ГроЭЛ
| HSPD1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | HSPD1 , CPN60, GROEL, HLD4, HSP-60, HSP60, HSP65, HuCHA60, SPG13, член 1 семейства белков теплового шока D (Hsp60) | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 118190 ; МГИ : 96242 ; Гомологен : 1626 ; Генные карты : HSPD1 ; ОМА : HSPD1 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||






В этой статье отсутствует информация о действии на бактерии и хлоропласты. ( декабрь 2020 г. ) |
GroEL — это белок, принадлежащий к шаперонинов семейству молекулярных шаперонов и обнаруженный во многих бактериях. [5] Он необходим для правильного сворачивания многих белков. Для правильного функционирования GroEL необходим белковый комплекс кошаперонина, похожий на крышку, GroES . У эукариот белки органелларные Hsp60 и Hsp10 структурно и функционально практически идентичны GroEL и GroES соответственно из-за их эндосимбиотического происхождения.
HSP60 участвует в импорте митохондриальных белков и сборке макромолекул. Он может способствовать правильному сворачиванию импортированных белков, а также может предотвращать неправильное сворачивание и способствовать повторному сворачиванию и правильной сборке развернутых полипептидов, образующихся в условиях стресса в митохондриальном матриксе. HSP60 взаимодействует с HRAS, а также с белком X HBV и белком p40tax HTLV-1. HSP60 принадлежит к семейству шаперонинов (HSP60). Примечание. Это описание может включать информацию из UniProtKB.
Альтернативные названия: шаперонин 60 кДа, шаперонин 60, CPN60, белок теплового шока 60, HSP-60, HuCHA60, белок митохондриального матрикса P1, белок лимфоцитов P60, HSPD1.
Белок теплового шока 60 (HSP60) представляет собой митохондриальный шаперонин , который обычно отвечает за транспортировку и рефолдинг белков из цитоплазмы в митохондриальный матрикс . Помимо своей роли белка теплового шока, HSP60 действует как шаперонин, помогая сворачивать линейные аминокислотные цепи в соответствующую трехмерную структуру. Благодаря обширному изучению groEL, бактериального гомолога HSP60, HSP60 был признан необходимым для синтеза и транспортировки незаменимых митохондриальных белков из цитоплазмы клетки в митохондриальный матрикс. Дальнейшие исследования связали HSP60 с диабетом , реакцией на стресс , раком и некоторыми типами иммунологических нарушений.
Открытие
[ редактировать ]О функции HSP60 известно немного. HSP60 млекопитающих впервые был описан как митохондриальный белок P1. Впоследствии он был клонирован и секвенирован Рэди Гуптой и его коллегами. [6] Аминокислотная последовательность показала сильную гомологию с GroEL. Первоначально считалось, что HSP60 функционирует только в митохондриях не существует эквивалентного белка и что в цитоплазме . Недавние открытия опровергли это утверждение и предположили, что существует заметная разница между HSP60 в митохондриях и цитоплазме. [7] Подобная белковая структура существует в хлоропластах некоторых растений. Наличие этого белка свидетельствует об эволюционной связи развития митохондрий и хлоропластов путем эндосимбиоза . [6]
Структура
[ редактировать ]В нормальных физиологических условиях HSP60 представляет собой олигомер массой 60 килодальтон, состоящий из мономеров, которые образуют комплекс, расположенный в виде двух сложенных друг на друга гептамерных колец. [8] Эта двойная кольцевая структура образует большую центральную полость, в которой развернутый белок связывается посредством гидрофобных взаимодействий. [9] Эта структура обычно находится в равновесии с каждым из своих отдельных компонентов: мономерами, гептамерами и тетрадекамерами. [10] Недавние исследования начали предполагать, что помимо типичного расположения в митохондриях, HSP60 также можно обнаружить в цитоплазме при нормальных физиологических условиях. [7]
Каждая субъединица HSP60 имеет три домена : апикальный домен, экваториальный домен и промежуточный домен. [11] Экваториальный домен содержит сайт связывания АТФ и другого гептамерного кольца. Промежуточный домен связывает экваториальный домен и апикальный домен вместе. [11] Промежуточный домен вызывает конформационные изменения при связывании АТФ, что позволяет чередовать гидрофильные и гидрофобные сайты связывания субстрата. [11] В неактивном состоянии белок находится в гидрофобном состоянии. При активации АТФ промежуточный домен претерпевает конформационные изменения, в результате которых обнажается гидрофильная область. Это обеспечивает точность связывания белка. [11] Шаперонин 10 помогает HSP60 сворачиваться, действуя как куполообразное покрытие на АТФ-активной форме HSP60. Это приводит к увеличению центральной полости и способствует сворачиванию белка. [11] Более подробную информацию о структуре смотрите на рисунке выше.
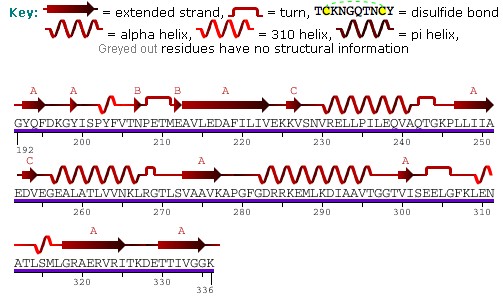
митохондриального HSP60 Последовательность содержит серию G-повторов на С-конце . [6] Структура и функция этой последовательности до конца не известны. N -конец содержит пре-последовательность гидроксилированных аминокислот , а именно аргинина , лизина , серина и треонина , которые служат проводниками импорта белка в митохондрии. [6]
Предсказанная структура HSP60 включает в себя несколько вертикальных синусоидальных волн , альфа-спиралей , бета-листов и поворотов на 90 градусов. Существуют области гидрофобности , где белок предположительно пересекает мембрану . Также имеются три сайта N-связанного гликозилирования в положениях 104, 230, 436. [9] Последовательность и вторичная структура митохондриального белка показаны на изображении выше, полученном из банка данных белков.
