Метаболизм наркотиков
Метаболизм наркотиков это метаболический распад наркотиков - живыми организмами , обычно с помощью специализированных ферментативных систем. В более общем смысле ксенобиотическое метаболизм (от греческого ксеноса «незнакомца» и биотического «связанного с живыми существами») - это набор метаболических путей , которые модифицируют химическую структуру ксенобиотиков , которые являются соединениями, чуждо для нормальной биохимии организма, например, любое лекарство, такое как любое лекарство . или яд . Эти пути являются формой биотрансформации , присутствующей во всех основных группах организмов, и считаются древним происхождением. Эти реакции часто действуют для детоксикации ядовитых соединений (хотя в некоторых случаях промежуточные соединения в ксенобиотическом метаболизме сами могут вызывать токсические эффекты). Изучение метаболизма лекарств является объектом фармакокинетики . Метаболизм является одним из стадий (см. Adme ) транзита препарата через организм, который включает в себя разрушение препарата, чтобы его можно было выделить организм.
Метаболизм фармацевтических препаратов является важным аспектом фармакологии и медицины . Например, скорость метаболизма определяет продолжительность и интенсивность фармакологического действия препарата. Метаболизм лекарственного средства также влияет на устойчивость к мультираге при инфекционных заболеваниях и химиотерапии при раке , а действие некоторых препаратов в качестве субстратов или ингибиторов ферментов, участвующих в ксенобиотическом метаболизме, являются общей причиной взаимодействия опасных лекарств . Эти пути также важны в науке о окружающей среде , с ксенобиотическим метаболизмом микроорганизмов, определяющих, будет ли загрязнитель разбит во время биоремедиации или сохраняется в окружающей среде. Ферменты ксенобиотического метаболизма, в частности, глутатион S-трансферазы также важны в сельском хозяйстве, поскольку они могут вызывать устойчивость к пестицидам и гербицидам .
Метаболизм наркотиков разделен на три этапа. На фазе I ферменты, такие как цитохром P450 оксидазы, вводят реактивные или полярные группы в ксенобиотики. Эти модифицированные соединения затем конъюгируют с полярными соединениями в реакциях фазы II. Эти реакции катализируются трансферовыми ферментами, такими как глутатион S-трансферазы. Наконец, на фазе III конъюгированные ксенобиотики могут быть дополнительно обработаны, прежде чем быть распознаваемыми транспортерами оттока и выкачаются из клеток. Метаболизм лекарств часто превращает липофильные соединения в гидрофильные продукты, которые более легко выделяются . [ Цитация необходима ]
Барьеры проницаемости и детоксикация
[ редактировать ]Точные соединения, на которые подвергается организму, будут в значительной степени непредсказуемы и могут сильно различаться с течением времени; Это основные характеристики ксенобиотического токсического стресса. [ 1 ] Основная проблема, с которой сталкиваются системы детоксикации ксенобиотиков, заключается в том, что они должны быть в состоянии удалить почти ограниченное количество ксенобиотических соединений из сложной смеси химических веществ, участвующих в нормальном метаболизме . Решение, которое развивалось для решения этой проблемы, представляет собой элегантную комбинацию физических барьеров и ферментативных систем с низкой специфичностью.
Все организмы используют клеточные мембраны в качестве барьеров гидрофобной проницаемости для контроля доступа к их внутренней среде. Полярные соединения не могут диффундировать по этим клеточным мембранам, а поглощение полезных молекул опосредовано через транспортные белки , которые специально выбирают субстраты из внеклеточной смеси. Это селективное поглощение означает, что большинство гидрофильных молекул не могут войти в клетки, поскольку они не распознаются какими -либо конкретными транспортерами. [ 2 ] Напротив, диффузия гидрофобных соединений в этих барьерах не может контролироваться, и, следовательно, организмы не могут исключать липидные ксенобиотики с использованием мембранных барьеров.
Тем не менее, существование барьера проницаемости означает, что организмы были способны развивать системы детоксикации, которые используют гидрофобность, общую для проницательности мембраны ксенобиотиков. Таким образом, эти системы решают проблему специфичности, обладая такой широкой субстратной специфичностью, что они усваивают практически любое неполярное соединение. [ 1 ] Полезные метаболиты исключены, поскольку они полярны, и в целом содержат одну или несколько заряженных групп.
