Каспаза 3
| КАСП3 | ||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | CASP3 , CPP32, CPP32B, SCA-1, каспаза 3 | |||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 600636 ; МГИ : 107739 ; Гомологен : 37912 ; Генные карты : CASP3 ; OMA : CASP3 — ортологи | |||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||



Каспаза-3 представляет собой белок каспазы , который взаимодействует с каспазой-8 и каспазой-9 . Он кодируется геном CASP3 . CASP3 Ортологи [ 4 ] были идентифицированы у многих млекопитающих , для которых доступны полные данные о геноме. Уникальные ортологи имеются также у птиц , ящериц , лисамфибий и костистых рыб .
Белок CASP3 - является членом цистеин ) аспарагиновой кислоты семейства протеаз ( каспаз . [ 5 ] Последовательная активация каспаз играет центральную роль в фазе апоптоза клеток . Каспазы существуют в виде неактивных проферментов , которые подвергаются протеолитическому процессингу консервативных остатков аспарагиновой кислоты с образованием двух субъединиц, большой и малой, которые димеризуются с образованием активного фермента . Этот белок расщепляет и активирует каспазы 6 и 7 ; а сам белок процессируется и активируется каспазами 8, 9 и 10 . Это преобладающая каспаза, участвующая в расщеплении белка-предшественника бета-амилоида 4А , который связан с гибелью нейронов при болезни Альцгеймера . [ 6 ] Альтернативный сплайсинг этого гена приводит к образованию двух вариантов транскрипта, кодирующих один и тот же белок. [ 7 ]
 |
 |
Каспаза-3 имеет многие типичные характеристики, общие для всех известных в настоящее время каспаз. Например, его активный сайт содержит остаток цистеина (Cys-163) и остаток гистидина (His-121), которые стабилизируют расщепление пептидной связи белковой последовательности на карбокси-концевой стороне аспарагиновой кислоты , когда она является частью определенного Последовательность из 4 аминокислот. [ 9 ] [ 10 ] Эта специфичность позволяет каспазам быть невероятно избирательными: аспарагиновая кислота в 20 000 раз предпочитает глутаминовую кислоту . [ 11 ] Ключевой особенностью каспаз в клетке является то, что они присутствуют в виде зимогенов , называемых прокаспазами, которые неактивны до тех пор, пока биохимические изменения не вызовут их активацию. Каждая прокаспаза имеет N-концевую большую субъединицу массой около 20 кДа, за которой следует меньшая субъединица массой около 10 кДа, называемая р20 и р10 соответственно. [ 12 ]
Специфичность субстрата
[ редактировать ]В норме каспазы распознают тетрапептидные последовательности на своих субстратах и гидролизуют пептидные связи после остатков аспарагиновой кислоты . Каспаза 3 и каспаза 7 имеют сходную субстратную специфичность, распознавая тетрапептидный мотив Asp-xx-Asp. [ 13 ] С-концевой Asp абсолютно необходим, тогда как вариации в трех других положениях допустимы. [ 14 ] Специфичность субстрата каспазы широко используется при разработке ингибиторов каспаз и лекарств. [ 15 ]
Структура
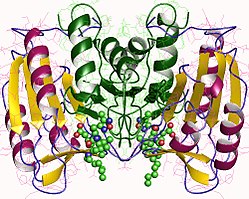
[ редактировать ]В частности, каспаза-3 (также известная как CPP32/Yama/апопаин) [ 16 ] [ 17 ] [ 18 ] образуется из зимогена массой 32 кДа, который расщепляется на субъединицы массой 17 кДа и 12 кДа. Когда прокаспаза расщепляется по определенному остатку, активный гетеротетрамер затем может образовываться за счет гидрофобных взаимодействий, в результате чего четыре антипараллельных бета-листа от p17 и два от p12 собираются вместе, образуя гетеродимер, который, в свою очередь, взаимодействует с другим гетеродимером. с образованием полной 12-нитевой структуры бета-листа, окруженной альфа-спиралями , уникальной для каспаз. [ 12 ] [ 19 ] Когда гетеродимеры выравниваются друг с другом «голова к хвосту», на каждом конце молекулы располагается активный центр, образованный остатками обеих участвующих субъединиц, хотя необходимые остатки Cys-163 и His-121 обнаруживаются на p17 (более крупные ) субъединица. [ 19 ]