Новая информация начала предполагать, что HSP60, обнаруженный в митохондриях, отличается от такового в цитоплазме. Что касается аминокислотной последовательности, цитоплазматический HSP60 имеет N-концевую последовательность, не обнаруженную в митохондриальном белке. [7] При анализе гель-электрофореза были обнаружены существенные различия в миграции цитоплазматического и митохондриального HSP60. Цитоплазматический HSP60 содержит сигнальную последовательность из 26 аминокислот на N-конце. Эта последовательность сильно вырождена и способна сворачиваться в амфифильную спираль . [7] Антитела против HSP60 нацелены как на митохондриальную, так и на цитоплазматическую форму. [7] Тем не менее, антитела против сигнальной последовательности нацелены только на цитоплазматическую форму. В нормальных физиологических условиях оба вещества обнаруживаются в относительно равных концентрациях. [7] Во время стресса или высокой потребности в HSP60 либо в цитоплазме, либо в митохондриях клетка способна компенсировать это за счет увеличения присутствия HSP60 в одном компартменте и снижения его концентрации в противоположном компартменте.
Функция
[ редактировать ]Общий
[ редактировать ]Белки теплового шока являются одними из наиболее эволюционно консервативных белков . [10] Значительная функциональная, структурная и последовательная гомология между HSP60 и его прокариотическим гомологом, groEL, демонстрирует этот уровень консервативности. Более того, аминокислотная последовательность HSP60 имеет сходство с его гомологом у растений , бактерий и человека . [13] Белки теплового шока несут основную ответственность за поддержание целостности клеточных белков, особенно в ответ на изменения окружающей среды. Такие стрессы, как температура, дисбаланс концентрации, изменение pH и токсины, могут стимулировать белки теплового шока поддерживать конформацию белков клетки. HSP60 способствует укладке и поддержанию конформации примерно 15–30% всех клеточных белков. [11] В дополнение к типичной роли HSP60 как белка теплового шока, исследования показали, что HSP60 играет важную роль в транспортировке и поддержании митохондриальных белков, а также в передаче и репликации митохондриальной ДНК .
Транспорт митохондриального белка
[ редактировать ]HSP60 несет две основные обязанности в отношении транспорта митохондриальных белков. Он катализирует сворачивание белков, предназначенных для матрикса, и поддерживает белок в развернутом состоянии для транспорта через внутреннюю мембрану митохондрий. [14] Многие белки предназначены для переработки в матриксе митохондрий, но затем быстро экспортируются в другие части клетки. Гидрофобная часть HSP60 отвечает за поддержание развернутой конформации белка для трансмембранного транспорта. [14] Исследования показали, как HSP60 связывается с поступающими белками и вызывает конформационные и структурные изменения. Последующие изменения концентрации АТФ гидролизуют связи между белком и HSP60, что сигнализирует белку о выходе из митохондрий. [14] HSP60 также способен различать белки, предназначенные для экспорта, и белки, которым суждено оставаться в митохондриальном матриксе, путем поиска амфифильной альфа-спирали из 15-20 остатков. [14] Наличие этой последовательности сигнализирует о том, что белок должен быть экспортирован, тогда как отсутствие сигнализирует о том, что белок должен оставаться в митохондриях. Точный механизм еще не до конца понятен.
Метаболизм ДНК
[ редактировать ]Помимо своей решающей роли в сворачивании белков, HSP60 участвует в репликации и передаче митохондриальной ДНК . В обширных исследованиях активности HSP60 у Saccharomyces cerevisiae ученые предположили, что HSP60 преимущественно связывается с одноцепочечной цепью матричной ДНК в тетрадекамероподобном комплексе. [15] Этот тетрадекамерный комплекс взаимодействует с другими транскрипционными элементами, служа регуляторным механизмом репликации и передачи митохондриальной ДНК. Мутагенные исследования также подтвердили регуляторное участие HSP60 в репликации и передаче митохондриальной ДНК. [16] Мутации в HSP60 повышают уровень митохондриальной ДНК и приводят к последующим дефектам передачи.
