Цикл мочевины (также известный как орнитиновый цикл ) — это цикл биохимических реакций, в ходе которых образуется мочевина (NH 2 ) 2 CO из аммиака (NH 3 ). Животные, использующие этот цикл, главным образом земноводные и млекопитающие, называются уреотелическими .
Цикл мочевины преобразует высокотоксичный аммиак в мочевину для выведения. [ 1 ] Этот цикл был первым метаболическим циклом, открытым Гансом Кребсом и Куртом Хензелейтом в 1932 году. [ 2 ] [ 3 ] [ 4 ] за пять лет до открытия цикла ТСА . Более подробно цикл мочевины был описан позже Ратнером и Коэном. Цикл мочевины протекает преимущественно в печени и, в меньшей степени, в почках .
Катаболизм аминокислот приводит к образованию отходов аммиака. Всем животным нужен способ выведения этого продукта. Большинство водных организмов , или аммонотелических организмов, выделяют аммиак, не преобразуя его. [ 1 ] Организмы, которые не могут легко и безопасно удалить азот в виде аммиака, преобразуют его в менее токсичное вещество, такое как мочевина , посредством цикла мочевины, который происходит главным образом в печени. Мочевина, вырабатываемая печенью, затем попадает в кровоток , откуда попадает в почки и в конечном итоге выводится с мочой . Цикл мочевины важен для этих организмов, потому что, если азот или аммиак не выводятся из организма, это может быть очень вредным. [ 5 ] У видов, включая птиц и большинство насекомых , аммиак превращается в мочевую кислоту или ее уратную соль, которая выводится из организма в твердой форме . Кроме того, цикл мочевины потребляет углекислый газ кислых отходов путем объединения его с основным аммиаком, помогая поддерживать нейтральный pH.
Весь процесс преобразует две аминогруппы, одну из NH. + 4 и один от аспартата и атом углерода от HCO − 3 , к относительно нетоксичному продукту выделения мочевины . [ 6 ] Это происходит за счет четырех «высокоэнергетических» фосфатных связей (3 АТФ гидролизуются до 2 АДФ и одной АМФ ). Преобразование аммиака в мочевину происходит в пять основных этапов. Первый необходим для поступления аммиака в цикл, а следующие четыре являются частью самого цикла. Для входа в цикл аммиак преобразуется в карбамоилфосфат . Цикл мочевины состоит из четырех ферментативных реакций: одной митохондриальной и трех цитозольных . [ 1 ] [ 7 ] Для этого используются 6 ферментов. [ 6 ] [ 7 ] [ 8 ]
Перед началом цикла мочевины аммиак превращается в карбамоилфосфат. Реакция катализируется карбамоилфосфатсинтетазой I и требует участия двух молекул АТФ . [ 1 ] Затем карбамоилфосфат поступает в цикл мочевины.
Аргинин расщепляется аргиназой с образованием мочевины и орнитина. Затем орнитин транспортируется обратно в митохондрии, чтобы снова начать цикл мочевины. [ 1 ] [ 7 ]
Поскольку фумарат получают удалением NH 3 из аспартата (посредством реакций 3 и 4), а PP i + H 2 O → 2 Pi , уравнение можно упростить следующим образом:
Обратите внимание, что реакции, связанные с циклом мочевины, также вызывают образование 2 НАДН , поэтому в целом реакция выделяет немного больше энергии, чем потребляет. НАДН производится двумя способами:
Одна молекула НАДН вырабатывается ферментом глутаматдегидрогеназой при превращении глутамата в аммоний и α-кетоглутарат . Глутамат является нетоксичным носителем аминогрупп. Это обеспечивает ион аммония, используемый при первоначальном синтезе карбамоилфосфата.
Два произведенных НАДН могут обеспечить энергию для образования 5 АТФ (цитозольный НАДН обеспечивает 2,5 АТФ с помощью малатно-аспартатного челнока в клетках печени человека), что является чистым производством двух высокоэнергетических фосфатных связей для цикла мочевины. Однако, если в цитозоле происходит глюконеогенез , последний восстанавливающий эквивалент используется для изменения стадии GAPDH вместо генерации АТФ.
Судьба оксалоацетата заключается либо в производстве аспартата посредством трансаминирования, либо в превращении в фосфоенолпируват , который является субстратом глюконеогенеза .
Как указано выше, многие позвоночные используют цикл мочевины для создания мочевины из аммония, чтобы аммоний не повреждал организм. Хотя это полезно, существуют и другие эффекты цикла мочевины. Например: потребление двух АТФ, производство мочевины, образование Н. + , объединение ОХО − 3 и NH + 4 в формы, где он может быть регенерирован, и, наконец, потребление НХ + 4 . [ 9 ]
Синтез карбамоилфосфата и цикл мочевины зависят от присутствия N -ацетилглутаминовой кислоты (NAcGlu), которая аллостерически активирует CPS1 . NAcGlu является облигатным активатором карбамоилфосфатсинтетазы. [ 10 ] Синтез NAcGlu с помощью N -ацетилглутаматсинтазы (NAGS) стимулируется как Arg, аллостерическим стимулятором NAGS, так и Glu, продуктом реакций трансаминирования и одним из субстратов NAGS, уровень обоих из которых повышается при свободных аминокислот повышении уровня . Таким образом, Glu не только является субстратом для NAGS, но также служит активатором цикла мочевины.
