Вирус краснухи
| Rubivirus rubellae | |
|---|---|
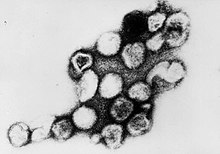
| |
| Электронная микрофотография вирионов вируса рубки | |
| Классификация вирусов | |
| (не вмешательство): | Вирус |
| Область : | Рибовирия |
| Королевство: | Orthornavirae |
| Филум: | Kitrinoviricota |
| Сорт: | Alsuviricetes |
| Заказ: | Hepelivirales |
| Семья: | Matonaviridae |
| Род: | Рубивирус |
| Разновидность: | Rubivirus rubellae
|
| Синонимы [ 1 ] | |
| |
Вирус краснухи ( RUV ) является патогенным агентом заболевания , передаваемой только между людьми по дыхательному пути, и является основной причиной синдрома врожденной краснухи, когда инфекция происходит в течение первых недель беременности .
Вирус краснухи, научное название rubellae , является членом рода Rubivirus и принадлежит к семейству Matonaviridae Rubivirus , чьи члены обычно имеют геном одноцепочечной РНК положительной полярности , которая заключена в икозаэдрическом капсиде .
По состоянию на 1999 [update] Молекулярная основа для причинности синдрома врожденной краснухи еще не была полностью ясной, но исследования in vitro с клеточными линиями показали, что вирус краснухи оказывает апоптотическое действие на определенные типы клеток. Существуют доказательства для P53 -зависимого механизма. [ 2 ]
Таксономия
[ редактировать ]Вирус краснухи ( Rubivirus rubellae ) назначен роду Rubivirus . [ 1 ]
Matonaviridae Семейство
[ редактировать ]До 2018 года рубивирусы были классифицированы как часть семьи Togaviridae , но с тех пор были изменены как единственный род семейства Matonaviridae . Эта семья названа в честь Джорджа де Матона, который в 1814 году впервые отличила краснуху от кори и скарлатины . [ 3 ] Изменение было внесено Международным комитетом по таксономии вирусов (ICTV), центральным руководящим органом для вирусной классификации . Matonaviridae остается частью царства, в которой он уже находился как Togaviridae , Riboviria , из -за его РНК -генома и РНК -зависимой РНК -полимеразы . [ 3 ]
Другие рубивирусы
[ редактировать ]В 2020 году вирус Рухугу и вирус Растрола присоединился к вирусу краснухи как секунды и думал только о трех членах Rubivirus рода [ 4 ] Ни один из них, как известно, заражает людей. [ 5 ]
Морфология
[ редактировать ]В то время как альфавирусные вирионы являются сферическими и содержат икосаэдрический нуклеокапсид , вирионы RUV являются плейоморфными и не содержат икосаэдрических нуклеокапсидов. [ 3 ]
Филогения
[ редактировать ]ICTV проанализировал последовательность RUV и сравнил его филогения с последованием тогавирусов. Они пришли к выводу:
Филогенетический анализ РНК-зависимой РНК-полимеразы альфавирусов, вируса краснухи и других вирусов РНК с положительным смыслом показывают два рода в тогавиридах не являются монофилетическими. В частности, вирусные группы более тесно связаны с членами семей Benyviridae , Hepeviridae и Alphatetraviridae , а также несколько неклассифицированных вирусов, чем с членами семейства Togaviridae, принадлежащих к роду альфавирус . [ 3 ]
Структура
[ редактировать ]Частицы сферических вирусов ( вирионы ) матонавиридов имеют диаметр от 50 до 70 нм и покрыты липидной мембраной ( вирусная оболочка ), полученная из мембраны клеток -хозяина. Существуют выдающиеся «шипы» (проекции) 6 нм, состоящие из белков вирусной оболочки E1 и E2, встроенных в мембрану. [ 6 ]
Гликопротеин E1 считается иммунодоминантным в гуморальном ответе, индуцированном против структурных белков, и содержит как нейтрализующие, так и гемагглютинирующие детерминанты.
| Род | Структура | Симметрия | Капсид | Геномное расположение | Геномная сегментация |
|---|---|---|---|---|---|
| Рубивирус | Икосаэдральный | T = 4 | Охвачен | Линейный | Монопартий |
Капсидный белок
[ редактировать ]Внутри липидной конверты диаметр 40 нм. Капсидный белок (CP) имеет разные функции. [ 8 ] Его основными задачами являются образование олигомеров гомо с образованием капсида и связывания геномной РНК. Кроме того, он ответственен за агрегацию РНК в капсиде, он взаимодействует с мембранными белками E1 и E2 и связывает белок-хозяин человека P32, который важен для репликации вируса у хозяина. [ 9 ]
В отличие от альфавирусов, капсид не подвергается автопротеолизу, скорее, он отрезан от остальной части полипротеина сигнальной пептидазой . Производство капсида происходит на поверхности внутриклеточных мембран одновременно с почкованием вируса. [ 10 ]
Геном
[ редактировать ]Геном РНК с положительной цепью имеет 9 762 нуклеотида и кодирует 2 Нтруктурные полипептиды (P150 и P90) в его 5'-концевых двух третях и 3 структурных полипептидах (C, E2 и E1) в пределах его 3'-концевой трети одной трети. [ 11 ] Оба белка оболочки E1 и E2 являются гликозилированными .