Цитоплазматический и митохондриальный HSP60
[ редактировать ]Помимо уже проиллюстрированных структурных различий между цитоплазматическим и митохондриальным HSP60, существуют заметные функциональные различия. Исследования показали, что HSP60 играет ключевую роль в предотвращении апоптоза в цитоплазме. Цитоплазматический HSP60 образует комплекс с белками, ответственными за апоптоз, и регулирует активность этих белков. [7] Цитоплазматическая версия также участвует в иммунном ответе и раке . [7] Эти два аспекта будут подробно рассмотрены позже. Совсем недавние исследования начали предполагать регуляторную корреляцию между HSP60 и гликолитическим ферментом 6- фосфофруктокиназой-1 . Хотя информации доступно немного, цитоплазматические концентрации HSP60 влияют на экспрессию 6-фосфофруктокиназы при гликолизе . [17] Несмотря на эти заметные различия между цитоплазматической и митохондриальной формой, экспериментальный анализ показал, что клетка способна быстро перемещать цитоплазматический HSP60 в митохондрии, если условия окружающей среды требуют более высокого присутствия митохондриального HSP60. [7]
Синтез и сборка
[ редактировать ]HSP60 обычно обнаруживается в митохондриях, а также в органеллах эндосимбиотического происхождения. Мономеры HSP60 образуют два гептамерных кольца, которые связываются с поверхностью линейных белков и катализируют их сворачивание в АТФ-зависимом процессе. [18] Субъединицы HSP60 кодируются ядерными генами и транслируются в цитозоль. Эти субъединицы затем перемещаются в митохондрии, где обрабатываются другими молекулами HSP60. [9] Несколько исследований показали, что белки HSP60 должны присутствовать в митохондриях для синтеза и сборки дополнительных компонентов HSP60. [9] Существует прямая положительная корреляция между наличием белков HSP60 в митохондриях и продукцией дополнительных белковых комплексов HSP60.
Кинетика . сборки субъединиц HSP60 в 2-гептамерные кольца занимает две минуты Последующий устойчивый к протеазе HSP60 образуется за период полувыведения 5–10 минут. [9] Этот быстрый синтез указывает на то, что существует АТФ-зависимое взаимодействие, при котором образовавшийся комплекс HSP60 стабилизирует промежуточное соединение комплекса сборки HSP60, эффективно служа катализатором. [9] Необходимость ранее существовавшего HSP60 для синтеза дополнительных молекул HSP60 подтверждает эндосимбиотическую теорию происхождения митохондрий . Должно быть, существовал рудиментарный прокариотический гомологичный белок, способный к подобной самосборке.
Иммунологическая роль
[ редактировать ]Как обсуждалось выше, HSP60 обычно известен как шаперонин, который способствует сворачиванию белков в митохондриях. Однако некоторые новые исследования показали, что HSP60, возможно, играет роль в иммунном ответе «каскада сигналов опасности» . [19] Появляется все больше доказательств того, что он играет роль в аутоиммунных заболеваниях.
Инфекция и болезнь оказывают на клетку чрезвычайно сильный стресс. Когда клетка находится в состоянии стресса, она естественным образом увеличивает выработку белков стресса, в том числе белков теплового шока , таких как HSP60. Чтобы HSP60 действовал как сигнал, он должен присутствовать во внеклеточной среде. В недавних исследованиях «выяснилось, что… шаперонин 60 можно обнаружить на поверхности различных прокариотических и эукариотических клеток и даже высвобождать из клеток». [11] Согласно недавним исследованиям, множество различных типов белков теплового шока ответа используется в передаче сигналов иммунного , но оказывается, что разные белки действуют и реагируют по-разному на другие сигнальные молекулы. Было показано, что HSP60 высвобождается из специфических клеток, таких как мононуклеарные клетки периферической крови (PBMC), когда присутствуют липополисахариды (LPS) или GroEL. Это говорит о том, что клетка имеет разные рецепторы и реакции на человеческий и бактериальный HSP60. [19] Кроме того, было показано, что HSP60 обладает способностью «активировать моноциты , макрофаги и дендритные клетки… а также индуцировать секрецию широкого спектра цитокинов ». [19] Тот факт, что HSP60 реагирует на другие сигнальные молекулы, такие как LPS или GroEL, и обладает способностью активировать определенные типы клеток, подтверждает идею о том, что HSP60 является частью каскада сигналов опасности, который участвует в активации иммунного ответа.
Однако в иммунологической роли HSP60 есть некоторая особенность. Как упоминалось выше, существует два разных типа белков HSP60: бактериальные и млекопитающие. Поскольку они очень похожи по последовательности, нельзя ожидать, что бактериальный HSP60 вызовет сильный иммунный ответ у людей. Иммунная система «сконструирована так, чтобы игнорировать «я», то есть компоненты хозяина; однако, как это ни парадоксально, с шаперонинами дело обстоит иначе». [11] Было обнаружено, что существует множество антител против шаперонина, которые связаны со многими аутоиммунными заболеваниями. По данным Рэнфорда и др. были проведены эксперименты, которые показали, что антитела , которые «генерируются человеком-хозяином после воздействия бактериальных белков-шаперонинов 60», могут перекрестно реагировать с белками-шаперонинами 60 человека. [11] Бактериальный HSP60 заставляет иммунную систему вырабатывать антитела против шаперонина, хотя бактериальный и человеческий HSP60 имеют схожие белковые последовательности. Эти новые антитела затем распознают и атакуют человеческий HSP60, вызывающий аутоиммунное заболевание. Это предполагает, что HSP60 может играть роль в аутоиммунитете , однако необходимо провести дополнительные исследования, чтобы более полно выяснить его роль в этом заболевании.
