Мох
| Мох | |
|---|---|
 | |
| Комки мха на земле и у основания деревьев в Национальном лесу Аллегейни , Пенсильвания, США. | |
| Научная классификация | |
| Королевство: | Растения |
| Клэйд : | Эмбриофиты |
| Клэйд : | Сетафита |
| Разделение: | Мохообразные Шимп. в строгом смысле |
| Классы [2] | |
| Синонимы | |
| |
Мхи — это небольшие бессосудистые без цветков растения из таксономического отдела Bryophyta ( / b r aɪ ˈ ɒ f ə t ə / , [3] / ˌ б р аɪ . ə ˈ f aɪ t ə / ) в строгом смысле . Bryophyta ( sensu lato , Schimp . 1879). [4] ) также может относиться к родительской группе мохообразных , в которую входят печеночники , мхи и роголистники . [5] Мхи обычно образуют плотные зеленые комки или коврики, часто во влажных или затененных местах. Отдельные растения обычно состоят из простых листьев толщиной всего в одну клетку, прикрепленных к стеблю , который может быть разветвленным или неразветвленным и играет лишь ограниченную роль в проведении воды и питательных веществ. Хотя у некоторых видов имеются проводящие ткани, они, как правило, плохо развиты и структурно отличаются от аналогичных тканей, встречающихся у сосудистых растений . [6] У мхов нет семян , и после оплодотворения развиваются спорофиты с неразветвленными стеблями, увенчанными одиночными капсулами, содержащими споры . Обычно они имеют высоту 0,2–10 см (0,1–3,9 дюйма), хотя некоторые виды намного крупнее. Даусония , самый высокий мох в мире, может вырасти до 50 см (20 дюймов) в высоту. Существует около 12 000 видов. [2]
Мхи обычно путают с печеночниками, роголистниками и лишайниками . [7] Хотя многие мхи часто называют бессосудистыми растениями , они имеют развитую сосудистую систему. [8] [9] Как и у печеночников и роголистников, гаплоидное гаметофитное поколение мхов является доминирующей фазой жизненного цикла . Это контрастирует с картиной у всех сосудистых растений ( семенных растений и птеридофитов ), где диплоидное доминирует поколение спорофитов. Лишайники могут внешне напоминать мхи и иногда имеют общие названия, включающие слово «мох» (например, « оленьий мох » или « исландский мох »), но они представляют собой грибковый симбиоз и не связаны со мхами. [7] : 3
Основное коммерческое значение мхов заключается в том, что они являются основным компонентом торфа (в основном рода Sphagnum ), хотя они также используются в декоративных целях, например, в садах и в цветочном торговле. Традиционное использование мхов включало в себя изоляцию и способность поглощать жидкости, в 20 раз превышающие их вес. Мох является ключевым родом и способствует восстановлению среды обитания и лесовосстановлению . [10]
Физические характеристики
[ редактировать ]Описание
[ редактировать ]
С ботанической точки зрения мхи относятся к бессосудистым растениям отдела наземных растений Bryophyta. Обычно это небольшие (несколько сантиметров в высоту) травянистые (недревесные) растения, которые поглощают воду и питательные вещества в основном через листья и собирают углекислый газ и солнечный свет для создания пищи путем фотосинтеза . [11] [12] За исключением древней группы Takakiopsida , ни один из известных мхов не образует микоризу . [13] но мохообразные грибы широко распространены во мхах и других мохообразных, где они обитают как сапротрофы, паразиты, возбудители болезней и мутуалисты, некоторые из них - эндофиты . [14] Мхи отличаются от сосудистых растений отсутствием водоносных ксилемных трахеид или сосудов . Как и у печеночников и роголистников , является генерация гаплоидных гаметофитов доминирующей фазой жизненного цикла . Это контрастирует с картиной у всех сосудистых растений ( семенных растений и птеридофитов ), где диплоидное поколение спорофитов доминирует . Мхи размножаются спорами , а не семенами , и не имеют цветков.

Гаметофиты мха имеют стебли, которые могут быть простыми или разветвленными, вертикальными (акрокарпий) или стелющимися (плеврокарпий). Ранние дивергентные классы Takakiopsida, Sphagnopsida, Andreaeopsida и Andreaeobriopsida либо лишены устьиц , либо имеют псевдоустьица, не образующие пор. В остальных классах устьица терялись более 60 раз. [15] Их листья простые, обычно состоят из одного слоя клеток без внутренних воздушных пространств, часто с более толстыми жилками (нервами). Нерв может выходить за край кончика листа и называется экскуррентным. Кончик листовой пластинки может быть удлинен в виде волоска, состоящего из бесцветных клеток. Они кажутся белыми на фоне темно-зеленых листьев. Край листа может быть гладким или иметь зубцы. Может существовать особый тип клеток, определяющих край листа, отличающийся по форме и/или цвету от других клеток листа. [16] У мха есть нитевидные ризоиды , которые прикрепляют его к субстрату, что можно сравнить с корневыми волосками , а не с более прочными корневыми структурами сперматофитов . [17] Мхи не поглощают воду и питательные вещества из субстрата через ризоиды. [ нужна ссылка ] Их можно отличить от печеночников ( Marchantiophyta или Hepaticae ) по многоклеточным ризоидам. Спороносные капсулы или спорангии мхов располагаются поодиночке на длинных неразветвленных стеблях, что отличает их от полиспорангиофитов , к которым относятся все сосудистые растения. Спорофиты, производящие споры (т.е. диплоидное многоклеточное поколение), недолговечны и обычно способны к фотосинтезу, но запасы воды и большинства или всех питательных веществ зависят от гаметофита. [18] Кроме того, у большинства мхов спороносная капсула увеличивается и созревает после удлинения ее стебля, тогда как у печеночников капсула увеличивается и созревает до удлинения ее стебля. [12] Другие различия не универсальны для всех мхов и всех печеночников, но наличие четко дифференцированного стебля с простыми по форме бессосудистыми листьями, расположенными не в три ряда, все указывает на то, что растение является мхом. [ нужна ссылка ]
Жизненный цикл
[ редактировать ]Сосудистые растения имеют два набора хромосом в вегетативных клетках и называются диплоидными , то есть каждая хромосома имеет партнера, который содержит ту же или сходную генетическую информацию. Напротив, мхи и другие мохообразные имеют только один набор хромосом и поэтому являются гаплоидными (т. е. каждая хромосома существует в клетке в уникальной копии). В жизненном цикле мхов есть период, когда у них действительно имеется двойной набор парных хромосом, но это происходит только на стадии спорофита .
