Рибонуклеотидредуктаза
| рибонуклеозид-дифосфатредуктаза | |||
|---|---|---|---|
 Рибонуклеозид-дифосфатредуктаза гетерооктамер, E.Coli | |||
| Идентификаторы | |||
| ЕС №. | 1.17.4.1 | ||
| CAS №. | 9047-64-7Sy | ||
| Базы данных | |||
| Intenz | Intenz View | ||
| Бренда | Бренда вход | ||
| Расширение | Вид Nicezyme | ||
| Кегг | Кегг вход | ||
| Метатический | Метаболический путь | ||
| Напрямую | профиль | ||
| PDB Структуры | RCSB PDB PDBE PDBSUM | ||
| Джин Онтология | Друг / Quickgo | ||
| |||
Рибонуклеотидредуктаза ( RNR ), также известная как рибонуклеозидфосфатредуктаза , представляет собой фермент , который катализирует образование дезоксирибонуклеотидов из рибонуклеотидов . [ 1 ] [ 2 ] Он катализирует это образование путем удаления 2'-гидроксильной группы рибозных кольца нуклеозидфосфатов (или трифосфатов в зависимости от класса RNR). Это восстановление производит дезоксирибонуклеотиды. [ 3 ] Дезоксирибонуклеотиды, в свою очередь, используются в синтезе ДНК . Реакция, катализируемая RNR, строго консервативна во всех живых организмах. [ 4 ] Кроме того, RNR играет критическую роль в регуляции общей скорости синтеза ДНК, так что ДНК к клеточной массе поддерживается при постоянном соотношении во время деления клеток и репарации ДНК . [ 5 ] Несколько необычная особенность фермента RNR заключается в том, что он катализирует реакцию, которая проходит через свободного радикала . механизм действия [ 6 ] [ 7 ] Субстраты для RNR являются ADP , ВВП , CDP и UDP . DTDP (дезокситимидинофосфат) синтезируется другим ферментом ( тимидилаткиназа ) из DTMP (Deoxytymidine Monophoshate).
Структура
[ редактировать ]Рибонуклеотид редуктазы делятся на три класса. Ферменты RNR класса построены из крупной альфа -субъединицы и небольших бета -субъединиц, которые связываются с образованием активного гетеродимерного тетрамера . Снижение NDP до 2'-DNDP, фермент катализирует синтез де-новорота дезоксирибонуклеотидов (DNTP), которые являются предшественниками синтеза ДНК и необходимы для пролиферации клеток . [ 8 ] RNR класса II продуцируют 5'-дезоксиаденозиловый радикал при гомолитическом расщеплении связи C-CO в аденозилкобаламине. Кроме того, RNR класса III содержат стабильный глицильный радикал. [ 9 ]
Люди несут класс I RNRS. Альфа -субъединица кодируется геном RRM1, в то время как существует две изоформы бета -субъединицы, кодируемых генами RRM2 и RRM2B:
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Каждый альфа -мономер класса I состоит из трех доменов : [ 10 ]
- Один в основном спиральный домен, включающий 220 N-концевых остатков,
- Вторая большая десятицепочечная α/β-структура, содержащая 480 остатков,
- и третья небольшая пятицепочечная α/β-структура, содержащая 70 остатков.
В PFAM второй домен был интерпретирован как два отдельных домена:
- более короткий все-альфа-N-концевой домен,
- и более длинный ствол С-концевой домен.
|
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


Класс-бета-субъединица обычно содержит диметаллический центр и стабильный тирозильный радикал . У людей бета-субъединица опирается на кофактор ди-железа. В E. coli тирозильный радикал расположен в положении 122 (Y122), обеспечивающем стабильный радикал для субъединиц RNR2 класса I. [ 14 ] В A. aegypti этот тирозильный радикал находится в позиции 184 (Y184). [ 15 ] Тирозильный радикал глубоко похоронен внутри белка в гидрофобной среде, расположенной вблизи железного центра, который используется при стабилизации тирозильного радикала. В структуре двух μ-oxo-связанных утюгов преобладают лиганды, которые служат в качестве сайтов связывания железа: четыре карбоксилаты [ аспартат (D146), глутамат (E177, E240 и E274) и два гистидина (H180 и H277). [ 15 ] Ассоциация возникает между С-конце RNR2 и C-концевым RNR1. [ 10 ] Ферментативная активность зависит от ассоциации субъединиц RNR1 и RNR2. Активный сайт состоит из активных групп дитиолов из RNR1, а также дифференциального центра и тирозильного радикала из субъединицы RNR2.