Стрессовая реакция
[ редактировать ]Было показано, что HSP60, как митохондриальный белок, также участвует в реакции на стресс. Реакция на тепловой шок — это гомеостатический механизм, который защищает клетку от повреждений путем усиления экспрессии генов, кодирующих HSP60. [20] Активация выработки HSP60 позволяет поддерживать другие клеточные процессы, происходящие в клетке, особенно во время стресса. В одном эксперименте исследователи вводили различных мышей L-ДОФА и обнаружили значительное усиление экспрессии HSP60 в митохондриях и экспрессии HSP70 в цитоплазме. Исследователи пришли к выводу, что сигнальный путь теплового шока служит «основным механизмом защиты от нейротоксичности , вызываемой свободными радикалами кислорода и азота, образующимися при старении и нейродегенеративных заболеваниях». [21] Несколько исследований показали, что HSP60 и другие белки теплового шока необходимы для выживания клеток в токсических или стрессовых условиях. [22]
Связь с раком
[ редактировать ]
Человеческий Hsp60, продукт гена HSPD1, представляет собой митохондриальный шаперонин группы I, филогенетически родственный бактериальному GroEL. Недавно сообщалось о присутствии Hsp60 вне митохондрий и вне клетки, например, в циркулирующей крови [1], [2]. Хотя предполагается, что внемитохондриальная молекула Hsp60 идентична митохондриальной, это еще не полностью выяснено. Несмотря на растущее количество экспериментальных данных, свидетельствующих о том, что Hsp60 находится вне клетки, пока неясно, насколько общим является этот процесс и каковы механизмы, ответственные за транслокацию Hsp60 вне клетки. Ни на один из этих вопросов нет однозначного ответа, хотя имеется некоторая информация о внеклеточном Hsp70. Этот шаперон также классически считался внутриклеточным белком, таким как Hsp60, но в последние несколько лет появились значительные доказательства его периклеточного и внеклеточного присутствия.
Было показано, что HSP60 влияет на апоптоз клеток опухолевых , что, по-видимому, связано с изменением уровня экспрессии. Существует некоторое несоответствие в том, что некоторые исследования показывают положительное выражение, в то время как другие исследования показывают отрицательное выражение, и, похоже, это зависит от типа рака. Существуют разные гипотезы, объясняющие эффекты позитивного и негативного выражения. Положительная экспрессия, по-видимому, ингибирует « апоптотическую и некротическую гибель клеток», тогда как отрицательная экспрессия, как полагают, играет роль «в активации апоптоза». [23] [24]
Было показано, что изменения уровня экспрессии HSP60 не только влияют на апоптоз, но и являются «новыми полезными биомаркерами для диагностических и прогностических целей». [23] По мнению Лебрета и др., потеря экспрессии HSP60 «указывает на плохой прогноз и риск развития опухолевой инфильтрации», особенно при мочевого пузыря карциноме , но это не обязательно справедливо для других типов рака. [25] Например, исследования опухолей яичников показали, что чрезмерная экспрессия коррелирует с лучшим прогнозом, тогда как снижение экспрессии коррелирует с агрессивной опухолью. [25] Все эти исследования показывают, что экспрессию HSP60 можно использовать для прогнозирования выживаемости при определенных типах рака и, следовательно, можно будет выявить пациентов, которым могут помочь определенные методы лечения. [24]
Механизм
[ редактировать ]В этой статье отсутствует информация о различных механизмах взаимодействия бактериального GroEL и HSPD1 — HSPD1 имеет несколько странных симметричных промежуточных продуктов. ( декабрь 2020 г. ) |
Внутри клетки процесс сворачивания белка, опосредованного GroEL/ES, включает несколько раундов связывания, инкапсуляции и высвобождения белка-субстрата. Развернутые белки-субстраты связываются с гидрофобным связывающим участком на внутреннем крае открытой полости GroEL, образуя бинарный комплекс с шаперонином. Связывание белка-субстрата таким образом, в дополнение к связыванию АТФ , индуцирует конформационные изменения , которые позволяют ассоциировать бинарный комплекс с отдельной структурой крышки, GroES . Связывание GroES с открытой полостью шаперонина заставляет отдельные субъединицы шаперонина вращаться так, что гидрофобный сайт связывания субстрата удаляется из внутренней части полости, в результате чего белок-субстрат выбрасывается из обода в теперь уже в значительной степени гидрофильный участок. камера. Гидрофильная среда камеры способствует захоронению гидрофобных остатков субстрата, вызывая сворачивание субстрата. Гидролиз АТФ и связывание нового белка-субстрата с противоположной полостью посылают аллостерический сигнал, вызывающий высвобождение GroES и инкапсулированного белка в цитозоль . Данный белок претерпевает несколько циклов сворачивания, каждый раз возвращаясь в исходное развернутое состояние, пока не будет достигнута нативная конформация или промежуточная структура, обеспечивающая достижение нативного состояния. Альтернативно, субстрат может подвергаться конкурирующей реакции, такой как неправильное сворачивание и агрегация с другими неправильно свернутыми белками. [26]
Термодинамика
[ редактировать ]Ограниченная природа внутренней части молекулярного комплекса сильно благоприятствует компактным молекулярным конформациям белка-субстрата. Свободные в растворе дальнодействующие неполярные взаимодействия могут происходить только с высокой ценой энтропии . В тесных помещениях комплекса GroEL относительная потеря энтропии гораздо меньше. Метод захвата также имеет тенденцию концентрировать неполярные сайты связывания отдельно от полярных сайтов. Когда неполярные поверхности GroEL удалены, вероятность того, что какая-либо данная неполярная группа встретит неполярный внутримолекулярный участок, намного выше, чем в объемном растворе. Гидрофобные сайты, находившиеся снаружи, собираются вместе вверху цис- домена и связываются друг с другом. Геометрия GroEL требует, чтобы полярные структуры были ведущими и охватывали неполярное ядро, когда оно выходит с транс- стороны.
