Кактус
| Кактус Временный диапазон: Поздний эоцен - недавний
| |
|---|---|
| Различные Cactaceae от Brockhaus 'Konversations-lexikon v. 2 1892 | |
| Научная классификация | |
| Королевство: | Plantae |
| Клада : | Трахеофиты |
| Клада : | Покрытосеменные |
| Клада : | Eudicots |
| Заказ: | Caryophylles |
| Семья: | Cactaceae Юсс. [ 1 ] |
| Подсемейства | |
|
Смотрите также классификацию Cactaceae | |
| Синонимы [ 2 ] | |
| |



Кактус кактус ( пл.: Кактусы , кактусы или , ) реже [ 3 ] является членом растений семейства Cactaceae ( / k æ K ˈ t eɪ s i . iː , - ˌ aɪ / ), [ А ] Семейство, состоящее из 127 родов с около 1750 известными видами ордена Caryophylles . [ 4 ] Слово кактус происходит через латынь, от древнегреческого слова κάκτος ( káktos ), название, первоначально используемое Теофрастом для колючего растения, чья идентичность теперь не уверена. [ 5 ] Кактусы встречаются в широком диапазоне форм и размеров. Они родом из Америки, начиная от Патагонии на юге до некоторых частей Западной Канады на севере, за исключением Rhipsalis Baccifera , которая также встречается в Африке и Шри -Ланке . Кактусы адаптированы для жизни в очень сухой среде, в том числе в пустыне Атакама , одно из самых сухих мест на земле. Из -за этого кактусы демонстрируют много адаптаций, чтобы сохранить воду. Например, почти все кактусы являются суккулентами , что означает, что они утолщенные, мясистые детали, адаптированные для хранения воды. В отличие от многих других суккулентов, стебель является единственной частью большинства кактусов, где происходит этот жизненно важный процесс. Большинство видов кактусов потеряли истинные листья, удерживая только шипы , которые являются сильно модифицированными листьями. Помимо защиты от травоядных животных , шипы помогают предотвратить потерю воды за счет уменьшения потока воздуха вблизи кактуса и обеспечивая некоторую тень. В отсутствие истинных листьев увеличенные стебли кактуи проводят фотосинтез .
Колючицы кактуса производятся из специализированных конструкций, называемых Areoles , своего рода сильно уменьшенной ветвей. Ареолы являются идентифицирующей особенностью кактусов. Наряду с шипами, ареолы дают цветы , которые обычно трубчаты и размножаются. Многие кактусы имеют короткие вегетационные времена и длинные общежития и могут быстро реагировать на любые осадки, которые помогают обширная, но относительно неглубокая корневая система, которая быстро поглощает любую воду, достигающую поверхности земли. Стебли кактуса часто ребристые или рифмы с несколькими ребрами, что соответствует числу в числах Фибоначчи (2, 3, 5, 8, 13, 21, 34 и т. Д.). Это позволяет им легко расширяться и сжиматься на быстрое поглощение воды после дождя, после чего задерживается в течение длительных засухи периодов . Как и другие суккулентные растения, большинство кактусов используют специальный механизм, называемый « метаболизм кислой кислоты » (CAM) как часть фотосинтеза. Транспирация , во время которого углекислый газ входит в растение и воду, не происходит в течение дня в то же время, что и фотосинтез, а вместо этого происходит ночью. Завод хранит углекислый газ, который принимает как Мололовая кислота , удерживая ее до возвращения дневного света, и только затем использование ее в фотосинтезе. Поскольку транспирация происходит во время прохладного, более влажного ночного часа, потеря воды значительно снижается.
Многие меньшие кактусы имеют штоки в форме шара, сочетающие максимально возможный объем для хранения воды с минимальной возможной площадью поверхности для потери воды от транспирации . Самый высокий [ B ] Отдельно стоящий кактус- Pachycereus pringlei , с максимальной зарегистрированной высотой 19,2 м (63 фута), [ 7 ] и наименьшим является Blossfeldia liliputiana , только около 1 см (0,4 дюйма) в диаметре при зрелости. [ 8 ] Говорят, что полностью выросший сагуаро ( Carnegiea gigantea ) способен поглощать до 200 галлонов США (760 л; 170 им. Гал) воды во время ливня. [ 9 ] Несколько видов значительно отличаются по внешнему виду от большей части семейства. По крайней мере, поверхностно, растения родов Leuenbergeria , Rhodocactus и Pereskia напоминают другие деревья и кустарники, растущие вокруг них. У них есть постоянные листья, и когда старые, покрытые корой стеблями. Их ареолы идентифицируют их как кактусы, и, несмотря на их внешность, у них тоже есть много адаптации к сохранению воды . Leuenbergeria считается близкой к наследственным видам, из которых эволюционировали все кактусы. В тропических регионах другие кактусы растут как лесные альпинисты и эпифиты (растения, которые растут на деревьях). Их стебли, как правило, сплющенные, почти листообразные по внешности, с меньшим количеством или даже без шипов, таких как известный рождественский кактус или кактус Дня благодарения (в роде Schlumbergera ).
Cacti have a variety of uses: many species are used as ornamental plants, others are grown for fodder or forage, and others for food (particularly their fruit). Cochineal is the product of an insect that lives on some cacti.
Many succulent plants in both the Old and New World – such as some Euphorbiaceae (euphorbias) – are also spiny stem succulents and because of this are sometimes incorrectly referred to as "cactus".[citation needed]
Morphology

The 1,500 to 1,800 species of cacti mostly fall into one of two groups of "core cacti": opuntias (subfamily Opuntioideae) and "cactoids" (subfamily Cactoideae). Most members of these two groups are easily recognizable as cacti. They have fleshy succulent stems that are major organs of photosynthesis. They have absent, small, or transient leaves. They have flowers with ovaries that lie below the sepals and petals, often deeply sunken into a fleshy receptacle (the part of the stem from which the flower parts grow). All cacti have areoles—highly specialized short shoots with extremely short internodes that produce spines, normal shoots, and flowers.[10]
The remaining cacti fall into only two groups: three tree-like genera, Leuenbergeria, Pereskia and Rhodocactus (all formerly placed in Pereskia), and the much smaller Maihuenia. These two groups are rather different from other cacti,[10] which means any description of cacti as a whole must frequently make exceptions for them. Species of the first three genera superficially resemble other tropical forest trees. When mature, they have woody stems that may be covered with bark and long-lasting leaves that provide the main means of photosynthesis. Their flowers may have superior ovaries (i.e., above the points of attachment of the sepals and petals) and areoles that produce further leaves. The two species of Maihuenia have succulent but non-photosynthetic stems and prominent succulent leaves.[10]
Growth habit
Cacti show a wide variety of growth habits, which are difficult to divide into clear, simple categories.
- Arborescent cacti
Cacti can be tree-like (arborescent), meaning they typically have a single more-or-less woody trunk topped by several to many branches. In the genera Leuenbergeria, Pereskia and Rhodocactus, the branches are covered with leaves, so the species of these genera may not be recognized as cacti. In most other cacti, the branches are more typically cactus-like, bare of leaves and bark and covered with spines, as in Pachycereus pringlei or the larger opuntias. Some cacti may become tree-sized but without branches, such as larger specimens of Echinocactus platyacanthus. Cacti may also be described as shrubby, with several stems coming from the ground or from branches very low down, such as in Stenocereus thurberi.[11]
- Columnar cacti
Smaller cacti may be described as columnar. They consist of erect, cylinder-shaped stems, which may or may not branch, without a very clear division into trunk and branches. The boundary between columnar forms and tree-like or shrubby forms is difficult to define. Smaller and younger specimens of Cephalocereus senilis, for example, are columnar, whereas older and larger specimens may become tree-like. In some cases, the "columns" may be horizontal rather than vertical. Thus, Stenocereus eruca can be described as columnar even though it has stems growing along the ground, rooting at intervals.[11]
- Globular cacti
Cacti whose stems are even smaller may be described as globular (or globose). They consist of shorter, more ball-shaped stems than columnar cacti. Globular cacti may be solitary, such as Ferocactus latispinus, or their stems may form clusters that can create large mounds. All or some stems in a cluster may share a common root.[11]
- Other forms
Other cacti have a quite different appearance. In tropical regions, some grow as forest climbers and epiphytes. Their stems are typically flattened and almost leaf-like in appearance, with few or even no spines. Climbing cacti can be very large; a specimen of Hylocereus was reported as 100 meters (330 ft) long from root to the most distant stem. Epiphytic cacti, such as species of Rhipsalis or Schlumbergera, often hang downwards, forming dense clumps where they grow in trees high above the ground.[11]
-
 Treelike habit (Pereskia aculeata)
Treelike habit (Pereskia aculeata) -
 Tall treelike habit (Pachycereus pringlei)
Tall treelike habit (Pachycereus pringlei) -
 Tall unbranched columnar habit (Cephalocereus)
Tall unbranched columnar habit (Cephalocereus) -
 Shorter clustered columnar habit (Ferocactus pilosus)
Shorter clustered columnar habit (Ferocactus pilosus) -
 Solitary globular habit (Ferocactus echidne)
Solitary globular habit (Ferocactus echidne) -
 Clustered globular habit (Rebutia species)
Clustered globular habit (Rebutia species) -
 Epiphytic cactus (Rhipsalis paradoxa)
Epiphytic cactus (Rhipsalis paradoxa) -
 There is a wide variety of cacti at the Desert Botanical Garden
There is a wide variety of cacti at the Desert Botanical Garden








Stems

The leafless, spiny stem is the characteristic feature of the majority of cacti (all belonging to the largest subfamily, the Cactoideae). The stem is typically succulent, meaning it is adapted to store water. The surface of the stem may be smooth (as in some species of Opuntia) or covered with protuberances of various kinds, which are usually called tubercles. These vary from small "bumps" to prominent, nipple-like shapes in the genus Mammillaria and outgrowths almost like leaves in Ariocarpus species. The stem may also be ribbed or fluted in shape. The prominence of these ribs depends on how much water the stem is storing: when full (up to 90% of the mass of a cactus may be water), the ribs may be almost invisible on the swollen stem, whereas when the cactus is short of water and the stems shrink, the ribs may be very visible.[11]
The stems of most cacti are some shade of green, often bluish or brownish green. Such stems contain chlorophyll and are able to carry out photosynthesis; they also have stomata (small structures that can open and close to allow passage of gases). Cactus stems are often visibly waxy.[11]
Areoles