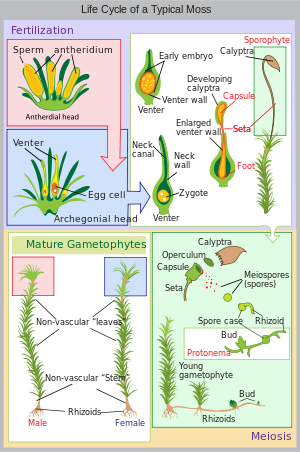
Жизненный цикл мха начинается с гаплоидной споры , которая прорастает, образуя протонему ( мн. protonemata), которая представляет собой либо массу нитевидных нитей, либо таллоид (плоский и таллоидный). Массивные протонемы мха обычно выглядят как тонкий зеленый войлок и могут расти на влажной почве, коре деревьев, камнях, бетоне или практически на любой другой достаточно устойчивой поверхности. Это переходный этап в жизни мха, но из протонемы вырастает гаметофор ( «носитель гамет»), структурно дифференцированный на стебли и листья. На одном коврике протонемат может образоваться несколько побегов гаметофора, в результате чего образуется комок мха.
Из кончиков стеблей или ветвей гаметофора развиваются половые органы мхов. Женские органы известны как архегонии ( sing.archegonium . ) и защищены группой видоизмененных листьев, известных как перихетум (множественное число, перихета) Архегонии представляют собой небольшие скопления клеток в форме колбы с открытой шейкой (брюшной частью), по которой плавают мужские сперматозоиды. Мужские органы известны как антеридии ( sing. antheridium ) и окружены видоизмененными листьями, называемыми перигониями ( мн. perigonia). Окружающие листья у некоторых мхов образуют чашу для разбрызгивания, позволяющую сперме, содержащейся в чашке, разбрызгиваться на соседние стебли падающими каплями воды. [19]
Рост кончика гаметофора нарушается грибным хитином . [20] [21] [22] Галотто и др. , 2020 применили хитооктаозу и обнаружили, что кончики обнаруживают это производное хитина и реагируют на него, изменяя экспрессию генов . [20] [21] [22] Они пришли к выводу, что эта защитная реакция, вероятно, у самого недавнего общего предка мохообразных сохранилась и трахеофитов . [20] Орр и др. , 2020 обнаружили, что микротрубочки растущих кончиковых клеток структурно подобны F-актину и служат той же цели. [21]
Мхи могут быть как двудомными (ср. двудомные у семенных растений), так и однодомными (ср. однодомные ). У двудомных мхов мужские и женские половые органы расположены на разных растениях-гаметофитах. У одноцветных (также называемых автоикными) мхов оба растения растут на одном и том же растении. В присутствии воды сперматозоиды из антеридиев доплывают до архегониев и происходит оплодотворение , приводящее к образованию диплоидного спорофита. Сперма мхов двужгутиковая, т. е. у них есть два жгутика, которые помогают двигаться. Поскольку сперматозоид должен доплыть до архегония, оплодотворение не может произойти без воды. Некоторые виды (например, Mnium Hornum или несколько видов Polytrichum ) держат свои антеридии в так называемых «чашечках», чашеобразных структурах на кончиках побегов, которые перемещают сперму на несколько дециметров при попадании капель воды, увеличивая расстояние оплодотворения. [19]
После оплодотворения незрелый спорофит выходит из вентральной части архегониала. требуется несколько месяцев спорофита Для созревания . Тело спорофита состоит из длинной ножки, называемой щетинкой, и капсулы, увенчанной шляпкой, называемой жаберной крышкой . Капсула и жаберная крышка, в свою очередь, покрыты гаплоидной калиптрой, которая представляет собой остатки вентральной архегониальной части. Калиптра обычно отпадает, когда капсула созревает. Внутри капсулы клетки, производящие споры, подвергаются мейозу с образованием гаплоидных спор, после чего цикл может начаться снова. Устье капсулы обычно окружено зубцами, называемыми перистомами. У некоторых мхов он может отсутствовать. [ нужна ссылка ]
Большинство мхов разносят споры ветром. У рода Sphagnum споры выбрасываются на высоту примерно 10–20 см (4–8 дюймов) от земли с помощью сжатого воздуха, содержащегося в капсулах; споры ускоряются примерно в 36 000 раз быстрее ускорения Земли . гравитационного [23] [24]

Недавно было обнаружено, что микроартроподы, такие как коллемболы и клещи , могут производить удобрение мха. [25] и что этот процесс опосредован ароматами, исходящими от мха. мужского и женского пола излучает разные и сложные летучие органические ароматы. огненный мох Например, [26] Женские растения выделяют больше соединений, чем мужские. Было обнаружено, что коллемболы предпочитают женские растения, и одно исследование показало, что коллемболы усиливают оплодотворение мха, что предполагает наличие опосредованных запахом отношений, аналогичных отношениям растение-опылитель, обнаруженным у многих семенных растений. [26] Вид вонючего мха Splachnum sphaericum далее развивает опыление насекомыми, привлекая мух к своим спорангиям сильным запахом падали и обеспечивая сильный визуальный сигнал в виде набухших красных воротников под каждой споровой капсулой. Мухи, привлеченные мхом, переносят его споры в свежий навоз травоядных животных, который является излюбленной средой обитания видов этого рода. [27]
У многих мхов, например, Ulota phyllantha , на листьях или ветвях образуются зеленые вегетативные структуры, называемые геммами , которые могут отрываться и образовывать новые растения без необходимости проходить цикл оплодотворения. Это способ бесполого размножения , и генетически идентичные особи могут привести к образованию клональных популяций.