Другие остатки RNR2, такие как аспартат (D273), триптофан (W48) и тирозин (Y356), еще больше стабилизируют тирозильный радикал активного сайте, что позволяет переносить электрон. [ 10 ] Эти остатки помогают в переносе радикального электрона из тирозина (Y122) RNR2 в цистеин (C439) RNR1. Перенос электрона начинается с тирозина RNR2 (Y122) и продолжается в RNR2, чтобы триптофан (W48), который отделен от тирозина RNR1 (Y731) на 2,5 нанометров . Перенос электрона от RNR2 в RNR1 происходит через тирозин (Y356 до Y731) и продолжается через тирозин (Y730) к цистеину (C439) в активном сайте. [ 16 ] Установленные сайты мутации первичной структуры RNR указывают на то, что все упомянутые выше остатки участвуют в передаче свободного радикала на большие расстояния в активное участок. [ 10 ]
У A. aegypti Mosquites RNR1 сохраняет большинство важных аминокислотных остатков, включая аспартат (D64) и валин (V292 или V284), которые необходимы для аллостерической регуляции ; активном остатки ; сайте гидрофобном в Остатки цистеина (C225, C436 и C451), которые участвуют в удалении атома водорода и переносе радикального электрона в активном участке; остатки цистеин (C225 и C436), аспарагин (N434) и глутамат (E441), которые связывают рибонуклеотидный субстрат; остатки тирозина (Y723 и Y743), которые определяют радикальный перенос; и остатки цистеина (C838 и C841), которые используются для регенерации групп дитиола в активном сайте. [ 15 ]
Дрожжи YIL066C Saccharomyces cerevisiae обладают незначительной изоформой большой субъединицы рибонуклеотид-дифосфатредуктазы при обозначении RNR3 или . на дрожжевой хромосоме IX [ 17 ] Это паралог дрожжей RNR1 , который, вероятно, возник из -за события дублирования целого генома . [ 18 ] Комплекс RNR катализирует этап ограничения скорости в синтезе DNTP , регулируемый путями контрольной точки повреждения ДНК и повреждения ДНК посредством локализации небольших субъединиц.
Функция
[ редактировать ]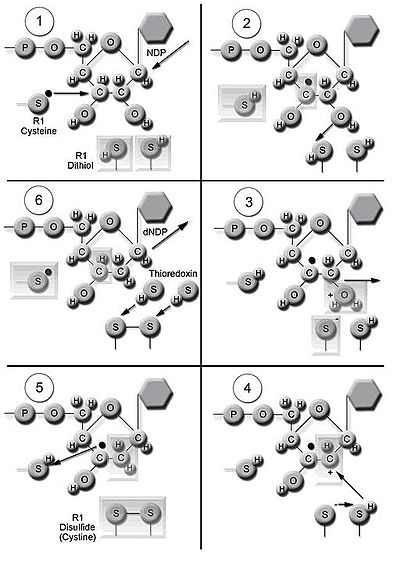
Фермент рибонуклеотидредуктаза (RNR) катализирует синтез De novo DNDP. [ 19 ] Катализ рибонуклеозида 5'-дифосфатов (NDP) включает в себя восстановление при 2'-углероде рибозы 5-фосфата , образуя 2'-дезокси-кровообращение с 2'-дезоксирибонуклеозидом 5'-дифосфатами (DNDP). Это сокращение инициируется генерацией свободного радикала. После одного восстановления RNR требует, чтобы электроны пожертвовали из групп дитиола белка тиоредоксина . Регенерация тиоредоксина происходит, когда никотинамид аденин -динуклеотидфосфат ( NADPH ) обеспечивает два атома водорода, которые используются для уменьшения дисульфидных групп тиоредоксина.
