В -клетка
| B лимфоцит клетка | |
|---|---|
 Анимация B -клеток | |
| Подробности | |
| Предшественник | Гематопоэтические стволовые клетки |
| Система | Иммунная система |
| Идентификаторы | |
| латинский | лимфоцит б |
| Сетка | D001402 |
| FMA | 62869 |
| Анатомические термины микроанатомии | |
-клетки , также известные как В -лимфоциты , являются типом лейкоцитов подтипа лимфоцитов В . [ 1 ] Они функционируют в гуморального иммунитета компоненте адаптивной иммунной системы . [ 1 ] В-клетки продуцируют молекулы антител , которые могут быть секретированы или вставлены в плазматическую мембрану, где они служат частью B-клеточных рецепторов . [ 2 ] Когда B-клетка или В-клетки памяти активируется антигеном, он пролиферирует и дифференцируется в эффекторную клетку, секретирующую антитело, известную как плазмаблу или плазматическую клетку. [ 2 ] Кроме того, В-клетки представляют антигены (они также классифицируются как профессиональные антиген-презентативные клетки, APC ) и секретируют цитокины . [ 1 ] У млекопитающих В -клетки созревают в костном мозге , который лежит в ядре большинства костей . [ 3 ] У птиц В -клетки созревают в бурсе Фабрициуса , лимфоидного органа, где они были впервые обнаружены Чангом и Гликом, [ 4 ] Вот почему B обозначает бурсу , а не костного мозга , как обычно считают.
В -клетки, в отличие от двух других классов лимфоцитов, Т -клеток и природных клеток -киллеров , экспрессируют рецепторы В -клеток (BCR) на их клеточной мембране . [ 1 ] BCR позволяют B -клетку связываться с иностранным антигеном , против которого он будет инициировать ответ антител. [ 1 ] Рецепторы В -клеток чрезвычайно специфичны, причем все BCR на В -клетках распознают тот же эпитоп . [ 5 ]
Разработка
[ редактировать ]
В -клетки развиваются из гематопоэтических стволовых клеток (HSC), которые происходят из костного мозга . [ 6 ] [ 7 ] HSCS сначала дифференцируется в мультипотентные клетки -предшественника (MPP), затем общие клетки лимфоидных предшественников (CLP). [ 7 ] Отсюда их развитие в B -клетки происходит на нескольких стадиях (показано на изображении справа), каждая из которых отмечена различными паттернами экспрессии генов и с цепью L и L -цепи расположением локусов генов , последняя из -за B -клеток, подвергающихся V (D) J Рекомбинация по мере развития. [ 8 ]
В -клетки подвергаются двум типам отбора при развитии в костном мозге, чтобы обеспечить правильное развитие, оба с участием рецепторов В -клеток (BCR) на поверхности клетки. Положительный отбор происходит через антиген-независимую передачу сигналов с участием как Pre-BCR, так и BCR. [ 9 ] [ 10 ] Если эти рецепторы не связываются с их лигандом , В -клетки не получают надлежащие сигналы и не перестают развиваться. [ 9 ] [ 10 ] Отрицательный отбор происходит через связывание самоантигена с BCR; Если BCR может сильно связываться с самоантигеном, то B-клетка подвергается одной из четырех существ: клональная делеция , редактирование рецепторов , анергию или невежество (В-клеток игнорирует сигнал и продолжает развитие). [ 10 ] Этот отрицательный процесс отбора приводит к состоянию центральной толерантности , в котором зрелые В -клетки не связывают самогенов, присутствующие в костном мозге. [ 8 ]
Для завершения развития незрелые В -клетки мигрируют из костного мозга в селезенку в виде переходных В -клеток , проходя через две переходные стадии: T1 и T2. [ 11 ] На протяжении всей миграции в селезенку и после въезда селезенки они считаются T1 -B -клетками. [ 12 ] Внутри селезенки T1 B -клетки переходят на T2 -B -клетки. [ 12 ] B -клетки T2 дифференцируются в фолликулярные (FO) B -клетки или B -клетки маргинальной зоны (MZ) в зависимости от сигналов, полученных через BCR и других рецепторов. [ 13 ] После дифференциации они теперь считаются зрелыми В -клетками или наивными В -клетками. [ 12 ]
Активация
[ редактировать ]
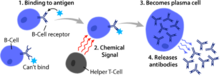