Детоксикация реактивных побочных продуктов нормального метаболизма не может быть достигнута с помощью систем, изложенных выше, поскольку эти виды получены из нормальных клеточных составляющих и обычно имеют свои полярные характеристики. Однако, поскольку эти соединения мало в количестве, конкретные ферменты могут распознавать и удалять их. Примерами этих специфических систем детоксикации являются система глиоксалазы , которая удаляет реактивную альдегид -метилглаксаль, [ 3 ] и различные антиоксидантные системы, которые устраняют реактивные формы кислорода . [ 4 ]
Фазы детоксикации
[ редактировать ]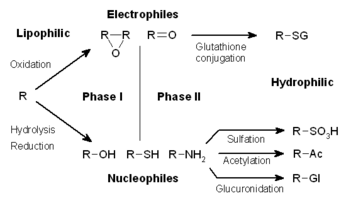
Метаболизм ксенобиотиков часто делится на три фазы: модификация, конъюгация и экскреция. Эти реакции действуют согласованно, чтобы детоксифицировать ксенобиотики и удалить их из клеток.
Фаза I - Модификация
[ редактировать ]На фазе I различные ферменты действуют для введения реактивных и полярных групп в свои субстраты. Одним из наиболее распространенных модификаций является гидроксилирование, катализируемое цитохромовой P-450-зависимой системой смешанной оксидазы смешанной функции . Эти ферментные комплексы действуют для включения атома кислорода в неактивированные углеводороды, что может привести к введению гидроксильных групп или N-, O- и S-диалкилирования субстратов. [ 5 ] Механизм реакции P-450 оксидаз проходит через восстановление цитохромового кислорода и генерации высокореактивных видов оксиферрила, согласно следующей схеме: [ 6 ]
- O 2 + nadph + h + + RH → NADP + + H 2 O + RAW
Реакции фазы I (также называемые несинтетическими реакциями) могут происходить путем окисления , восстановления , гидролиза , циклизации , дециклирования и добавления кислорода или удаления водорода, выполняемых с помощью смешанных оксидаз, часто в печени. Эти окислительные реакции обычно включают цитохромум монооксигеназу цитохрома P450 (часто сокращенная CYP), NADPH и кислород. Классы фармацевтических препаратов, которые используют этот метод для их метаболизма, включают фенотиазины , парацетамол и стероиды. Если метаболиты реакций фазы I достаточно полярно, они могут быть легко выделены на данный момент. Тем не менее, многие продукты фазы I не устраняются быстро и подвергаются последующей реакции, в которой эндогенный субстрат в сочетании с вновь включенной функциональной группой с образованием высокополярного конъюгата.
Общее окисление фазы I включает преобразование связи CH в C-OH. Эта реакция иногда превращает фармакологически неактивное соединение ( пролекарство ) в фармакологически активное. Точно так же, фаза I может превратить нетоксичную молекулу в ядовитую ( токсификацию ). Простой гидролиз в желудке обычно является безобидной реакцией, однако есть исключения. Например, метаболизм фазы I превращает ацетонитрил в хох 2 CN, который быстро диссоциирует в формальдегид и цианид водорода . [ 7 ]
Фаза I Метаболизм кандидатов на лекарства может быть смоделирован в лаборатории с использованием негменных катализаторов. [ 8 ] Этот пример биомиметической реакции имеет тенденцию давать продукты, которые часто содержат метаболиты фазы I. Например, основной метаболит фармацевтического тримебутина , desmethyltrimebutine (nor-trimebutine), может быть эффективно продуцирован путем окисления коммерчески доступного лекарственного средства in vitro. Гидроксилирование N-метильной группы приводит к изгнанию молекулы формальдегида , в то время как окисление О-метиловых групп происходит в меньшей степени.
Окисление
[ редактировать ]- Система монооксигеназы цитохрома P450
- Флавинсодержащая монооксигеназная система
- Алкогольдегидрогеназа и альдегиддегидрогеназа
- Моноаминоксидаза
- Коооогание пероксидазами
Снижение
[ редактировать ]Cytochrome P450 reductase, also known as NADPH:ferrihemoprotein oxidoreductase, NADPH:hemoprotein oxidoreductase, NADPH:P450 oxidoreductase, P450 reductase, POR, CPR, CYPOR, is a membrane-bound enzyme required for electron transfer to cytochrome P450 in the microsome of the eukaryotic ячейка из причуда и FMN-содержащий фермент NADPH: цитохром P450 редуктаза Общая схема потока электронов в системе POR/P450: Надф → Увлечение → FMN → P450 → O 2
Во время реакций восстановления химическое вещество может войти в бесполезный велосипед , в котором он получает свободный радикальный электрон, а затем быстро теряет его из-за кислорода (с образованием супероксидного аниона ).