Структура
[ редактировать ]Структурно GroEL представляет собой двухкольцевой тетрадекамер, в котором цис- и транс - кольца состоят из семи субъединиц каждое. Конформационные изменения, которые происходят внутри центральной полости GroEL, приводят к тому, что внутренняя часть GroEL становится гидрофильной, а не гидрофобной, и, вероятно, это способствует сворачиванию белка.
 ГроЭЛ (сбоку)
ГроЭЛ (сбоку) ГроЭЛ (вверху)
ГроЭЛ (вверху) Комплекс ГроЭС/ГроЭЛ (сбоку)
Комплекс ГроЭС/ГроЭЛ (сбоку) Комплекс ГроЭС/ГроЭЛ (вверху)
Комплекс ГроЭС/ГроЭЛ (вверху)




Ключ к активности GroEL кроется в структуре мономера. Мономер Hsp60 имеет три отдельных участка, разделенных двумя шарнирными областями. Апикальный сайтов отдел содержит множество гидрофобных связывания развернутых белковых субстратов . Многие глобулярные белки не связываются с апикальным доменом, поскольку их гидрофобные части сгруппированы внутри, вдали от водной среды, поскольку это термодинамически оптимальная конформация. Таким образом, эти «сайты-субстраты» будут связываться только с белками, которые не свернуты оптимально. Апикальный домен также имеет сайты связывания мономеров Hsp10 GroES.
Экваториальный домен имеет щель вблизи шарнирной точки для связывания АТФ , а также две точки прикрепления другой половины молекулы GroEL. Остальная часть экваториального отдела умеренно гидрофильна.
Добавление АТФ и GroES оказывает радикальное влияние на конформацию цис- домена. Этот эффект вызван сгибанием и вращением двух шарнирных точек мономеров Hsp60. Промежуточный домен сгибается вниз и внутрь примерно на 25° на нижнем шарнире. Этот эффект, умноженный за счет совместного изгибания всех мономеров, увеличивает экваториальный диаметр клетки GroEL. Но апикальный домен поворачивается на полные 60° вверх и наружу на верхнем шарнире, а также поворачивается на 90° вокруг оси шарнира. Это движение очень широко открывает клетку в верхней части цис- домена, но полностью удаляет сайты связывания субстрата изнутри клетки.
Взаимодействия
[ редактировать ]Было показано, что GroEL взаимодействует с GroES . [27] [28] АЛДГ2 , [28] Каспаза 3 [27] [29] и дигидрофолатредуктаза . [30]
Морфогенез фага Т4
[ редактировать ]Гены ( фага бактериофага ) Т4 , кодирующие белки , определяющие структуру фага Т4, были идентифицированы с использованием условно-летальных мутантов . [31] Большинство из этих белков оказались либо основными, либо второстепенными структурными компонентами завершенной фаговой частицы. Однако среди генных продуктов (gps), необходимых для сборки фага, Snustad [32] идентифицировали группу gps, которые действуют каталитически, а не включаются в структуру фага. Эти каталитические GPS включали gp31. Бактерия E. coli является хозяином фага Т4, и кодируемый фагом белок gp31, по-видимому, функционально гомологичен шапаронному белку GroES E. coli и способен заменять его при сборке вирионов фага Т4 во время инфекции. [5] Роль белка gp31, кодируемого фагом, по-видимому, заключается во взаимодействии с белком GroEL, кодируемым хозяином E. coli, для содействия правильному сворачиванию и сборке основного капсидного белка головки фага, gp23. [5]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000144381 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000025980 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Перейти обратно: а б с Зейлстра-Райаллс Дж., Файет О., Георгопулос С. (1991). «Универсально консервативные шаперонины GroE (Hsp60)». Анну. Преподобный Микробиол . 45 : 301–25. дои : 10.1146/annurev.mi.45.100191.001505 . ПМИД 1683763 .
- ^ Перейти обратно: а б с д Гупта Р.С. (январь 1995 г.). «Эволюция семейств шаперонинов (Hsp60, Hsp10 и Tcp-1) белков и происхождение эукариотических клеток» . Мол. Микробиол . 15 (1): 1–11. дои : 10.1111/j.1365-2958.1995.tb02216.x . ПМИД 7752884 .
- ^ Перейти обратно: а б с д и ж г час я Ито Х., Комацуда А., Отани Х. и др. (декабрь 2002 г.). «HSP60 млекопитающих быстро сортируется в митохондрии в условиях обезвоживания». Евро. Дж. Биохим . 269 (23): 5931–8. дои : 10.1046/j.1432-1033.2002.03317.x . ПМИД 12444982 .
- ^ Ченг М.Ю., Хартл Ф.У., Хорвич А.Л. (ноябрь 1990 г.). «Митохондриальный шаперонин hsp60 необходим для собственной сборки». Природа . 348 (6300): 455–8. Бибкод : 1990Natur.348..455C . дои : 10.1038/348455a0 . ПМИД 1978929 . S2CID 28394330 .
- ^ Перейти обратно: а б с д и ж Фентон В.А. и др. (октябрь 1994 г.). «Остатки шаперонина GroEL необходимы для связывания и высвобождения полипептидов». Природа . 371 (6498): 614–9. Бибкод : 1994Natur.371..614F . дои : 10.1038/371614a0 . ПМИД 7935796 . S2CID 23840816 .
- ^ Перейти обратно: а б Хабич С. и др. (март 2007 г.). «Белок теплового шока 60: регуляторная роль в клетках врожденного иммунитета» . Клетка. Мол. Наука о жизни . 64 (6): 742–51. дои : 10.1007/s00018-007-6413-7 . ПМЦ 11138415 . ПМИД 17221165 . S2CID 24067484 .