Механизм
[ редактировать ]Каталитический сайт каспазы-3 включает тиоловую группу Cys-163 и имидазольное кольцо His-121. His-121 стабилизирует карбонильную группу ключевого остатка аспартата, тогда как Cys-163 атакует, в конечном итоге разрывая пептидную связь. Cys-163 и Gly-238 также стабилизируют тетраэдрическое переходное состояние комплекса субстрат-фермент посредством водородных связей . [ 19 ] In vitro было обнаружено, что каспаза-3 предпочитает пептидную последовательность DEVDG (Asp-Glu-Val-Asp-Gly), расщепление которой происходит на карбоксильной стороне второго остатка аспарагиновой кислоты (между D и G). [ 11 ] [ 19 ] [ 20 ] Каспаза-3 активна в широком диапазоне pH , который немного выше (более основной), чем многие другие каспазы-палачи. Этот широкий диапазон указывает на то, что каспаза-3 будет полностью активна в нормальных и апоптотических клеточных условиях. [ 21 ]

Активация
[ редактировать ]Каспаза-3 активируется в апоптотической клетке как внешним (лиганд смерти), так и внутренним (митохондриальным) путями. [ 12 ] [ 22 ] Зимогенная особенность каспазы-3 необходима, потому что, если ее не регулировать, активность каспазы будет убивать клетки без разбора. [ 23 ] В качестве каспазы-палача зимоген каспазы-3 практически не обладает активностью до тех пор, пока он не будет расщеплен каспазой-инициатором после того, как произошли события апоптотической передачи сигнала. [ 24 ] Одним из таких сигнальных событий является введение гранзима B киллерами , который может активировать инициаторные каспазы, в клетки, нацеленные на апоптоз Т-клетками- . [ 25 ] [ 26 ] Эта внешняя активация затем запускает характерный каспазный каскад, характерный для апоптотического пути, в котором каспаза-3 играет доминирующую роль. [ 10 ] При внутренней активации цитохром c из митохондрий работает в сочетании с каспазой-9 , фактором активации апоптоза 1 ( Apaf-1 ) и АТФ , обрабатывая прокаспазу-3. [ 20 ] [ 26 ] [ 27 ] Этих молекул достаточно для активации каспазы-3 in vitro, но in vivo необходимы и другие регуляторные белки . [ 27 ] Было показано, что экстракт мангустина ( Garcinia mangostana ) ингибирует активацию каспазы 3 в обработанных B-амилоидом нейрональных клетках человека. [ 28 ]
Торможение
[ редактировать ]Одним из способов ингибирования каспаз является семейство белков IAP (ингибитор апоптоза), которое включает c-IAP1, c-IAP2, XIAP и ML-IAP. [ 19 ] XIAP связывает и ингибирует инициатор каспазу-9, которая принимает непосредственное участие в активации каспазы-исполнителя-3. [ 27 ] Однако во время каспазного каскада каспаза-3 ингибирует активность XIAP, расщепляя каспазу-9 в определенном сайте, не позволяя XIAP связываться и ингибировать активность каспазы-9. [ 29 ]
Взаимодействия
[ редактировать ]Было показано, что каспаза 3 взаимодействует с:
Биологическая функция
[ редактировать ]Было обнаружено, что каспаза-3 необходима для нормального развития мозга , а также ее типичная роль в апоптозе, где она отвечает за конденсацию хроматина и фрагментацию ДНК . [ 20 ] Повышенный уровень фрагмента каспазы-3, р17, в кровотоке является признаком недавнего инфаркта миокарда . [ 51 ] В настоящее время показано, что каспаза-3 может играть роль в дифференцировке эмбриональных и гемопоэтических стволовых клеток . [ 52 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000031628 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Филогенетический маркер OrthoMaM: кодирующая последовательность CASP3» . Архивировано из оригинала 3 марта 2016 г. Проверено 20 декабря 2009 г.
- ^ Алнемри Э.С., Ливингстон Д.Д., Николсон Д.В., Салвесен Дж., Торнберри Н.А., Вонг В.В., Юань Дж. (октябрь 1996 г.). «Номенклатура человеческих протеаз ICE/CED-3» . Клетка . 87 (2): 171. doi : 10.1016/S0092-8674(00)81334-3 . ПМИД 8861900 . S2CID 5345060 .