Areoles are structures unique to cacti. Although variable, they typically appear as woolly or hairy areas on the stems from which spines emerge. Flowers are also produced from areoles. In the genus Leuenbergeria, believed similar to the ancestor of all cacti, the areoles occur in the axils of leaves (i.e. in the angle between the leaf stalk and the stem).[12] In leafless cacti, areoles are often borne on raised areas on the stem where leaf bases would have been.
Areoles are highly specialized and very condensed shoots or branches. In a normal shoot, nodes bearing leaves or flowers would be separated by lengths of stem (internodes). In an areole, the nodes are so close together, they form a single structure. The areole may be circular, elongated into an oval shape, or even separated into two parts; the two parts may be visibly connected in some way (e.g. by a groove in the stem) or appear entirely separate (a dimorphic areole). The part nearer the top of the stem then produces flowers, the other part spines. Areoles often have multicellular hairs (trichomes) that give the areole a hairy or woolly appearance, sometimes of a distinct color such as yellow or brown.[11]
In most cacti, the areoles produce new spines or flowers only for a few years and then become inactive. This results in a relatively fixed number of spines, with flowers being produced only from the ends of stems, which are still growing and forming new areoles. In Pereskia, a genus close to the ancestor of cacti, areoles remain active for much longer; this is also the case in Opuntia and Neoraimondia.[11]
Leaves
The great majority of cacti have no visible leaves; photosynthesis takes place in the stems (which may be flattened and leaflike in some species). Exceptions occur in three (taxonomically, four) groups of cacti. All the species of Leuenbergeria, Pereskia and Rhodocactus are superficially like normal trees or shrubs and have numerous leaves with a midrib and a flattened blade (lamina) on either side. This group is paraphyletic, forming two taxonomic clades. Many cacti in the opuntia group (subfamily Opuntioideae) also have visible leaves, which may be long-lasting (as in Pereskiopsis species) or produced only during the growing season and then lost (as in many species of Opuntia).[11] The small genus Maihuenia also relies on leaves for photosynthesis.[13] The structure of the leaves varies somewhat between these groups. Opuntioids and Maihuenia have leaves that appear to consist only of a midrib.[14]
Even those cacti without visible photosynthetic leaves do usually have very small leaves, less than 0.5 mm (0.02 in) long in about half of the species studied and almost always less than 1.5 mm (0.06 in) long. The function of such leaves cannot be photosynthesis; a role in the production of plant hormones, such as auxin, and in defining axillary buds has been suggested.[15]
Spines
Botanically, "spines" are distinguished from "thorns": spines are modified leaves, and thorns are modified branches. Cacti produce spines, always from areoles as noted above. Spines are present even in those cacti with leaves, such as Pereskia, Pereskiopsis and Maihuenia, so they clearly evolved before complete leaflessness. Some cacti only have spines when young, possibly only when seedlings. This is particularly true of tree-living cacti, such as Rhipsalis and Schlumbergera, but also of some ground-living cacti, such as Ariocarpus.[11]
The spines of cacti are often useful in identification, since they vary greatly between species in number, color, size, shape and hardness, as well as in whether all the spines produced by an areole are similar or whether they are of distinct kinds. Most spines are straight or at most slightly curved, and are described as hair-like, bristle-like, needle-like or awl-like, depending on their length and thickness. Some cacti have flattened spines (e.g. Sclerocactus papyracanthus). Other cacti have hooked spines. Sometimes, one or more central spines are hooked, while outer spines are straight (e.g., Mammillaria rekoi).[11]
In addition to normal-length spines, members of the subfamily Opuntioideae have relatively short spines, called glochids, that are barbed along their length and easily shed. These enter the skin and are difficult to remove due to being very fine and easily broken, causing long-lasting irritation.[11]
-
 Varied spines of a Ferocactus
Varied spines of a Ferocactus -
 Hooked central spine (cf. Mammillaria rekoi)
Hooked central spine (cf. Mammillaria rekoi) -
 Unusual flattened spines of Sclerocactus papyracanthus
Unusual flattened spines of Sclerocactus papyracanthus -
 Glochids of Opuntia microdasys
Glochids of Opuntia microdasys




Roots
Most ground-living cacti have only fine roots, which spread out around the base of the plant for varying distances, close to the surface. Some cacti have taproots; in genera such as Ariocarpus, these are considerably larger and of a greater volume than the body. Taproots may aid in stabilizing the larger columnar cacti.[16] Climbing, creeping and epiphytic cacti may have only adventitious roots, produced along the stems where these come into contact with a rooting medium.[11]
Flowers


Like their spines, cactus flowers are variable. Typically, the ovary is surrounded by material derived from stem or receptacle tissue, forming a structure called a pericarpel. Tissue derived from the petals and sepals continues the pericarpel, forming a composite tube—the whole may be called a floral tube, although strictly speaking only the part furthest from the base is floral in origin. The outside of the tubular structure often has areoles that produce wool and spines. Typically, the tube also has small scale-like bracts, which gradually change into sepal-like and then petal-like structures, so the sepals and petals cannot be clearly differentiated (and hence are often called "tepals").[11] Some cacti produce floral tubes without wool or spines (e.g. Gymnocalycium)[17] or completely devoid of any external structures (e.g. Mammillaria).[11] Unlike the flowers of most other cacti, Pereskia flowers may be borne in clusters.[12]
Cactus flowers usually have many stamens, but only a single style, which may branch at the end into more than one stigma. The stamens usually arise from all over the inner surface of the upper part of the floral tube, although in some cacti, the stamens are produced in one or more distinct "series" in more specific areas of the inside of the floral tube.[11]
The flower as a whole is usually radially symmetrical (actinomorphic), but may be bilaterally symmetrical (zygomorphic) in some species. Flower colors range from white through yellow and red to magenta.[11]
Adaptations for water conservation
All cacti have some adaptations to promote efficient water use. Most cacti—opuntias and cactoids—specialize in surviving in hot and dry environments (i.e. are xerophytes), but the first ancestors of modern cacti were already adapted to periods of intermittent drought.[10] A small number of cactus species in the tribes Hylocereeae and Rhipsalideae have become adapted to life as climbers or epiphytes, often in tropical forests, where water conservation is less important.
Leaves and spines
The absence of visible leaves is one of the most striking features of most cacti. Pereskia (which is close to the ancestral species from which all cacti evolved) does have long-lasting leaves, which are, however, thickened and succulent in many species.[10] Other species of cactus with long-lasting leaves, such as the opuntioid Pereskiopsis, also have succulent leaves.[18] A key issue in retaining water is the ratio of surface area to volume. Water loss is proportional to surface area, whereas the amount of water present is proportional to volume. Structures with a high surface area-to-volume ratio, such as thin leaves, necessarily lose water at a higher rate than structures with a low area-to-volume ratio, such as thickened stems.
Spines, which are modified leaves, are present on even those cacti with true leaves, showing the evolution of spines preceded the loss of leaves. Although spines have a high surface area-to-volume ratio, at maturity they contain little or no water, being composed of fibers made up of dead cells.[14] Spines provide protection from herbivores and camouflage in some species, and assist in water conservation in several ways. They trap air near the surface of the cactus, creating a moister layer that reduces evaporation and transpiration. They can provide some shade, which lowers the temperature of the surface of the cactus, also reducing water loss. When sufficiently moist air is present, such as during fog or early morning mist, spines can condense moisture, which then drips onto the ground and is absorbed by the roots.[11]
Stems

The majority of cacti are stem succulents, i.e., plants in which the stem is the main organ used to store water. Water may form up to 90% of the total mass of a cactus. Stem shapes vary considerably among cacti. The cylindrical shape of columnar cacti and the spherical shape of globular cacti produce a low surface area-to-volume ratio, thus reducing water loss, as well as minimizing the heating effects of sunlight. The ribbed or fluted stems of many cacti allow the stem to shrink during periods of drought and then swell as it fills with water during periods of availability.[11] A mature saguaro (Carnegiea gigantea) is said to be able to absorb as much as 200 U.S. gallons (760 L; 170 imp gal) of water during a rainstorm.[9] The outer layer of the stem usually has a tough cuticle, reinforced with waxy layers, which reduce water loss. These layers are responsible for the grayish or bluish tinge to the stem color of many cacti.[11]
The stems of most cacti have adaptations to allow them to conduct photosynthesis in the absence of leaves. This is discussed further below under Metabolism.
Roots
Many cacti have roots that spread out widely, but only penetrate a short distance into the soil. In one case, a young saguaro only 12 cm (4.7 in) tall had a root system with a diameter of 2 m (7 ft), but no more than 10 cm (4 in) deep.[16] Cacti can also form new roots quickly when rain falls after a drought. The concentration of salts in the root cells of cacti is relatively high.[19] All these adaptations enable cacti to absorb water rapidly during periods of brief or light rainfall. Thus, Ferocactus cylindraceus reportedly can take up a significant amount of water within 12 hours from as little as 7 mm (0.3 in) of rainfall, becoming fully hydrated in a few days.[11]
Although in most cacti, the stem acts as the main organ for storing water, some cacti have in addition large taproots.[11] These may be several times the length of the above-ground body in the case of species such as Copiapoa atacamensis,[11] which grows in one of the driest places in the world, the Atacama Desert in northern Chile.[20]
Metabolism
Photosynthesis requires plants to take in carbon dioxide gas (CO2). As they do so, they lose water through transpiration. Like other types of succulents, cacti reduce this water loss by the way in which they carry out photosynthesis. "Normal" leafy plants use the C3 mechanism: during daylight hours, CO2 is continually drawn out of the air present in spaces inside leaves and converted first into a compound containing three carbon atoms (3-phosphoglycerate) and then into products such as carbohydrates. The access of air to internal spaces within a plant is controlled by stomata, which are able to open and close. The need for a continuous supply of CO2 during photosynthesis means the stomata must be open, so water vapor is continuously being lost. Plants using the C3 mechanism lose as much as 97% of the water taken up through their roots in this way.[21] A further problem is that as temperatures rise, the enzyme that captures CO2 starts to capture more and more oxygen instead, reducing the efficiency of photosynthesis by up to 25%.[22]
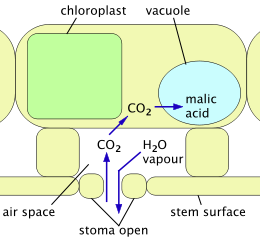