- ^ Перейти обратно: а б с д и ж г час я Рэнфорд Дж.С. и др. (сентябрь 2000 г.). «Шаперонины — это белки, передающие клеточные сигналы: биология молекулярных шаперонов» . Эксперт преподобный Мол Мед . 2 (8): 1–17. дои : 10.1017/S1462399400002015 . ПМИД 14585136 . S2CID 38110138 .
- ^ PDB : 1SRV ; Уолш М.А. и др. (июнь 1999 г.). «Доведя MAD до крайности: сверхбыстрое определение структуры белка». Акта Кристаллогр. Д. 55 (6): 1168–73. Бибкод : 1999AcCrD..55.1168W . дои : 10.1107/S0907444999003698 . ПМИД 10329779 .
- ^ Джонсон Р.Б. и др. (2003). «Клонирование и характеристика гена дрожжевого шаперонина HSP60». Генетика . 84 (2): 295–300. дои : 10.1016/0378-1119(89)90503-9 . ПМИД 2575559 .
- ^ Перейти обратно: а б с д Колл Х. и др. (март 1992 г.). «Антифолдинговая активность белка hsp60, связывающего импорт в митохондриальный матрикс с экспортом в межмембранное пространство» (PDF) . Клетка . 68 (6): 1163–75. дои : 10.1016/0092-8674(92)90086-R . ПМИД 1347713 . S2CID 7430067 .
- ^ Кауфман, бакалавр. Исследования нуклеоидов митохондриальной ДНК Saccharomyces cerevisiae: идентификация бифункциональных белков. Доктор генетики и развития , Юго-западный медицинский центр UT в Далласе, Даллас, Техас. 241 стр.
- ^ Кауфман, бакалавр (2003). «Функция митохондриального шаперонина Hsp60 в структуре и передаче нуклеоидов митохондриальной ДНК у Saccharomyces cerevisiae» . Журнал клеточной биологии . 163 (3): 457–461. дои : 10.1083/jcb.200306132 . ISSN 0021-9525 . ПМК 2173642 . ПМИД 14597775 .
- ^ Колл Х. и др. (1992). «Антифолдинговая активность импорта белков пар HSP60 в митохондриальный матрикс с экспортом в межмембранное пространство» (PDF) . Клетка . 68 (6): 1163–75. дои : 10.1016/0092-8674(92)90086-R . ПМИД 1347713 . S2CID 7430067 .
- ^ Ито Х. и др. (декабрь 2002 г.). «HSP60 млекопитающих быстро сортируется в митохондрии в условиях обезвоживания». Евро. Дж. Биохим . 269 (23): 5931–8. дои : 10.1046/j.1432-1033.2002.03317.x . ПМИД 12444982 .
- ^ Перейти обратно: а б с Хансен Дж. Дж., Бросс П., Вестергаард М. и др. (январь 2003 г.). «Геномная структура генов митохондриальных шаперонинов человека: HSP60 и HSP10 локализованы лицом к лицу на хромосоме 2, разделенной двунаправленным промотором». Хм. Жене . 112 (1): 71–7. дои : 10.1007/s00439-002-0837-9 . ПМИД 12483302 . S2CID 25856774 .
- ^ Варгас-Парада Л., Солис С. (2001). «Тепловой шок и реакция на стресс Taenia solium и T. crassiceps ». Паразитология . 122 (5): 583–8. дои : 10.1017/s0031182001007764 . ПМИД 11393832 . S2CID 41092962 .
- ^ Калабрезе В., Манкузо С., Раванья А. и др. (май 2007 г.). «In vivo индукция белков теплового шока в черной субстанции после введения L-ДОФА связана с повышенной активностью митохондриального комплекса I и нитрозативным стрессом у крыс: регуляция с помощью окислительно-восстановительного состояния глутатиона» . Дж. Нейрохем . 101 (3): 709–17. дои : 10.1111/j.1471-4159.2006.04367.x . ПМИД 17241115 . S2CID 6512400 .
- ^ Росси М.Р., Сомджи С., Гарретт Ш., Сенс М.А., Нат Дж., Сенс Д.А. (декабрь 2002 г.). «Экспрессия генов ответа на стресс hsp 27, hsp 60, hsc 70 и hsp 70 в культивируемых уротелиальных клетках человека (UROtsa), подвергшихся воздействию летальных и сублетальных концентраций арсенита натрия» . Окружающая среда. Перспектива здоровья . 110 (12): 1225–32. дои : 10.1289/ehp.021101225 . ПМК 1241110 . ПМИД 12460802 .
- ^ Перейти обратно: а б Каппелло Ф., Ди Стефано А., Дэвид С. и др. (ноябрь 2006 г.). «Снижение регуляции Hsp60 и Hsp10 предсказывает канцерогенез бронхиального эпителия у курильщиков с хронической обструктивной болезнью легких» . Рак . 107 (10): 2417–24. дои : 10.1002/cncr.22265 . ПМИД 17048249 .
- ^ Перейти обратно: а б Урусибара М., Кагеяма Ю., Акаши Т. и др. (январь 2007 г.). «HSP60 может предсказать хороший патологический ответ на неоадъювантную химиолучевую терапию при раке мочевого пузыря» . Япония. Дж. Клин. Онкол . 37 (1): 56–61. дои : 10.1093/jjco/hyl121 . ПМИД 17095522 .