Гидролиз
[ редактировать ]Фаза II - сопряжение
[ редактировать ]В последующих реакциях фазы II эти активированные ксенобиотические метаболиты конъюгированы с заряженными видами, такими как глутатион (GSH), сульфат , глицин или глюкуроновая кислота . Сайты на лекарствах, где возникают реакции конъюгации, включают группы карбокси (-coOH), гидрокси (-OH), амино (NH 2 ) и тиол (-SH). Продукты реакций конъюгации имеют повышенную молекулярную массу и имеют тенденцию быть менее активными, чем их субстраты, в отличие от реакций фазы I, которые часто продуцируют активные метаболиты . Добавление крупных анионных групп (таких как GSH) детоксифицирует реактивные электрофилы и производит больше полярных метаболитов, которые не могут диффундировать по мембранам и, следовательно, могут активно транспортироваться.
Эти реакции катализируются большой группой трансфератов широкой специфичности, которые в комбинации могут метаболизировать практически любое гидрофобное соединение, которое содержит нуклеофильные или электрофильные группы. [ 1 ] Одним из наиболее важных классов этой группы является классы глутатионе S-трансферазы (GST).
| Механизм | Вовлеченное фермент | Коафактор | Расположение | Источники |
|---|---|---|---|---|
| метилирование | метилтрансфераза | S-Аденозил-L-метионин | печень, почка, легкие, CNS | [ 9 ] |
| сульфатация | сульфотрансферазы | 3'-фосфоаденозин-5'-фосфосульфат | печень, почка, кишечник | [ 9 ] |
| ацетилирование | ацетил коэнзим а | печень, легкая, селезенка, слизистая оболочка желудка, эритроциты , лимфоциты | [ 9 ] | |
| глюкуронидация | UDP-глюкуронозилтрансферазы | UDP-глюкуроновая кислота | печень, почка, кишечник, легкие, кожа, простата, мозг | [ 9 ] |
| Глутатионное сопряжение | глутатион S-трансферазы | глутатион | печень, почка | [ 9 ] |
| Спряжение глицина | Два шага процесса:
|
глицин | печень, почка | [ 10 ] |
Фаза III - дальнейшая модификация и экскреция
[ редактировать ]После реакций фазы II ксенобиотические конъюгаты могут быть далее метаболизированы. Распространенным примером является обработка конъюгатов глутатиона в конъюгаты ацетилцистеина (меркаптуриновая кислота). [ 11 ] Здесь остатки γ-глутамата и глицина в молекуле глутатиона удаляются гамма-глутамил-трансептидазой и дипептидазами . На последнем этапе остаток цистеина в конъюгате ацетилирован .
Конъюгаты и их метаболиты могут быть выделены из клеток в фазе III их метаболизма, при этом анионные группы выступают в качестве аффинных меток для различных мембранных транспортеров семейства белка с резистентностью с множественной лекарственной стойкой (MRP). [ 12 ] Эти белки являются членами семейства АТФ-связывающих кассет-транспортеров и могут катализировать АТФ-зависимый транспорт огромного разнообразия гидрофобных анионов, [ 13 ] и, таким образом, действуйте для удаления продуктов фазы II во внеклеточную среду, где они могут быть далее метаболизированы или выводится. [ 14 ]
Эндогенные токсины
[ редактировать ]Детоксикация эндогенных реактивных метаболитов, таких как пероксиды и реактивные альдегиды, часто не может быть достигнута с помощью системы, описанной выше. Это является результатом того, что эти виды будут получены из нормальных клеточных составляющих и обычно разделяют их полярные характеристики. Однако, поскольку эти соединения мало в числе, ферментативные системы могут использовать специфическое молекулярное распознавание для распознавания и удаления их. Сходство этих молекул с полезными метаболитами означает, что различные ферменты детоксикации обычно требуются для метаболизма каждой группы эндогенных токсинов. Примерами этих специфических систем детоксикации являются система глиоксалазы , которая действует для утилизации реактивного альдегида метилглаксаля и различных антиоксидантных систем, которые удаляют активные формы кислорода .