- ^ Жерве Ф.Г., Сюй Д., Робертсон Г.С., Вайланкур Дж.П., Чжу Ю., Хуан Дж., Леблан А., Смит Д., Ригби М., Ширман М.С., Кларк Э.Э., Чжэн Х., Ван дер Плог Л.Х., Руффоло СК, Торнберри Н.А., Ксантудакис С. Замбони Р.Дж., Рой С., Николсон Д.В. (апрель 1999 г.). «Участие каспаз в протеолитическом расщеплении белка-предшественника бета-амилоида Альцгеймера и образовании амилоидогенного бета-пептида» . Клетка . 97 (3): 395–406. дои : 10.1016/s0092-8674(00)80748-5 . ПМИД 10319819 . S2CID 17524567 .
- ^ «Ген Энтреза: каспаза 3 CASP3, цистеинпептидаза, связанная с апоптозом» .
- ^ Харрингтон Х.А., Хо К.Л., Гош С., Тунг К.К. (2008). «Построение и анализ модульной модели активации каспаз при апоптозе» . Теоретическая биология и медицинское моделирование . 5 (1): 26. дои : 10.1186/1742-4682-5-26 . ПМК 2672941 . ПМИД 19077196 .
- ^ Уилли АХ (1997). «Апоптоз: обзор» . Британский медицинский бюллетень . 53 (3): 451–65. doi : 10.1093/oxfordjournals.bmb.a011623 . ПМИД 9374030 .
- ^ Перейти обратно: а б Перри Д.К., Смит М.Дж., Стеннике Х.Р., Салвесен Г.С., Дюрье П., Пуарье Г.Г., Ханнун Ю.А. (июль 1997 г.). «Цинк является мощным ингибитором апоптотической протеазы каспазы-3. Новая мишень для цинка в ингибировании апоптоза» . Журнал биологической химии . 272 (30): 18530–3. дои : 10.1074/jbc.272.30.18530 . ПМИД 9228015 .
- ^ Перейти обратно: а б Стеннике Х.Р., Ренатус М., Мелдал М., Салвесен Г.С. (сентябрь 2000 г.). «Внутренне погашенные флуоресцентные пептидные субстраты раскрывают предпочтения субсайтов человеческих каспаз 1, 3, 6, 7 и 8» . Биохимический журнал . 350 (2): 563–8. дои : 10.1042/0264-6021:3500563 . ПМК 1221285 . ПМИД 10947972 .
- ^ Перейти обратно: а б с Салвесен Г.С. (январь 2002 г.). «Каспасы: открывание ящиков и толкование стрелок». Смерть клеток и дифференцировка . 9 (1): 3–5. дои : 10.1038/sj.cdd.4400963 . ПМИД 11803369 . S2CID 31274387 .
- ^ Агнисвами Дж., Фанг Б., Вебер И.Т. (сентябрь 2007 г.). «Пластичность карманов специфичности S2-S4 каспазы-палача-7, выявленная с помощью структурного и кинетического анализа». Журнал ФЭБС . 274 (18): 4752–65. дои : 10.1111/j.1742-4658.2007.05994.x . ПМИД 17697120 . S2CID 1860924 .
- ^ Фанг Б., Боросс П.И., Тожер Дж., Вебер И.Т. (июль 2006 г.). «Структурный и кинетический анализ каспазы-3 показывает роль сайта связывания s5 в распознавании субстрата». Журнал молекулярной биологии . 360 (3): 654–66. дои : 10.1016/j.jmb.2006.05.041 . ПМИД 16781734 .
- ^ Вебер И.Т., Фанг Б., Агнисвами Дж. (октябрь 2008 г.). «Каспазы: структурно-ориентированная разработка лекарств для контроля гибели клеток». Мини-обзоры по медицинской химии . 8 (11): 1154–62. дои : 10.2174/138955708785909899 . ПМИД 18855730 .
- ^ Фернандес-Алнемри Т., Литвак Г., Алнемри Э.С. (декабрь 1994 г.). «CPP32, новый апоптозный белок человека, гомологичен белку гибели клеток Caenorhabditis elegans Ced-3 и бета-превращающему ферменту интерлейкина-1 млекопитающих» . Журнал биологической химии . 269 (49): 30761–4. дои : 10.1016/S0021-9258(18)47344-9 . ПМИД 7983002 .