- ^ Перейти обратно: а б Лебре Т., Уотсон Р.В., Молинье В. и др. (сентябрь 2003 г.). «Белки теплового шока HSP27, HSP60, HSP70 и HSP90: экспрессия при карциноме мочевого пузыря». Рак . 98 (5): 970–7. дои : 10.1002/cncr.11594 . ПМИД 12942564 . S2CID 30820581 .
- ^ Хорвич А.Л., Фентон В.А., Чепмен Э., Фарр Г.В. (2007). «Два семейства шаперонинов: физиология и механизм». Анну. Преподобный Cell Dev. Биол . 23 : 115–45. doi : 10.1146/annurev.cellbio.23.090506.123555 . ПМИД 17489689 .
- ^ Перейти обратно: а б Самали А., Кай Дж., Животовский Б., Джонс Д.П., Оррениус С. (апрель 1999 г.). «Наличие преапоптотического комплекса прокаспазы-3, Hsp60 и Hsp10 в митохондриальной фракции клеток юрката» . ЭМБО Дж . 18 (8): 2040–8. дои : 10.1093/emboj/18.8.2040 . ПМЦ 1171288 . ПМИД 10205158 .
- ^ Перейти обратно: а б Ли К.Х., Ким Х.С., Чон Х.С., Ли Ю.С. (октябрь 2002 г.). «Шаперонин GroESL опосредует сворачивание белка митохондриальной альдегиддегидрогеназы печени человека в Escherichia coli». Биохим. Биофиз. Рез. Коммун . 298 (2): 216–24. дои : 10.1016/S0006-291X(02)02423-3 . ПМИД 12387818 .
- ^ Ксантудакис С., Рой С., Распер Д., Хеннесси Т., Обин Ю., Кэссиди Р., Тава П., Руэл Р., Розен А., Николсон Д.В. (апрель 1999 г.). «Hsp60 ускоряет созревание прокаспазы-3 с помощью вышестоящих протеаз-активаторов во время апоптоза» . ЭМБО Дж . 18 (8): 2049–56. дои : 10.1093/emboj/18.8.2049 . ПМЦ 1171289 . ПМИД 10205159 .
- ^ Мэйхью М., да Силва AC, Мартин Дж., Эрджюмент-Бромаж Х., Темпст П., Хартл Ф.У. (февраль 1996 г.). «Складывание белка в центральной полости комплекса шаперонинов GroEL-GroES». Природа . 379 (6564): 420–6. Бибкод : 1996Natur.379..420M . дои : 10.1038/379420a0 . ПМИД 8559246 . S2CID 4310511 .
- ^ Эдгар Р.С., Эпштейн Р.Х. (февраль 1965 г.). «Генетика бактериального вируса». Научный американец . 212 (2): 70–8. Бибкод : 1965SciAm.212b..70E . doi : 10.1038/scientificamerican0265-70 . ПМИД 14272117 .
- ^ Снустад Д.П. (август 1968 г.). «Взаимодействия доминирования в клетках Escherichia coli, смешанно инфицированных бактериофагом T4D дикого типа и янтарными мутантами, и их возможные последствия в отношении типа функции гена-продукта: каталитическая или стехиометрическая». Вирусология . 35 (4): 550–63. дои : 10.1016/0042-6822(68)90285-7 . ПМИД 4878023 .
Дальнейшее чтение
[ редактировать ]- Табибзаде С., Брум Дж. (1999). «Белки теплового шока в эндометрии человека на протяжении менструального цикла» . Infect Dis Obstet Gynecol . 7 (1–2): 5–9. doi : 10.1002/(SICI)1098-0997(1999)7:1/2<5::AID-IDOG2>3.0.CO;2-Y . ПМК 1784709 . ПМИД 10231001 .
- Шефер С., Уильямс Дж. А. (2000). «Стресс-киназы и белки теплового шока в поджелудочной железе: возможная роль в нормальном функционировании и заболеваниях». Дж. Гастроэнтерол . 35 (1): 1–9. дои : 10.1080/003655200750024443 . hdl : 2027.42/42441 . ПМИД 10632533 . S2CID 9706591 .
- Мозли П. (2000). «Стрессовые белки и иммунный ответ». Иммунофармакология . 48 (3): 299–302. дои : 10.1016/S0162-3109(00)00227-7 . ПМИД 10960671 .
- Лю Ю, Штайнакер Дж. М. (2001). «Изменения в белках теплового шока скелетных мышц: патологическое значение» . Передний. Биосци . 6 : Д12-25. дои : 10.2741/Лю . ПМИД 11145923 .
- Ван Маеле Б., Дебизер З. (2005). «Интеграция ВИЧ-1: взаимодействие между интегразой ВИЧ-1, клеточными и вирусными белками». СПИД преп . 7 (1): 26–43. ПМИД 15875659 .
- Хохштрассер Д.Ф., Фрутигер С., Паке Н., Байрох А., Равье Ф., Паскуали С., Санчес Х.К., Тиссо Дж.Д., Бьелквист Б., Варгас Р. (1992). «Карта белков печени человека: справочная база данных, созданная путем микросеквенирования и сравнения гелей». Электрофорез . 13 (12): 992–1001. дои : 10.1002/elps.11501301201 . ПМИД 1286669 . S2CID 23518983 .
- Икава С., Вайнберг Р.А. (1992). «Взаимодействие между p21ras и белком теплового шока hsp60, шаперонином» . Учеб. Натл. акад. наук. США . 89 (6): 2012–6. Бибкод : 1992ПНАС...89.2012И . дои : 10.1073/pnas.89.6.2012 . ПМК 48586 . ПМИД 1347942 .