Сайты
[ редактировать ]Количественно гладкая эндоплазматическая ретикулума клеток печени является основным органом лекарственного метаболизма, хотя каждая биологическая ткань обладает некоторой способностью метаболизировать лекарства. Факторы, ответственные за вклад печени в метаболизм лекарственного средства, включают, что это большой орган, что он является первым органом, перфузированным химическими веществами, поглощенными в кишечнике , и что существуют очень высокие концентрации большинства лекарственных метаболизирующих ферментных систем относительно других органов. Если препарат принимается в желудочно-кишечный тракт, где он входит в печеночную циркуляцию через портальную вену , он становится хорошо метаболизированным и, как говорят, показывает первый эффект прохода .
Другие участки метаболизма лекарственного средства включают эпителиальные клетки желудочно -кишечного тракта , легких , почек и кожи . Эти сайты обычно ответственны за локализованные реакции токсичности.
Факторы, влияющие на метаболизм наркотиков
[ редактировать ]Продолжительность и интенсивность фармакологического действия большинства липофильных препаратов определяются скоростью, которую они метаболизируются в неактивные продукты. Система монооксигеназы цитохрома P450 является важным путем в этом отношении. В целом, все, что увеличивает скорость метаболизма (например, индукция фермента ) фармакологически активного метаболита, уменьшит продолжительность и интенсивность действия лекарственного средства. Противоположность также верна, как при ингибировании ферментов . Однако в тех случаях, когда фермент отвечает за метаболизацию пролевки в лекарственное средство, индукция фермента может ускорить эту конверсию и повысить уровень лекарственного средства, потенциально вызывая токсичность. [ Медицинская цитата необходима ]
Различные физиологические и патологические факторы также могут влиять на метаболизм лекарств. Физиологические факторы, которые могут влиять на метаболизм лекарственного средства, включают возраст, индивидуальные вариации (например, фармакогенетика ), энтерогепатическую кровообращение , питание , половые различия или кишечная микробиота . [ Медицинская цитата необходима ] Этот последний фактор имеет значение, поскольку микроорганизмы кишечника способны химически модифицировать структуру лекарств посредством процессов деградации и биотрансформации, тем самым изменяя активность и токсичность лекарств. Эти процессы могут снизить эффективность лекарств, как и случай дигоксина в присутствии Eggerthella Lenta в микробиоте. [ 15 ] Генетическая вариация ( полиморфизм ) учитывает некоторую вариабельность влияния лекарств. [ 15 ]
В целом, лекарства метаболизируются медленнее у плода , неонатальных и пожилых людей и животных , чем у взрослых . Унаследованные генетические различия в ферментах с метаболизированием лекарств приводят к их различным уровням каталитической активности. Например, N-ацетилтрансферазы (участвующие в реакциях фазы II ), индивидуальные вариации создают группу людей, которые медленно ацетилируют ( медленные ацетиляторы ), и тех, кто быстро ацетилит ( быстрые ацетиляторы ), расщепляются примерно на 50:50 в популяции Канады. Тем не менее, изменчивость распределения аллелей Nat2 по разным популяциям высока, а некоторые этнические группы имеют более высокую долю медленных ацетиляторов. [ 16 ] Это изменение в метаболизирующей способности может иметь драматические последствия, поскольку медленные ацетиляторы более подвержены дозе токсичности. Фермент NAT2 является первичным метаболизмом противотуберкулеза ( изониазид ), некоторых антигипертензивных ( гидралазин ), антиарритмических препаратов ( прокаинамид ), антидепрессанты ( фенользин ) и многие другие [ 17 ] и повышенная токсичность, а также побочные реакции лекарств в медленных ацетилаторах широко сообщаются. Подобные явления измененного метаболизма из-за наследственных вариаций были описаны для других ферментов лекарственного средства, таких как CYP2D6 , CYP3A4 , DPYD , UGT1A1 . DPYD и UGT1A1 в настоящее время требуется перед введением соответствующих соединений субстрата ( 5-FU и капецитабин для DPYD и иринотекана для UGT1A1) для определения активности DPYD и UGT1A1 Полем [ 18 ]
Доза, частота, путь введения, распределение тканей и связывание белка препарата влияют на его метаболизм. [ Медицинская цитата необходима ] Патологические факторы также могут влиять на метаболизм лекарств, включая печень , почки или сердечные заболевания. [ Медицинская цитата необходима ]
В моделировании и моделировании силико и моделировании позволяют предсказать метаболизм лекарств в виртуальных популяциях пациентов до проведения клинических исследований у людей. [ 19 ] Это может быть использовано для идентификации людей, наиболее подверженных риску от побочной реакции.