Активация В -клеток происходит во вторичных лимфоидных органах (SLO), таких как селезенка и лимфатические узлы . [ 1 ] После того, как В -клетки созревают в костном мозге, они мигрируют через кровь в SLO, которые получают постоянное поставку антигена посредством циркулирующей лимфы . [ 14 ] При SLO активация B -клеток начинается, когда B -клетка связывается с антигеном через его BCR. [ 15 ] Хотя события, происходящие сразу после активации, еще не были полностью определены, считается, что В -клетки активируются в соответствии с моделью кинетической сегрегации [ Цитация необходима ] Первоначально определяется в Т -лимфоцитах. Эта модель обозначает, что до стимуляции антигена рецепторы диффундируют через мембрану, вступающую в контакт с LCK и CD45 на одинаковой частоте, что делает чистое равновесие фосфорилирования и нефосфорилирования. Только когда клетка вступает в контакт с антигеном, представляющей клетку, более крупный CD45 смещен из -за близкого расстояния между двумя мембранами. Это обеспечивает чистое фосфорилирование BCR и инициация пути передачи сигнала [ Цитация необходима ] Полем Из трех подмножества B-клеток FO B-клетки преимущественно подвергаются Т-клеткам-зависимой активации, в то время как B-клетки MZ и B1-B-клетки преимущественно подвергаются активации, независимой от Т-клеток. [ 16 ]
Активация В -клеток усиливается благодаря активности CD21 , поверхностного рецептора в комплексе с поверхностными белками CD19 и CD81 (все три вместе известны как комплекс корецептора B -клеток). [ 17 ] Когда BCR связывает антиген, помеченный фрагментом белка комплемента C3, CD21 связывает фрагмент C3, совместно лигирует со связанным BCR, а сигналы трансдуцируются через CD19 и CD81, чтобы снизить порог активации клетки. [ 18 ]
Т-клеточная активация
[ редактировать ]Антигены, которые активируют В-клетки с помощью Т-клеток, известны как антигены, зависящие от Т-клеток (ТД) и включают в себя иностранные белки. [ 1 ] Они названы как таковые, потому что они не могут вызвать гуморальный ответ у организмов, в которых отсутствуют Т -клетки. [ 1 ] В B-клеточные реакции на эти антигены занимают несколько дней, хотя генерируемые антитела имеют более высокую аффинность и являются более функционально универсальными, чем те, которые генерируются из-за независимой от Т-клеток активации. [ 1 ]
После того, как BCR связывает антиген TD, антиген попадает в B-клетку с помощью рецепторного опосредованного эндоцитоза , деградируется и представляется Т-клеткам в виде пептидных кусочков в комплексе с молекулами MHC-II на клеточной мембране. [ 19 ] T Herper (T H Клетки ) , обычно фолликулярные T-Helper (T FH ) клетки распознают и связывают эти комплексы MHC-II-пептид через их рецептор Т-клеток (TCR) . [ 20 ] После связывания TCR-MHC-II-пептида Т-клетки экспрессируют поверхностный белок CD40L , а также цитокины, такие как IL-4 и IL-21 . [ 20 ] CD40L служит необходимым костимуляторным фактором для активации В-клеток путем связывания рецептора B-клеточной поверхности CD40 , который способствует пролиферации В-клеток , переключению класса иммуноглобулина и соматической гипермутации , а также поддерживает рост и дифференцировку Т-клеток. [ 1 ] Цитокины, полученные из Т-клеток, связанные с рецепторами цитокинов В-клеток, также способствуют пролиферации В-клеток, переключению класса иммуноглобулина и соматической гипермутации, а также направляющей дифференциации. [ 20 ] После того, как В -клетки получают эти сигналы, они считаются активированными. [ 20 ]

После активации В-клетки участвуют в двухступенчатом процессе дифференцировки, который дает как кратковременные плазмабласты для немедленной защиты, так и долгоживущих плазматических клеток и В-клеток памяти для постоянной защиты. [ 16 ] Первый шаг, известный как экстрафолликулярный отклик, происходит наружу лимфоидные фолликулы, но все еще в SLO. [ 16 ] Во время этой стадии активированные В -клетки пролиферируются, могут подвергаться переключению класса иммуноглобулина и дифференцируются на плазмабласты, которые производят ранние слабые антитела в основном из класса IgM. [ 21 ]

Второй этап состоит из активированных В -клеток, входящих в лимфоидный фолликул и образуя зародышевый центр (ГХ) , который представляет собой специализированное микроокружение, где В -клетки подвергаются обширной пролиферации, переключению класса иммуноглобулина и созреванием аффинности, направленным соматической гиперметацией. [ 22 ] Эти процессы облегчаются клетками T FH в GC и генерируют как B-клетки памяти с высокоаффинной памяти, так и долгоживущие плазматические клетки. [ 16 ] Результирующие плазматические клетки выделяют большое количество антител и либо остаются в SLO, либо, более предпочтительно, мигрируют в костный мозг. [ 22 ]