- Брудзинский К., Мартинес В., Гупта Р.С. (1992). «Иммуноцитохимическая локализация белка, родственного белку теплового шока 60, в секреторных гранулах бета-клеток и его измененное распределение у мышей с диабетом, не страдающих ожирением» . Диабетология . 35 (4): 316–24. дои : 10.1007/BF00401198 . ПМИД 1516759 .
- Доусон С.Дж., Уайт, Лос-Анджелес (1992). «Лечение эндокардита, вызванного Haemophilus aphrophilus, ципрофлоксацином». Дж. Заразить . 24 (3): 317–20. дои : 10.1016/S0163-4453(05)80037-4 . ПМИД 1602151 .
- Сингх Б., Патель Х.В., Ридли Р.Г., Фриман К.Б., Гупта Р.С. (1990). «Митохондриальный импорт белка шаперонина человека (HSP60)». Биохим. Биофиз. Рез. Коммун . 169 (2): 391–6. дои : 10.1016/0006-291X(90)90344-M . ПМИД 1972619 .
- Веннер Т.Дж., Сингх Б., Гупта Р.С. (1990). «Нуклеотидные последовательности и новые структурные особенности семейств генов hsp60 (шаперонина) человека и китайского хомячка». ДНК Клеточная Биол . 9 (8): 545–52. дои : 10.1089/dna.1990.9.545 . ПМИД 1980192 .
- Уорд Л.Д., Хонг Дж., Уайтхед Р.Х., Симпсон Р.Дж. (1990). «Разработка базы данных аминокислотных последовательностей белков карциномы толстой кишки человека, разделенных двумерным электрофорезом в полиакриламидном геле». Электрофорез . 11 (10): 883–91. дои : 10.1002/elps.1150111019 . ПМИД 2079031 . S2CID 21541503 .
- Джиндал С., Дудани А.К., Сингх Б., Харли С.Б., Гупта Р.С. (1989). «Первичная структура митохондриального белка человека, гомологичного бактериальным и растительным шаперонинам и микобактериальному антигену массой 65 килодальтон» . Мол. Клетка. Биол . 9 (5): 2279–83. дои : 10.1128/mcb.9.5.2279 . ПМК 363030 . ПМИД 2568584 .
- Уолдингер Д., Эккерскорн С., Лотспейх Ф., Клив Х. (1988). «Гомология аминокислотной последовательности полиморфного клеточного белка из лимфоцитов человека и шаперонинов Escherichia coli (groEL) и хлоропластов (Рубиско-связывающий белок)». Биол. хим. Хоппе-Сейлер . 369 (10): 1185–9. дои : 10.1515/bchm3.1988.369.2.1185 . ПМИД 2907406 .
- Крайзель В., Хильдебрандт Х., Шильц Э., Келер Г., Спамер С., Дитц С., Месснер В., Хайльманн С. (1994). «Иммуно-электронно-микроскопическое обнаружение белка теплового шока 60 (hsp60) в митохондриях гепатоцитов и миокардиоцитов крысы». Акта Гистохим . 96 (1): 51–62. дои : 10.1016/s0065-1281(11)80009-7 . ПМИД 7518175 .
- Корбетт Дж.М., Уиллер Ч.С., Бейкер К.С., Якуб М.Х., Данн М.Дж. (1994). «База данных двумерных гелевых белков миокарда человека: обновление 1994 г.». Электрофорез . 15 (11): 1459–65. дои : 10.1002/elps.11501501209 . ПМИД 7895732 . S2CID 33359306 .
- Бака-Эстрада М.Э., Гупта Р.С., Стед Р.Х., Кроитору К. (1994). «Кишечная экспрессия и клеточные иммунные реакции на белок теплового шока человека 60 при болезни Крона». Копать. Дис. Наука . 39 (3): 498–506. дои : 10.1007/BF02088334 . ПМИД 7907543 . S2CID 22032288 .
- Велес-Гранелл К.С., Ариас А.Е., Торрес-Руис Х.А., Бендаян М. (1994). «Молекулярные шапероны в ткани поджелудочной железы: наличие cpn10, cpn60 и hsp70 в отдельных отсеках секреторного пути ацинарных клеток». Дж. Клеточная наука . 107 (3): 539–49. дои : 10.1242/jcs.107.3.539 . ПМИД 7911805 .
- Мэйхью М., да Силва А.С., Мартин Дж., Эрджюмент-Бромаж Х., Темпст П., Хартл Ф.У. (1996). «Складывание белка в центральной полости комплекса шаперонинов GroEL-GroES». Природа . 379 (6564): 420–6. Бибкод : 1996Natur.379..420M . дои : 10.1038/379420a0 . ПМИД 8559246 . S2CID 4310511 .
- Табибзаде С., Конг Ч.Ф., Сатьясваруп П.Г., Бабакния А. (1996). «Белки теплового шока в эндометрии человека на протяжении менструального цикла» . Хм. Репродукция . 11 (3): 633–40. дои : 10.1093/humrep/11.3.633 . ПМИД 8671282 .
Внешние ссылки
[ редактировать ]- GroEL+Protein в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- «Палеосбактерии: Кусочки: GroEL» . Архивировано из оригинала 26 апреля 2007 г. (Права не защищены)
- 3D макромолекулярные структуры GroEL в EMDB