Карликовые самцы
[ редактировать ]Самцы моховых карликов (также известные как наннандри или филлодиоидность ) происходят из разносимых ветром мужских спор , которые оседают и прорастают на женских побегах, где их рост ограничивается несколькими миллиметрами. У некоторых видов карликовость детерминирована генетически: все мужские споры становятся карликовыми. [28] Чаще всего это обусловлено окружающей средой: мужские споры, попадающие на самку, становятся карликовыми, а те, которые попадают в другое место, развиваются в крупных самцов размером с самку. [28] [29] [30] [31] В последнем случае карликовые самцы, пересаженные от самок на другой субстрат, развиваются в крупные побеги, что позволяет предположить, что самки выделяют вещество, тормозящее рост прорастающих самцов и, возможно, ускоряющее также наступление у них полового созревания. [30] [31] Природа такого вещества неизвестна, но фитогормон ауксин. возможно, в этом участвует [28]
Ожидается, что рост самцов как карликов на самке повысит эффективность оплодотворения за счет минимизации расстояния между мужскими и женскими репродуктивными органами. Соответственно, было замечено, что частота оплодотворения положительно связана с присутствием карликовых самцов у некоторых филлодиоидных видов. [32] [33]
Самцы-карлики встречаются в нескольких неродственных линиях. [33] [34] и может быть более распространенным, чем считалось ранее. [33] Например, по оценкам, от четверти до половины всех двудомных плеврокарпов имеют карликовых самцов. [33]
восстановление ДНК
[ редактировать ]Мох Physcomitrium patens использовался в качестве модельного организма для изучения того, как растения восстанавливают повреждения своей ДНК, особенно механизм восстановления, известный как гомологичная рекомбинация . Если растение не может восстановить повреждения ДНК, например двухцепочечные разрывы , в своих соматических клетках , клетки могут потерять нормальные функции или погибнуть. Если это произойдет во время мейоза (часть полового размножения), они могут стать бесплодными. Геном P. patens был секвенирован, что позволило идентифицировать несколько генов, участвующих в репарации ДНК. [35] Мутанты P. patens , дефектные на ключевых этапах гомологичной рекомбинации, были использованы для выяснения того, как функционирует механизм репарации в растениях. Например, исследование мутантов P. patens , дефектных по Rp RAD51, гену, который кодирует белок, лежащий в основе реакции рекомбинационной репарации, показало, что гомологичная рекомбинация необходима для восстановления двухцепочечных разрывов ДНК у этого растения. [36] Аналогичным образом, исследования мутантов, дефектных по Ppmre11 или Pprad50 (которые кодируют ключевые белки комплекса MRN , основного сенсора двухцепочечных разрывов ДНК), показали, что эти гены необходимы для восстановления повреждений ДНК, а также для нормального роста и развития. [37]
Классификация
[ редактировать ]Совсем недавно мхи были сгруппированы с печеночниками и роголистниками в отдел Bryophyta ( мохообразные , или Bryophyta sensu lato ). [5] [38] [39] [40] [41] [42] [43] [44] [45] [ чрезмерное цитирование ] Сам отдел мохообразных включает три (бывших) отдела: Bryophyta (мхи), Marchantiophyta (печеночники) и Anthocerotophyta (роголистники); было предложено перевести эти последние подразделения в классы Bryopsida, Marchantiopsida и Anthocerotopsida соответственно. [5] Сейчас мхи и печеночники считаются принадлежащими к кладе Setaphyta . [39] [46] [47]
Мхи (Bryophyta sensu stricto) делятся на восемь классов:
отдел мохообразных
|
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Современная филогения и состав мохообразных. [2] [48] | ||||||||||||||||||||||||||||||||||||||||||||||||||||

Шесть из восьми классов содержат только один или два рода каждый. Polytrichopsida включает 23 рода, а Bryopsida включает большую часть разнообразия мхов, причем к этому классу принадлежит более 95% видов мхов.
Sphagnopsida, торфяные мхи, включают два современных рода Ambuchanania и Sphagnum , а также ископаемые таксоны. Сфагнум – разнообразный, широко распространенный и экономически важный вид. Эти крупные мхи образуют обширные кислые болота на торфяных болотах. В листьях сфагнума крупные мертвые клетки чередуются с живыми фотосинтезирующими клетками. Мертвые клетки помогают хранить воду. Помимо этого признака, уникальное ветвление, слоевищное (плоское и расширенное) протонема и взрыворазрывающийся спорангий выделяют его среди других мхов.
Andreaeopsida и Andreaeobriopsida отличаются двурядными (два ряда клеток) ризоидами, многорядной (многорядность клеток) протонемой и спорангием, расщепляющимся по продольным линиям. У большинства мхов есть капсулы, открывающиеся сверху.
У Polytrichopsida листья с набором параллельных пластинок, створок клеток, содержащих хлоропласты, которые выглядят как плавники на радиаторе. Они осуществляют фотосинтез и могут помочь сохранить влагу, частично закрывая поверхности газообмена. Polytrichopsida отличаются от других мхов и другими деталями своего развития и анатомии, а также могут становиться крупнее большинства других мхов, например, Polytrichum commune образует подушки высотой до 40 см (16 дюймов). Самый высокий наземный мох, представитель Polytrichidae, вероятно, Dawsonia superba , произрастающий в Новой Зеландии и других частях Австралазии .
Геологическая история
[ редактировать ]
Летопись окаменелостей мхов скудна из-за их мягкостенной и хрупкой природы. Однозначные окаменелости мхов были обнаружены еще в пермском периоде Антарктиды и России, а также были сделаны предположения о каменноугольных мхах. [49] Далее утверждалось, что трубчатые окаменелости из силура представляют собой мацерированные останки моховых калиптр . [50] Мхи также развиваются в 2–3 раза медленнее, чем папоротники, голосеменные и покрытосеменные растения . [51]
Недавние исследования показывают, что древний мох может объяснить, почему произошли ордовикские ледниковые периоды. Когда предки современного мха начали распространяться по суше 470 миллионов лет назад, они поглощали CO 2 из атмосферы и извлекали минералы, выделяя органические кислоты, которые растворяли камни, на которых они росли. Эти химически измененные породы, в свою очередь, вступили в реакцию с атмосферным CO 2 и образовали новые карбонатные породы в океане за счет выветривания ионов кальция и магния из силикатных пород. Выветренные породы также выпустили значительное количество фосфора и железа, которые оказались в океанах, где вызвали массовое цветение водорослей, что привело к захоронению органического углерода и извлечению большего количества углекислого газа из атмосферы. Мелкие организмы, питающиеся питательными веществами, создали большие территории без кислорода, что вызвало массовое вымирание морских видов, а уровень CO 2 упал во всем мире, что привело к образованию ледяных шапок на полюсах. [52] [53]
Экология
[ редактировать ]среда обитания
[ редактировать ] Плотные колонии мха в прохладном прибрежном лесу
Плотные колонии мха в прохладном прибрежном лесу Прохладный моховой лес на большой высоте/широте; лесная подстилка покрыта мхом под хвойными деревьями
Прохладный моховой лес на большой высоте/широте; лесная подстилка покрыта мхом под хвойными деревьями Мох колонизирует базальтовый поток в Исландии.
Мох колонизирует базальтовый поток в Исландии. Мох, растущий вдоль выходов и источников в недавно отложенных базальтовых породах, Исландия.
Мох, растущий вдоль выходов и источников в недавно отложенных базальтовых породах, Исландия. Мох, растущий вдоль ручья из карстового источника ; Травертин откладывается из воды ручья, и мох зарастает его, образуя хребет с ручьем наверху.
Мох, растущий вдоль ручья из карстового источника ; Травертин откладывается из воды ручья, и мох зарастает его, образуя хребет с ручьем наверху. Мох со спорофитами на кирпиче
Мох со спорофитами на кирпиче Молодые спорофиты мха обыкновенного Tortula muralis (винтовой мох настенный)
Молодые спорофиты мха обыкновенного Tortula muralis (винтовой мох настенный) Подпорная стенка, покрытая мхом
Подпорная стенка, покрытая мхом Небольшой комок мха под хвойным деревом (тенистое, обычно сухое место)
Небольшой комок мха под хвойным деревом (тенистое, обычно сухое место) Мох на бетонной стене
Мох на бетонной стене Мох (Bryophyta) на лесной подстилке в Брокен-Боу, Оклахома.
Мох (Bryophyta) на лесной подстилке в Брокен-Боу, Оклахома.











мха Гаметофиты автотрофны и требуют солнечного света для фотосинтеза . [54] Тенеустойчивость варьируется в зависимости от вида, как и у высших растений. В большинстве районов мхи растут в основном во влажных, затененных местах, таких как лесные массивы и по берегам ручьев, но они могут расти где угодно в прохладном, влажном и пасмурном климате, а некоторые виды адаптированы к солнечным, сезонно засушливым местам, таким как альпийские горы. скалы или стабилизированные песчаные дюны.