История
[ редактировать ]Исследования о том, как люди трансформируют вещества, которые они принимают, начались в середине девятнадцатого века, когда химики обнаружили, что органические химические вещества, такие как бензальдегид, могут быть окислены и конъюгированы с аминокислотами в организме человека. [ 20 ] В течение оставшейся части девятнадцатого века было обнаружено несколько других основных реакций детоксикации, таких как метилирование , ацетилирование и сульфонация .
В начале двадцатого века работа перешла к исследованию ферментов и путей, которые были ответственны за производство этих метаболитов. Эта область стала определенной как отдельная область обучения с публикацией Ричардом Уильямсом книги из механизмов детоксикации в 1947 году. [ 21 ] Это современное биохимическое исследование привело к идентификации глютатионов S -трансфераз в 1961 году, [ 22 ] Затем следует открытие цитохрома P450S в 1962 году, [ 23 ] и реализация их центральной роли в ксенобиотическом метаболизме в 1963 году. [ 24 ] [ 25 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный в Jakoby WB, Ziegler DM (декабрь 1990 г.). «Ферменты детоксикации» . Дж. Биол. Химический 265 (34): 20715–8. doi : 10.1016/s0021-9258 (17) 45272-0 . PMID 2249981 . Архивировано из оригинала на 2009-06-21 . Получено 2012-12-29 .
- ^ Mizuno N, Niwa T, Yotsumoto Y, Sugiyama Y (сентябрь 2003 г.). «Влияние исследований транспортера лекарств на обнаружение и развитие лекарств». Фармакол. Преподобный 55 (3): 425–61. doi : 10.1124/pr.55.3.1 . PMID 12869659 . S2CID 724685 .
- ^ Thornalley PJ (июль 1990 г.). «Система гликсалазы: новые разработки в отношении функциональной характеристики метаболического пути, фундаментального к биологической жизни» . Биохимия. Дж . 269 (1): 1–11. doi : 10.1042/bj2690001 . PMC 1131522 . PMID 2198020 .
- ^ SIES H (март 1997 г.). «Окислительный стресс: окислители и антиоксиданты» . Эксплуат Физиол . 82 (2): 291–5. doi : 10.1113/expphysiol.1997.sp004024 . PMID 9129943 .
- ^ Guengerich FP (июнь 2001 г.). «Общие и необычные реакции цитохрома P450, связанные с метаболизмом и химической токсичностью». Химический Резерв Токсиколовый . 14 (6): 611–50. doi : 10.1021/tx0002583 . PMID 11409933 .
- ^ Schlichting I, Berendzen J, Chu K, Stock AM, Maves SA, Benson DE, Sweet RM, Ringe D, Petsko GA, Sligar SG (март 2000 г.). «Каталитический путь цитохрома P450CAM при атомной разрешении». Наука . 287 (5458): 1615–22. Bibcode : 2000sci ... 287.1615s . doi : 10.1126/science.287.5458.1615 . PMID 10698731 .
- ^ «Ацетонитрил (EHC 154, 1993)» . www.inchem.org . Архивировано из оригинала 2017-05-22 . Получено 2017-05-03 .
- ^ Akagah B, Lormier AT, Fournet A, Ficadère B (декабрь 2008 г.). «Окисление антипаразитовых 2-замещенных хинолинов с использованием металлопорфириновых катализаторов: масштабирование биомиметической реакции для выработки метаболита кандидатов на лекарственные средства». Орг Биомол. Химический 6 (24): 4494–7. doi : 10.1039/b815963g . PMID 19039354 .
- ^ Jump up to: а беременный в дюймовый и Liston HL, Markowitz JS, Devane CL (октябрь 2001 г.). «Глюкуронидирование лекарств при клинической психофармакологии». J Clin Psychopharmacol . 21 (5): 500–15. doi : 10.1097/00004714-200110000-00008 . PMID 11593076 . S2CID 6068811 .