Т-клеточная независимая активация
[ редактировать ]Антигены, которые активируют В-клетки без помощи Т-клеток, известны как Т-клеточные (Ti) антигены [ 1 ] и включают иностранные полисахариды и неметилированную ДНК CPG. [ 16 ] Они названы как таковые, потому что они способны вызывать гуморальный ответ у организмов, в которых отсутствуют Т -клетки. [ 1 ] В B-клеточный ответ на эти антигены быстрый, хотя генерируемые антитела имеют тенденцию иметь более низкую аффинность и являются менее функционально универсальными, чем те, которые генерируются в зависимости от активации T-клеток. [ 1 ]
Как и в случае с антигенами TD, В-клетки, активируемые антигенами Ti, нуждаются в дополнительных сигналах для полной активации, но вместо того, чтобы получать их от Т-клеток, они обеспечиваются либо распознаванием, так и связыванием общей микробной компоненты с рецепторами, подобными Toll, (TLR) или Обширное сшивание BCRS с повторными эпитопами на бактериальной клетке. [ 1 ] В-клетки, активируемые антигенами Ti, продолжают пролиферировать внешние лимфоидные фолликулы, но все еще в SLO (GC не образуются), возможно, подвергаются переключению класса иммуноглобулина и дифференцируются в недолговечь Некоторые популяции долгоживущих плазматических клеток. [ 23 ]
Активация В -клеток памяти
[ редактировать ]Активация В -клеток памяти начинается с обнаружения и связывания их мишенивого антигена, который разделяется их родительской B -клеткой. [ 24 ] Некоторые В-клетки памяти могут быть активированы без помощи Т-клеток, например, определенные вирус-специфические В-клетки памяти, но другие нуждаются в помощи Т-клеток. [ 25 ] При связывании антигена В-клетка памяти принимает антиген посредством рецепторного эндоцитоза, разлагает его и представляет его Т-клеткам в виде пептидных кусочков в комплексе с молекулами MHC-II на клеточной мембране. [ 24 ] Клетки T-Helper (T H ), обычно фолликулярные T-Helper (T FH ) клетки памяти, которые были получены из Т-клеток, активируемых с одним и тем же антигеном, распознают и связывают эти комплексы MHC-II-пептидов через их TCR. [ 24 ] После связывания TCR-MHC-II-пептида и реле других сигналов из ячейки памяти T FH- клетки активируются и дифференцируются либо на плазмаблуаслы, так и плазматические клетки посредством экстрафолликулярного отклика или вводите реакцию зародышевой центральной части, где они генерируют плазму. ячейки и больше В -клеток памяти. [ 24 ] [ 25 ] Неясно, подвергаются ли В -клетки памяти дальнейшее созревание аффинности в этих вторичных GC. [ 24 ] in vitro Активация B-клеток B-клеток может быть достигнута путем стимуляции различными активаторами, такими как митоген POKEWEED или анти- CD40 моноклональные антитела , однако исследование обнаружило, что комбинация R-848 и R-848 и рекомбинантным человеком IL-2 рекомбинантного IL-2 является наиболее эффективным активатором Полем [ 26 ]
Типы В -клеток
[ редактировать ]
- Plasmablast
- Недолгая пролиферирующая антитело, секретирующая клетка, возникающая в результате дифференцировки В-клеток. [ 1 ] Плазмабласты генерируются на ранних стадиях инфекции, а их антитела, как правило, имеют более слабую сродство к их мишени антиген по сравнению с плазматической клеткой. [ 16 ] Плазмабласты могут возникнуть в результате Т-клеточной активации B-клеток или внефолликулярного ответа от T-клеток-зависимой активации B-клеток. [ 1 ]
- Плазматическая ячейка
- Долгоживая, непролификационная антитело, секретирующая клетка, возникающая в результате дифференцировки В-клеток. [ 1 ] Существуют доказательства того, что В-клетки сначала дифференцируются в плазмаблускую клетку, а затем дифференцируются в плазматическую клетку. [ 16 ] Плазматические клетки генерируются позже при инфекции и, по сравнению с плазмабластами, имеют антитела с более высокой аффинностью к их мишщему антигену из -за созревания аффинности в зародышевом центре (GC) и продуцируют больше антител. [ 16 ] Плазматические клетки обычно возникают в результате реакции зародышевого центра, зависящей от T-клеток, активации B-клеток, хотя они также могут возникнуть в результате T-клеток-независимой активации B-клеток. [ 23 ]
- Лимфоплазмацитоидные клетки