Выбор субстрата также зависит от вида. Виды мхов можно классифицировать как растущие на: камнях, обнаженной минеральной почве, нарушенных почвах, кислой почве, известковой почве, выходах скал и участках с брызгами водопадов, берегах ручьев, затененной перегнойной почве, поваленных бревнах, сгоревших пнях, основаниях стволов деревьев, верхних стволах деревьев. и ветках деревьев или в болотах . часто специфичны в отношении видов деревьев, на которых они растут, например, предпочитают хвойные породы широколиственным деревьям , дубы ольхе Виды мха, растущие на деревьях или под ними , или наоборот. [12] Хотя мхи часто растут на деревьях как эпифиты , они никогда не паразитируют на дереве.
Мхи также встречаются в трещинах между брусчаткой на сырых городских улицах и на крышах. Некоторые виды, приспособленные к нарушенным солнечным местам, хорошо приспособлены к городским условиям и обычно встречаются в городах. Примерами могут служить Rhytidiadelphus sqarrosus , садовый сорняк в районах Ванкувера и Сиэтла; Bryum argenteum , космополитический тротуарный мох, и Ceratodon purpureus , красный кровельный мох, еще один космополитный вид. Некоторые виды полностью водные, например Fontinalis antipyretica , водяной мох обыкновенный; а другие, такие как Sphagnum, обитают в болотах, болотах и очень медленно текущих водных путях. [12] Такие водные или полуводные мхи могут значительно превышать нормальный диапазон длины, наблюдаемый у наземных мхов. распространены отдельные растения длиной 20–30 см (8–12 дюймов) и более Sphagnum Например, у видов . Но даже водным видам мхов и других мохообразных необходимо, чтобы их зрелые капсулы подвергались воздействию воздуха за счет удлинения щетинок или сезонного понижения уровня воды, чтобы иметь возможность размножаться. [55]
Где бы они ни встречались, мхам требуется жидкая вода, по крайней мере, часть года для завершения оплодотворения. Многие мхи могут пережить высыхание , иногда в течение нескольких месяцев, возвращаясь к жизни в течение нескольких часов после регидратации. [54]
Принято считать, что в Северном полушарии северная сторона деревьев и скал обычно имеет более обильный рост мха, чем другие стороны. [56] Предполагается, что причина в том, что солнечный свет на южной стороне вызывает сухость окружающей среды. Обратное было бы справедливо в Южном полушарии . Некоторые натуралисты считают, что мхи растут на более влажной стороне деревьев и камней. [11] В некоторых случаях, например, в солнечном климате умеренных северных широт, это будет затененная северная сторона дерева или камня. На крутых склонах это может быть подъем. Для мхов, растущих на ветвях деревьев, это обычно верхняя сторона ветки на горизонтально растущих участках или возле промежности. В прохладном, влажном и пасмурном климате все стороны стволов деревьев и камней могут быть одинаково влажными, что способствует росту мха. Каждому виду мха требуется определенное количество влаги и солнечного света, поэтому он растет на определенных участках одного и того же дерева или камня.
Некоторые мхи растут под водой или полностью затоплены. Многие предпочитают хорошо дренированные места. Есть мхи, которые преимущественно растут на камнях и стволах деревьев различного химического состава. [57]
Связь с цианобактериями
[ редактировать ]В бореальных лесах некоторые виды мхов играют важную роль в обеспечении экосистемы азотом благодаря их связи с азотфиксирующими цианобактериями . Цианобактерии колонизируют мох и получают убежище в обмен на фиксированный азот. Мох выделяет фиксированный азот вместе с другими питательными веществами в почву «при таких нарушениях, как высыхание, повторное увлажнение и пожары», делая его доступным для всей экосистемы. [58]
Выращивание
[ редактировать ]

Мох часто считают сорняком на газонах, но его намеренно поощряют выращивать в соответствии с эстетическими принципами, примером которых является японское садоводство . В старых храмовых садах мох может покрыть лесной пейзаж. Считается, что мох добавляет ощущение спокойствия, возраста и тишины в садовую сцену. Мох также используется в бонсай , чтобы покрыть почву и усилить впечатление возраста. [59] Правила выращивания широко не установлены. Сборы мха нередко начинают с образцов, пересаженных из дикой природы в водоудерживающий мешок. Некоторые виды мха чрезвычайно сложно содержать вдали от их естественного места обитания из-за их уникальных требований к сочетанию света, влажности, химического состава субстрата, укрытия от ветра и т. д.
Выращивание мха из спор еще менее контролируется. Споры мха падают под постоянным дождем на открытые поверхности; те поверхности, которые благоприятны для определенного вида мха, обычно заселяются этим мхом в течение нескольких лет под воздействием ветра и дождя. Пористые и влагоудерживающие материалы, такие как кирпич , дерево и некоторые грубые бетонные смеси, благоприятствуют мху. Поверхности также можно подготовить с помощью кислых веществ, включая пахту , йогурт , мочу и слегка протертые смеси образцов мха, воды и верескового компоста .
На прохладном, влажном и пасмурном северо-западе Тихого океана мху иногда позволяют расти естественным путем в виде мохового газона , который практически не нужно стричь, удобрять или поливать. В этом случае трава считается сорняком. [60] Ландшафтные дизайнеры в районе Сиэтла иногда собирают валуны и поваленные бревна, из которых растет мох, для установки в садах и ландшафтах. Лесные сады во многих частях мира могут иметь ковер из натуральных мхов. [54] Заповедник Блодел на острове Бейнбридж, штат Вашингтон, известен своим мховым садом. Сад мха был создан путем удаления кустарникового подлеска и травянистого почвопокровного покрова, прореживания деревьев и обеспечения естественного зарастания мхов. [61]
Зеленые крыши и стены
[ редактировать ]
Мхи иногда используются в зеленых крышах . Преимущества мхов перед высшими растениями в зеленых крышах заключаются в снижении весовых нагрузок, повышенном водопоглощении, отсутствии необходимости в удобрениях, высокой засухоустойчивости. Поскольку у мхов нет настоящих корней, им требуется меньше посадочной среды, чем высшим растениям с обширной корневой системой. При правильном выборе видов для местного климата мхи на зеленых крышах не требуют орошения и не требуют особого ухода. [62] Мхи также используются на зеленых стенах .
Моссери
[ редактировать ]Преходящее увлечение сбором мха в конце 19 века привело к созданию мховых ферм во многих британских и американских садах. Мох обычно строится из решетчатого дерева с плоской крышей, открытой на северную сторону (сохраняет тень). В щели между деревянными планками были установлены образцы мха. Затем весь мох регулярно увлажняли для поддержания роста.