- ^ Badenhorst CP, Van der Sluis R, Erasmus E, Van Dijk AA (сентябрь 2013 г.). «Конъюгация глицина: важность в метаболизме, роль глициновой н-ацилтрансферазы и факторы, которые влияют на межиндивидуальные вариации». Опытное мнение о метаболизме лекарств и токсикологии . 9 (9): 1139–1153. doi : 10.1517/17425255.2013.796929 . PMID 23650932 . S2CID 23738007 .
Конъюгация глицина митохондриальной ацил-коас, катализируемое глицином N-ацилтрансферазой (глиат, EC 2.3.1.13), является важным метаболическим путем, ответственным за поддержание адекватных уровней свободного коэнзима A (Coash). Однако из -за небольшого числа фармацевтических препаратов, которые конъюгированы с глицином, путь еще не был охарактеризован подробно. Здесь мы рассмотрим причины и возможные последствия межиндивидуальных изменений в пути конъюгации глицина. ...
Рисунок 1. Конъюгация глицина бензойной кислоты. Путь конъюгации глицина состоит из двух шагов. Первый бензоат лигируется в коаши, чтобы сформировать высокоэнергетический бензоил-коа тиоэстер. Эта реакция катализируется кислотой средней цепей HXM-A и HXM-B: COA-лигаз и требует энергии в форме АТФ. ... Бензоил-КоА затем конъюгируют с глицином глиатом с образованием гиппорочной кислоты, выпуская коашу. В дополнение к факторам, перечисленным в коробках, уровни АТФ, коаша и глицина могут влиять на общую скорость пути конъюгации глицина. - ^ Boyland E, Chasseaud LF (1969). «Роль глутатионе и глутатион S-трансферазы в биосинтезе меркуттуровой кислоты». Адвла Фермол. Относительно Области мол. Биол . Достижения в области фермеров - и связанных с ними областей молекулярной биологии. 32 : 173–219. doi : 10.1002/9780470122778.CH5 . ISBN 9780470122778 Полем PMID 4892500 .
- ^ Homolya L, Váradi A, Sarkadi B (2003). «Многородные белки, связанные с резистентностью: экспортные насосы для конъюгатов с глутатионом, глюкуронатом или сульфатом». Биофофакторы . 17 (1–4): 103–14. doi : 10.1002/biof.5520170111 . PMID 12897433 . S2CID 7744924 .
- ^ König J, Nies AT, Cui Y, Leier I, Keppler D (декабрь 1999 г.). «Конъюгатные экспортные насосы семейства белковых белков с множественной лекарственной устойчивостью (MRP): локализация, специфичность субстрата и MRP2-опосредованная лекарственная устойчивость» . Биохим. Биофиз. Акт . 1461 (2): 377–94. doi : 10.1016/s0005-2736 (99) 00169-8 . PMID 10581368 .
- ^ Commandeur JN, Stijntjes GJ, Vermeulen NP (июнь 1995 г.). «Ферменты и транспортные системы, участвующие в формировании и расположении глутатиона S-конъюгатов. Роль в биоактивации и механизмах детоксикации ксенобиотиков». Фармакол. Преподобный 47 (2): 271–330. PMID 7568330 .
- ^ Jump up to: а беременный Heinken A, Hertel J, Acharya G, et al. (19 января 2023 г.). «Метаболическая реконструкция генома в масштабе 7 302 микроорганизма человека для персонализированной медицины» . Nature Biotechnology . 41 (9): 1320–1331. doi : 10.1038/s41587-022-01628-0 . PMC 10497413 . PMID 36658342 .
- ^ Гутьеррес-Вирген, Хорхе Э.; Пинья-Поза, Марисела; Эрнандес-Табия, Эстер А.; Таджа-Чейб, Люсия; Лопес-Гонсалес, Массачусетс. Лурда; Meraz-Ríos, Marco A.; Гомес, Роцио (2023-04-06). "NAT2 Global Landsca: разнообразие и аварация обзора системной магистрации " плюс OU 18 (4): E0283726. Bibcode : 2023ption .... 1883726g doi : 10,1371/журнал . ISSN 1932-6203 . PMC 10079069 . PMID 37023111 .
