливает условное предпочтение среды для окружающей среды, в сочетании с сексуальным опытом (Hedges et al., 2009). ...
Таблица 1 - ^ День JJ, Carelli RM (апрель 2007 г.). «Ядро прилегает и павловийское вознаграждение» . Нейробиолог . 13 (2): 148–59. doi : 10.1177/1073858406295854 . PMC 3130622 . PMID 17404375 .
В соответствии с другими сообщениями (Nicola и др. 2004; Taha and Fields 2006), преобладающим ответом нейронов NAC на инфузии сахарозы было снижение активности (рис. 2). Как видно на рисунке 2, те же нейроны демонстрировали противоположные ответы, когда агрессивный раствор хинина был доставлен внутриорологически. Одна гипотеза предполагает, что ингибирования, наблюдаемые во время доставки вознаграждений, происходят среди нейронов NAC, содержащих ГАМК, которые проецируются на важные моторные области, такие как вентральный паллидум (VP).
- ^ Carlezon WA, Thomas MJ (2009). «Биологические субстраты вознаграждения и отвращения: гипотеза активности ядра прикуса» . Нейрофармакология . 56 (Suppl 1): 122–32. doi : 10.1016/j.neuropharm.2008.06.075 . PMC 2635333 . PMID 18675281 .
При рассмотрении вместе, эти исследования предоставили два критических доказательства, которые сыграли заметную роль в составлении нашей нынешней рабочей гипотезы: во-первых, что дофаминозависимое вознаграждение ослабляется блокадой D2-подобных рецепторов, которые являются ингибирующими рецепторами, экспрессируемыми в основном экспрессируемыми в NAC на MSN непрямого пути; и во-вторых, эти события, которые, как ожидается, снизит общую возбудимость NAC (например, стимуляция опиоидных рецепторов, связанных с GI, снижение стимуляции возбуждающих NMDA-рецепторов, снижение возбуждающего ввода), достаточны для вознаграждения. Эта интерпретация привела к разработке модели вознаграждения, в которой критическим событием снижается активация MSN в NAC
- ^ Коста В.Д., Ланг П.Дж., Сабатинелли Д., Версаче Ф., Брэдли М.М. (сентябрь 2010 г.). «Эмоциональные образы: оценка удовольствия и возбуждения в схеме вознаграждения мозга» . Картирование человеческого мозга . 31 (9): 1446–57. doi : 10.1002/hbm.20948 . PMC 3620013 . PMID 20127869 .
- ^ Sabatinelli D, Bradley MM, Lang PJ, Costa VD, Versace F (сентябрь 2007 г.). «Удовольствие, а не значимость активирует укусывание ядра человека и медиальную префронтальную кору». Журнал нейрофизиологии . 98 (3): 1374–9. doi : 10.1152/jn.00230.2007 . PMID 17596422 . S2CID 14938485 .
- ^ Могенсон Г.Дж., Джонс Д.Л., Им Сай (1980). «От мотивации к действию: функциональный интерфейс между лимбической системой и моторной системой». Прогресс в нейробиологии . 14 (2–3): 69–97. doi : 10.1016/0301-0082 (80) 90018-0 . PMID 6999537 . S2CID 14029796 .
- ^ Hart G, Leung Bk, Balleine BW (февраль 2014 г.). «Спинные и вентральные потоки: четкая роль стриатальных субрегионов в приобретении и выполнении целенаправленных действий» . Нейробиология обучения и памяти . 108 : 104–18. doi : 10.1016/j.nlm.2013.11.003 . PMC 4661143 . PMID 24231424 .
- ^ Castro DC, Cole SL, Berridge KC (2015). «Боковой гипоталамус, прилежащие ядра и роли вентрального паллидума в еде и голоде: взаимодействие между гомеостатическими и вознагражденными схемами» . Границы в системах нейробиологии . 9 : 90. DOI : 10.3389/fnsys.2015.00090 . PMC 4466441 . PMID 26124708 .
- ^ Берридж К.С., Крингельбах М.Л. (июнь 2013 г.). «Нейронаука аффекта: мозговые механизмы удовольствия и неудовольствия» . Современное мнение о нейробиологии . 23 (3): 294–303. doi : 10.1016/j.conb.2013.01.017 . PMC 3644539 . PMID 23375169 .
- ^ Инь Х.Х., Остлунд С.Б., Баллейн Б.В. (октябрь 2008 г.). «Обучение под руководством вознаграждений за пределами дофамина в прилежащем ядре: интегративные функции сетей кортико-басальных ганглиев» . Европейский журнал нейробиологии . 28 (8): 1437–48. doi : 10.1111/j.1460-9568.2008.06422.x . PMC 2756656 . PMID 18793321 .