Акваскейпинг
[ редактировать ]В акваскейпинге используется множество водных мхов. Они лучше всего растут при низких уровнях питательных веществ, света и тепла и довольно легко размножаются. Они помогают поддерживать химический состав воды, подходящий для аквариумных рыб. [63] Они растут медленнее, чем многие аквариумные растения, и довольно выносливы. [64]
Замедление роста
[ редактировать ]Мох может стать неприятным сорняком в контейнерных питомниках и теплицах. [65] Энергичный рост мха может препятствовать появлению всходов и проникновению воды и удобрений к корням растений.
Рост мха можно остановить несколькими способами:
- Уменьшение доступности воды через дренаж .
- Увеличение количества прямых солнечных лучей.
- Увеличивается количество и ресурсы, доступные для конкурентоспособных растений, таких как травы .
- Повышение рН почвы внесением извести .
- Интенсивное движение или вручную рыхление мха граблями.
- Применение химикатов, таких как сульфат железа (например, на газонах) или отбеливатель (например, на твердых поверхностях).
- В контейнерных питомниках грубые минеральные материалы, такие как песок, гравий и каменная крошка, используются в качестве быстро дренируемой подкормки в контейнерах для растений, чтобы препятствовать росту мха.
Применение средств, содержащих сульфат железа или сульфат железа-аммония, убьет мох; эти ингредиенты обычно содержатся в коммерческих продуктах и удобрениях для борьбы с мхом . Сера и железо являются важными питательными веществами для некоторых конкурирующих растений, таких как травы. Уничтожение мха не предотвратит возобновление роста, если не будут изменены условия, благоприятные для его роста. [66]
Использование
[ редактировать ]
Традиционный
[ редактировать ]Доиндустриальные общества использовали мхи, растущие на их территориях.
Саамы , североамериканские племена и другие приполярные народы использовали мхи в качестве подстилки. [11] [54] Мхи также использовались в качестве утеплителя как для жилищ, так и для одежды. Традиционно сушеный мох использовался в некоторых странах Северной Европы и России в качестве изолятора между бревнами в бревенчатых хижинах , а племена северо-востока США и юго-востока Канады использовали мох для заполнения щелей в деревянных длинных домах. [54] Приполярные и альпийские народы использовали мх для утепления сапог и рукавиц. У Эци Ледяного человека были ботинки, покрытые мхом. [54]
Способность сушеных мхов поглощать жидкости сделала их практичными как в медицинских, так и в кулинарных целях. Племенные народы Северной Америки использовали мх для изготовления подгузников, повязок на раны и всасывания менструальных выделений. [54] Племена северо-запада Тихого океана в США и Канаде использовали мох для очистки лосося перед его сушкой и упаковывали влажный мох в ямы-печи для пропаривания луковиц камас . Корзины для хранения продуктов и корзины для варки также были набиты мхом. [54]
Недавние исследования останков неандертальцев, обнаруженных в Эль-Сидроне, предоставили доказательства того, что их рацион состоял в основном из кедровых орехов, мха и грибов. Этому противоречат данные из других европейских стран, которые указывают на более плотоядную диету. [67]
В Финляндии торфяной мох использовался для приготовления хлеба во время голода . [68]
Коммерческий
[ редактировать ]
Существует значительный рынок мхов, собранных в дикой природе. Нетронутый мох в основном используется в цветочной торговле и для украшения дома. Разлагающийся мох рода Sphagnum также является основным компонентом торфа , который «добывают» для использования в качестве топлива , в качестве добавки к почве в садоводстве и для курения солода при производстве шотландского виски .
Мох сфагнум , обычно виды S. cristatum и S. subnitens , собирают во время роста и высушивают для использования в питомниках и садоводстве в качестве среды для выращивания растений.
Некоторые мхи -сфагнумы могут поглощать воды в 20 раз больше собственного веса. [69] В Первую мировую войну мох - сфагнум использовался в качестве повязок для оказания первой помощи солдатам, так как этот мох, как говорят, впитывает жидкость в три раза быстрее, чем хлопок, лучше удерживает жидкость, лучше распределяет жидкость равномерно по себе, а также более прохладный, мягкий и меньше раздражать. [69] Также утверждается, что он обладает антибактериальными свойствами. [70] Коренные американцы были одним из народов, использовавших сфагнум для изготовления подгузников и менструальных прокладок , что до сих пор делается в Канаде . [71]
В сельской местности Fontinalis Великобритании antipyretica традиционно использовалась для тушения пожаров, поскольку в значительных количествах ее можно было найти в медленно текущих реках, а мох удерживал большие объемы воды, что помогало тушить пламя. Это историческое использование отражено в его специфическом латинско - греческом названии, приблизительное значение которого — «против огня».
В Мексике мох используют в качестве рождественского украшения.
Physcomitrium patens все чаще используется в биотехнологии . Яркими примерами являются идентификация генов мха.с последствиями для улучшения урожая или здоровья человека [72] и безопасное производство сложных биофармацевтических препаратов в моховом биореакторе , разработанном Ральфом Рески и его коллегами. [73]
В Лондоне установили несколько сооружений под названием «Городские деревья»: стены, покрытые мхом, каждая из которых, как утверждается, обладает «способностью очищать воздух, как 275 обычных деревьев», поглощая оксиды азота и другие виды загрязнений воздуха и производя кислород. [74]
Ссылки
[ редактировать ]- ^ Хуберс, М.; Керп, Х. (2012). «Самые старые известные мхи обнаружены в слоях Миссисипи (поздневизейского периода) в Германии». Геология . 40 (8): 755–58. Бибкод : 2012Geo....40..755H . дои : 10.1130/G33122.1 .
- ^ Jump up to: а б с Гоффине, Бернар; Уильям Р. Бак (2004). «Систематика мохообразных (мхов): от молекул к пересмотренной классификации». Монографии по систематической ботанике . Молекулярная систематика мохообразных. Том. 98. Издательство Ботанического сада Миссури. стр. 205–239. ISBN 978-1-930723-38-2 .
- ^ «Мохообразные» . Словарь Merriam-Webster.com .
- ^ Шимпер, WP (1879). «Мохообразные». В Зиттеле, К.А. (ред.). Справочник по палеонтологии . Том 2. Р. Ольденбург.
- ^ Jump up to: а б с де Соуза, Филипе; и др. (2019). «Филогения ядерных белков поддерживает монофилию трех групп мохообразных (Bryophyta Schimp.)» . Новый фитолог . 222 (1): 565–75. дои : 10.1111/nph.15587 . hdl : 1983/0b471d7e-ce54-4681-b791-1da305d9e53b . ПМИД 30411803 . S2CID 53240320 .
- ^ Лигроне, Р.; Дакетт, Дж. Г.; Рензалья, К.С. (2000). «Проводящие ткани и филетические взаимоотношения мохообразных» . Philos Trans R Soc Lond B Biol Sci . 355 (1398): 795–813. дои : 10.1098/rstb.2000.0616 . ПМЦ 1692789 . ПМИД 10905610 .
- ^ Jump up to: а б Лишайники Северной Америки, Ирвин М. Бродо, Сильвия Дюран Шарнофф, ISBN 978-0-300-08249-4 , 2001 г.