- ^ Фукуканага, Коя; Като, Кен; Окусака, Такуджи; Сайто, Такео; Икеда, Масаши; Йошида, Терухико; Зембусу, Хитоши; Ивата, Накао; Муширода, Тайзей (2021-03-18). «Функциональная характеристика влияния аллелей N-ацетилтрансферазы 2 на N-ацетилирование восьми лекарств и разнообразие по всему миру » Границы в генетике 12 : 652704. DOI : 10.3389/ fgene.2021.652704 ISSN 1664-8 8012690PMC 33815485PMID
- ^ Малдун, Меган; Бек, Молли; Sebree, Nichlas; Йодер, Робин; Риттер, Стейси; Аллен, Иосия Д.; Алькахтани, Зухайр; Грунд, Хайме; Филипс, Брук; Гессе, Кристина; Эль -Руби, Нихал (февраль 2024 г.). «Реальное внедрение фармакогенетического тестирования DPYD и UGT1A1 в раковом центре на уровне сообщества» . Клиническая и трансляционная наука . 17 (2). doi : 10.1111/cts.13704 . ISSN 1752-8054 . PMC 10818131 .
- ^ Ростами-Ходжеган А., Такер Г.Т. (февраль 2007 г.). «Моделирование и прогнозирование метаболизма лекарств in vivo в популяциях человека из in vitro данных ». Nat Rev Drug Discov . 6 (2): 140–8. doi : 10.1038/nrd2173 . PMID 17268485 . S2CID 205476485 .
- ^ Мерфи П.Дж. (июнь 2001 г.). «Ксенобиотический метаболизм: взгляд от прошлого в будущее» . Метабовка для наркотиков. Расположенный 29 (6): 779–80. PMID 11353742 . Архивировано из оригинала на 2009-06-21 . Получено 2012-12-29 .
- ^ Neuberger A, Smith RL (1983). «Ричард Теквин Уильямс: Человек, его работа, его влияние». Метабовка для наркотиков. Преподобный 14 (3): 559–607. doi : 10.3109/03602538308991399 . PMID 6347595 .
- ^ Бут Дж., Бойланд Э, Симс П (июнь 1961 г.). «Фермент из катализации печени крысы с глутатионом» . Биохимия. Дж . 79 (3): 516–24. doi : 10.1042/bj0790516 . PMC 1205680 . PMID 16748905 .
- ^ Омура Т., Сато Р (апрель 1962 г.). «Новый цитохром в микросомах печени» . Дж. Биол. Химический 237 (4): 1375–6. doi : 10.1016/s0021-9258 (18) 60338-2 . PMID 14482007 . Архивировано из оригинала на 2009-06-21 . Получено 2012-12-29 .
- ^ Estabrook RW (декабрь 2003 г.). «Страсть к P450S (воспоминания о ранней истории исследований по цитохрому P450)». Метабовка для наркотиков. Расположенный 31 (12): 1461–73. doi : 10.1124/dmd.31.12.1461 . PMID 14625342 .
- ^ Estabrook RW, Cooper Dy, Rosenthal O (1963). «Обратимое ингибирование монооксида углерода системы стероидной гидроксилазы в коре надпочечников» в коре надпочечников ». Biochem z . 338 : 741–55. PMID 14087340 .
Дальнейшее чтение
[ редактировать ]- Parvez H, Reiss C (2001). Молекулярные ответы на ксенобиотики . Elsevier. ISBN 0-345-42277-5 .
- Ioannides C (2001). Ферментные системы, которые метаболизируют лекарства и другие ксенобиотики . Джон Уайли и сыновья. ISBN 0-471-89466-4 .
- Ричардсон М. (1996). Экологические ксенобиотики . Taylor & Francis Ltd. ISBN 0-7484-0399-X .
- Ioannides C (1996). Цитохромы P450: метаболические и токсикологические аспекты . CRC Press Inc. ISBN 0-8493-9224-1 .
- Awasthi YC (2006). Токсикология глутатионина S-трансферессы . CRC Press Inc. ISBN 0-8493-2983-3 .
Внешние ссылки
[ редактировать ]- Базы данных
- Метаболизм наркотиков
- Микробная биодеградация
- История
- История ксенобиотического метаболизма на машине Wayback (архивировано 13 июля 2007 г.)