Три класса RNR имеют сходные механизмы для восстановления НДП, но различаются по домену, который генерирует свободный радикал, специфический металл в структуре металлопротеинов и донорами электронов. Все классы используют свободную химию. [ 10 ] Класс I Редактазы используют железный центр с преобразованием железа в железу для генерации тирозильного свободного радикала. Снижение субстратов НДП происходит в аэробных условиях. Класс I редуктазы делятся на IA и IB из -за различий в регулировании. Класс -редуктазы распределяются у эукариот , эубактерий , бактериофагов и вирусов . Рредуктазы класса IB встречаются в Eubacteria. Рредуктазы класса IB также могут использовать радикал, генерируемый при стабилизации бинуклеарного марганцевого центра. свободного радикала Класс II редуктазы генерируют 5'-дезоксиденозиловый радикал из кобаламина (коэнзим B12) и имеют более простую структуру, чем редуктазы класса I и класса III. Снижение НДП или рибонуклеотида 5'-трифосфатов (NTP) происходит в аэробных или анаэробных условиях. Рредуктазы класса II распространяются в Archaebacteria , Eubacteria и бактериофагах. Класс III редуктазы используют глициновый радикал, генерируемый с помощью S-аденозилметионина и центра серы железа. Снижение NTPS ограничено анаэробными условиями. Рредуктазы класса III распределены в Archaebacteria, Eubacteria и бактериофагах. [ 10 ] [ 15 ] Организмы не ограничиваются наличием одного класса ферментов. Например, E. coli имеют как RNR класса I, так и класса III.
Механизм каталитического восстановления
[ редактировать ]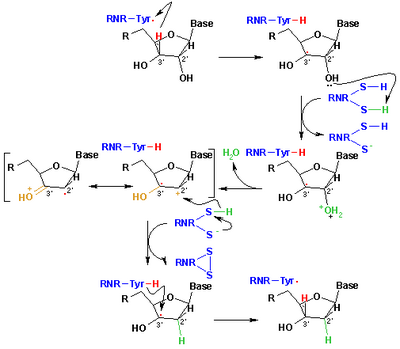
Механизм, который в настоящее время принимается для восстановления рибонуклеотидов до дезоксирибонуклеотидов, изображен в следующей схеме. Первый шаг включает в себя абстракцию 3'-h субстрата 1 радикальным Cys439. Впоследствии реакция включает в себя элиминацию одной молекулы воды из углерода C-2 'рибонуклеотида, катализируемой Cys225 и Glu441. На третьем этапе происходит перенос атома водорода от Cys225 к углероду C-2 '2'-кетольного радикала 3 после предыдущего переноса протона от Cys462 до Cys225. В конце этого этапа получаются радикальный анионный дисульфидский мост и промежуточный кетон с закрытым оболочкой. Это промежуточное соединение было идентифицировано во время преобразования нескольких двухзамещенных аналогов субстрата, а также с естественным субстратом [ 20 ] Взаимодействие с ферментными мутантами. Следующим шагом является окисление анионного дисульфидного моста с сопутствующим восстановлением субстрата, генерируя 5. Плотность вращения смещается от атомов серы к атому C-3 'подложки с одновременным переносом протона от Glu441 к углероду C -3 '. Последний шаг-это обратная задача первого шага и включает перенос водорода от Cys439 на C-3 ', восстанавливая начальный радикал и приводя к конечному продукту 6.