- ^ Тевари М., Куан Л.Т., О'Рурк К., Деснуайерс С., Зенг З., Бейдлер Д.Р., Пуарье Г.Г., Салвесен Г.С., Диксит В.М. (июнь 1995 г.). «Yama/CPP32 beta, гомолог CED-3 млекопитающих, представляет собой CrmA-ингибируемую протеазу, которая расщепляет субстрат смерти поли(АДФ-рибоза) полимеразу» . Клетка . 81 (5): 801–9. дои : 10.1016/0092-8674(95)90541-3 . ПМИД 7774019 . S2CID 18866447 .
- ^ Николсон Д.В., Али А., Торнберри Н.А., Вайанкур Дж.П., Дин С.К., Галлант М., Гаро Ю., Гриффин П.Р., Лабелль М., Лазебник Ю.А. (июль 1995 г.). «Идентификация и ингибирование протеазы ICE/CED-3, необходимой для апоптоза млекопитающих». Природа . 376 (6535): 37–43. Бибкод : 1995Natur.376...37N . дои : 10.1038/376037a0 . ПМИД 7596430 . S2CID 4240789 .
- ^ Перейти обратно: а б с д и Лаврик И.Н., Голкс А., Краммер П.Х. (октябрь 2005 г.). «Каспазы: фармакологическое манипулирование гибелью клеток» . Журнал клинических исследований . 115 (10): 2665–72. дои : 10.1172/JCI26252 . ПМЦ 1236692 . ПМИД 16200200 .
- ^ Перейти обратно: а б с Портер АГ, Янике РУ (февраль 1999 г.). «Новая роль каспазы-3 в апоптозе» . Смерть клеток и дифференцировка . 6 (2): 99–104. дои : 10.1038/sj.cdd.4400476 . ПМИД 10200555 .
- ^ Стеннике Х.Р., Салвесен Г.С. (октябрь 1997 г.). «Биохимическая характеристика каспаз-3, -6, -7 и -8» . Журнал биологической химии . 272 (41): 25719–23. дои : 10.1074/jbc.272.41.25719 . ПМИД 9325297 .
- ^ Гавами С., Хашеми М., Анде С.Р., Йеганех Б., Сяо В., Эшраги М., Бус С.Дж., Кадхода К., Вечек Э., Халайко А.Дж. , Лос М. (август 2009 г.). «Апоптоз и рак: мутации в генах каспаз» . Журнал медицинской генетики . 46 (8): 497–510. дои : 10.1136/jmg.2009.066944 . ПМИД 19505876 .
- ^ Боатрайт К.М., Салвесен Г.С. (декабрь 2003 г.). «Механизмы активации каспаз». Современное мнение в области клеточной биологии . 15 (6): 725–31. дои : 10.1016/j.ceb.2003.10.009 . ПМИД 14644197 .
- ^ Уолтерс Дж., Поп С., Скотт Ф.Л., Драг М., Шварц П., Маттос С., Салвесен Г.С., Кларк AC (декабрь 2009 г.). «Конститутивно активный и неингибируемый зимоген каспазы-3 эффективно индуцирует апоптоз» . Биохимический журнал . 424 (3): 335–45. дои : 10.1042/BJ20090825 . ПМК 2805924 . ПМИД 19788411 .
- ^ Галлахер Б.В., Хилле Р., Райл К., Кисс В. (сентябрь 2001 г.). «Апоптоз: жить или умереть — в любом случае тяжелая работа!». Гормональные и метаболические исследования . 33 (9): 511–9. дои : 10.1055/s-2001-17213 . ПМИД 11561209 . S2CID 36623826 .
- ^ Перейти обратно: а б Катунума Н., Мацуи А., Ле КТ, Уцуми К., Салвесен Г., Охаши А. (2001). «Новый каскад активации прокаспазы-3, опосредованный лизоапоптазами, и его биологическое значение в апоптозе». Достижения в регуляции ферментов . 41 (1): 237–50. дои : 10.1016/S0065-2571(00)00018-2 . ПМИД 11384748 .
- ^ Перейти обратно: а б с Ли П., Ниджхаван Д., Ван Икс (январь 2004 г.). «Митохондриальная активация апоптоза» . Клетка . 116 (2 доп): С57–9, 2 п после С59. дои : 10.1016/S0092-8674(04)00031-5 . ПМИД 15055583 . S2CID 5180966 .