Crassulacean acid metabolism (CAM) is a mechanism adopted by cacti and other succulents to avoid the problems of the C3 mechanism. In full CAM, the stomata open only at night, when temperatures and water loss are lowest. CO2 enters the plant and is captured in the form of organic acids stored inside cells (in vacuoles). The stomata remain closed throughout the day, and photosynthesis uses only this stored CO2. CAM uses water much more efficiently at the price of limiting the amount of carbon fixed from the atmosphere and thus available for growth.[23] CAM-cycling is a less water-efficient system whereby stomata open in the day, just as in plants using the C3 mechanism. At night, or when the plant is short of water, the stomata close and the CAM mechanism is used to store CO2 produced by respiration for use later in photosynthesis. CAM-cycling is present in Pereskia species.[10]
By studying the ratio of 14C to 13C incorporated into a plant—its isotopic signature—it is possible to deduce how much CO2 is taken up at night and how much in the daytime. Using this approach, most of the Pereskia species investigated exhibit some degree of CAM-cycling, suggesting this ability was present in the ancestor of all cacti.[10] Pereskia leaves are claimed to only have the C3 mechanism with CAM restricted to stems.[24] More recent studies show that "it is highly unlikely that significant carbon assimilation occurs in the stem"; Pereskia species are described as having "C3 with inducible CAM."[10] Leafless cacti carry out all their photosynthesis in the stem, using full CAM. As of February 2012[update], it is not clear whether stem-based CAM evolved once only in the core cacti, or separately in the opuntias and cactoids;[10] CAM is known to have evolved convergently many times.[23]
To carry out photosynthesis, cactus stems have undergone many adaptations. Early in their evolutionary history, the ancestors of modern cacti (other than Leuenbergeria species) developed stomata on their stems and began to delay developing bark. However, this alone was not sufficient; cacti with only these adaptations appear to do very little photosynthesis in their stems. Stems needed to develop structures similar to those normally found only in leaves. Immediately below the outer epidermis, a hypodermal layer developed made up of cells with thickened walls, offering mechanical support. Air spaces were needed between the cells to allow carbon dioxide to diffuse inwards. The center of the stem, the cortex, developed "chlorenchyma" – a plant tissue made up of relatively unspecialized cells containing chloroplasts, arranged into a "spongy layer" and a "palisade layer" where most of the photosynthesis occurs.[25]
Taxonomy and classification


(below) A Melocactus, likely the first genus seen by Europeans
Naming and classifying cacti has been both difficult and controversial since the first cacti were discovered for science. The difficulties began with Carl Linnaeus. In 1737, he placed the cacti he knew into two genera, Cactus and Pereskia. However, when he published Species Plantarum in 1753—the starting point for modern botanical nomenclature—he relegated them all to one genus, Cactus. The word "cactus" is derived through Latin from the Ancient Greek κάκτος (kaktos), a name used by Theophrastus for a spiny plant,[26] which may have been the cardoon (Cynara cardunculus).[27]
Later botanists, such as Philip Miller in 1754, divided cacti into several genera, which, in 1789, Antoine Laurent de Jussieu placed in his newly created family Cactaceae. By the early 20th century, botanists came to feel Linnaeus's name Cactus had become so confused as to its meaning (was it the genus or the family?) that it should not be used as a genus name. The 1905 Vienna botanical congress rejected the name Cactus and instead declared Mammillaria was the type genus of the family Cactaceae. It did, however, conserve the name Cactaceae, leading to the unusual situation in which the family Cactaceae no longer contains the genus after which it was named.[28]
The difficulties continued, partly because giving plants scientific names relies on "type specimens". Ultimately, if botanists want to know whether a particular plant is an example of, say, Mammillaria mammillaris, they should be able to compare it with the type specimen to which this name is permanently attached. Type specimens are normally prepared by compression and drying, after which they are stored in herbaria to act as definitive references. However, cacti are very difficult to preserve in this way; they have evolved to resist drying and their bodies do not easily compress.[29] A further difficulty is that many cacti were given names by growers and horticulturalists rather than botanists; as a result, the provisions of the International Code of Nomenclature for algae, fungi, and plants (which governs the names of cacti, as well as other plants) were often ignored. Curt Backeberg, in particular, is said to have named or renamed 1,200 species without one of his names ever being attached to a specimen, which, according to David Hunt, ensured he "left a trail of nomenclatural chaos that will probably vex cactus taxonomists for centuries."[30]
Classification
In 1984, it was decided that the Cactaceae Section of the International Organization for Succulent Plant Study should set up a working party, now called the International Cactaceae Systematics Group (ICSG), to produce consensus classifications down to the level of genera. Their system has been used as the basis of subsequent classifications. Detailed treatments published in the 21st century have divided the family into around 125–130 genera and 1,400–1,500 species, which are then arranged into a number of tribes and subfamilies.[31][32][33] The ICSG classification of the cactus family recognized four subfamilies, the largest of which was divided into nine tribes. The subfamilies were:[31]



- Subfamily Pereskioideae K. Schumann
- The only genus in the ICSG classification was Pereskia. It has features considered closest to the ancestors of the Cactaceae. Plants are trees or shrubs with leaves; their stems are smoothly round in cross section, rather than being ribbed or having tubercles.[31] Two systems may be used in photosynthesis, both the "normal" C3 mechanism and crassulean acid metabolism (CAM)—an "advanced" feature of cacti and other succulents that conserves water.[10]
- Molecular phylogenetic studies showed that when broadly circumscribed, Pereskia was not monophyletic,[34][33] and it has been split into three genera, Leuenbergeria, Rhodocactus and a narrowly circumscribed Pereskia.[35][36] Leuenbergeria is then placed on its own in a separate monogeneric subfamily, Leuenbergerioideae.[37]
- Subfamily Opuntioideae K. Schumann
- Some 15 genera are included in this subfamily. They may have leaves when they are young, but these are lost later. Their stems are usually divided into distinct "joints" or "pads" (cladodes).[31] Plants vary in size from the small cushions of Maihueniopsis[38] to treelike species of Opuntia, rising to 10 m (33 ft) or more.[39]
- Subfamily Maihuenioideae P. Fearn
- The only genus is Maihuenia, with two species, both of which form low-growing mats.[13] It has some features that are primitive within the cacti. Plants have leaves, and crassulean acid metabolism is wholly absent.[31]
- Subfamily Cactoideae
- Divided into nine tribes, this is the largest subfamily, including all the "typical" cacti. Members are highly variable in habit, varying from tree-like to epiphytic. Leaves are normally absent, although sometimes very reduced leaves are produced by young plants. Stems are usually not divided into segments, and are ribbed or tuberculate. Two of the tribes, Hylocereeae and Rhipsalideae, contain climbing or epiphytic forms with a rather different appearance; their stems are flattened and may be divided into segments.[31]
Molecular phylogenetic studies have supported the monophyly of three of these subfamilies (not Pereskioideae),[33][34] but have not supported all of the tribes or even genera below this level; indeed, a 2011 study found only 39% of the genera in the subfamily Cactoideae sampled in the research were monophyletic.[33] Classification of the cacti currently remains uncertain and is likely to change.
Phylogeny and evolution
Phylogeny

A 2005 study suggested the genus Pereskia as then circumscribed (Pereskia sensu lato) was basal within the Cactaceae, but confirmed earlier suggestions it was not monophyletic, i.e., did not include all the descendants of a common ancestor. The Bayesian consensus cladogram from this study is shown below with subsequent generic changes added.[34][35][36]
| Cactaceae |
| ||||||||||||||||||||||||
A 2011 study using fewer genes but more species also found that Pereskia s.l. was divided into the same clades, but was unable to resolve the members of the "core cacti" clade. It was accepted that the relationships shown above are "the most robust to date."[33]
Leuenbergeria species (Pereskia s.l. Clade A) always lack two key features of the stem present in most of the remaining "caulocacti": like most non-cacti, their stems begin to form bark early in the plants' life and also lack stomata—structures that control admission of air into a plant and hence control photosynthesis. By contrast, caulocacti, including species of Rhodocactus and the remaining species of Pereskia s.s., typically delay forming bark and have stomata on their stems, thus giving the stem the potential to become a major organ for photosynthesis. (The two highly specialized species of Maihuenia are something of an exception.)[34][36]
The first cacti are thought to have been only slightly succulent shrubs or small trees whose leaves carried out photosynthesis. They lived in tropical areas that experienced periodic drought. If Leuenbergeria is a good model of these early cacti, then, although they would have appeared superficially similar to other trees growing nearby, they had already evolved strategies to conserve water (some of which are present in members of other families in the order Caryophyllales). These strategies included being able to respond rapidly to periods of rain, and keeping transpiration low by using water very efficiently during photosynthesis. The latter was achieved by tightly controlling the opening of stomata. Like Pereskia species today, early ancestors may have been able to switch from the normal C3 mechanism, where carbon dioxide is used continuously in photosynthesis, to CAM cycling, in which when the stomata are closed, carbon dioxide produced by respiration is stored for later use in photosynthesis.[10]
The clade containing Rhodocactus and Pereskia s.s. marks the beginnings of an evolutionary switch to using stems as photosynthetic organs. Stems have stomata and the formation of bark takes place later than in normal trees. The "core cacti" show a steady increase in both stem succulence and photosynthesis accompanied by multiple losses of leaves, more-or-less complete in the Cactoideae. One evolutionary question at present unanswered is whether the switch to full CAM photosynthesis in stems occurred only once in the core cacti, in which case it has been lost in Maihuenia, or separately in Opuntioideae and Cactoideae, in which case it never evolved in Maihuenia.[10]
Understanding evolution within the core cacti clade is difficult as of February 2012[update], since phylogenetic relationships are still uncertain and not well related to current classifications. Thus, a 2011 study found "an extraordinarily high proportion of genera" were not monophyletic, so were not all descendants of a single common ancestor. For example, of the 36 genera in the subfamily Cactoideae sampled in the research, 22 (61%) were found not monophyletic.[33] Nine tribes are recognized within Cactoideae in the International Cactaceae Systematics Group (ICSG) classification; one, Calymmantheae, comprises a single genus, Calymmanthium.[31] Only two of the remaining eight – Cacteae and Rhipsalideae – were shown to be monophyletic in a 2011 study by Hernández-Hernández et al. For a more detailed discussion of the phylogeny of the cacti, see Classification of the Cactaceae.
Parts of this article (those related to "Phylogeny") need to be updated. (May 2021) |
Evolutionary history
Никаких известных окаменелостей из кактусов не существует, чтобы пролить свет на свою эволюционную историю. [40] However, the geographical distribution of cacti offers some evidence. Except for a relatively recent spread of Rhipsalis baccifera to parts of the Old World, cacti are plants of South America and mainly southern regions of North America. This suggests the family must have evolved after the ancient continent of Gondwana split into South America and Africa, which occurred during the Early Cretaceous, around 145 to 101 million years ago.[41] Precisely when after this split cacti evolved is less clear. Older sources suggest an early origin around 90 – 66 million years ago, during the Late Cretaceous. More recent molecular studies suggest a much younger origin, perhaps in very Late Eocene to early Oligocene periods, around 35–30 million years ago.[40][42] Based on the phylogeny of the cacti, the earliest diverging group (Leuenbergeria) may have originated in Central America and northern South America, whereas the caulocacti, those with more-or-less succulent stems, evolved later in the southern part of South America, and then moved northwards.[34] Core cacti, those with strongly succulent stems, are estimated to have evolved around 25 million years ago.[40] Возможный стимул их эволюции, возможно, возвышался в центральных Андах , около 25–20 миллионов лет назад, что было связано с увеличением и различной засушкой. [ 34 ] Тем не менее, считается, что нынешнее разнообразие видов кактусов возникло только за последние 10–5 миллионов лет (от позднего миоцена до плиоцена ). Другие суккулентные растения, такие как Aizoaceae в Южной Африке, Didiereaceae на Мадагаскаре и род Agave в Америке, по -видимому, диверсифицировали одновременно, что совпало с глобальным расширением засушливых сред. [ 40 ]
Распределение

