Вирус краснухи
| Рубивирус краснухи | |
|---|---|
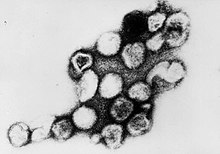 | |
| Трансмиссионная электронная микрофотография вирионов вируса краснухи | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область : | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Китриновирикота |
| Сорт: | Алсувирицетес |
| Заказ: | Гепеливирусы |
| Семья: | Матонавирусиды |
| Род: | Рубивирус |
| Разновидность: | Рубивирус краснухи |
| Синонимы [1] | |
| |
Вирус краснухи ( РуВ ) — возбудитель заболевания краснуха , передается между людьми только респираторным путем и является основной причиной синдрома врожденной краснухи при заражении в первые недели беременности .
Вирус краснухи, научное название Rubellae , является членом рода Rubivirus и принадлежит к семейству Matonaviridae Rubivirus , представители которого обычно имеют геном одноцепочечной РНК положительной полярности , окруженный икосаэдрическим капсидом .
По состоянию на 1999 год [update] Молекулярная основа возникновения синдрома врожденной краснухи еще не была полностью ясна, но исследования in vitro на клеточных линиях показали, что вирус краснухи оказывает апоптотическое действие на определенные типы клеток. Имеются данные о p53 -зависимом механизме. [2]
Таксономия
[ редактировать ]Вирус краснухи ( Rubivirus Rubellae ) отнесен к роду Rubivirus . [1]
матонавирусов Семейство
[ редактировать ]До 2018 года рубивирусы классифицировались как часть семейства Togaviridae , но с тех пор были изменены и стали единственным родом семейства Matonaviridae . Это семейство названо в честь Жоржа де Матона, который в 1814 году впервые отличил краснуху от кори и скарлатины . [3] Изменение было внесено Международным комитетом по таксономии вирусов (ICTV), центральным руководящим органом по классификации вирусов . Matonviridae остается частью области, в которой они уже находились как Togaviridae , Riboviria , из-за своего РНК-генома и РНК-зависимой РНК-полимеразы . [3]
Другие рубивирусы
[ редактировать ]В 2020 году вирус Рухугу и вирус Рустрела присоединились к вирусу краснухи как второй и третий из трех представителей рода Rubivirus . [4] Ни один из них, как известно, не заражает людей. [5]
Морфология
[ редактировать ]Вирионы альфавируса имеют сферическую форму и содержат икосаэдрический нуклеокапсид , вирионы RuV плейоморфны и не содержат икосаэдрических нуклеокапсидов. [3]
Филогения
[ редактировать ]ICTV проанализировала последовательность RuV и сравнила его филогению с филогенией тогавирусов. Они пришли к выводу:
Филогенетический анализ РНК-зависимой РНК-полимеразы альфавирусов, вируса краснухи и других РНК-вирусов с положительным смыслом показывает, что эти два рода внутри Togaviridae не являются монофилетическими. В частности, вирус краснухи более тесно группируется с членами семейств Benyviridae , Hepeviridae и Alphatetraviridae , а также с несколькими неклассифицированными вирусами, чем с членами семейства Togaviridae, принадлежащими к роду Alphavirus . [3]
Структура
[ редактировать ]Сферические вирусные частицы ( вирионы ) матонавирусов имеют диаметр от 50 до 70 нм и покрыты липидной мембраной ( вирусной оболочкой ), происходящей из мембраны клетки-хозяина. Имеются заметные «шипы» (выступы) размером 6 нм, состоящие из белков вирусной оболочки E1 и E2, встроенных в мембрану. [6]
Гликопротеин Е1 считается иммунодоминантным в гуморальном ответе, индуцированном против структурных белков, и содержит как нейтрализующие, так и гемагглютинирующие детерминанты.
| Род | Структура | Симметрия | Капсид | Геномное расположение | Геномная сегментация |
|---|---|---|---|---|---|
| Рубивирус | икосаэдрический | Т=4 | обернутый | Линейный | Однодольный |
Капсидный белок
[ редактировать ]Внутри липидной оболочки находится капсид диаметром 40 нм. Капсидный белок (CP) выполняет различные функции. [8] Его основными задачами являются образование гомоолигомеров для формирования капсида и связывание геномной РНК. Кроме того, он отвечает за агрегацию РНК в капсиде, взаимодействует с мембранными белками Е1 и Е2 и связывает человеческий белок-хозяин p32, который важен для репликации вируса в хозяине. [9]
В отличие от альфавирусов капсид не подвергается аутопротеолизу, а отсекается от остальной части полипротеина сигнальной пептидазой . Образование капсида происходит на поверхности внутриклеточных мембран одновременно с почкованием вируса. [10]
Геном
[ редактировать ]Геном с положительной цепью РНК состоит из 9762 нуклеотидов и кодирует 2 неструктурных полипептида (p150 и p90) в пределах своих 5'-концевых двух третей и 3 структурных полипептида (C, E2 и E1) в пределах своей 3'-концевой трети. [11] Оба белка оболочки E1 и E2 гликозилированы .