- ^ Мунгкарнди П., Срисават С., Саетун П., Джантаравинид Дж., Пирапиттайамонгкол С., Сой-ампорнкул Р., Джунну С., Синчайкул С., Чен С.Т., Чароэнсилп П., Тонгбункерд В., Неунгтон Н. (май 2010 г.). «Защитное действие экстракта мангустина против цитотоксичности, индуцированной бета-амилоидом, окислительного стресса и измененного протеома в клетках SK-N-SH» (PDF) . Журнал исследований протеома . 9 (5): 2076–86. дои : 10.1021/pr100049v . ПМИД 20232907 .
- ^ Дено Ж.Б., Экельман Б.П., Шин Х., Поп С., Салвесен Г.С. (июль 2007 г.). «Каспаза 3 ослабляет XIAP (X-связанный ингибитор белка апоптоза), опосредованное ингибирование каспазы 9» . Биохимический журнал . 405 (1): 11–9. дои : 10.1042/BJ20070288 . ЧВК 1925235 . ПМИД 17437405 .
- ^ Го Й., Сринивасула С.М., Друиле А., Фернандес-Алнемри Т., Алнемри Э.С. (апрель 2002 г.). «Каспаза-2 индуцирует апоптоз, высвобождая проапоптотические белки из митохондрий» . Журнал биологической химии . 277 (16): 13430–7. дои : 10.1074/jbc.M108029200 . ПМИД 11832478 .
- ^ Сринивасула С.М., Ахмад М., Фернандес-Алнемри Т., Литвак Г., Алнемри Э.С. (декабрь 1996 г.). «Молекулярное упорядочение пути Fas-апоптоза: протеаза Fas/APO-1 Mch5 представляет собой CrmA-ингибируемую протеазу, которая активирует несколько Ced-3/ICE-подобных цистеиновых протеаз» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (25): 14486–91. Бибкод : 1996PNAS...9314486S . дои : 10.1073/pnas.93.25.14486 . ПМК 26159 . ПМИД 8962078 .
- ^ Сельвакумар, П.; Шарма, РК. (май 2007 г.). «Роль кальпаина и каспазной системы в регуляции N-миристоилтрансферазы при раке толстой кишки человека (обзор)» . Int J Mol Med . 19 (5): 823–7. дои : 10.3892/ijmm.19.5.823 . ПМИД 17390089 .
- ^ Шу Х.Б., Халпин Д.Р., Гёддел Д.В. (июнь 1997 г.). «Casper является индуктором апоптоза, связанным с FADD и каспазой» . Иммунитет . 6 (6): 751–63. дои : 10.1016/S1074-7613(00)80450-1 . ПМИД 9208847 .
- ^ Хан Д.К., Чаудхари П.М., Райт М.Е., Фридман С., Траск Б.Дж., Ридель Р.Т., Баскин Д.Г., Шварц С.М., Худ Л. (октябрь 1997 г.). «MRIT, новый белок, содержащий домен эффектора смерти, взаимодействует с каспазами и BclXL и инициирует гибель клеток» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (21): 11333–8. Бибкод : 1997PNAS...9411333H . дои : 10.1073/pnas.94.21.11333 . ПМК 23459 . ПМИД 9326610 .
- ^ Форсет С, Йе Х, Грейнджер Л, Корсет В, Шин Х, Бредесен Д.Е., Мелен П. (март 2001 г.). «Рецептор зависимости DCC (удаленный при колоректальном раке) определяет альтернативный механизм активации каспаз» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (6): 3416–21. Бибкод : 2001PNAS...98.3416F . дои : 10.1073/pnas.051378298 . ПМК 30668 . ПМИД 11248093 .
- ^ Самали А., Кай Дж., Животовский Б., Джонс Д.П., Оррениус С. (апрель 1999 г.). «Наличие преапоптотического комплекса прокаспазы-3, Hsp60 и Hsp10 в митохондриальной фракции клеток юрката» . Журнал ЭМБО . 18 (8): 2040–8. дои : 10.1093/emboj/18.8.2040 . ПМЦ 1171288 . ПМИД 10205158 .
- ^ Ксантудакис С., Рой С., Распер Д., Хеннесси Т., Обин Ю., Кэссиди Р., Тава П., Руэл Р., Розен А., Николсон Д.В. (апрель 1999 г.). «Hsp60 ускоряет созревание прокаспазы-3 с помощью вышестоящих протеаз-активаторов во время апоптоза» . Журнал ЭМБО . 18 (8): 2049–56. дои : 10.1093/emboj/18.8.2049 . ПМЦ 1171289 . ПМИД 10205159 .