- Клетка с смесью морфологических особенностей лимфоцитов и плазматических клеток, которые, как считается, тесно связаны или подтипом плазматических клеток. Этот тип клеток обнаруживается в дискразии до знакомых и злокачественных плазматических клеток , которые связаны с секрецией IgM моноклональных белков ; Эти дискразии включают моноклональную гаммопатию IGM неопределенной значимости и макроглобулинемию Вальденстрема . [ 27 ]
- Мюмо -ячейка
- Дерные В -клеток, возникающие в результате дифференцировки В -клеток. [ 1 ] Их функция состоит в том, чтобы циркулировать через организм и инициировать более сильный, более быстрый ответ антител (известный как анамнечный ответ вторичного антитела), если они обнаруживают антиген, который активировал их родительскую В -клеток (В -клетки памяти и их родительские В -клетки имеют одинаковый BCR, таким образом, они обнаруживают тот же антиген). [ 25 ] В-клетки памяти могут быть получены из-за зависимой от Т-клеток активации как через экстрафолликулярный отклик, так и реакцию зародышевой центральной части, а также от, независимой от Т-клеток активации клеток B1. [ 25 ]
- B-2 ячейка
- FO B -клетки и MZ B -клетки. [ 28 ]
- Фолликулярная (Fo) B-клетка (также известная как клетка B-2)
- Наиболее распространенный тип B -клеток и, когда не циркулируют через кровь, встречается в основном в лимфоидных фолликулах вторичных лимфоидных органов (SLO). [ 16 ] Они несут ответственность за создание большинства антител с высокой аффинностью во время инфекции. [ 1 ]
- B-клетка с маргинальной зоной (MZ)
- Обнаружено в основном в предельной зоне селезенки и служит первой линией защиты от переносимых кровь патогенов, поскольку маргинальная зона получает большое количество крови от общей циркуляции. [ 29 ] Они могут подвергаться как независимой от Т-клеток, так и зависимой от Т-клеток активации, но преимущественно подвергаться активации, независимой от Т-клеток. [ 16 ]
- B-1 ячейка
- Возникает из пути развития, отличного от F -клеток и B -клеток MZ. [ 28 ] У мышей они преимущественно заполняют брюшную полость и плевральную полость , генерируют природные антитела (антитела, продуцируемые без инфекции), защищают от патогенов слизистой оболочки и в первую очередь демонстрируют активацию, независимую от Т-клеток. [ 28 ] Истинный гомолог мышиных клеток B-1 не был обнаружен у людей, хотя были описаны различные клеточные популяции, сходные с клетками B-1. [ 28 ]
- Регуляторная клетка B (BREG)
- Иммуносупрессивный тип B - клеток, который останавливает расширение патогенных, провоспалительных лимфоцитов посредством секреции IL-10, IL-35 и TGF-β. [ 30 ] Кроме того, он способствует генерации регуляторных клеток T (Treg) , непосредственно взаимодействуя с Т -клетками, чтобы перекосить их дифференцировку по отношению к Tregs. [ 30 ] Никакой общей идентичности клеток BREG не было описано, и многие подмножества клеток BREG, разделяющие регуляторные функции, были обнаружены как у мышей, так и у людей. [ 30 ] В настоящее время неизвестно, связаны ли подмножества клеток BREG, и как именно происходит дифференциация в ячейку BREG. [ 30 ] Существуют доказательства того, что почти все типы В -клеток могут дифференцироваться в клетку Брега посредством механизмов, включающих воспалительные сигналы и распознавание BCR. [ 30 ]
Патология, связанная с В-клетками
[ редактировать ]Аутоиммунное заболевание может быть результатом аномального распознавания В-клеток самоантигенов с последующим производством аутоантител. [ 31 ] Аутоиммунные заболевания, в которых активность заболевания коррелирует с активностью В-клеток, включают склеродермию , рассеянный склероз , системную волчанку эритематозу , диабет 1 типа , пост-инфекционные СРК и ревматоидный артрит . [ 31 ]
Злокачественная трансформация В-клеток и их предшественников может вызвать множество раковых заболеваний , включая хронический лимфоцитарный лейкоз (CLL) , острый лимфобластный лейкоз (все) , лейкоз волосатых клеток , фолликулярную лимфому , лимфому не-смодкина , лимфома Ходжкина и клетки плазмы и клетки плазмы, клетки и клетки плазмы . Как множественная миелома , макроглобулинемия Уолденстром и определенные формы амилоидоза . [ 32 ] [ 33 ]
Аномальные В-клетки могут быть относительно большими, а некоторые заболевания включают это в свои названия, такие как диффузные большие В-клеточные лимфомы (DLBCLS) и внутрисосудистая большая В-клеточная лимфома .