У матонавирусов есть три сайта, которые являются высококонсервативными: структура «стебель-петля» на 5'-конце генома, консервативная последовательность из 51 нуклеотида вблизи 5'-конца генома и консервативная последовательность из 20 нуклеотидов на 5'-конце генома. Стартовый сайт субгеномной РНК. Гомологичные последовательности присутствуют в геноме краснухи. [11]
Геном кодирует несколько некодирующих структур РНК; среди них есть 3'-цис-действующий элемент вируса краснухи , который содержит несколько стеблевых петель , одна из которых, как было обнаружено, необходима для репликации вируса. [12]
Единственная значительная область гомологии между краснухой и альфавирусами расположена на NH2-конце неструктурного белка 3. Эта последовательность обладает геликазной и репликазной активностью. В геноме краснухи они встречаются в ориентации, противоположной той, что обнаружена в альфавирусах, что указывает на то, что произошла перестройка генома.
Геном имеет самое высокое содержание G+C среди всех известных в настоящее время вирусов с одноцепочечной РНК (~70%). [13] Несмотря на такое высокое содержание GC, использование его кодонов аналогично использованию его человека-хозяина.
Репликация
[ редактировать ]Вирусы прикрепляются к поверхности клетки через специфические рецепторы и поглощаются эндосомой образующейся . При нейтральном pH вне клетки белок оболочки E2 покрывает белок E1. Падение pH внутри эндосомы освобождает внешний домен E1 и вызывает слияние вирусной оболочки с мембраной эндосомы. Таким образом, капсид достигает цитозоля , распадается и освобождает геном.
+оцРНК ( положительная одноцепочечная РНК ) сначала действует только как матрица для трансляции неструктурных белков, которые синтезируются в виде большого полипротеина , а затем разрезаются на отдельные белки. Последовательности структурных белков сначала реплицируются вирусной РНК-полимеразой (репликазой) через комплементарную -оцРНК в качестве матрицы и транслируются как отдельная короткая мРНК. Эта короткая субгеномная РНК дополнительно упакована в вирион. [14]
Трансляция структурных белков приводит к образованию большого полипептида (110 кДа ). Затем его эндопротеолитически разрезают на E1, E2 и капсидный белок. Е1 и Е2 представляют собой трансмембранные белки типа I , которые транспортируются в эндоплазматический ретикулум (ЭР) с помощью N-концевой сигнальной последовательности. Из ЭР гетеродимерный комплекс Е1·Е2 достигает аппарата Гольджи , где происходит отпочкование новых вирионов (в отличие от альфа-вирусов, у которых отпочкование происходит на плазматической мембране).С другой стороны, капсидные белки остаются в цитоплазме и взаимодействуют с геномной РНК, вместе образуя капсид. [15]
| Род | Сведения о хосте | Тканевой тропизм | Детали входа | Подробности выпуска | Сайт репликации | Монтажный участок | Передача инфекции |
|---|---|---|---|---|---|---|---|
| Рубивирус | Люди | Никто | Клатрин-опосредованный эндоцитоз | Секреция | Цитоплазма | Цитоплазма | Аэрозоль |
Передача инфекции
[ редактировать ]RuV передается между людьми через дыхание. [3]
Эпидемиология
[ редактировать ]На основании различий в последовательности белка Е1 описаны два генотипа, различающиеся на 8–10%. Они были разделены на 13 признанных генотипов - 1a, 1B, 1C, 1D, 1E, 1F, 1G, 1h, 1i, 1j, 2A, 2B и 2C.
Для типирования ВОЗ рекомендует минимальное окно, включающее нуклеотиды с 8731 по 9469. [16]
Генотипы 1a, 1E, 1F, 2A и 2B были выделены в Китае .
Генотип 1j был выделен только в Японии и на Филиппинах .
Генотип 1Е встречается в Африке , Америке , Азии и Европе .
Генотип 1G выделен в Беларуси , Кот-д'Ивуаре и Уганде .
Генотип 1С эндемичен только в Центральной и Южной Америке.
Генотип 2В был выделен в Южной Африке .
Генотип 2С выделен в России .
Литература
[ редактировать ]- Дэвид М. Найп, Питер М. Хоули и др. (ред.): Fields Virology 4. Auflage, Филадельфия, 2001 г.
- CM Fauquet, MA Mayo и др.: Восьмой отчет Международного комитета по таксономии вирусов , Лондон, Сан-Диего, 2005 г.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Беннетт А.Дж., Паски А.С., Эбингер А., Кун Дж.Х., Бишоп-Лилли К.А., Бир М., Голдберг Т.Л. (31 июля 2020 г.). «Создайте два новых вида и переименуйте один вид в род Rubivirus ( Hepelivirales : Matonaviridae )» (docx) . Международный комитет по таксономии вирусов (ICTV). Архивировано из оригинала 21 декабря 2022 года . Проверено 15 мая 2021 г.