Кактусы обитают в различных регионах, от прибрежных равнин до высоких гор. За одним исключением, они родом из Америки , где их диапазон простирается от Патагонии до Британской Колумбии и Альберты в Западной Канаде . Существует ряд центров разнообразия. Для кактусов, адаптированных к засухе, тремя основными центрами являются Мексика и юго -запад США; юго -западные Анды , где они находятся в Перу , Боливии , Чили и Аргентине ; и Восточная Бразилия , вдали от бассейна Амазонки . Эпифитные и лазавливые и лазавливые кактуры, которые обязательно имеют разные центры разнообразия, поскольку они требуют влажной среды. Они в основном находятся в прибрежных горах и атлантических лесах юго -восточной Бразилии; в Боливии, которая является центром разнообразия для подсемейства Rhipsalideae ; и в лесных регионах Центральной Америки , где скалолазание Hylocereeae наиболее разнообразны. [ 43 ]
Rhipsalis Baccifera является исключением; Он родом как в Америке, так и в Старом Свете , где он встречается в тропической Африке , Мадагаскаре и Шри -Ланке . чтобы он был распространен в качестве семян в пищеварительных трактах перелетных Одна теория состоит в том , птиц ; Семена Рипсалиса адаптированы для распределения птиц. Популяции Старого Света являются полиплоидными и считаются отдельными подвидом, подтверждая идею о том, что распространение не было недавним. [ 44 ] Альтернативная теория - это вид, который первоначально пересек Атлантику на европейских кораблях, торговляя между Южной Америкой и Африкой, после чего птицы могли распространяться более широко. [ 45 ]
Натурализованные виды
Многие другие виды стали натурализованными за пределами Америки после того, как были представлены людьми, особенно в Австралии , Гавайях и Средиземноморье . В Австралии в 19 -м веке были введены виды Opuntia , особенно Opuntia Stricta для использования в качестве естественных сельскохозяйственных заборов и в попытке создать кохинеальную промышленность. Они быстро стали серьезной проблемой сорняков, но в настоящее время контролируются биологическими агентами, в частности, Cactorum моли . [ 46 ] Однако потенциал сорняков видов Opuntia в Австралии продолжается, что приводит ко всем опунтиоидным кактуи, за исключением того, что O. ficus-indica объявляется сорняками национального значения Австралийским комитетом сорняков в апреле 2012 года.
Аравийский полуостров имеет широкий спектр постоянно растущих, введенных популяций кактусов. Некоторые из них культивируются, [ 47 ] [ 48 ] Некоторые из них убегают от культивирования, а некоторые являются инвазивами, которые, как предполагается, являются декоративными побегами. [ 49 ] [ 50 ] [ 51 ]
Репродуктивная экология


Цветы кактусов опыляются насекомыми, птицами и летучими мышами. Известно, что ни один из них насыщенным ветром, и самоопыление происходит только у очень немногих видов; Например, цветы некоторых видов Fraiea не открываются ( Cleistogamy ). [ 52 ] Необходимость привлечения опылителей привела к эволюции синдромов опыления , которые определяются как группы «цветочных признаков, включая награды, связанные с привлечением и использованием определенной группы животных в качестве опылителей». [ 53 ]
Пчелы являются наиболее распространенными опылителями кактусов; Считается, что пчелоноводство было первым, кто развивался. [ 52 ] Дневные бабочки и ночные мотыльки связаны с различными синдромами опыления. Цветы с бабочкой обычно ярко окрашены, открываются днем, тогда как цветы, топливаемые на моли часто белые или бледные, открываются только вечером и ночью. [ 54 ] Например, Lophocereus schottii опыляется определенным видом моли, Upiga virescens , который также откладывает свои яйца среди развивающихся семян, его гусеницы позже потребляют. [ 54 ] Цветы этого кактуса имеют воронку в форме, белые до темно-розовые, длиной до 4 см (1,6 дюйма) и открыты ночью. [ 55 ]
Колибри являются значительными опылителями кактусов. Виды, показывающие типичный синдром, блюд-палочки колибри, имеют цветы с цветами в направлении красного конца спектра, пыльники и тычинки, которые выступают из цветов, и форма, которая не является радиально симметричной , с нижней губой, которая наклоняется вниз; Они производят большое количество нектара с относительно низким содержанием сахара. [ 56 ] Виды Schlumbergera , такие как S. truncata , имеют цветы, которые тесно связаны с этим синдромом. [ 57 ] Другие блюда, подавленные колибри, включают Cleistocactus и Disocactus . [ 52 ]
Известно, что развевание летучих мышей относительно редко встречается, но около четверти родов кактусов, как известно, опыляются летучими мышами -необычайно высокая доля, превышаемая среди эвдикотов только двумя другими семействами, оба с очень небольшим количеством родов. Сторонные кактусы, растущие в полусетровых областях, являются одними из тех, кто наиболее вероятен, будучи разбитыми летучими мышами; Это может быть связано с тем, что летучие мыши могут пройти значительные расстояния, поэтому эффективные опылители растений, которые широко растут друг от друга. Синдром опыления, связанный с летучими мышами, включает в себя тенденцию открываться цветами вечером и ночью, когда летучие мыши активны. Другие функции включают относительно скучный цвет, часто белый или зеленый; радиально симметричная форма, часто трубчатая; запах, описанный как «затхлый»; и производство большого количества богатого сахаром нектара. Carnegiea gigantea является примером сложившегося летучих мышей, как и многие виды Pachycereus и Pilosocereus . [ 58 ]

Фрукты , произведенные кактусы после оплодотворения, значительно различаются; Многие мясисты, хотя некоторые сухие. Все содержат большое количество семян. Мятистые, красочные и сладкие фрукты связаны с рассеиванием семян птицами. Семена проходят через их пищеварительные системы и осаждаются в их помете. Фрукты, которые падают на землю, могут быть съедены другими животными; Сообщается, что гигантские черепахи распространяют семена Опунтии на Галапагосских островах. Муравьи , по -видимому, рассеивают семена нескольких родов, таких как Blossfeldia . Сухая колючие фрукты могут цепляться за мех млекопитающих или перемещаться по ветру. [ 59 ]
Использование
Ранняя история
По состоянию на март 2012 года [update]По -прежнему существуют противоречия в отношении точных дат, когда люди впервые вошли в те области нового мира, где обычно встречаются кактусы, и, следовательно, когда они могли бы сначала их использовать. Археологическое место в Чили была датирована около 15 000 лет назад, [ 60 ] Предполагая, что кактусы встречались бы до этого. Ранние доказательства использования кактусов включают в себя картины пещер в Серра да Капиваре в Бразилии , а также семена, найденные в древних средних (отходы) в Мексике и Перу , с датами, оцениваемыми в 12 000–9 000 лет назад. Охотники-собиратели, вероятно, собрали фрукты кактуса в дикой природе и вернули их в свои лагеря. [ 61 ]


Неизвестно, когда кактусы были сначала культивированы. Opuntias (Dockly Pears) использовали для различных целей ацтеками , империя которого, продолжительностью от 14 до 16 -го века, имела сложную систему садоводства. Их столицей 15 -го века был Теночтитлан (ныне Мехико ); Одним из объяснений происхождения имени является то, что оно включает в себя Nahuatl Word nōchtli , ссылаясь на плод Opuntia. [ 62 ] Герб Мексики показывает орела, расположенного на кактусе, держа змею, изображение в центре мифа о основании Теночтитлана. [ 63 ] Ацтеки символически связывали спелые красные плоды опунтии с человеческими сердцами; Точно так же, как фрукты гасит жажду, так и предлагая человеческим сердцам, чтобы Бог Солнце обеспечил продолжение движения. [ 64 ]
Европейцы впервые столкнулись с кактусами, когда они прибыли в новый мир в конце 15 -го века. Их первые выручки были в Вест -Индии , где найдено относительно мало родов кактусов; Одним из наиболее распространенных является род Melocactus . [ 65 ] Таким образом, Мелокакти, возможно, были среди первых кактусов, которые видели европейцы. Виды Melocactus присутствовали в английских коллекциях кактусов до конца 16 -го века (к 1570 году в соответствии с одним источником, [ 66 ] ) там, где их называли echinomelocactus , позже сокращенным до Melocactus Джозефом Питтоном де Турневиллем в начале 18 -го века. [ 67 ] Кактусы, как чисто декоративные виды, так и те, у кого есть съедобные фрукты, продолжали прибывать в Европу, поэтому Карл Линнеус смог назвать 22 вида к 1753 году. Один из них, его кактус Опунтиа (ныне часть Opuntia ficus-indica ), был описан как « Fructu Majore ... Nunc in Hispania et Lusitania » (с большими фруктами ... теперь в Испании и Португалии), что свидетельствует о его раннем использовании в Европе. [ 68 ] [ 69 ]
Еда