- ^ Soares-Cunha C, Coimbra B, Sousa N, Rodrigues AJ (сентябрь 2016 г.). «Переосмысление стриатальных D1- и D2-нейронов в награде и отвращении» (PDF) . Нейробиологии и биобиоэвиральные обзоры . 68 : 370–386. doi : 10.1016/j.neubiorev.2016.05.021 . HDL : 1822/47044 . PMID 27235078 . S2CID 207092810 .
- ^ Soares-Cunha C, Coimbra B, Domingues AV, Vasconcelos N, Sousa N, Rodrigues AJ (19 апреля 2018 года). «Ядро accumbens microcircuit, лежащее в основе D2-MSN, повышение мотивации» . Eneuro . 5 (2): Eneuro.0386–18.2018. doi : 10.1523/eneuro.0386-18.2018 . PMC 5957524 . PMID 29780881 .
Оптогенетическая активация D2-MSN снижала активность вентрального паллидума (VP), снижая ингибирующий тонус до VTA, что привело к повышению дофаминергической активности. Важно отметить, что оптогенетическая активация терминалов D2-MSN в VP была достаточной для повторного использования повышения мотивации
- ^ Ferris CF, Kulkarni P, Sullivan JM, Harder JA, Messenger TL, Febo M (январь 2005 г.). «Suckling Suckling более полезен, чем кокаин: доказательства функциональной магнитно-резонансной томографии и трехмерного вычислительного анализа» . Журнал нейробиологии . 25 (1): 149–56. Arxiv : 1510.02343 . doi : 10.1523/jneurosci.3156-04.2005 . PMC 6725197 . PMID 15634776 .
- ^ Numan M (январь 2007 г.). «Мотивационные системы и нейронная схема материнского поведения у крысы» . Психобиология развития . 49 (1): 12–21. doi : 10.1002/dev.20198 . PMID 17186513 .
- ^ Glocker ML, Langleben DD, Ruparel K, Loughead JW, Valdez JN, Griffin MD, Sachser N, Gur RC (июнь 2009 г.). «Детская схема модулирует систему вознаграждений мозга у нулипарных женщин» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (22): 9115–9. Bibcode : 2009pnas..106.9115G . doi : 10.1073/pnas.0811620106 . JSTOR 40482823 . PMC 2690007 . PMID 19451625 .
- ^ Jump up to: а беременный Hyman SE, Malenka RC, Nestler EJ (2006). «Нейронные механизмы зависимости: роль обучения и памяти, связанного с вознаграждением». Ежегодный обзор нейробиологии . 29 : 565–98. doi : 10.1146/annurev.neuro.29.051605.113009 . PMID 16776597 .
- ^ Штайнер Х, Ван Уэз В. (январь 2013 г.). «Регуляция генов, связанная с зависимостью: риск воздействия когнитивных усилителей по сравнению с другими психостимулянтами» . Прогресс в нейробиологии . 100 : 60–80. doi : 10.1016/j.pneurobio.2012.10.001 . PMC 3525776 . PMID 23085425 .
- ^ Jump up to: а беременный Ruffle JK (ноябрь 2014). «Молекулярная нейробиология зависимости: о чем все (δ) FOSB?». Американский журнал по вопросам злоупотребления наркотиками и алкоголем . 40 (6): 428–37. doi : 10.3109/00952990.2014.933840 . PMID 25083822 . S2CID 19157711 .
ΔFOSB является важным транскрипционным фактором, участвующим в молекулярных и поведенческих путях зависимости после повторного воздействия лекарственного средства. Образование ΔFOSB в нескольких областях мозга, и молекулярный путь, ведущий к образованию комплексов AP-1, хорошо понятно. Создание функциональной цели для ΔFOSB позволило дополнительно определить некоторые из ключевых аспектов его молекулярных каскадов, включающих такие эффекторы, как GluR2 (87,88), CDK5 (93) и NFKB (100). Более того, многие из этих идентифицированных молекулярных изменений в настоящее время напрямую связаны с структурными, физиологическими и поведенческими изменениями, наблюдаемыми после хронического воздействия лекарственного средства (60,95,97,102). Новые границы исследований, изучающих молекулярные роли ΔFOSB, были открыты эпигенетическими исследованиями, и последние достижения иллюстрировали роль ΔFOSB, действующего на ДНК и гистоны, действительно как Молекулярный переключатель (34).
- ^ Канехиса лаборатории (29 октября 2014 г.). «Алкоголизм - Homo Sapiens (человек)» . Кегг путь . Получено 31 октября 2014 года .