Теоретические модели некоторых этапов этих механизмов с использованием полной модели белка R1 могут быть обнаружены в исследованиях, проведенных Cerqueira et al. Полем [ 21 ] [ 22 ]
Регулирование
[ редактировать ]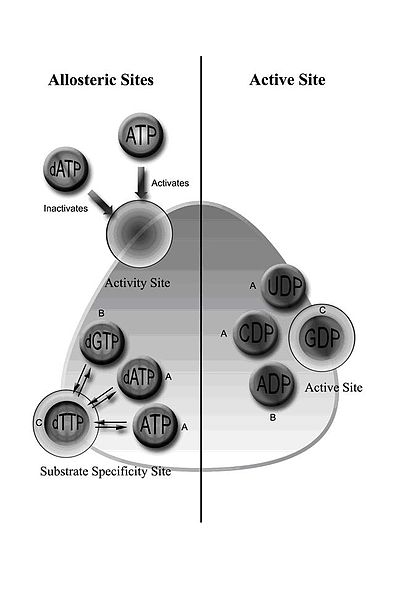
RNR класса I включает в себя субъединицы RNR1 и RNR2, которые могут связать с образованием гетеродимерного тетрамера. [ 6 ] RNR1 содержит оба аллостерические сайты, опосредуя регуляцию субстратной специфичности и активности. [ 12 ] В зависимости от аллостерической конфигурации, один из четырех рибонуклеотидов связывается с активным сайтом.
Регулирование RNR предназначено для поддержания сбалансированного количества DNTP. Связывание эффекторных молекул либо увеличивает, либо уменьшает активность RNR. Когда АТФ связывается с сайтом аллостерической активности, он активирует RNR. Напротив, когда DATP связывается с этим сайтом, он деактивирует RNR. [ 10 ] В дополнение к контролирующей активности, аллостерический механизм также регулирует специфичность субстрата и гарантирует, что фермент производит равное количество каждого DNTP для синтеза ДНК. [ 10 ] Во всех классах связывание АТФ или DATP с аллостерическим сайтом индуцирует восстановление цитидина 5'-дифосфата (CDP) и уридина 5'-дифосфата (UDP); 2'-дезоксигуанозин 5'-трифосфат (DGTP) индуцирует восстановление аденозина 5'-дифосфата (ADP); и 2'-дезокситимидин 5'-трифосфат (DTTP) индуцирует восстановление гуанозина 5'-дифосфата (ВВП) (рис. 1).
Рредуктазы класса IB не ингибируются DATP, потому что им не хватает приблизительно 50 N-концевых аминокислот, необходимых для сайта аллостерической активности. [ 23 ] Кроме того, важно, чтобы активность рибонуклеотидредуктазы находилась под контролем транскрипции и посттранскрипции, поскольку синтез ДНК без повреждений зависит от сбалансированного пула дезоксирибонуклеотидов. [ 24 ] Эукариотические клетки с редуктазами класса IA имеют механизм негативного контроля, чтобы отключить синтез DNTP, когда они накапливаются. Этот механизм защищает клетку от токсичных и мутагенных эффектов, которые могут возникнуть в результате перепроизводства DNTP, поскольку изменения в сбалансированных пулах DNTP приводят к повреждению ДНК и гибели клеток. [ 25 ] [ 26 ] Несмотря на то, что перепроизводство DNTP или несбалансированное предложение из них может привести к неправильному обучению нуклеотидов в ДНК, поставка снабжения DNTPS может позволить восстановить ДНК. P53R2 - это небольшая субъединица рибонуклеотидредуктазы, которая может вызвать такое восстановление. Изменения в рамках этого индуцированного p53 гомолога R2 могут вызвать истощение в митохондриальной ДНК, а следовательно, p53R2 служит основным фактором при постановке DNTP. [ 27 ]
RNR может использовать Morpheein модель аллостерической регуляции . [ 28 ]
Ингибиторы RNR1 и RNR2
[ редактировать ]Обычно ингибиторы RNR класса могут быть разделены на три основные группы: ингибиторы трансляции, которые блокируют синтез фермента; ингибиторы димеризации, которые предотвращают связь двух субъединиц RNR (R1 и R2); и каталитические ингибиторы, которые инактивируют субъединицу R1 и/или субъединицу R2. [ 21 ]