- ^ Белл, Н. Э. и Хивонен, Дж. (2010). «Филогения класса мхов Polytrichopsida (BRYOPHYTA): структура родового уровня и несоответствующие генные деревья» . Молекулярная филогенетика и эволюция . 55 (2): 381–398. дои : 10.1016/j.ympev.2010.02.004 . ПМИД 20152915 .
- ^ Бодрибб, Ти Джей; и др. (2020). «У широко распространенного мха обнаружена расширенная сосудистая функция». Природные растения . 6 (3): 273–279. дои : 10.1038/s41477-020-0602-x . ПМИД 32170283 . S2CID 212641738 .
- ^ Рошфор, Линия (2000). «Сфагнум: ключевой род в восстановлении среды обитания» . Бриолог . 103 (3): 503–508. doi : 10.1639/0007-2745(2000)103[0503:SAKGIH]2.0.CO;2 . ISSN 0007-2745 . JSTOR 3244138 . S2CID 55699307 .
- ^ Jump up to: а б с Мэтьюз, Дэниел (1994). Каскадно-Олимпийский Природоведение . Портленд, Орегон: общество Одюбона Портлендское / Raven Editions. ISBN 978-0-9620782-0-0 .
- ^ Jump up to: а б с д Пожар и Маккиннон (1994). Растения северо-западного побережья Тихого океана . Ванкувер, Британская Колумбия: Издательство Lone Pine. ISBN 978-1-55105-040-9 .
- ^ Наличие трех микоризных генов у общего предка наземных растений предполагает ключевую роль микоризы в колонизации суши растениями.
- ^ Использование секвенирования 454 для изучения разнообразия, специфичности хозяина и тканевой специфичности грибов рода Galerina, связанных с четырьмя бореальными мхами.
- ^ При более чем 60 независимых потерях устьица становятся расходным материалом для мхов.
- ^ Атертон, Ян; Бозанке, Сэм; Лоули, Марк (2010). Мхи и печеночники Британии и Ирландии — полевой справочник . Британское бриологическое общество. п. 848. ИСБН 9780956131010 .
- ^ Уотсон, Э. Вернон (1981). Британские мхи и печеночники (3-е изд.). Издательство Кембриджского университета. п. 519. ИСБН 052129472X .
- ^ Будке, Джессика М; Бернар, Эрнест С; Грей, Деннис Дж; Хуттунен, Санна; Пьечулла, Биргит; Триджано, Роберт Н. (2018). «Введение в специальный выпуск о мохообразных». Критические обзоры по наукам о растениях . 37 (2–3): 102–112. дои : 10.1080/07352689.2018.1482396 .
- ^ Jump up to: а б ван дер Вельде, М.; Во время, HJ; ван де Занде, Л.; Бейлсма, Р. (2001). «Репродуктивная биология Polytrichum formosum: клональная структура и отцовство, выявленное с помощью микросателлитов». Молекулярная экология . 10 (10): 2423–2434. Бибкод : 2001MolEc..10.2423V . дои : 10.1046/j.0962-1083.2001.01385.x . ПМИД 11742546 . S2CID 19716812 .
- ^ Jump up to: а б с Дело, Пьер-Марк; Шорнак, Себастьян (19 февраля 2021 г.). «Эволюция растений, обусловленная взаимодействием с симбиотическими и патогенными микробами» (PDF) . Наука . 371 (6531). Американская ассоциация развития науки (AAAS): 1–10. дои : 10.1126/science.aba6605 . ISSN 0036-8075 . ПМИД 33602828 . S2CID 231955632 .
- ^ Jump up to: а б с Бибо, Джеффри П.; Галотто, Джулия; Ву, Мин; Тюзель, Эркан; Видали, Луис (6 апреля 2021 г.). «Количественная клеточная биология роста кончиков мха» . Молекулярная биология растений . 107 (4–5). Спрингер : 227–244. дои : 10.1007/s11103-021-01147-7 . ISSN 0167-4412 . ПМЦ 8492783 . ПМИД 33825083 .
- ^ Jump up to: а б Сунь, Гуйлин; Бай, Шэнлун; Гуань, Яньлун; Ван, Шуанхуа; Ван, Ця; Лю, Ян; Лю, Хуан; Гоффине, Бернар; Чжоу, Юн; Паолетти, Матье; Ху, Сянъян; Хаас, Фабиан Б.; Фернандес-Посо, Ноэ; Чырт, Алия; Солнце, Ханг; Ренсинг, Стефан А.; Хуан, Цзиньлин (31 июля 2020 г.). «Связаны ли геномные области грибного происхождения с антагонизмом по отношению к грибам во мхах?» . Новый фитолог . 228 (4). Фонд новых фитологов ( Уайли ): 1169–1175. дои : 10.1111/nph.16776 . ISSN 0028-646X . PMID 32578878 . S2CID 220047618 .
- ^ Йохан Л. ван Леувен (23 июля 2010 г.). «Запущен при 36 000 г ». Наука . 329 (5990): 395–6. дои : 10.1126/science.1193047 . ПМИД 20651138 . S2CID 206527957 .
- ^ Дуайт К. Уитакер и Джоан Эдвардс (23 июля 2010 г.). « Мох сфагнум разносит споры с помощью вихревых колец». Наука . 329 (5990): 406. Бибкод : 2010Sci...329..406W . дои : 10.1126/science.1190179 . ПМИД 20651145 . S2CID 206526774 .
- ^ Кронберг, Н.; Начева Р.; Хедлунд, К. (2006). «Микроартроподы опосредуют передачу спермы у мхов». Наука . 313 (5791): 1255. doi : 10.1126/science.1128707 . ПМИД 16946062 . S2CID 11555211 .
- ^ Jump up to: а б Розенстиль, Теннесси; Шортлидж, EE; Мельниченко А.Н.; Панков, Дж. Ф.; Эппли, С.М. (2012). «Летучие соединения с разным полом влияют на оплодотворение мха, опосредованное микроартроподами». Природа . 489 (7416): 431–433. Бибкод : 2012Natur.489..431R . дои : 10.1038/nature11330 . ПМИД 22810584 . S2CID 4419337 .
- ^ Вайзи, младший (1890). «К морфологии спорофита Splachnum luteum ». Анналы ботаники . 1 : 1–8. doi : 10.1093/oxfordjournals.aob.a090623 .
- ^ Jump up to: а б с Уне, Кодзи (1985). «Половой диморфизм японских видов Macromitrium Brid. (Musci: Orthotrichaceae)» . Журнал ботанической лаборатории Хаттори, посвященный бриологии и лихенологии . 59 : 487–513.
- ^ Блэксток, TH (1987). «Мужские гаметофоры Leucobryum glaucum (Hedw.) Ångstr. и L. juniperoideum (Brid.) C. Muell. В двух лесах Уэльса». Журнал бриологии . 14 (3): 535–541. Бибкод : 1987JBryo..14..535B . дои : 10.1179/jbr.1987.14.3.535 .
- ^ Jump up to: а б Лавленд, Хью Фрэнк (1956). Половой диморфизм у мхов рода Dicranum Hedw. (Диссертация) . Мичиганский университет.