Есть три сайта, которые высоко консервативны в матонавирусах: структура стволовых и петли на 5 'конце генома, консервативная последовательность 51-нуклеотидов вблизи 5' конца генома и консервативная последовательность с 20 нуклеотидом в Субгеномный сайт начала РНК. Гомологичные последовательности присутствуют в геноме краснухи. [ 11 ]
Геном кодирует несколько некодирующих РНК- структур; Среди них элемент цис-действий вируса рубзы 3 ' , который содержит несколько петлей стеблей , один из которых был признан необходимым для репликации вируса. [ 12 ]
Единственная значимая область гомологии между краснухой и альфавирусами расположена на конце NH2 не структурного белка 3. Эта последовательность обладает геликазы и репликазы активностью . В геноме краснухи они встречаются в противоположной ориентации к тому, что встречается в альфавирусах, что указывает на то, что произошла перестройка генома.
Геном имеет самое высокое содержание G+C у всех в настоящее время известного одноцепочечного РНК -вируса (~ 70%). [ 13 ] Несмотря на это высокое содержание GC, его использование кодона аналогично содержимому его человеческому хозяину.
Репликация
[ редактировать ]Вирусы прикрепляются к клеточной поверхности через специфические рецепторы и поднимаются с эндосомой образованной . При нейтральном рН за пределами клетки белок E2 Envelope охватывает белок E1. Падение pH внутри эндосомы освобождает внешний домен E1 и вызывает слияние вирусной оболочки с эндосомальной мембраной. Таким образом, капсид достигает цитозоля , распадает и высвобождает геном
+SSRNA ( положительная одноцепочечная РНК ) сначала действует только как матрица для трансляции неструктурных белков, которые синтезируются как большой полипротеин и затем разрезают в отдельные белки. Последовательности для структурных белков сначала реплицируются вирусной РНК -полимеразой (репликазой) через комплементарную -ссРНК в качестве матрицы и транслируются как отдельная короткая мРНК. Эта короткая субгеномная РНК дополнительно упакована в вирионе. [ 14 ]
Перевод структурных белков продуцирует большой полипептид (110 кДа ). Затем это эндопротеолитически разрезается на E1, E2 и капсидный белок. E1 и E2 являются трансмембранными белками I типа , которые транспортируются в эндоплазматический ретикулум (ER) с помощью N-концевой сигнальной последовательности. Из ER гетеродимерный E1 · E2-Complex достигает аппарата Гольджи , где возникает подавление новых вирионов (в отличие от альфа-вирусов, где на плазматической мембране возникает почкуение. Капсидные белки, с другой стороны, остаются в цитоплазме и взаимодействуют с геномной РНК, вместе образуя капсид. [ 15 ]
| Род | Детали хоста | Ткань тропизм | Детали записи | Выпустить детали | Сайт репликации | Сайт сборки | Передача инфекции |
|---|---|---|---|---|---|---|---|
| Рубивирус | Люди | Никто | Клатрин-опосредованный эндоцитоз | Секреция | Цитоплазма | Цитоплазма | Аэрозоль |
Передача инфекции
[ редактировать ]RUV передается путем дыхания между людьми. [ 3 ]
Эпидемиология
[ редактировать ]На основании различий в последовательности белка E1 были описаны два генотипа, которые различаются на 8-10%. Они были подразделены на 13 распознанных генотипов - 1a, 1b, 1c, 1d, 1e, 1f, 1g, 1h, 1i, 1j, 2a, 2b и 2c.
Для печати ВОЗ рекомендует минимальное окно, которое включает нуклеотиды от 8731 до 9469. [ 16 ]
Генотипы 1a, 1e, 1f, 2a и 2b были изолированы в Китае .
Генотип 1J был выделен только из Японии и Филиппин .
Генотип 1E встречается в Африке , Америке , Азии и Европе .
Генотип 1G был изолирован в Беларуси , Кот -д'Ивуаре и Уганде .
Генотип 1C является эндемичным только в Центральной и Южной Америке.
Генотип 2B был изолирован в Южной Африке .
Генотип 2C был изолирован в России .