RNR класса может быть ингибирован пептидами, сходными с С-конце RNR2. Эти пептиды могут конкурировать с RNR2 за связывание с RNR1, и в результате RNR1 не образует ферментативно активного комплекса с RNR2. [ 29 ] [ 30 ] Хотя С-конце белков RNR2 различен по видам, RNR2 может взаимодействовать с RNR1 по разным видам. [ 31 ] Когда C-концерт мыши rnr2 был заменен на аминокислотные остатки E.coli RNR2 (7 или 33), химерная субъединица RNR2 все еще связывается с мышиными субъединицами RNR1. Тем не менее, им не хватает ферментативной активности, вероятно, из -за устранения остатков, участвующих в переносе электрона свободного радикала от RNR2 в субъединицу RNR1. [ 30 ]
Маленькие пептиды могут специфически ингибировать субъединицы RNR2 от связывания с RNR1, когда они имеют значительное сходство с нормальным C-конце RNR2. [ 32 ] Это ингибирование связывания RNR2 с RNR1 было успешно проверено в RNR вируса простого герпеса (HSV). Когда 7-аминокислотный олигомер (Gavvndl) усечен с C-конце субъединицы RNR2, использовался в конкурентных анализах, это препятствовало образованию нормального активального комплекса с RNR1. [ 33 ] Другие мелкие пептидные ингибиторы, сходные с C-конце RNR2, также успешно использовались для ингибирования ферментативной активности HSV RNR и, следовательно, репликации HSV. [ 34 ] Сообщается, что у мышей моделей стромального кератита и неоваскуляризации роговицы ( HSV- глазного заболевания ) небольшой аналоговый аналоговый бильд 1263 RNR2 ингибирует RNR и эффективен в предотвращении этих заболеваний. [ 35 ] В некоторых случаях, хотя лечение небольшими С-концевыми аналогами может не остановить распространение заболеваний, они все равно могут помочь в заживлении. Сообщалось, что в ацикловире -резистентном HSV (PAAR5) небольшой ингибитор пептидов Bild 1633 был в 5-10 раз больше мощного, чем BILD 1263 против кожной инфекции PAAR5. [ 36 ] Подход комбинированной терапии (Bild 1633 и ацикловир) более эффективен для лечения местных поражений у мышей. Эти данные свидетельствуют о том, что небольшие ингибиторы пептидов, которые конкурируют с RNR2 за связывание с RNR1, полезны для предотвращения распространения HSV.
Галлий ингибирует RNR2 путем замены Fe 3+ на активном сайте. Малтолат в галлия является перорально биодоступной формой галлия, которая использует эту ингибирующую активность для лечения рака, инфекций и других заболеваний. [ 37 ]
Наркотики гидроксимочевина [ 38 ] и мотексафин гадолиний мешает действию этого фермента. [ 39 ]
Ссылки
[ редактировать ]- ^ Hofer A, Crona M, Logan DT, Sjöberg BM (2012). «Строительные блоки ДНК: хранение контроля над производством» . Критические обзоры биохимии и молекулярной биологии . 47 (1): 50–63. doi : 10.3109/10409238.2011.630372 . PMC 3267527 . PMID 22050358 .
- ^ Элледж С.Дж., Чжоу З, Аллен Дж.Б. (март 1992 г.). «Рибонуклеотидредуктаза: регуляция, регуляция, регуляция». Тенденции в биохимических науках . 17 (3): 119–23. doi : 10.1016/0968-0004 (92) 90249-9 . PMID 1412696 .
- ^ Sneeden JL, Loeb LA (сентябрь 2004 г.). «Мутации в субъединице R2 рибонуклеотидредуктазы, которые обеспечивают устойчивость к гидроксимочевине» . Журнал биологической химии . 279 (39): 40723–8. doi : 10.1074/jbc.m402699200 . PMID 15262976 .
- ^ Торренты Е., Алой П., Гиберт I, Родригес-Треллы F (август 2002 г.). «Рибонуклеотид редуктазы: дивергентная эволюция древнего фермента» . Журнал молекулярной эволюции . 55 (2): 138–52. Bibcode : 2002jmole..55..138t . doi : 10.1007/s00239-002-2311-7 . PMID 12107591 . S2CID 24603578 .
- ^ Herrick J, Sclavi B (январь 2007 г.). «Рибонуклеотидредуктаза и регуляция репликации ДНК: старая история и древнее наследие» . Молекулярная микробиология . 63 (1): 22–34. doi : 10.1111/j.1365-2958.2006.05493.x . PMID 17229208 . S2CID 9473163 .