Остальные ферменты цикла контролируются концентрациями их субстратов. Таким образом, унаследованный дефицит ферментов цикла, отличных от ARG1, не приводит к значительному снижению выработки мочевины (если какой-либо фермент цикла полностью отсутствует, смерть наступает вскоре после рождения). Скорее, субстрат дефицитного фермента накапливается, увеличивая скорость реакции дефицита до нормальной.
Однако аномальное накопление субстрата не обходится без затрат. Концентрации субстрата повышаются на всем пути обратного цикла до NH. + 4 , что приводит к гипераммониемии (повышение [ NH + 4 ] П ).
Хотя основная причина NH + 4 токсичность до конца не выяснена, высокая [ NH + 4 ] оказывает огромную нагрузку на NH + 4- система очистки, особенно в головном мозге (симптомы недостаточности ферментов цикла мочевины включают умственную отсталость и вялость ). Эта система очистки включает GLUD1 и GLUL , которые уменьшают пулы 2-оксоглутарата (2OG) и Glu. Мозг наиболее чувствителен к истощению этих пулов. Истощение 2OG снижает уровень TCAC , тогда как Glu является одновременно нейромедиатором и предшественником GABA , другого нейромедиатора. [1] (стр.734)
Цикл мочевины и цикл лимонной кислоты являются независимыми циклами, но связаны между собой. Один из атомов азота в цикле мочевины получается в результате переаминирования оксалоацетата в аспартат. [ 11 ] Фумарат, образующийся на третьем этапе, также является промежуточным продуктом цикла лимонной кислоты и возвращается в этот цикл. [ 11 ]
Нарушения цикла мочевины встречаются редко и затрагивают примерно одного из 35 000 человек в Соединенных Штатах . [ 12 ] Могут возникнуть генетические дефекты ферментов, участвующих в цикле, которые обычно проявляются в течение нескольких дней после рождения. [ 5 ] Недавно родившийся ребенок обычно испытывает различные приступы рвоты и периоды летаргии . [ 5 ] В конечном итоге младенец может впасть в кому и получить повреждение головного мозга . [ 5 ] Новорожденные с UCD подвергаются гораздо более высокому риску осложнений или смерти из-за несвоевременных скрининговых тестов и неправильно диагностированных случаев. Наиболее частым ошибочным диагнозом является неонатальный сепсис . Признаки UCD могут присутствовать в течение первых 2–3 дней жизни, но существующий метод получения подтверждения по результатам анализов может занять слишком много времени. [ 13 ] Это потенциально может вызвать такие осложнения, как кома или смерть. [ 13 ]
Нарушения цикла мочевины также могут быть диагностированы у взрослых, и симптомы могут включать делирия эпизоды , летаргию и симптомы, подобные симптомам инсульта . [ 14 ] Помимо этих симптомов, если цикл мочевины в печени начинает нарушаться , у пациента может развиться цирроз печени . [ 15 ] Это также может привести к саркопении (потере мышечной массы). [ 15 ] Мутации приводят к дефициту различных ферментов и транспортеров, участвующих в цикле мочевины, и вызывают нарушения цикла мочевины. [ 1 ] Если люди с дефектом любого из шести ферментов, используемых в цикле, потребляют аминокислоты сверх того, что необходимо для минимальной суточной потребности, то образующийся аммиак не сможет преобразоваться в мочевину. У этих людей может наблюдаться гипераммониемия или накопление промежуточного продукта цикла.
Все дефекты цикла мочевины, за исключением дефицита ОТС, наследуются по аутосомно-рецессивному типу. Дефицит ОТК наследуется как Х-сцепленное рецессивное заболевание, хотя у некоторых женщин могут проявляться симптомы. Большинство нарушений цикла мочевины связаны с гипераммониемией , однако аргининемия и некоторые формы аргининоянтарной ацидурии не сопровождаются повышенным содержанием аммиака.
Arc.Ask3.Ru Номер скриншота №: d2daa57b97556fe95948b099c6b6fc2f__1721903280 URL1:https://arc.ask3.ru/arc/aa/d2/2f/d2daa57b97556fe95948b099c6b6fc2f.html Заголовок, (Title) документа по адресу, URL1: Urea cycle - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)
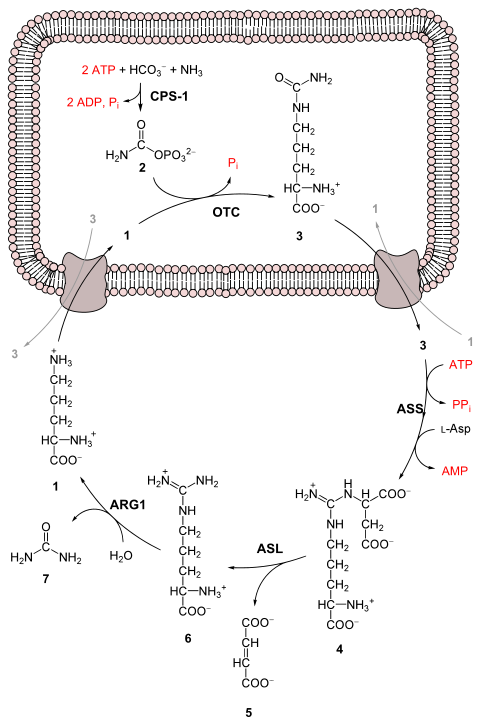
 Цикл мочевины.
Цикл мочевины. Цветной цикл мочевины.
Цветной цикл мочевины.