Литература
[ редактировать ]- Дэвид М. Книпп, Питер М. Хоули и соавт. (Ред.): Fields Viologiery 4. Auflage, Филадельфия 2001
- CM Fauquet, Ma Mayo et al.: Восьмой доклад Международного комитета по таксономии вирусов , Лондон Сан -Диего 2005
Ссылки
[ редактировать ]- ^ Jump up to: а беременный Bennett AJ, Paskey AC, Ebinger A, Kuhn JH, Bishop-Lilly KA, Beer M, Goldberg TL (31 июля 2020 года). «Создайте два новых вида и переименуйте один вид в роде Rubivirus ( Hepelivirales : Matonaviridae )» (Docx) . Международный комитет по таксономии вирусов (ICTV). Архивировано из оригинала 21 декабря 2022 года . Получено 15 мая 2021 года .
- ^ Megyei K, Beencsi K, Halazonets TD, et al. (Июнь 1999 г.). «Участие p53-зависимого пути в апоптоз, вызванный вирусом краснухи » Вирусология 259 (1): 74–8 Doi : 10.1006/ viro.1999.9 PMID 103644491
- ^ Jump up to: а беременный в дюймовый и ICTV. «Создайте новое семейство Matonaviridae, чтобы включить род Rubivirus, удаленный из семейства выпуска Togaviridae» . Получено 29 октября 2019 года . [ мертвая ссылка ]
- ^ Беннетт, Эндрю Дж.; Паски, Адриан С.; Эбингер, Арнт; Пфафф, Флориан; Пример, Грит; Хёпер, Дирк; Breithaupt, Angele; Heuser, Elisa; Ульрих, Рейнер Г.; Kuhn, Jens H.; Епископ-Лейли, Кимберли А. (7 октября 2020 г.). «Родственники вируса краснухи у разнообразных млекопитающих» . Природа : 1–5. doi : 10.1038/s41586-020-2812-9 . ISSN 1476-4687 . PMC 7572621 . PMID 33029010 .
- ^ Гиббонс, Энн (7 октября 2020 года). «Недавно обнаруженные вирусы предполагают, что« Немецкая корь »прыгнула с животных к людям» . наука . doi : 10.1126/science.abf1520 . Архивировано из оригинала 8 декабря 2022 года . Получено 30 июня 2022 года .
- ^ Барделетти Г., Кесслер Н., Аймард-Хенри М. (1975). «Морфология, биохимический анализ и нейрораминидазная активность вируса краснухи». Архи Вирол 49 (2–3): 175–86. doi : 10.1007/bf01317536 . PMID 1212096 .
- ^ Jump up to: а беременный «Вирусная зона» . Расширение. Архивировано из оригинала 24 июня 2015 года . Получено 15 июня 2015 года .
- ^ Chen MH, Icenogle JP (апрель 2004 г.). «Капсидный белок вируса рубки модулирует репликацию вирусного генома и инфекционность вируса» . Журнал вирусологии . 78 (8): 4314–22. doi : 10.1128/jvi.78.8.4314-4322.2004 . PMC 374250 . PMID 15047844 .
- ^ Beatch MD, Everitt JC, Law LJ, Hobman TC (август 2005 г.). «Взаимодействие между капсидом вируса краснухи и белком -хозяином P32 важны для репликации вируса» . J. Virol . 79 (16): 10807–20. doi : 10.1128/jvi.79.16.10807-10820.2005 . PMC 1182682 . PMID 16051872 .
- ^ Beatch MD, Hobman TC (июнь 2000 г.). «Капсид вируса рубки ассоциируется с клеточным белком -хозяином P32 и локализуется в митохондриях» . J. Virol . 74 (12): 5569–76. doi : 10.1128/jvi.74.12.5569-5576.2000 . PMC 112044 . PMID 10823864 .
- ^ Jump up to: а беременный Домингес Г., Ван Сай, Фрей Т.К. (июль 1990 г.). «Последовательность РНК генома вируса краснухи: свидетельство генетической перестройки во время эволюции тогавируса» . Вирусология . 177 (1): 225–38. doi : 10.1016/0042-6822 (90) 90476-8 . PMC 7131718 . PMID 2353453 .
- ^ Чен, MH; Фрей Т.К. (1999). «Мутагенный анализ 3 'цис-действий элементов генома вируса рубки» . J Virol . 73 (4): 3386–3403. PMC 104103 . PMID 10074193 .
- ^ Zhou Y, Chen X, Ushijima H, Frey TK (2012) Анализ использования основания и кодона вирусом краснухи. Арх Вирол
- ^ «Togaviridae- классификация и таксономия» . Архивировано из оригинала 5 августа 2012 года . Получено 10 мая 2009 года .
- ^ Garbutt M, Law LM, Chan H, Hobman TC (май 1999). «Роль гликопротеиновых доменов вируса рубки в сборке вирус-подобных частиц» . J. Virol . 73 (5): 3524–33. PMC 104124 . PMID 10196241 .
- ^ «Стандартизация номенклатуры для генетических характеристик вирусов краснухи дикого типа» (PDF) . Wkly Epidemiol Rec . 80 (14): 126–132. 2005. PMID 15850226 . Архивировано из оригинала (PDF) 18 мая 2009 года.