Пациенты с алимфоцитозом В -клеток предрасположены к инфекциям. [ 34 ]
Эпигенетика
[ редактировать ]Исследование, в котором изучалось метилем В-клеток вдоль цикла дифференцировки, с использованием секвенирования бисульфита всего генома (WGBS), показало, что существует гипометилирование с самых ранних стадий до самых дифференцированных стадий. Самая большая разница метилирования находится между стадиями B -клеток зародышевого центра и B -клетками памяти. Кроме того, это исследование показало, что существует сходство между опухолями В-клеток и долгоживущими В-клетками в их сигнатурах метилирования ДНК . [ 35 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий с Мерфи К (2012). Иммунобиология Джейнвея (8 -е изд.). Нью -Йорк: Гарлендская наука. ISBN 9780815342434 .
- ^ Jump up to: а беременный Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). «В -клетки и антитела» . Молекулярная биология клетки (4 -е изд.). Гарлендская наука.
- ^ Купер MD (март 2015 г.). «Ранняя история В -клеток» . Природные обзоры. Иммунология . 15 (3): 191–197. doi : 10.1038/nri3801 . PMID 25656707 .
- ^ Глик, Брюс; Чанг, Тимоти С.; Джаап, Р. Джордж (1956-01-01). «Бурса производства Фабриса и антител» . Наука птицы . 35 (1): 224–225. doi : 10.3382/ps.0350224 . ISSN 0032-5791 .
- ^ Jespersen, Мартин Клистер; Махаджан, Свапнил; Петерс, Бьён; Нильсен, Мортен; Marcatili, Paolo (2019). «Антитела специфичные B-клеточные прогнозы эпитопа: использование информации из белковых комплексов антител-антиген» . Границы в иммунологии . 10 : 298. doi : 10.3389/fimmu.2019.00298 . PMC 6399414 . PMID 30863406 .
- ^ Фишер У, Ян Дж.Дж., Икава Т., Хейн Д., Висенте-Дуэнс С., Боркхардт А., Санчес-Гарсия I (ноябрь 2020 г.). «Решения о судьбе клеток: роль транскрипционных факторов в раннем развитии B-клеток и лейкемии» . Открытие рака крови . 1 (3): 224–233. doi : 10.1158/2643-3230.bcd-20-0011 . PMC 7774874 . PMID 33392513 .
- ^ Jump up to: а беременный Кондо М (ноябрь 2010 г.). «Лимфоидные и миелоидные приверженность линии у мультипотентных гематопоэтических предшественников» . Иммунологические обзоры . 238 (1): 37–46. doi : 10.1111/j.1600-065x.2010.00963.x . PMC 2975965 . PMID 20969583 .
- ^ Jump up to: а беременный Pelanda R, Torres RM (апрель 2012 г.). «Центральная B-клеточная толерантность: где начинается выбор» . Перспективы Cold Spring Harbor в биологии . 4 (4): A007146. doi : 10.1101/cshperspect.a007146 . PMC 3312675 . PMID 22378602 .
- ^ Jump up to: а беременный Mårtensson IL, Almqvist N, Grimsholm O, Bernardi AI (июнь 2010 г.). «Контрольная точка рецептора до-B» . Письма Febs . 584 (12): 2572–2579. doi : 10.1016/j.febslet.2010.04.057 . PMID 20420836 . S2CID 43158480 .
- ^ Jump up to: а беременный в Lebien TW, Tedder TF (сентябрь 2008 г.). «B -лимфоциты: как они развиваются и функционируют» . Кровь . 112 (5): 1570–1580. doi : 10.1182/blood-2008-02-078071 . PMC 2518873 . PMID 18725575 .