- ^ Руззене М., Пензо Д., Пинна Л.А. (май 2002 г.). «Ингибитор протеинкиназы CK2 4,5,6,7-тетрабромбензотриазол (TBB) индуцирует апоптоз и каспазозависимую деградацию специфического для клеток гемопоэтической линии белка 1 (HS1) в клетках Jurkat» . Биохимический журнал . 364 (Часть 1): 41–7. дои : 10.1042/bj3640041 . ПМЦ 1222543 . ПМИД 11988074 .
- ^ Чен Ю.Р., Кори Р., Джон Б., Тан Т.Х. (ноябрь 2001 г.). «Каспаза-опосредованное расщепление актин-связывающих и содержащих SH3-домен белков кортактина, HS1 и HIP-55 во время апоптоза». Связь с биохимическими и биофизическими исследованиями . 288 (4): 981–9. дои : 10.1006/bbrc.2001.5862 . ПМИД 11689006 .
- ^ Тамм И., Ван Й., Сосвилл Э., Скудьеро Д.А., Винья Н., Ольтерсдорф Т., Рид Дж.К. (декабрь 1998 г.). «Белок сурвивин семейства IAP ингибирует активность каспаз и апоптоз, индуцированные Fas (CD95), Bax, каспазами и противораковыми препаратами». Исследования рака . 58 (23): 5315–20. ПМИД 9850056 .
- ^ Шин С., Сунг Б.Дж., Чо Ю.С., Ким Х.Дж., Ха NC, Хван Джи, Чунг К.В., Юнг Ю.К., О Б.Х. (январь 2001 г.). «Антиапоптотический белок человека сурвивин является прямым ингибитором каспаз-3 и -7». Биохимия . 40 (4): 1117–23. дои : 10.1021/bi001603q . ПМИД 11170436 .
- ^ Ли З.Х., Ли С.Э., Квак К., Йео В., Ли Т.Х., Бэ С.С., Су П.Г., Ким Х.Х. (март 2001 г.). «Каспаза-опосредованное расщепление TRAF3 в FasL-стимулированных клетках Jurkat-T». Журнал биологии лейкоцитов . 69 (3): 490–6. дои : 10.1189/jlb.69.3.490 . ПМИД 11261798 . S2CID 34256107 .
- ^ Лео Э., Деверо К.Л., Бухгольц К., Уэлш К., Мацудзава С., Стеннике Х.Р., Салвесен Г.С., Рид Дж.К. (март 2001 г.). «TRAF1 является субстратом каспаз, активируемых во время апоптоза, индуцированного рецептором фактора некроза опухоли альфа» . Журнал биологической химии . 276 (11): 8087–93. дои : 10.1074/jbc.M009450200 . ПМИД 11098060 .
- ^ Сузуки Ю., Накабаяши Ю., Такахаши Р. (июль 2001 г.). «Активность убиквитин-протеинлигазы Х-связанного ингибитора белка апоптоза способствует протеосомной деградации каспазы-3 и усиливает ее антиапоптотический эффект при Fas-индуцированной гибели клеток» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (15): 8662–7. Бибкод : 2001PNAS...98.8662S . дои : 10.1073/pnas.161506698 . ПМК 37492 . ПМИД 11447297 .
- ^ Силке Дж., Хокинс С.Дж., Экерт П.Г., Чу Дж., Дэй С.Л., Пакуш М., Верхаген А.М., Во Д.Л. (апрель 2002 г.). «Антиапоптотическая активность XIAP сохраняется при мутации как сайтов взаимодействия с каспазой 3, так и с каспазой 9» . Журнал клеточной биологии . 157 (1): 115–24. дои : 10.1083/jcb.200108085 . ПМК 2173256 . ПМИД 11927604 .
- ^ Ридл С.Дж., Ренатус М., Шварценбахер Р., Чжоу К., Сунь С., Фесик С.В., Лиддингтон Р.К., Салвесен Г.С. (март 2001 г.). «Структурная основа ингибирования каспазы-3 XIAP» . Клетка . 104 (5): 791–800. дои : 10.1016/S0092-8674(01)00274-4 . ПМИД 11257232 . S2CID 17915093 .