Растение, в настоящее время известное как Opuntia ficus-Indica , или индийский кактус, давно является важным источником пищи. Считается, что первоначальный вид прибыл из Центральной Мексики, хотя теперь это неясно, потому что коренные народы южной части Северной Америки развили и распространяли ряд садоводческих сортов ( сортов ), включая формы видов и гибридов с другими окунтами. Как фрукты, так и прокладки едят, первые часто под испанским тунцом , последним под названием Nopal . Культивируемые формы часто являются значительно менее колючими или даже бескласными. [ 70 ] Говорят, что индустрия Nopal в Мексике стоила 150 миллионов долларов США в 2007 году. [ 71 ] Индийский фиговый кактус, вероятно, уже присутствовал на Карибском бассейне, когда испанцы прибыли, и вскоре после этого был доставлен в Европу. Он быстро распространился в средиземноморской области, как естественно, так и при введении - настолько, что ранние ботаники предположили, что он был родным для этого района. За пределами Америки индийский фигнический кактус является важной коммерческой культурой на Сицилии , Алжире и других странах Северной Африки. [ 69 ] Плоды других опунтов также едят, как правило, под тем же названием, тунцом . Цветочные бутоны, особенно видов цилиндропунты , также потребляются. [ 72 ]
Почти любой мясистый фрукт кактуса съедобен. Слово Питайя или Питахая (обычно считалось, что было взято на испанский из гаитянского креольского [ 73 ] ) может быть применен в диапазоне «чешуйчатых фруктов», особенно с столбчатыми кактусами. Плод Сагуаро ( Carnegiea gigantea ) уже давно важен для коренных народов северо -западной Мексики и юго -западных Соединенных Штатов, включая пустыню Сонора . Его можно сохранить путем кипения для производства сиропа и сушки. Сироп также может быть ферментирован, чтобы произвести алкогольный напиток. Фрукты видов Stenocereus также были важными источниками пищи в аналогичных частях Северной Америки; Stenocereus Queretaroensis выращивается для его фруктов. В более тропических южных районах альпинист Selenicereus undatus предоставляет Pitahaya Orejona , теперь широко выращенный в Азии под названием Dragon Fruit. Другие кактусы, обеспечивающие съедобные плоды, включают виды Echinocereus , Ferocactus , Mammillaria , Myrtillocactus , Pachycereus , Peniocereus и Selenicereus . Тела кактусов, кроме Opuntias, едят реже, хотя Андерсон сообщил, что Neowerdermannia vorwerkii готовится и едят, как картофель в Голлендской Боливии. [ 74 ]
-
 Сбор Сагуаро в 1907 году фруктов
Сбор Сагуаро в 1907 году фруктов -
 Съедобный плод Сагуаро
Съедобный плод Сагуаро -
 Фрукты некоторых витокакта съедобны.
Фрукты некоторых витокакта съедобны. -
 Драконы фрукты на продажу на Тайване
Драконы фрукты на продажу на Тайване -
 Фрукты, приготовленные из stenocereus Queretaroensis
Фрукты, приготовленные из stenocereus Queretaroensis -
 Салат, включая нарезанные нопалес ( подушки Opuntia )
Салат, включая нарезанные нопалес ( подушки Opuntia )






Психоактивные агенты



Было показано, что ряд видов кактусов содержат психоактивные агенты , химические соединения, которые могут вызвать изменения в настроении, восприятии и познании благодаря их влиянию на мозг. Два вида имеют долгую историю использования коренными народами Америки: Пейоте, Лофофора Уильямси , в Северной Америке, и Сан -Педротус, Трихоцерес Макрогон Вар. Пачаной , в Южной Америке. Оба содержат мескалин . [ 75 ]
L. Williamsii родом из северной Мексики и южного Техаса. Отдельные стебли составляют около 2–6 см (0,8–2,4 дюйма) высотой с диаметром 4–11 см (1,6–4,3 дюйма) и могут быть обнаружены в комков до 1 м (3 фута) шириной. [ 76 ] Большая часть стебля обычно находится под землей. Мескалин сосредоточен в фотосинтетической части стебля над землей. Центр стебля, который содержит точку растущего (апикальная меристема ), затоплен. Опытные коллекционеры Peyote удаляют тонкий ломтик с верхней части растения, оставляя точку растущей точки нетронутой, что позволяет растению регенерировать. [ 77 ] Данные указывают на то, что Пейот использовался более 5500 лет назад; Сушеные кнопки Peyote, предположительно, находятся с сайта в Рио -Гранде, штат Техас, были радиоуглеродом, датированным около 3780–3660 гг. До н.э. [ 78 ] Пейот воспринимается как средство доступа к духовному миру. Попытки римско -католической церкви подавить его использование после того, как испанское завоевание были в значительной степени безуспешными, и к середине 20 -го века Пейот был более широко использован, чем когда -либо коренными народами на севере, до Канады. Сейчас он формально используется церковью коренных американцев . [ 75 ]
Trichocereus macrogonus var. Pachanoi (Syn. Echinopsis pachanoi ) является родом из Эквадора и Перу. Это очень отличается от внешнего вида от Л. Уильямсии . Он имеет высокие стебли, высотой до 6 м (20 футов), с диаметром 6–15 см (2,4–5,9 дюйма), который разветвляется от основания, придавая всем растению кустарник или дерево, похожий на дерево. [ 79 ] Археологические доказательства использования этого кактуса, по -видимому, датируются 2000–2300 годами назад, с резьбой и керамическими объектами, показывающими столбчатые кактусы. [ 80 ] Хотя церковные власти под испанским пытались подавить его использование, это потерпело неудачу, как показано христианским элементом в общем названии «Сан -Педротус» - Святого Петратуса. Андерсон приписывает название убеждению, что, как и Святой Петр, держит ключи от небес, последствия кактуса позволяют пользователям «достигать небес, пока все еще на земле». [ 75 ] Он продолжает использоваться для его психоактивных эффектов, как для духовных, так и для целей исцеления, часто в сочетании с другими психоактивными агентами, такими как Datura Ferox и Tobacco. [ 80 ] Несколько других видов Echinopsis , включая E. peruviana и E. lageniformis , также содержат мескалин. [ 75 ]
Декоративные растения



Кактусы культивировались как декоративные растения с того времени, когда они были впервые привезены из нового мира. К началу 1800 -х годов у энтузиастов в Европе были большие коллекции (часто включая другие суккуленты вместе с кактусами). Редкие растения были проданы по очень высоким ценам. Поставщики кактусов и других суккулентов использовали коллекционеров для получения растений из дикой природы, в дополнение к выращиванию собственного. В конце 1800 -х годов коллекционеры обратились к орхидеям, и кактусы стали менее популярными, хотя никогда не исчезали от культивирования. [ 81 ]
Кактусы часто выращивают в теплицах, особенно в регионах, не связанных с выращиванием кактусов на открытом воздухе, таких как северные части Европы и Северной Америки. Здесь они могут храниться в горшках или выращивать в земле. Кактусы также выращиваются в качестве комнатных растений , многие из которых терпимы к часто сухой атмосфере. Кактусы в горшках могут быть помещены на улицу летом в орнаментные сады или патио, а затем держатся под прикрытием зимой. [ 82 ] Менее устойчивые к засухе эпифиты , такие как гибриды Epiphyllum , Schlumbergera (День благодарения или рождественский кактус) и Хатиора (Пасхальный Кактус), широко выращиваются в качестве комнатных.

Кактусы также могут быть посажены на открытом воздухе в регионах с подходящим климатом. Забота о сохранении воды в засушливых регионах привела к продвижению садов, требующих меньшего водопада ( Xeriscaping ). Например, в Калифорнии муниципальный район в Восточном заливе спонсировал публикацию книги о растениях и ландшафтах для летнего сухого климата. [ 83 ] Кактусы-одна группа устойчивых к засухе растений, рекомендованных для сухого ландшафтного садоводства. [ 84 ]
Другое использование
Кактусов имеет много других применений. Они используются для человеческой пищи и в качестве корма для животных, обычно после сжигания шипов. [ 85 ] В дополнение к их использованию в качестве психоактивных агентов, некоторые кактусы работают в травяной медицине . Таким образом, практика использования различных видов Opuntia распространилась из Америки, где они, естественно, встречаются в других регионах, где они растут, такие как Индия . [ 86 ]
Cochineal - это красный краситель, продуцируемый масштабным насекомым , которое живет на видах Opuntia . Давно используемый народами Центральной и Северной Америки, спрос быстро упал, когда европейские производители начали производить синтетические красители в середине 19 -го века. Коммерческое производство в настоящее время увеличилось после роста спроса на натуральные красители. [ 87 ]
Кактусы используются в качестве строительных материалов. Живые заборы кактуса используются в качестве баррикад вокруг зданий, чтобы не дать людям входить. Они также привыкли к набору животных. Деревянные части кактусов, такие как Cereus repandus и Echinopsis atacamensis , используются в зданиях и в мебели. Рамки домов на плетении и магистрали, построенных сери народа Мексики, могут использовать части Сагуаро ( Carnegiea gigantea ). Очень тонкие шипии и волосы (трихомы) некоторых кактусов использовались в качестве источника волокна для заполнения подушек и в ткаке. [ 88 ]
Сохранение