- ^ Лодер Ф., Мушлер Б., Рэй Р.Дж., Пейдж С.Дж., Сидерас П., Торрес Р. и др. (Июль 1999). «Развитие В-клеток в селезенке происходит на дискретных шагах и определяется качеством сигналов, полученных из рецептора,» . Журнал экспериментальной медицины . 190 (1): 75–89. doi : 10.1084/jem.190.1.75 . PMC 2195560 . PMID 10429672 .
- ^ Jump up to: а беременный в Chung JB, Silverman M, Monroe JG (июнь 2003 г.). «Переходные В -клетки: шаг за шагом к иммунной компетентности». Тенденции в иммунологии . 24 (6): 343–349. doi : 10.1016/s1471-4906 (03) 00119-4 . PMID 12810111 .
- ^ Cerutti A, Cols M, Puga I (февраль 2013 г.). «B-клетки маргинальной зоны: достоинства врожденных антитело, продуцирующих лимфоциты» . Природные обзоры. Иммунология . 13 (2): 118–132. doi : 10.1038/nri3383 . PMC 3652659 . PMID 23348416 .
- ^ Harwood NE, Batista FD (2010-01-01). «Ранние события в активации В -клеток». Ежегодный обзор иммунологии . 28 (1): 185–210. doi : 10.1146/Annurev-Immunol-030409-101216 . PMID 20192804 .
- ^ Юсефф М.И., Пистобон П., Реверсии А., Леннон-Дюменл А.М. (июль 2013 г.). «Как В -клетки захватывают, обрабатывают и представляют антигены: важная роль для клеточной полярности». Природные обзоры. Иммунология . 13 (7): 475–486. doi : 10.1038/nri3469 . PMID 23797063 . S2CID 24791216 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж Nutt SL, Hodgkin PD, Tarlinton DM, Corcoran LM (март 2015 г.). «Генерация плазматических клеток, секретирующих антител». Природные обзоры. Иммунология . 15 (3): 160–171. doi : 10.1038/nri3795 . PMID 25698678 . S2CID 9769697 .
- ^ Asokan R, Banda NK, Szakonyi G, Chen XS, Holers VM (январь 2013 г.). «Человеческий рецептор комплемента 2 (CR2/CD21) в качестве рецептора для ДНК: последствия для его роли в иммунном ответе и патогенез системной волчанки (SLE)» . Молекулярная иммунология . 53 (1–2): 99–110. doi : 10.1016/j.molimm.2012.07.002 . PMC 3439536 . PMID 22885687 .
- ^ Забель М.Д., Вейс Дж. Х. (март 2001 г.). «Клеточная регуляция гена CD21». Международная иммунофармакология . Раскрытие механизмов и открытие новых ролей для дополнения. 1 (3): 483–493. doi : 10.1016/s1567-5769 (00) 00046-1 . PMID 11367532 .
- ^ Blum JS, Worsch PA, Cresswell P (2013-01-01). «Пути обработки антигена» . Ежегодный обзор иммунологии . 31 (1): 443–473. doi : 10.1146/Annurev-Immunol-032712-095910 . PMC 4026165 . PMID 23298205 .
- ^ Jump up to: а беременный в дюймовый Crotty S (март 2015 г.). «Краткая история Т -клеток помогает В -клеткам» . Природные обзоры. Иммунология . 15 (3): 185–189. doi : 10.1038/nri3803 . PMC 4414089 . PMID 25677493 .
- ^ MacLennan IC, Toellner KM, Cunningham AF, Serre K, Sze DM, Zúñiga E, et al. (Август 2003 г.). «Экстрафолликулярные реакции антител». Иммунологические обзоры . 194 : 8–18. doi : 10.1034/j.1600-065x.2003.00058.x . PMID 12846803 . S2CID 2455541 .
- ^ Jump up to: а беременный Shlomchik MJ, Weisel F (май 2012 г.). «Выбор зародышевого центра и развитие памяти B и плазматических клеток» . Иммунологические обзоры . 247 (1): 52–63. doi : 10.1111/j.1600-065x.2012.01124.x . PMID 22500831 . S2CID 5362003 .