- ^ Jump up to: а беременный Эклунд Х., Эрикссон М., Ухлин У, Нордлунд П., Логан Д. (август 1997 г.). «Рибонуклеотидредуктаза - структурные исследования радикального фермента». Биологическая химия . 378 (8): 821–5. doi : 10.1515/bchm.1997.378.8.815 . PMID 9377477 .
- ^ Stubbe J, Riggs-Gelasco P (ноябрь 1998 г.). «Использование свободных радикалов: образование и функция тирозильного радикала в рибонуклеотидредуктазе». Тенденции в биохимических науках . 23 (11): 438–43. doi : 10.1016/s0968-0004 (98) 01296-1 . PMID 9852763 .
- ^ Fairman JW, Wijerathna SR, Ahmad MF, Xu H, Nakano R, Jha S, Prendergast J, Welin RM, Flodin S, Roos A, Nordlund P, Li Z, Walz T, Deadwis CG (март 2011 г.). «Структурная основа для аллостерической регуляции рибонуклеотидредуктазы человека с помощью нуклеотид-индуцированной олигомеризации» . Природа структурная и молекулярная биология . 18 (3): 316–22. doi : 10.1038/nsmb.2007 . PMC 3101628 . PMID 21336276 .
- ^ Ларссон К.М., Джордан А., Элиассон Р., Рейхард П., Логан Д.Т., Нордлунд П (ноябрь 2004 г.). «Структурный механизм регуляции специфичности аллостерической субстратной в рибонуклеотидредуктазе». Природа структурная и молекулярная биология . 11 (11): 1142–9. doi : 10.1038/nsmb838 . PMID 15475969 . S2CID 1025702 .
- ^ Jump up to: а беременный в дюймовый и фон глин час Джордан А., Рейхард П. (1998). «Рибонуклеотид редуктазы» . Ежегодный обзор биохимии . 67 (1): 71–98. doi : 10.1146/annurev.biochem.67.1.71 . PMID 9759483 .
- ^ Jump up to: а беременный ВВП : 1PEU ; Uppsten M, Färnegårdh M, Jordan A, Eliasson R, Eklund H, Uhlin U (июнь 2003 г.). «Структура большой субъединицы рибонуклеотид -редуктазы класса IB из сальмонеллы тайфимурия и его комплексов с аллостерическими эффекторами». Журнал молекулярной биологии . 330 (1): 87–97. doi : 10.1016/s0022-2836 (03) 00538-2 . PMID 12818204 .
- ^ Jump up to: а беременный Uhlin U, Eklund H (август 1994 г.). «Структура рибонуклеотид -редуктазы белка R1». Природа . 370 (6490): 533–9. Bibcode : 1994natur.370..533u . doi : 10.1038/370533a0 . PMID 8052308 . S2CID 8940689 .
- ^ Нордлунд П., Эклунд Х (июль 1993 г.). «Структура и функция белка рибонуклеотида escherichia coli рибонуклеотидредуктазы R2». Журнал молекулярной биологии . 232 (1): 123–64. doi : 10.1006/jmbi.1993.1374 . PMID 8331655 .
- ^ Högbom M, Andersson ME, Nordlund P (март 2001 г.). «Кристаллические структуры окисленных динуклеарных марганцевых центров в Mn-замещенной рибонуклеотидредуктазе класса I из Escherichia coli: карбоксилатные сдвиги с последствиями для активации O2 и генерации радикалов». Журнал биологической неорганической химии . 6 (3): 315–23. doi : 10.1007/s00775000000205 . PMID 11315567 . S2CID 20748553 .
- ^ Jump up to: а беременный в дюймовый Pham DQ, Blachuta BJ, Nichol H, Winzerling JJ (сентябрь 2002 г.). «Субъединицы рибонуклеотид редуктазы от комара желтой лихорадки, Aedes aegypti: клонирование и выражение». Биохимия насекомых и молекулярная биология . 32 (9): 1037–44. doi : 10.1016/s0965-1748 (02) 00041-3 . PMID 12213240 .
- ^ Chang MC, Yee CS, Stubbe J, Nocera DG (май 2004 г.). «Включение рибонуклеотидредуктазы инициированной легкой аминокислотной генерацией» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (18): 6882–7. Bibcode : 2004pnas..101.6882c . doi : 10.1073/pnas.0401718101 . PMC 406436 . PMID 15123822 .