- ^ Медьери К., Беренци К., Халазонетис Т.Д. и др. (июнь 1999 г.). «Участие p53-зависимого пути в апоптозе, индуцированном вирусом краснухи» . Вирусология . 259 (1): 74–84. дои : 10.1006/виро.1999.9757 . ПМИД 10364491 .
- ^ Перейти обратно: а б с д и ICTV. «Создать новое семейство Maonaviridae, включив в него род Rubivirus, удаленный из семейства Togaviridae Release» . Проверено 29 октября 2019 г. [ мертвая ссылка ]
- ^ Беннетт, Эндрю Дж.; Паски, Адриан К.; Эбингер, Арнт; Пфафф, Флориан; Праймер, Грит; Хопер, Дирк; Брайтаупт, Анжеле; Хойзер, Элиза; Ульрих, Райнер Г.; Кун, Йенс Х.; Бишоп-Лилли, Кимберли А. (7 октября 2020 г.). «Родственники вируса краснухи у различных млекопитающих» . Природа : 1–5. дои : 10.1038/s41586-020-2812-9 . ISSN 1476-4687 . ПМЦ 7572621 . ПМИД 33029010 .
- ^ Гиббонс, Энн (7 октября 2020 г.). «Недавно обнаруженные вирусы позволяют предположить, что «германская корь» передалась от животных к человеку» . наука . дои : 10.1126/science.abf1520 . Архивировано из оригинала 8 декабря 2022 года . Проверено 30 июня 2022 г.
- ^ Барделетти Г., Кесслер Н., Эймар-Генри М. (1975). «Морфология, биохимический анализ и нейраминидазная активность вируса краснухи». Арх. Вирол . 49 (2–3): 175–86. дои : 10.1007/BF01317536 . ПМИД 1212096 .
- ^ Перейти обратно: а б «Вирусная зона» . ЭксПАСи. Архивировано из оригинала 24 июня 2015 года . Проверено 15 июня 2015 г.
- ^ Чен М.Х., Icenogle JP (апрель 2004 г.). «Капсидный белок вируса краснухи модулирует репликацию вирусного генома и инфекционность вируса» . Журнал вирусологии . 78 (8): 4314–22. doi : 10.1128/jvi.78.8.4314-4322.2004 . ПМК 374250 . ПМИД 15047844 .
- ^ Битч, доктор медицинских наук, Эверитт Дж.С., Лоу Л.Дж., Хобман Т.К. (август 2005 г.). «Взаимодействие между капсидом вируса краснухи и белком-хозяином p32 важно для репликации вируса» . Дж. Вирол . 79 (16): 10807–20. doi : 10.1128/JVI.79.16.10807-10820.2005 . ПМЦ 1182682 . ПМИД 16051872 .
- ^ Битч, доктор медицинских наук, Хобман, Т.К. (июнь 2000 г.). «Капсид вируса краснухи связывается с белком p32 клетки-хозяина и локализуется в митохондриях» . Дж. Вирол . 74 (12): 5569–76. doi : 10.1128/JVI.74.12.5569-5576.2000 . ПМЦ 112044 . ПМИД 10823864 .
- ^ Перейти обратно: а б Домингес Дж., Ван С.И., Фрей Т.К. (июль 1990 г.). «Последовательность геномной РНК вируса краснухи: доказательства генетической перестройки в ходе эволюции тогавируса» . Вирусология . 177 (1): 225–38. дои : 10.1016/0042-6822(90)90476-8 . ПМК 7131718 . ПМИД 2353453 .
- ^ Чен, Миннесота; Фрей Т.К. (1999). «Мутагенный анализ 3'-цис-действующих элементов генома вируса краснухи» . Дж Вирол . 73 (4): 3386–3403. ПМК 104103 . ПМИД 10074193 .
- ^ Чжоу Ю, Чен X, Ушиджима Х, Фрей ТК (2012)Анализ использования оснований и кодонов вирусом краснухи. Арка Вирол
- ^ «Тогавирусы – Классификация и систематика» . Архивировано из оригинала 5 августа 2012 года . Проверено 10 мая 2009 г.
- ^ Гарбутт М., Лоу Л.М., Чан Х., Хобман Т.С. (май 1999 г.). «Роль гликопротеиновых доменов вируса краснухи в сборке вирусоподобных частиц» . Дж. Вирол . 73 (5): 3524–33. ПМК 104124 . ПМИД 10196241 .
- ^ «Стандартизация номенклатуры генетических характеристик вирусов краснухи дикого типа» (PDF) . Еженедельная эпидемиологическая рекомендация . 80 (14): 126–132. 2005. PMID 15850226 . Архивировано из оригинала (PDF) 18 мая 2009 года.