- ^ Jump up to: а беременный Bortnick A, Chernova I, Quinn WJ, Mugnier M, Cancro MP, Allman D (июнь 2012 г.). «Долгоживущие плазматические клетки костного мозга индуцируются на ранних этапах в ответ на независимые от Т-клеток или Т-клеточные антигены» . Журнал иммунологии . 188 (11): 5389–5396. doi : 10.4049/jimmunol.1102808 . PMC 4341991 . PMID 22529295 .
- ^ Jump up to: а беременный в дюймовый и Mcheyzer-Williams M, Okitsu S, Wang N, Mcheyzer-Williams L (декабрь 2011 г.). «Молекулярное программирование памяти В -клеток» . Природные обзоры. Иммунология . 12 (1): 24–34. doi : 10.1038/nri3128 . PMC 3947622 . PMID 22158414 .
- ^ Jump up to: а беременный в дюймовый Kurosaki T, Kometani K, Ise W (март 2015 г.). «В -клетки памяти». Природные обзоры. Иммунология . 15 (3): 149–159. doi : 10.1038/nri3802 . PMID 25677494 . S2CID 20825732 .
- ^ Jahnmatz, Maja; Кеса, пистолет; Netterlid, Eva; Buisman, Anne-Marie; Торстенссон, Ригмор; Альборг, Никлас (2013-05-31). «Оптимизация анализа IgG B-клеток IgG B-клеток для анализа вакцин-индуцированных B-клеточных реакций» . Журнал иммунологических методов . 391 (1): 50–59. doi : 10.1016/j.jim.2013.02.009 . ISSN 0022-1759 . PMID 23454005 .
- ^ Ribourtout B, Zandecki M (июнь 2015 г.). «Морфология плазматических клеток при множественной миеломе и связанных с ним расстройствах». Морфология . 99 (325): 38–62. doi : 10.1016/j.morpho.2015.02.001 . PMID 25899140 . S2CID 1508656 .
- ^ Jump up to: а беременный в дюймовый Baumgarth N (январь 2011 г.). «Двойной срок службы ячейки B-1: самореактивность выбирает для защитных эффекторных функций». Природные обзоры. Иммунология . 11 (1): 34–46. doi : 10.1038/nri2901 . PMID 21151033 . S2CID 23355423 .
- ^ Pillai S, Cariappa A, Moran St (2005-01-01). «B -клетки маргинальной зоны». Ежегодный обзор иммунологии . 23 (1): 161–196. doi : 10.1146/annurev.immunol.23.021704.115728 . PMID 15771569 .
- ^ Jump up to: а беременный в дюймовый и Rosser EC, Mauri C (апрель 2015 г.). «Регуляторные В -клетки: происхождение, фенотип и функция» . Иммунитет . 42 (4): 607–612. doi : 10.1016/j.immuni.2015.04.005 . PMID 25902480 .
- ^ Jump up to: а беременный Yanaba K, Bouaziz JD, Matsushita T, Magro CM, St Clair EW, Tedder TF (июнь 2008 г.). «Вклад B-лимфоцитов в аутоиммунное заболевание человека». Иммунологические обзоры . 223 (1): 284–299. doi : 10.1111/j.1600-065x.2008.00646.x . PMID 18613843 . S2CID 11593298 .
- ^ Shaffer AL, Young RM, Staudt LM (2012-01-01). «Патогенез лимфом В -клеток человека» . Ежегодный обзор иммунологии . 30 (1): 565–610. doi : 10.1146/Annurev-Immunol-020711-075027 . PMC 7478144 . PMID 22224767 .
- ^ Кастильо JJ (декабрь 2016 г.). «Расстройства плазменных клеток». Первичная помощь . 43 (4): 677–691. doi : 10.1016/j.pop.2016.07.002 . PMID 27866585 .
- ^ Grammatikos Александрос, Донати Мэтью, Джонстон Сара Л., Гоммельс Марк М. Дефицит периферических В -клеток и предрасположенность к вирусным инфекциям: парадигма иммунных дефицитов. Границы в иммунологии (12) 2021 https://www.frontiersin.org/articles/10.3389/fimmu.2021.731643 doi = 10.3389/fimmu.2021.731643
- ^ Кулис М., Меркель А., Хит С., Кейрос А.С., Шайлер Р.П., Кастеллано Г. и др. (Июль 2015). «Отпечаток пальца всего генома ДНК-метилома во время дифференцировки В-клеток человека» . Природа генетика . 47 (7): 746–756. doi : 10.1038/ng.3291 . PMC 5444519 . PMID 26053498 .