- ^ «RNR3 Рибонуклеотид-дифосфатредуктаза субъединица RNR3 [Saccharomyces cerevisiae S288C]» . Получено 2024-08-01 .
- ^ "Rnr3 | sgd" .
- ^ Кокс М., Нельсон Д.Р. (2008). Лехнингер Принципы биохимии . Сан -Франциско: WH Freeman. ISBN 978-0-7167-7108-1 .
- ^ Cerqueira NM, Fernandes PA, Eriksson LA, Ramos MJ (декабрь 2004 г.). «Активация рибонуклеотидов ферментом рибонуклеотидредуктазы: понимание роли фермента». Журнал вычислительной химии . 25 (16): 2031–7. doi : 10.1002/jcc.20127 . PMID 15481089 . S2CID 19665974 .
- ^ Jump up to: а беременный Cerqueira NM, Pereira S, Fernandes PA, Ramos MJ (2005). «Обзор ингибиторов рибонуклеотидредуктазы: привлекательная мишень при противоопухолевой терапии». Текущая лекарственная химия . 12 (11): 1283–94. doi : 10.2174/0929867054020981 . PMID 15974997 .
- ^ Cerqueira NM, Fernandes PA, Eriksson LA, Ramos MJ (март 2006 г.). «Обезвоживание рибонуклеотидов, катализируемых рибонуклеотидредуктазой: роль фермента» . Биофизический журнал . 90 (6): 2109–19. Bibcode : 2006bpj .... 90.2109C . doi : 10.1529/biophysj.104.054627 . PMC 1386789 . PMID 16361339 .
- ^ Eliasson R, Pontis E, Jordan A, Reichard P (октябрь 1996 г.). «Аллостерская регуляция третьей рибонуклеотидредуктазы (NRDEF Enzyme) из Enterobacteriaceae» . Журнал биологической химии . 271 (43): 26582–7. doi : 10.1074/jbc.271.43.26582 . PMID 8900130 .
- ^ Телендер Л (июнь 2007 г.). «Рибонуклеотидредуктаза и синтез митохондриальной ДНК». Природа генетика . 39 (6): 703–4. doi : 10.1038/ng0607-703 . PMID 17534360 . S2CID 22565931 .
- ^ Кунц Б.А. (1988). «Мутагенез и дезоксирибонуклеотидный дисбаланс бассейна». Мутационные исследования . 200 (1–2): 133–47. doi : 10.1016/0027-5107 (88) 90076-0 . PMID 3292903 .
- ^ Meuth M (апрель 1989 г.). «Молекулярная основа мутаций, индуцированных дезоксирибонуклеозидным дисбалансом пула в клетках млекопитающих». Экспериментальные исследования клеток . 181 (2): 305–16. doi : 10.1016/0014-4827 (89) 90090-6 . PMID 2647496 .
- ^ Бурдон А., Минай Л., Серре В., Джайс Дж.П., Сарзи Е., Ауберт С., Кретиен Д., Де Лонл П., Пакис-Флюклингер В., Аракава Х., Накамура Ю., Муннич А., Рётиг А (июнь 2007 г.). «Мутация RRM2B, кодирующая p53-контролируемая рибонуклеотидредуктаза (p53R2), вызывает тяжелое истощение митохондриальной ДНК». Природа генетика . 39 (6): 776–80. doi : 10.1038/ng2040 . PMID 17486094 . S2CID 22103978 .
- ^ Selwood T, Jaffe EK (март 2012 г.). «Динамические диссоциирующие гомоолигомеры и контроль функции белка» . Архивы биохимии и биофизики . 519 (2): 131–43. doi : 10.1016/j.abb.2011.11.020 . PMC 3298769 . PMID 22182754 .
- ^ Clul I, Sjöberg BM, Huang Cy (май 1991). «Карбоксил-концевые пептиды в качестве зондов для эсшерихийской коли рибонуклеотидредуктазы взаимодействие субъединицы: кинетический анализ исследований ингибирования». Биохимия . 30 (21): 5164–71. doi : 10.1021/bi00235a008 . PMID 2036382 .