- ^ Рой Н., Деверо К.Л., Такахаши Р., Салвесен Г.С., Рид Дж.К. (декабрь 1997 г.). «Белки c-IAP-1 и c-IAP-2 являются прямыми ингибиторами специфических каспаз» . Журнал ЭМБО . 16 (23): 6914–25. дои : 10.1093/emboj/16.23.6914 . ПМК 1170295 . ПМИД 9384571 .
- ^ Деверо К.Л., Такахаши Р., Салвесен Г.С., Рид Дж.К. (июль 1997 г.). «X-связанный IAP является прямым ингибитором протеаз клеточной гибели» . Природа . 388 (6639): 300–4. Бибкод : 1997Natur.388..300D . дои : 10.1038/40901 . ПМИД 9230442 . S2CID 4395885 .
- ^ Сузуки Ю., Накабаяши Ю., Наката К., Рид Дж.К., Такахаши Р. (июль 2001 г.). «Х-связанный ингибитор белка апоптоза (XIAP) ингибирует каспазу-3 и -7 разными способами» . Журнал биологической химии . 276 (29): 27058–63. дои : 10.1074/jbc.M102415200 . ПМИД 11359776 .
- ^ Оцубо Т., Камада С., Миками Т., Мураками Х., Цудзимото Ю. (сентябрь 1999 г.). «Идентификация NRF2, члена семейства транскрипционных факторов NF-E2, в качестве субстрата для каспазы-3(-подобных) протеаз» . Смерть клеток и дифференцировка . 6 (9): 865–72. дои : 10.1038/sj.cdd.4400566 . ПМИД 10510468 .
- ^ Агосто М., Азрин М., Сингх К., Джаффе А.С., Лян Б.Т. (январь 2011 г.). «Фрагмент p17 каспазы-3 в сыворотке повышен у пациентов с инфарктом миокарда с подъемом сегмента ST: новое наблюдение» . Журнал Американского колледжа кардиологов . 57 (2): 220–1. дои : 10.1016/j.jacc.2010.08.628 . ПМИД 21211695 .
- ^ Абдул-Гани М., Мегени Л.А. (июнь 2008 г.). «Реабилитация контрактного убийцы: каспаза-3 управляет дифференцировкой стволовых клеток» . Клеточная стволовая клетка . 2 (6): 515–6. дои : 10.1016/j.stem.2008.05.013 . ПМИД 18522841 .
Дальнейшее чтение
[ редактировать ]- Коэн GM (август 1997 г.). «Каспазы: палачи апоптоза» . Биохимический журнал . 326 (Часть 1): 1–16. дои : 10.1042/bj3260001 . ПМЦ 1218630 . ПМИД 9337844 .
- Ройг Дж., Трау Дж.А. (2001). Цитостатическая протеинкиназа гамма-PAK, активируемая белком p21 G. Витамины и гормоны. Том. 62. стр. 167–98. дои : 10.1016/S0083-6729(01)62004-1 . ISBN 9780127098623 . ПМИД 11345898 .
- Чжао LJ, Чжу Х (декабрь 2004 г.). «Структура и функция вспомогательного регуляторного белка Vpr ВИЧ-1: новые ключи к разработке лекарств». Текущие цели по борьбе с наркотиками. Иммунные, эндокринные и метаболические нарушения . 4 (4): 265–75. дои : 10.2174/1568008043339668 . ПМИД 15578977 .
- Ле Рузик Э, Бенишу С (2006). «Белок Vpr ВИЧ-1: различные роли в жизненном цикле вируса» . Ретровирусология . 2 (1): 11. дои : 10.1186/1742-4690-2-11 . ПМК 554975 . ПМИД 15725353 .
- Сайкс MC, Моубрей А.Л., Джо Х (февраль 2007 г.). «Обратимое глутатиолирование каспазы-3 глутаредоксином как новый окислительно-восстановительный сигнальный механизм при гибели клеток, индуцированной фактором некроза опухоли-альфа». Исследование кровообращения . 100 (2): 152–4. дои : 10.1161/01.RES.0000258171.08020.72 . ПМИД 17272816 . S2CID 12684325 .
Внешние ссылки
[ редактировать ]- Онлайн-база данных MEROPS Архивировано по пептидазам и их ингибиторам: C14.003. 3 марта 2016 г. на Wayback Machine.
- Апоптоз и каспаза 3 – Карта протеолиза – анимация
галерея PDB |
|---|