Все кактусы включены в Приложение II Конвенции о международной торговле исчезающими видами дикой фауны и флоры (CITE), которые «перечисляют виды, которые теперь не обязательно находятся под угрозой исчезновения, но это может стать таковой, если торговля не контролируется». Контроль осуществляется путем проведения международной торговли в большинстве образцов кактусов незаконными, если не были выданы разрешения, по крайней мере, для экспорта. [ 89 ] Некоторые исключения разрешены, например, для «натурализованных или искусственно распространенных растений». [ 90 ] Некоторые кактусы, такие как все виды ArioCarpus и Discocactus , включены в более ограничительное приложение I, [ 90 ] Используется для «наиболее находящихся под угрозой исчезновения» вида. Они могут быть перемещены только между странами для некоммерческих целей, и только тогда, когда они сопровождаются разрешениями как на экспорт, так и импорт. [ 89 ]
Три основные угрозы для кактусов в дикой природе-это развитие, выпас скота и чрезмерное собрание. Развитие принимает множество форм. Строительство плотины возле Зимапана , Мексика , вызвало разрушение большой части естественной среды обитания Echinocactus grusonii . Городское развитие и автомагистрали уничтожили места обитания кактуса в некоторых частях Мексики, Нью -Мексико и Аризоны , включая пустыню Сонора . Преобразование земли в сельское хозяйство повлияло на популяции ариокарпуса Котсхубеянуса в Мексике, где были вспахнуты сухие равнины для выращивания кукурузы, а также в Copiapoa и Eulychnia в Чили , где склоны долины были посажены виноградными лозами. [ 91 ] Выпас, во многих областях, введенными животными, такими как козы, нанес серьезный ущерб популяциям кактусов (а также других растений); Два примера, приведенные Андерсоном, - это Галапагосские острова, как правило, и влияние на Браунгинге канделяри в Перу . Чрезмерное собрание кактусов для продажи значительно повлияло на некоторые виды. Например, типовая местность Pelecyphora strobiliformis возле Микухуаны , Мексика, была практически обнаружена из растений, которые были вырыты на продажу в Европе. Незаконный сбор кактусов из дикой природы продолжает представлять угрозу. [ 92 ] [ 93 ]
Сохранение кактусов может быть in situ или ex situ . Сохранение на месте включает в себя сохранение привычек посредством обеспечения правовой защиты и создания специально охраняемых областей, таких как национальные парки и запасы. Примеры таких охраняемых районов в Соединенных Штатах включают Национальный парк Биг Бенд , штат Техас; Национальный парк Джошуа Три , Калифорния; и национальный парк Сагуаро , Аризона. Латиноамериканские примеры включают Parque Nacional Del Pinacate , Sonora, Мексика и Национальный парк Пан де Азукар , Чили. Сохранение ex Situ направлена на сохранение растений и семян за пределами их естественной среды обитания, часто с намерением более поздних реинтродукции. Ботанические сады играют важную роль в ex situ сохранении ; Например, семена кактусов и других суккулентов хранятся в долгосрочном хранении в пустынном ботаническом саду , штат Аризона. [ 94 ]
Культивирование

Популярность кактусов означает, что многие книги посвящены их выращиванию. Кактусов естественным образом встречаются в широком диапазоне среды обитания, а затем выращиваются во многих странах с различным климатом, поэтому точно воспроизводить условия, в которых обычно растет вид, обычно не практично. [ 81 ] Можно провести широкое различие между кактусами Semidesert и эпифитными кактусами, которые нуждаются в разных условиях и лучше всего выращиваются отдельно. [ 95 ] Этот раздел в первую очередь связан с выращиванием кактусотую семидерта в контейнерах и под защитой, например, в теплице или в доме, а не культивировании снаружи в земле в тех климатических условиях, которые позволяют это. Для выращивания эпифитных кактусов см. В культивировании Schlumbergera (Рождественские или Дня Благодарения) и выращивание гибридов Epiphyllum .
Выращивание среда

Целью растущей среды является обеспечение поддержки и хранения воды, кислорода и растворенных минералов для кормления растения. [ 96 ] В случае кактусов существует общее согласие, что открытая среда с высоким содержанием воздуха важна. Когда кактусы выращиваются в контейнерах, рекомендации относительно того, как это следует достичь, сильно различаются; Майлз Андерсон говорит, что если бы попросили описать идеальную среду выращивания, «десять производителей дадут 20 различных ответов». [ 97 ] Роджер Браун предполагает смесь двух частей коммерческой выросшей среды , одной части гидропонной глины и одной части грубой пемзы или перлита , с добавлением почвы из понижений дождевого червя. [ 96 ] Общая рекомендация в размере 25–75% материала на основе органического основания, остальное, неорганическое, такое как пемза, перлит или зернистость, поддерживается другими источниками. [ 97 ] [ 98 ] [ 99 ] [ 100 ] Однако использование органического материала полностью отвергается другими; Хехт говорит, что кактусы (кроме эпифитов) «хотят почвы, которая имеет низкую или не имеющую гумуса », и рекомендует грубый песок в качестве основы растущей среды. [ 101 ]
Полив
Полудистые кактусы нуждаются в тщательном поливании. Общие советы трудно дать, поскольку требуемая частота полива зависит от того, где выращиваются кактусы, природу выращивания и первоначальную среду обитания кактусов. [ 102 ] Браун говорит, что больше кактусов теряется благодаря «безвременному применению воды, чем по любой другой причине», и что даже в течение бездейного зимнего сезона Хитчи нуждаются в некоторой воде. [ 103 ] Другие источники говорят, что вода может быть удержана зимой (с ноября по марш в северном полушарии). [ 95 ] Другая проблема - твердость воды; Там, где необходимо использовать жесткую воду , рекомендуется регулярное переосмысление , чтобы избежать накопления солей. [ 103 ] Общий совет, данный, заключается в том, что в течение вегетационного периода кактусы должны быть разрешены высыхание между тщательным поливом. [ 103 ] [ 104 ] [ 95 ] Счетчик воды может помочь в определении, когда почва сухая. [ 104 ]
Свет и температура
Несмотря на то, что полуотдельные кактусы могут подвергаться воздействию высоких уровней освещения в дикой природе, им все равно может потребоваться некоторое затенение, когда летом подвергается более высоким уровням света и температурой теплицы. [ 105 ] [ 106 ] Разрешение температуры поднимается выше 32 ° C (90 ° F), не рекомендуется. [ 106 ] Минимальная необходимая зимняя температура во многом зависит от вовлеченных видов кактуса. Для смешанной сбора часто предлагается минимальная температура между 5 ° C (41 ° F) и 10 ° C (50 ° F), за исключением холодных чувствительных родов, таких как Melocactus и Discocactus . [ 107 ] [ 95 ] Некоторые кактусы, особенно из высоких Анд , полностью заморозили, когда остаются сухими (например, опрокидывание Minuscula переживает температуру до -9 ° C (16 ° F) в выращивании [ 108 ] ) и может цвететь лучше при воздействии периода простуды. [ 109 ]
Распространение
Кактусы могут быть распространены семенами, черенками или прививками . Семена, посеянные в начале года, создают саженцы, которые выигрывают от более длительного растущего периода. [ 110 ] Семена поселяют в влажной среде для выращивания, а затем сохраняются в покрытой среде, до 7–10 дней после прорастания, чтобы избежать высыхания. [ 111 ] Очень влажная среда для выращивания может привести к гниению как семена, так и саженцев. [ 112 ] Температурный диапазон 18–30 ° C (64–86 ° F) предлагается для прорастания; Температура почвы около 22 ° C (72 ° F) способствует наилучшему росту корня. Низкие уровни освещения достаточны во время прорастания, но впоследствии полуудельные кактусы нуждаются в более высоких уровнях освещения, чтобы вызвать сильный рост, хотя акклиматизация необходима для условий в теплице, таких как более высокие температуры и сильный солнечный свет. [ 111 ]

Воспроизведение черенками использует части растения, которые могут выращивать корни. Некоторые кактусы производят «прокладки» или «суставы», которые можно отделить или чисто отрезать. Другие кактусы производят смещения, которые можно удалить. [ 110 ] В противном случае можно сделать черенки стебля, в идеале из относительно нового роста. Рекомендуется, чтобы любые срезанные поверхности могли высохнуть в течение нескольких дней до нескольких недель, пока не образуется мозоль над поверхностью разреза. Затем укоренение может происходить в соответствующей среде для выращивания при температуре около 22 ° C (72 ° F). [ 110 ] [ 111 ]