- ^ Jump up to: а беременный Hamann CS, Lentainge S, Li LS, Salem JS, Yang FD, Cooperman BS (март 1998 г.). «Химерные маленькие ингибиторы субъединиц рибонуклеотид-редуктазы млекопитающих: двойная функция для C-конца R2?» Полем Протеиновая инженерия . 11 (3): 219–24. doi : 10.1093/белок/11.3.219 . PMID 9613846 .
- ^ Cosentino G, Lavallée P, Rakhit S, Plante R, Gaudette Y, Lawetz C, Whitehead PW, Duceppe JS, Lépine-Frenette C, Dansereau N (январь 1991 г.). «Специфическое ингибирование рибонуклеотидредуктаз пептидами, соответствующим С-конце их второй субъединицы». Биохимия и клеточная биология . 69 (1): 79–83. doi : 10.1139/O91-011 . PMID 2043345 .
- ^ Cooperman BS (2003). «Олигопептидное ингибирование рибонуклеотид -редуктаз класса I». Биополимеры . 71 (2): 117–31. doi : 10.1002/bip.10397 . PMID 12767114 . S2CID 25196379 .
- ^ Филатов Д., Ингемрсон Р., Граллонд А., Телендер Л. (август 1992). «Роль вируса простого герпеса рибонуклеотидредуктазы небольшой субъединицы карбоксильной термина при взаимодействии субъединицы и образовании структуры железного тирозильного центра» . Журнал биологической химии . 267 (22): 15816–22. doi : 10.1016/s0021-9258 (19) 49608-7 . PMID 1322407 .
- ^ Cohen EA, Gaudreau P, Brazeau P, Langelier Y (1986). «Специфическое ингибирование герпесвирусной рибонуклеотидредуктазы непептида, полученного из карбокси -конца субъединицы 2». Природа . 321 (6068): 441–3. Bibcode : 1986natur.321..441c . doi : 10.1038/3214441A0 . PMID 3012360 . S2CID 4238076 .
- ^ Брандт К.Р., Спенсер Б., Имш П., Гарно М., Дезиэль Р (май 1996). «Оценка ингибитора пептидомиметической рибонуклеотидредуктазы с мышиной моделью герпестного вируса вируса вируса 1 типа 1» . Антимикробные агенты и химиотерапия . 40 (5): 1078–84. doi : 10.1128/aac.40.5.1078 . PMC 163269 . PMID 8723444 .
- ^ Duan J, Liuzzi M, Paris W, Lambert M, Lawetz C, Moss N, Jaramillo J, Gauthier J, Déziel R, Cordingley MG (июль 1998 г.). «Противовирусная активность селективного ингибитора рибонуклеотидредуктазы против ацикловир-резистентного герпеса вируса простого вируса типа 1 in vivo» . Антимикробные агенты и химиотерапия . 42 (7): 1629–35. doi : 10.1128/aac.42.7.1629 . PMC 105657 . PMID 9660995 .
- ^ Бернштейн Л.Р. (декабрь 1998 г.). «Механизмы терапевтической активности для галлия» (PDF) . Фармакологические обзоры . 50 (4): 665–82. PMID 9860806 .
- ^ «Информация о EC 1.17.4.1-рибонуклеозид-дифосфатредуктаза» . Бренда . Получено 25 июля 2015 года .
- ^ Hashemy Si, Ungerstedt JS, Zahedi Avval F, Holmgren A (апрель 2006 г.). «Мотексафин гадолиний, опухолевая селективная препарат, нацеленная на тиоредоксинкредуктазу и рибонуклеотидредуктазу» . Журнал биологической химии . 281 (16): 10691–7. doi : 10.1074/jbc.m511373200 . PMID 16481328 .
Внешние ссылки
[ редактировать ]- Рибонуклеотид+редуктазы в Национальной библиотеке медицины Медицинской библиотеки Медицинской библиотеки (сетка)
- База данных рибонуклеотид редуктазы (RNRDB) Архивирована 2018-10-15 на машине Wayback