- ^ Jump up to: а б Уоллес, Миннесота (1970). Морфология развития и половой диморфизм у Homalothecium megaptilum (Sull.) Robins. (Диссертация) . Университет штата Вашингтон.
- ^ Сагмо Солли, международный мастер; Сёдерстрем, Ларс; Баккен, Сольвейг; Флатберг, Хьелл Ивар; Педерсен, Борд (1998). «Исследование плодовитости Dicranum majus в двух популяциях с контрастным производством спорофитов». Журнал бриологии . 22 (1): 3–8. дои : 10.1179/jbr.2000.22.1.3 . S2CID 85349694 .
- ^ Jump up to: а б с д Хеденас, Ларс; Бисанг, Ирен (2011). «Незамеченные самцы-карлики во мхах — уникальные среди зеленых наземных растений». Перспективы экологии, эволюции и систематики растений . 13 (2): 121–135. дои : 10.1016/j.ppees.2011.03.001 .
- ^ Рамзи, Хелен П.; Берри, ГК (1982). «Определение пола у мохообразных». Журнал Ботанической лаборатории Хаттори . 52 : 255–274.
- ^ Ренсинг, Южная Каролина; Ланг, Д; Циммер, AD; Терри, А.; Саламов А; Шапиро, Х.; Нисияма, Т.; и др. (январь 2008 г.). «Геном Physcomitrella раскрывает эволюционное понимание завоевания суши растениями» (PDF) . Наука . 319 (5859): 64–9. Бибкод : 2008Sci...319...64R . дои : 10.1126/science.1150646 . hdl : 11858/00-001M-0000-0012-3787-A . ПМИД 18079367 . S2CID 11115152 .
- ^ Маркманн-Мулиш У., Венделер Э., Зобелл О., Швин Г., Штайнбисс Х.Х., Рейсс Б. (октябрь 2007 г.). «Дифференциальные требования к RAD51 при развитии Physcomitrella patens и Arabidopsis thaliana и восстановлении повреждений ДНК» . Растительная клетка . 19 (10): 3080–9. дои : 10.1105/tpc.107.054049 . ПМК 2174717 . ПМИД 17921313 .
- ^ Камисуги Й., Шефер Д.Г., Козак Дж., Шарло Ф., Врилинк Н., Хола М., Анжелис К.Дж., Куминг А.С., Ноге Ф (апрель 2012 г.). «MRE11 и RAD50, но не NBS1, необходимы для нацеливания на гены мха Physcomitrella patens» . Нуклеиновые кислоты Рез . 40 (8): 3496–510. дои : 10.1093/nar/gkr1272 . ПМЦ 3333855 . ПМИД 22210882 .
- ^ Кокс, Саймон Дж.; и др. (2014). «Противоречивые филогении ранних наземных растений вызваны предвзятостью состава синонимических замен» . Систематическая биология . 63 (2): 272–279. дои : 10.1093/sysbio/syt109 . ПМЦ 3926305 . ПМИД 24399481 .
- ^ Jump up to: а б Путтик, Марк Н.; и др. (2018). «Взаимоотношения наземных растений и природа предкового эмбриофита». Современная биология . 28 (5): 733–745.e2. дои : 10.1016/j.cub.2018.01.063 . hdl : 1983/ad32d4da-6cb3-4ed6-add2-2415f81b46da . ПМИД 29456145 . S2CID 3269165 .
- ^ Либенс-Мак, Джеймс Х.; и др. (2019). «Тысяча растительных транскриптомов и филогеномика зеленых растений» . Природа . 574 (7780): 679–685. дои : 10.1038/s41586-019-1693-2 . ПМК 6872490 . PMID 31645766 .
- ^ Чжан, Цзянь; и др. (2020). «Геном роголистника и ранняя эволюция наземных растений» . Природные растения . 6 (2): 107–118. дои : 10.1038/s41477-019-0588-4 . ПМК 7027989 . ПМИД 32042158 .
- ^ Харрис, Броган Дж.; и др. (2020). «Филогеномные доказательства монофилии мохообразных и редуктивной эволюции устьиц». Современная биология . 30 (11): P2201–2012.E2. дои : 10.1016/j.cub.2020.03.048 . hdl : 1983/fbf3f371-8085-4e76-9342-e3b326e69edd . ПМИД 32302587 . S2CID 215798377 .
- ^ Ли, Фэй-Вэй; и др. (2020). «Геномы Anthoceros проливают свет на происхождение наземных растений и уникальную биологию роголистников» . Природные растения . 6 (3): 259–272. дои : 10.1038/s41477-020-0618-2 . hdl : 10261/234303 . ПМЦ 8075897 . ПМИД 32170292 .
- ^ Соуза, Филипе; и др. (2020). «Филогения наземных растений хлоропластов: анализ с использованием более подходящих моделей неоднородного состава деревьев и участков» . Границы в науке о растениях . 11 : 1062. дои : 10.3389/fpls.2020.01062 . ПМЦ 7373204 . ПМИД 32760416 .
- ^ Су, Данян; и др. (2021). «Крупномасштабный филогеномный анализ выявляет монофилию мохообразных и неопротерозойское происхождение наземных растений» . Молекулярная биология и эволюция . 38 (8): 3332–3344. дои : 10.1093/molbev/msab106 . ПМЦ 8321542 . ПМИД 33871608 .
- ^ Соуза, Филипе; и др. (2020). «Митохондриальная филогения наземных растений демонстрирует поддержку Setaphyta в рамках моделей гетерогенного замещения по составу» . ПерДж . 8 (4): е8995. дои : 10.7717/peerj.8995 . ПМК 7194085 . ПМИД 32377448 .
- ^ Кокс, Саймон Дж. (2018). «Молекулярная филогенетика наземных растений: обзор с комментариями по оценке несоответствия между филогениями». Критические обзоры по наукам о растениях . 37 (2–3): 113–127. Бибкод : 2018CRvPS..37..113C . дои : 10.1080/07352689.2018.1482443 . hdl : 10400.1/14557 . S2CID 92198979 .
- ^ Бак, Уильям Р. и Бернар Гоффине. (2000). «Морфология и классификация мхов», страницы 71–123 в книге А. Джонатана Шоу и Бернарда Гоффине (ред.), Bryophyte Biology . (Кембридж: Издательство Кембриджского университета). ISBN 0-521-66097-1 .
- ^ Томас, бакалавр (1972). «Вероятный мох из нижнего карбона леса Дин, Глостершир». Анналы ботаники . 36 (1): 155–161. doi : 10.1093/oxfordjournals.aob.a084568 . ISSN 1095-8290 . JSTOR 42752024 .
- ^ Коднер, РБ; Грэм, Ле (2001). «Высокотемпературные кислотно-гидролизованные останки Polytrichum (Musci, Polytrichaceae) напоминают загадочные силурийско-девонские трубчатые микроокаменелости». Американский журнал ботаники . 88 (3): 462–466. дои : 10.2307/2657111 . JSTOR 2657111 . ПМИД 11250824 .