- ^ Ким Й., Тейлан М.А., Барон М., Сэндс А., Нэрн А.С., Грингард П (февраль 2009 г.). «Индуцированное метилфенидатом образование дендритного позвоночника и экспрессия DeltaFosB в прилежащем ядре» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (8): 2915–20. Bibcode : 2009pnas..106.2915K . doi : 10.1073/pnas.0813179106 . PMC 2650365 . PMID 19202072 .
- ^ Кувшины KK, Vialou V, Nestler EJ, Laviolette SR, Lehman MN, Coolen LM (февраль 2013 г.). «Природные и лекарственные вознаграждения действуют на общие механизмы нейронной пластичности с ΔFOSB в качестве ключевого медиатора» . Журнал нейробиологии . 33 (8): 3434–42. doi : 10.1523/jneurosci.4881-12.2013 . PMC 3865508 . PMID 23426671 .
Препараты злоупотребления индуцируют нейропластичность в пути естественного вознаграждения, в частности, ядра (NAC), что вызывает развитие и экспрессию привыкания. ... Вместе эти результаты демонстрируют, что лекарства от злоупотребления и природного поведения вознаграждения действуют на общие молекулярные и клеточные механизмы пластичности, которые контролируют уязвимость к наркомании, и что эта повышенная уязвимость опосредована ΔFOSB и его нижестоящими транскрипционными целями. ... сексуальное поведение очень полезно (Tenk et al., 2009), а сексуальный опыт вызывает сенсибилизированное поведение, связанное с наркотиками, включая перекрестную сенсибилизацию к амфетаминам (AMPH), индуцированной локомоторной активностью (Bradley and Meisel, 2001; Piterers et al. ., 2010a) и улучшенное вознаграждение AMPH (Pithers et al., 2010a). Кроме того, сексуальный опыт вызывает нервную пластичность в NAC, аналогичную тем, что вызвано психостимулянтным воздействием, включая повышенную плотность дендритного позвоночника (Meisel and Mullins, 2006; Piterers et al., 2010a), измененный перевозка рецептора глутамата и снижение синоптической силы в префронтальной коре). - -отвечающие нейроны NAC Shell (Piterers et al., 2012). Наконец, периоды воздержания от сексуального опыта были обнаружены решающими для усиленного вознаграждения AMPH, Spinogenesis NAC (Pithers et al., 2010a) и транспорта рецепторов глутамата (Pithers et al., 2012). Эти результаты показывают, что природное и лекарственное вознаграждение испытывает общие механизмы нейронной пластичности
- ^ Мозговые электроды помогают лечить депрессию, архивируя 16 марта 2012 года в The Wayback Machine , Technology Review , 26 апреля 2007 г.
- ^ Bewernick BH, Hurlemann R, Matusch A, Kayser S, Grubert C, Hadrysiewicz B, Axmacher N, Lemke M, Cooper-Mahkorn D, Cohen MX, Brockmann H, Lenartz D, Sturm V, Schlaepfer Te (январь 2010 г.). «Ядро присыпает глубокую стимуляцию мозга снижает оценки депрессии и тревоги при устойчивой к лечению депрессии». Биологическая психиатрия . 67 (2): 110–6. doi : 10.1016/j.biopsych.2009.09.013 . PMID 19914605 . S2CID 26242126 .
- ^ OOMS P, Mantione M, Figee M, Schuurman PR, Van Den Munckhof P, Denys D (февраль 2014 г.). «Глубокая стимуляция мозга при обсессивно-компульсивных расстройствах: долгосрочный анализ качества жизни». Журнал неврологии, нейрохирургии и психиатрии . 85 (2): 153–8. doi : 10.1136/jnnp-2012-302550 . PMID 23715912 . S2CID 54572260 .
- ^ «Спорный хирургический вмешательство по поводу наркомании сгорает центр удовольствий мозга», автор Maia Szalavitz. 13 декабря 2012 г.
- ^ «Китай запрещает необратимую процедуру мозга» автор Zamiska Nicholas. 28 апреля 2008 года. The Wall Street Journal
- ^ Скотт DJ, Stohler CS, Egnatuk CM, Wang H, Koeppe RA, Zubieta JK (июль 2007 г.). «Индивидуальные различия в реагировании на вознаграждение объясняют плацебо-индуцированные ожидания и эффекты» . Нейрон . 55 (2): 325–36. doi : 10.1016/j.neuron.2007.06.028 . PMID 17640532 .
- «Область мозга центральная для эффекта плацебо выявлена» . Эврикалерт! (Пресс-релиз). 18 июля 2007 г.
Внешние ссылки
[ редактировать ]- Роль ядра приходится в цепи вознаграждения. Часть «мозг сверху донизу». на Brain.mcgill.ca
- Accumbens ядра - база данных с центром клеток
- Окрашенные изображения срезов мозга, которые включают в себя «ядро%20 -аккумбобы» в проекте Brainmaps