Прививка используется для видов, которые трудно выращивать в выращивании или не могут расти независимо, например, некоторые формы, не содержащие хлорофилла с белыми, желтыми или красными телами, или некоторые формы, которые показывают аномальный рост (например, кристат или чудовищные формы). Для растения -хозяина ( запас ) производители выбирают тот, который сильно растет при выращивании и совместимо с размножением растения: Scion . Произведение производителя делает сокращения как на складе, так и на Scion и соединяет их, связывая их вместе, пока они объединяются. Используются различные виды трансплантата - Flat Grafts, где как Scion, так и запасы имеют аналогичные диаметры, и расщелину трансплантатов, где меньший Scion вставлен в расщелину, сделанную в запасе. [ 113 ]
Коммерчески, огромное количество кактусов производится ежегодно. Например, в 2002 году только в Корее было распространено 49 миллионов заводов, стоимость которого составляет почти 9 миллионов долларов США. Большинство из них (31 миллион растений) были распространены путем прививки. [ 114 ]
Вредители и болезни
Ряд вредителей атакует кактусы в культивирование. Те, кто питается SAP, включают в себя Mealybugs , живущие как на стеблях, так и на корнях; масштабные насекомые , как правило, встречаются только на стеблях; белости , которые, как говорят, являются «нечастым» вредителем кактусов; [ 115 ] Красные пауки , которые очень маленькие, но могут происходить в большом количестве, построив прекрасную сеть вокруг себя и плохо отмечают кактус посредством их сосания SAP, даже если они не убивают его; и трипсы , которые особенно атакуют цветы. Некоторые из этих вредителей устойчивы ко многим инсектицидам , хотя есть биологический контроль . Корни кактусов могут быть съедены личинками Sciarid Mhlies и грибных комаров . Слизняки и улитки также едят кактусы. [ 116 ] [ 117 ]
Грибы, бактерии и вирусы атакуют кактусы, первые два, особенно когда растения переосмыслились. Гнитная гниль Fusarium может получить вход через рану и вызвать гниение, сопровождаемое красной фиолетовой плесенью. « Гельминоспорийная вызвана биполярисом » гниль [ 118 ] ); Виды Phytophthora также вызывают сходные гнибы у кактусов. Фунгициды могут иметь ограниченную ценность при борьбе с этими заболеваниями. [ 119 ] Несколько вирусов были обнаружены в клетках, в том числе кактус вирус X. По -видимому, они вызывают только ограниченные видимые симптомы, такие как хлоротические (бледно -зеленые) пятна и мозаичные эффекты (полосы и участки бледного цвета). [ 120 ] Однако у вида агавы был показан вирус кактуса X, особенно когда корни сухие. [ 121 ] Там нет обработки от вирусных заболеваний. [ 119 ]
Примечания
- ^ Хотя написания ботанических семей были в значительной степени стандартизированы, между ботаниками мало согласны с тем, как эти имена должны быть произносили. Суффикс -Aceae ˈ быть произнесен / . eɪ s i может iː / ( ay -see -ee ), / ˈ eɪ s i ˌ eɪ / ( ay -see -ay ), или / ˈ eɪ s iː / ( ay -see ).
- ^ Самый высокий живой кактус - это образец Pachycereus pringlei . Самым высоким кактусом, когда -либо измеряемым, был бесплодным кактусом Сагуаро , который взорвался в бурте в июле 1986 года; Это было 78 футов (24 м) ростом. [ 6 ]
Ссылки
- ^ Обновление классификации групп ), « 2009 ( покрытоперма Группа филогения покрытоперма филогения .1095-8339.2009.00996.x , HDL : 10654/18083
- ^ Org, реестр-миграция.gbif (14 февраля 2017 г.), , ( набор , Секретариат GBIF: таксономия магистра ) " Cactaceae данных " gbif.org 16 апреля 2017 года
- ^ «Кактус», онлайн-словарь Мерриам-Уэбстера , архивировав с оригинала 2012-02-02 , извлечен 2012-02-13
- ^ Christenhusz, MJM & Byng, JW (2016), «Количество известных видов растений в мире и его годового увеличения» , Phytotaxa , 261 (3): 201–17, doi : 10.11646/phytotaxa.261.3.1 , архивировано от Оригинал 2016-07-29
- ^ Джонсон, в; Smith, Ha & Stockdale, AP (2019), Упрощенные названия растений: их произношение, деривация и значение , Шеффилд, Йоркшир: 5M Publishing, p. 26, ISBN 9781910455067
- ^ Windstorm Fells 78-футовый кактус-самый большой в мире , август 1986 года, архивировал оригинал 2015-10-29 , извлечен 2015-08-04
- ^ Salak, M. (2000), «В поисках самого высокоготуса», кактус и суккулент , 72 (3)
- ^ Mauseth, James D., Mauseth Cactus Research: Blossfeldia Liliputiana , архивировав с оригинала 2012-01-31 , извлеченные 2012-02-13
- ^ Jump up to: а беременный Взгляд на национальные парки: Стоп № 3-Сагуаро (Carnegiea gigantea) , Служба национальных парков, Министерство внутренних дел США, архивное из оригинала 2011-10-26 , извлечено 2012-02-19
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л Edwards, EJ & Donoghue, MJ (2006), « Переския и происхождение жизненной формы кактуса» (PDF) , американский натуралист , 167 (6): 777–793, doi : 10.1086/504605 , pmid 16649155 , s2cid 832909 , архивировано из оригинала (PDF) 2012-02-13 , извлеченным 2012-02-08
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий с Т в v В Андерсон (2001) , с. 15–37
- ^ Jump up to: а беременный Андерсон (2001) , с. 566
- ^ Jump up to: а беременный Андерсон (2001) , с. 398
- ^ Jump up to: а беременный Mauseth (2007) , p
- ^ Mauseth, James D. (2007), «Крошечная, но сложная листвой листвы оставляет Cccur во многих« бесстыдных »кактуках (Cactaceae)», Международный журнал наук о растениях , 168 (6): 845–853, doi : 10.1086/518273 , S2CID 84247622 , с. 845
- ^ Jump up to: а беременный Биология кактусов , Университет Далхаузи, архивирована из оригинала 2012-02-20 , извлечен 2012-02-13
- ^ Андерсон (2001) , с. 347–348
- ^ Андерсон (2001) , с. 572
- ^ Гибсон, Артур С. и Нобель, Парк С. (1990), The Cactus Primer , издательство Гарвардского университета, ISBN 978-0-674-08991-4
- ^ Андерсон (2001) , с. 174
- ^ Raven, JA & Edwards, D. (2001), «Корни: эволюционное происхождение и биогеохимическая значимость», Journal of Experimental Botany , 52 (90001): 381–401, doi : 10.1093/jexbot/52. suppl_1.381 , pmid 11326045
- ^ Sharkey, Thomas (1988), «Оценка скорости фотосписка в листьях», Physiologia plantarum , 73 (1): 147–152, doi : 10.1111/j.1399-3054.1988.tb09205.x
- ^ Jump up to: а беременный 2003 , Rundel Keeley E. & ( ) Jon W. , Philip . 85186850 , Archived (PDF) из оригинала 2012-04-27 , получен 2012-02-19
- ^ Андерсон (2001) , с. 37
- ^ Edwards, Nyffeler & Donoghue (2005) , p. 1184
- ^ Johnson, AT & Smith, HA (1972), Имена растений упрощенные: их вывод и смысл произношения , Бакенхилл, Херефордшир: Книжный магазин Landsmans, ISBN 978-0-900513-04-6 , с. 19
- ^ Sonnante, G.; Pignone, D. & Hammer, K (2007), «Приручение артишока и кардона: от римского времени до геномного возраста», Анналы ботаники , 100 (5): 1095–1100, doi : 10.1093/aob/mcm127 , PMC 2759203 , PMID 17611191
- ^ Андерсон (2001) , с. 96
- ^ Андерсон (2001) , с. 93–94
- ^ Андерсон (2001) , с. 98
- ^ Jump up to: а беременный в дюймовый и фон глин Андерсон (2001) , с. 99–103
- ^ Охота, доктор, изд. (2006), Новый Lexicon (два тома) , Milborne Port: DH Books, ISBN 978-0-9538134-4-5 , цитируется в Bárcenas, Yesson & Hawkins
- ^ Jump up to: а беременный в дюймовый и фон Bárcenas, Rolando T.; Yesson, Chris & Hawkins, Julie A. (2011), «Мелекулярная система Cactaceae», ( 5 ) : 470–489 doi : 10.1111111111111111. , Cladistics 27 , S2CID
- ^ Jump up to: а беременный в дюймовый и фон Эдвардс, Эрика Дж.; Nyffeler, Reto & Donoghue, Michael J. (2005), «Базальная филогения кактуса: последствия Перескии (Cactaceae) для перехода к форме жизни кактуса», American Journal of Botany , 92 (7): 1177–1188, doi : 10.3732/ajb.92.7.1177 , PMID 21646140
- ^ Jump up to: а беременный 97 International , Кактус-приключения французском языке : ( ) на
- ^ Jump up to: а беременный в ( ) Rhodocactus Isssak Asai , Kazunoro Miytata & 2016 , : 7–12, archived , 2019-06-09 , получен 2021-04-04-2
- ^ , Интеграция рода Leuenbergeria Lodé на его собственной семье , 6–7 , , retrieved 2015-01-2
- ^ Андерсон (2001) , с. 399
- ^ Андерсон (2001) , с. 485
- ^ Jump up to: а беременный в дюймовый Аракаки, Моника; Кристин, Паскаль-Антуан; Nyffeler, Reto; Лендель, Анита; Эггли, урс; Огберн, Р. Мэтью; Сприггс, Элизабет; Мур, Майкл Дж. И Эдвардс, Эрика Дж. (2011-05-17), «Современные и недавние излучения основных мировых линий сочных растений», Труды Национальной академии наук , 108 (20): 8379–8384, Bibcode : 2011pnas..108.8379a , doi : 10.1073/pnas.1100628108 , PMC 3100969 , PMID 21536881
- ^ Андерсон (2001) , с. 37–38
- ^ Nyffeler, Reto (2002), «Филогенетические отношения в семействе кактусов (Cactaceae) на основе данных из Trnk/ matk и trnl-trnf последовательностей », American Journal of Botany , 89 (2): 312–326, doi : 10.3732/ ajb .89.2.312 , PMID 21669740
- ^ Андерсон (2001) , с. 39–40
- ^ Андерсон (2001) , с. 611
- ^ 2010 ) ( (PDF), Polibotanica, 29. , архивировано (PDF с 107–129 2013-10-2 ) 117–118
- ^ «Идентификация сорняков-колючая груша (общая)», Weeds Australia , австралийский комитет сорняков, архивный из оригинала 2012-05-04 , извлеченные 2012-02-14
- ^ Belgacem, Azaiez Ould; Аль-Фарси, Сафаа Мухаммед; Wawi, Hayel AL; Аль-Яфей, Хади Абдулла Шайф; Аль-Шарари, М.; Аль-Хамуди, Ахмед Мохамед; Лухаичи, Мунир (26–30 марта 2017 г.). Бесспинкий кактус на Аравийском полуострове: адаптивное поведение и производственные характеристики . Международный конгресс IX потуса и кохинеаль - «кулачковые культуры для более горячего и более сухого мира». Coquimbo , Чили: CGIAR . HDL : 20.500.11766/9182 . S2CID 199636444 .