- ^ Стенойен, Гонконг (2008). «Медленная молекулярная эволюция генов 18S рДНК, rbcL и nad5 мхов по сравнению с высшими растениями» . Журнал эволюционной биологии . 21 (2): 566–571. дои : 10.1111/j.1420-9101.2007.01479.x . ПМИД 18205784 .
- ^ «Первые наземные растения погрузили Землю в ледниковый период» . Newscientist.com. Архивировано из оригинала 23 сентября 2013 года . Проверено 11 сентября 2013 г.
- ^ «Первые растения вызвали ледниковые периоды, показывают новые исследования» . Sciencedaily.com. 1 февраля 2012 года. Архивировано из оригинала 3 октября 2013 года . Проверено 11 сентября 2013 г.
- ^ Jump up to: а б с д и ж г час Киммерер, Робин Уолл (2003). Сбор мха . Корваллис, Орегон: Издательство Университета штата Орегон. ISBN 978-0-87071-499-3 .
- ^ Давление отбора на эволюцию устьиц - Интернет-библиотека Wiley
- ^ Порли, Рон; Ходжеттс, Ник (2005). Мхи и печеночники . Лондон: Коллинз. стр. 80–81. ISBN 978-0-00-220212-1 .
- ^ Флетчер, Майкл (2006). Справочник производителей мха (3-е изд.). Чтение Беркшира: SevenTy Press. ISBN 0-9517176-0-Х .
- ^ Руск, Кэтрин; Джонс, Дэйви Л.; ДеЛука, Томас Х. (1 января 2013 г.). «Мхово-цианобактериальные ассоциации как биогенные источники азота в бореальных лесных экосистемах» . Границы микробиологии . 4 : 150. дои : 10.3389/fmicb.2013.00150 . ISSN 1664-302X . ПМЦ 3683619 . ПМИД 23785359 .
- ^ Чан, Питер (1993). Мастер-класс по бонсай . Нью-Йорк: ISBN Sterling Publishing Co. 978-0-8069-6763-9 .
- ^ Смит, Салли В. (1998). Решение проблем заката в западном саду . Менло-Парк, Калифорния: Книги о закате. ISBN 978-0-376-06132-4 .
- ^ «Резерв Блоделя» . Архивировано из оригинала 16 апреля 2011 года . Проверено 24 апреля 2011 г.
- ^ «Сад на Крыше» . Архивировано из оригинала 24 апреля 2011 года . Проверено 22 мая 2011 г.
- ^ «Руководство по хранению и выращиванию водного мха» . Акваскейп Любовь . 12 апреля 2016 г.
- ^ «Мхи» . www.aquasabi.com .
- ^ Хаглунд, Уильям А.; Рассел и Холланд (лето 1981 г.). «Борьба с мхом в саженцах хвойных деревьев, выращенных в контейнерах» (PDF) . Записки плантатора деревьев (USFS) . 32 (3): 27–29. Архивировано (PDF) из оригинала 19 июля 2011 года . Проверено 24 апреля 2011 г.
- ^ Стив Уитчер; Мастер-садовник (1996). «Борьба с мхом на газонах» . Садоводство в Западном Вашингтоне . Университет штата Вашингтон. Архивировано из оригинала 5 февраля 2007 года . Проверено 10 февраля 2007 г.
- ^ Вейрих, Лаура С.; Дюшен, Себастьян; Субрие, Жюльен; Арриола, Луис; Ламас, Бастьен; Брин, Джеймс; Моррис, Алан Г.; Альт, Курт В.; Карамелли, Дэвид; Дрезели, Вейт; Фаррелл, Милли; Фаррер, Эндрю Г.; Франкен, Майкл; Галли, Невилл; Хаак, Вольфганг; Харди, Карен; Харвати, Катерина; Держись, Петра; Холмс, Эдвард К.; Кайдонис, Джон; Лалуэса-Фокс, Карлес; де ла Расилья, Марко; Росас, Антонио; Семаль, Патрик; Солтысяк, Аркадиуш; Таунсенд, Грант; Усай, Донателла; Валь, Иоахим; Хьюсон, Дэниел Х.; и др. (2017). «Поведение, диета и болезни неандертальцев, выведенные на основе древней ДНК зубного камня» (PDF) . Природа . 544 (7650): 357–361. Бибкод : 2017Natur.544..357W . дои : 10.1038/nature21674 . hdl : 10261/152016 . ПМИД 28273061 . S2CID 4457717 .
- ^ Энгман, Макс; Д. Г. Кирби (1989). Финляндия: люди, нация, государство . К. Херст и Ко. с. 45. ISBN 0-253-32067-4 .
- ^ Jump up to: а б Подземный мир растений, сфагнум и вода, Австралийский ботанический сад. Архивировано 17 февраля 2014 г. в Wayback Machine.
- ^ Сталхейм, Т.; Балланс, С.; Кристенсен, Бельгия; Гранум, ЧП (1 марта 2009 г.). «Сфагнан — пектиноподобный полимер, выделенный из мха сфагнум, может подавлять рост некоторых типичных бактерий, вызывающих порчу пищевых продуктов и пищевых отравлений, путем снижения pH». Журнал прикладной микробиологии . 106 (3): 967–976. дои : 10.1111/j.1365-2672.2008.04057.x . ISSN 1365-2672 . ПМИД 19187129 . S2CID 1545021 .
- ^ Хотсон, JW (1921). «Сфагнум использовался в качестве хирургической повязки в Германии во время мировой войны (завершение)». Бриолог . 24 (6): 89–96. doi : 10.1639/0007-2745(1921)24[89:suasdi]2.0.co;2 . JSTOR 3237483 .
- ^ Ральф Рески и Вольфганг Франк (2005): мха ( Physcomitrella patens ) Функциональная геномика – открытие генов и разработка инструментов, имеющих последствия для сельскохозяйственных культур и здоровья человека. Брифинги по функциональной геномике и протеомике 4, 48–57.
- ^ Декер, Эл.; Рески, Р. (2007). «Мховые биореакторы для производства улучшенных биофармацевтических препаратов». Современное мнение в области биотехнологии . 18 (5): 393–398. дои : 10.1016/j.copbio.2007.07.012 . ПМИД 17869503 .
- ^ Лэндон, Алекс (7 января 2020 г.). «Городские деревья: в Лондоне появились новые искусственные деревья, которые пожирают загрязнения» . Тайный Лондон . Проверено 9 февраля 2020 г.
Дальнейшее чтение
[ редактировать ]- Киммерер, Робин Уолл (2003). Сбор мха: естественная и культурная история мхов . Издательство Университета штата Орегон. ISBN 0-87071-499-6 .
Внешние ссылки
[ редактировать ]- Информация, схемы и фотографии
- Справочник выращивателя мха (PDF-файл 2,3, 9 МБ)
- Британское бриологическое общество
- Картинная галерея мхов
- Мир мхов - Акварельные картины мха Роберта Мумы.
Классификация Archaeplastida или Plantae sl |
|---|