- ^ «Кактус колючей груши-это« чудо »урожай для сухих областей-эксперты» , Saudi Gazette , 2017-11-27 , извлечен 2021-03-29
- ^ «Источники кактуса Файфа Ландшафт» , Saudi Gazette , 2009-05-21, архивировав с оригинала на 2009-05-21 , извлечен 2021-03-29
- ^ Аль-Робай, Сами Ассир; Хоуладар, Саад Мохаммед; Мохамед, Хайдар Абдальгадир; Ахмед, Абделазим Али (2018), « Cylindropuntia rosea (DC.) Backeb, (Cactacae): новая общая чужую запись во флоре Саудовской Аравии», журнал о биоразнообразии Азиатско-Тихоокеанского региона , 11 (2): 320–323, Doi : 10.1016/j.japb.2018.04.001 , ISSN 2287-884X , S2CID 89705346
- ^ El-Hawary, Redaly S.; ) , sp. nov., a new record of the subfamily Phthiriinae (Bombyliidae, Diptera) from Saudi Arabia", 2014-10-10 , 3872 (4), –392 , doi : 10.11646 / Zotax , PMID 25544092, S2CID .
- ^ Jump up to: а беременный в Андерсон (2001) , с. 33.
- ^ Window et al. (2004) , с. 376
- ^ Jump up to: а беременный Хартманн, Стефани; и Офуализации моли Senita -Cactus -Senita. Nason, John D. ), «Филогенетическое происхождение & Bhattacharya, Debashish ( Lophocereus Cactaceae ) ( 2002 Ajb.89.7.1085 , PMID 21665708
- ^ Андерсон (2001) , с. 537.
- ^ Фенстер, Чарльз Б.; Армбрустер, В. Скотт; Уилсон, Пол; Dudash, Michele R. & Thomson, James D. (2004), «Синдромы опыления и специализация цветов», Ежегодный обзор экологии, эволюции и систематики , 35 : 375–403, doi : 10.1146/annurev.ecolsys.34.011802.132347 , JSTOR 30034121
- ^ McMilllan & Horobin (1995) , p. 499 и далее.
- ^ Флеминг, Теодор Х; Geiselman, Cullen & Kress, W. John (2009), «Эволюция опыления летучих мышей: филогенетическая перспектива», Annals of Botany , 104 (6): 1017–1043, doi : 10.1093/aob/mcp197 , pmc 2766192 , pmid 19789175
- ^ Андерсон (2001) , с. 35–36.
- ^ Гебель, Тед; Уотерс, Майкл Р. и О'Рурк, Деннис Х. (2008), «Позднее плейстоценовое рассеяние современных людей в Америке» (PDF) , Science , 319 (5869): 1497–1502, Bibcode : 2008sci ... 319.1497G , Citeseerx 10.1.1.398.9315 , doi : 10.1126/science.1153569 , pmid 18339930 , S2CID 36149744 , Archived (PDF) из оригинала на 2017-09-22
- ^ Андерсон (2001) , с. 43
- ^ Эндрюс, Дж. Ричард (2003), Введение в классический Nahuatl (пересмотренный изд.), Университет Оклахома Пресс, ISBN 978-0-8061-3452-9 , с. 502 (цитируется на весах: Tenochtitlan )
- ^ Авенни, AF; ( ) Ee Calnek Hartunt & , H. 1988 , , , 162323851
- ^ Barroqueiro, Silvério A., Aztecs: доколумбовая история , Институт учителей Йельского Нью-Хейвена, архивированный с оригинала 2012-05-20 , извлечен 2012-03-07
- ^ Innes (1995) , p. 17
- ^ Роули, Гордон Д. (1997), История сочных растений , Милл -Вэлли, Калифорния: Strawberry Press, OCLC 37830942 , с. 43, цитируется в Anderson 2001 , p. 96
- ^ Андерсон (2001) , с. 456–459
- ^ Linnaeus, Charles (1753), видовые растения Том 1 , Стокгольм: напечатано Лоуренсом Сальвии , извлечен 2012-03-08 , с. 466-470 . Современные названия типов, взятые из синонимов в индексе Anderson 2001 .
- ^ Jump up to: а беременный Гриффит, М. Патрик (2004), «Происхождение важной культуры кактуса, Opuntia ficus-indica (Cactaceae): новые молекулярные доказательства», Американский журнал ботаники , 91 (11): 1915–1921, doi : 10.3732/ajb .91.11.1915 , PMID 21652337 , S2CID 10454390
- ^ Андерсон (2001) , с. 51–54
- ^ Даниэль, Фрэнк Джек (2007-02-19), «Моль, питающаяся кактус, угрожает любимой мексиканской кухне» , Reuters , архивировав с оригинала 2012-09-04 , извлеченного 2012-03-07
- ^ Андерсон (2001) , с. 57–58
- ^ «Pitahaya», Collins English Dictionary , Collins, 2011, архивировал оригинал 2012-06-10 , извлечен 2012-03-13
- ^ Андерсон (2001) , с. 55–59
- ^ Jump up to: а беременный в дюймовый Андерсон (2001) , с. 45–49
- ^ Андерсон (2001) , с. 397
- ^ Zimmerman, Allan D. & Parfitt, Bruce D., «Lophophora williamsii», во Флоре редакционной комитета Флоры Северной Америки (ред.), Флора Северной Америки , архивирована из оригинала 2012-03-11 , извлеченного 2012-03-03-03-03-03 16
- ^ SEEDI, HR; Де Смет, Пенсильвания; Бек, О.; Obsnert, G. & Bruhn, JG (2005), «Доисторическое использование Peoote: алкалоидный анализ и радиоуглеродное датирование археологических образцов Lophophora из Техаса», Journal of Ethnopharmacology , 101 (1–3): 238–242, DOI : 10.1016/. J.Jep.2005.04.022 , PMID 15990261
- ^ Андерсон (2001) , с. 277
- ^ Jump up to: а беременный Bussmann, RW & Sharon, D. (2006), «Традиционное использование лекарственного растения в северном Перу: отслеживание двух тысяч лет культуры исцеления», Journal of Ethnobiology и Ethnomedicine , 2 (1): 47–64, doi : 10.1186/1746 -4269-2-47 , PMC 1637095 , PMID 17090303
- ^ Jump up to: а беременный Keen (1990) , p. 15
- ^ Андерсон (1999) , с. 24–41
- ^ Harlow, Nora & Coate, Barrie D. (2004), Растения и пейзажи для летнего сухого климата , Окленд, Калифорния: Муниципальный утилитный район Ист-Бэй, ISBN 978-0-9753231-0-6
- ^ Рекомендуемый список растений для пустынных ландшафтов (PDF) , Desert Botanical Garden (Phoenix, Аризона), архивировав из оригинала (PDF) 2012-12-03 , извлеченный 2012-03-21
- ^ Шетти, Аноп; Rana, M. & Preetham, S. (2011), «Кактус: лекарственная пища», Журнал пищевой науки и технологии , 49 (5): 530–536, doi : 10.1007/s13197-011-0462-5 , PMC 3550841 , PMID 24082263
- ^ Андерсон (2001) , с. 61–62
- ^ «Культивирование кохинеального в Оахаке», информационный бюллетень Go-Oaxaca , архивировав с оригинала на 2008-06-08 , извлечен 2012-03-21
- ^ Андерсон (2001) , с. 69–72
- ^ Jump up to: а беременный Приложения CITES , CITES , архивируя оригинал 2012-04-14 , извлеченные 2012-04-16
- ^ Jump up to: а беременный Приложения I, II и III , цитируют , архивировали оригинал 2012-10-09 , извлечены 2012-04-16 ; Смотрите "Cactaceae" и связанные сноски
- ^ Андерсон (2001) , с. 73–75
- ^ Андерсон (2001) , с. 77–79
- ^ Маргулис, Джаред Д. (2023). Охотники за кактусами: желание и вымирание в незаконной суккулентной торговле . Миннеаполис: Университет Миннесоты Пресс. ISBN 978-1-4529-6998-5 .
- ^ Андерсон (2001) , с. 79–81
- ^ Jump up to: а беременный в дюймовый Иннес (1995) , с. 22
- ^ Jump up to: а беременный Браун (2001) , с. 87
- ^ Jump up to: а беременный Андерсон (1999) , с. 217
- ^ Хьюитт (1993) , П. 147
- ^ Innes (1995) , p. 23
- ^ Keen (1990) , с. 27–28
- ^ Прикреплено (1994) , с. 140
- ^ Pilbeam (1987) , p. 10
- ^ Jump up to: а беременный в Браун (2001) , с. 88
- ^ Jump up to: а беременный Хьюитт (1993) , с. 151
- ^ Браун (2001) , с. 85
- ^ Jump up to: а беременный Хьюитт (1993) , с. 150
- ^ Pilbeam (1987) , p. 11
- ^ Amos, Robert (2012), «Show Reports: Malvern Show», Альпийский садовник , 80 (1): 80–83
- ^ Sheader, Martin (2012), «Show Reports: Summer Show South», Альпийский садовник , 80 (1): 88–91
- ^ Jump up to: а беременный в Иннес (1995) , с. 28
- ^ Jump up to: а беременный в Браун (2001) , с. 92
- ^ Innes (1995) , p. 27
- ^ Innes (1995) , p. 29
- ^ Ионг, Мён Ил; CHO, Chang-Hui & Lee, Jung-Myung (2009), Производство и разведение кактусов для прививки в Корее , Gyeonggi-Do Services Research & Extension Services, архивные из оригинала на 2013-05-28 , извлечены 2012-03-03- 28
- ^ Innes (1995) , p. 32
- ^ Innes (1995) , с. 31–32
- ^ Браун (2001) , с. 90–91
- ^ «Bipolaris Cactivora (Petr.) Alcorn», виды Fungorum , архивировав с оригинала 2013-05-14 , извлеченные 2012-03-30
- ^ Jump up to: а беременный Прилагается (1994) , с. 152
- ^ Вода, LML; Александр, Мав; Ривас, EB; Харакава, Р.; Sr & Dover, MM (2008), Cactaceae Stort State в , Бразилии в « Potexvirus » 2012-09-09-14 Galleti , 2012-03-30
- ^ Izaguirre-Mayoral, Maria Luisa; Мэрис, Эдглорис; Olivares, Elizabeth & Oropeza, Tamara (1995), «Влияние сезонной засухи и вирусной инфекции кактуса X на метаболизм агавы сисалана , растущие в неотропической саванне» , 46 ( 6): 639–646 , doi : 10.1093/jxb/46.6.639
Библиография
- Андерсон, Эдвард Ф. (2001), Семья кактусов , Пентленд, Орегон: Timber Press, ISBN 978-0-88192-498-5
- Андерсон, Майлз (1999), кактусы и суккуленты: иллюстрированная энциклопедия , Оксфорд: Себастьян Келли, ISBN 978-1-84081-253-4
- Браун, Роджер, «Культивирование кактусов», в Андерсоне (2001) , с. 85–92
- Hecht, Hans (1994), кактусы и суккуленты (P/B Ed.), Нью -Йорк: Стерлинг, ISBN 978-0-8069-0549-5
- Hewitt, Terry (1993), Полная книга кактусов и суккулентов , Лондон: Книги Ковент -Гарден, ISBN 978-1-85605-402-7
- Innes, Clive (1995), «кактусы», в Innes, Clive & Wall, Bill (Eds.), Dacti, Succulents and Bromeliads , Лондон: Cassell для Королевского садоводческого общества, стр. 11–70, ISBN 978-0-304-32076-9
- Keen, Bill (1990), кактусы и суккуленты: шаг за шагом к растущему успеху , Marlborough, Wiltshire: Crowood Press, ISBN 978-1-85223-264-1
- Макмиллан, AJS; Horobin, JF (1995), Рождественские кактусы: род Schlumbergera и ее гибриды (P/B Ed.), Шерборн, Дорсет: Дэвид Хант, ISBN 978-0-9517234-6-3
- Pilbeam, John (1987), кактусы для ценителя , Лондон: Баттсфорд, ISBN 978-0-7134-4861-0
Внешние ссылки
- «Программа Cactaceae» , Rhodes University , 2018-05-14 , получен 2021-12-02
- Наблюдения Cactaceae на iNaturalist
Семейства цветущих растений ( APG IV ) |
|---|