Теории общего анестезирующего действия

Общий анестетик (или анестетик ) — препарат , вызывающий обратимую потерю сознания . [2] Эти препараты обычно вводятся анестезиологом/ анестезиологом для индукции или поддержания общей анестезии для облегчения операции .
Общие анестетики получили широкое применение в хирургии с 1842 года, когда Кроуфорд Лонг впервые ввел диэтиловый эфир больному и провел безболезненную операцию. Долгое время считалось, что общие анестетики оказывают свое действие (обезболивание, потеря сознания, неподвижность). [3] через мембрано-опосредованный механизм или путем прямой модуляции активности мембранных белков в мембране нейрона. существует множество неисключающих молекулярных мишеней В целом, разные анестетики обладают разными механизмами действия, так что на всех уровнях интеграции в центральной нервной системе . [4] Однако считается, что для некоторых внутривенных анестетиков, таких как пропофол и этомидат , основной молекулярной мишенью является ГАМК А рецептор , причем решающую роль играют определенные β-субъединицы. [5] [6] [7]
Концепция специфических взаимодействий между рецепторами и лекарствами, впервые предложенная Паулем Эрлихом в 1897 году. [8] утверждает, что лекарства действуют только тогда, когда они связаны со своими мишенями (рецепторами). [1] Идентификация конкретных молекулярных мишеней для общих анестетиков стала возможной только благодаря современному развитию методов молекулярной биологии для выявления одиночных аминокислотных мутаций в белках генно -инженерных мышей . [1] [5] [6] [7]
Корреляция растворимости в липидах и эффективности анестезии (корреляция Мейера-Овертона)
[ редактировать ]
Неспецифический механизм действия общего анестетика был впервые предложен Эмилем Харлессом и Эрнстом фон Биброй в 1847 году. [9] Они предположили, что общие анестетики могут действовать, растворяясь в жировой фракции клеток головного мозга и удаляя из них жировые компоненты, изменяя тем самым активность клеток головного мозга и вызывая анестезию. В 1899 году Ганс Хорст Мейер опубликовал первые экспериментальные доказательства того, что эффективность анестетика связана с растворимостью в липидах. [10] [11] Два года спустя аналогичная теория была независимо опубликована Чарльзом Эрнестом Овертоном . [12]
Мейер сравнил эффективность многих агентов, определяемую как обратную молярную концентрацию, необходимую для индукции анестезии у головастиков, с их коэффициентом распределения оливковое масло/вода . Он обнаружил почти линейную зависимость между эффективностью и коэффициентом распределения для многих типов молекул анестетика, таких как спирты , альдегиды , кетоны , простые и сложные эфиры .Концентрация анестетика, необходимая для индукции анестезии у 50% популяции животных ( ЕС 50 ), не зависела от способа доставки анестетика, т.е. от газовой или водной фазы. [10] [11] [13]
Мейер и Овертон обнаружили поразительную корреляцию между физическими свойствами молекул общих анестетиков и их эффективностью: чем выше растворимость в липидах соединения в оливковом масле, тем выше его анестезирующая эффективность. [13] Эта корреляция справедлива для широкого спектра анестетиков с липидорастворимостью более 4-5 порядков, если в качестве масляной фазы используется оливковое масло. Эта корреляция может быть значительно улучшена как с точки зрения качества корреляции, так и с точки зрения расширения спектра анестетиков, если объем октанола [14] или полностью гидратированный жидкий липидный бислой [15] [16] [17] [18] используется в качестве «масляной» фазы. Было также отмечено, что эффект ингаляционных анестетиков аддитивен (смесь половинных доз двух различных ингаляционных анестетиков давала тот же анестезирующий эффект, что и полная доза любого препарата по отдельности).
Наиболее изученный участок анестетика, учитывающий корреляцию Мейера-Овертона, находится в упорядоченных липидных доменах. Анестетики неспецифически прикрепляются к поверхности участка специфического связывания пальмитата внутри липидной мембраны, вытесняя пальмитат из упорядоченных липидов GM1. Этот процесс приводит к возникновению компонента мембранно-опосредованной анестезии . [19] Аналогичный механизм был показан для люциферазы. [20] Анестетики неспецифически связывались с гидрофобной поверхностью и превосходили специфическое связывание люциферина. Однако люцифераза не имеет физиологического значения для позвоночных, поскольку она не экспрессируется у позвоночных эндогенно.
Ранние липидные гипотезы действия общих анестетиков
[ редактировать ]
На основании корреляции между растворимостью в липидах и анестезирующей активностью Мейер и Овертон предположили единый механизм общей анестезии. Они предположили, что солюбилизация липофильного анестетика в липидном бислое нейрона вызывает его нарушение и анестезирующий эффект при достижении критической концентрации анестетика. Позже, в 1973 году, Миллер и Смит предложили гипотезу критического объема, также называемую гипотезой расширения липидного бислоя. [21] Они предположили, что объемные и гидрофобные молекулы анестетика накапливаются внутри гидрофобных (или липофильных) участков липидной мембраны нейронов, вызывая ее искажение и расширение (утолщение) за счет смещения объема. Накопление критических количеств анестетика вызывает утолщение мембраны, достаточное для обратимого изменения функции мембранных ионных каналов, обеспечивая тем самым анестезирующий эффект. Реальная химическая структура анестетика сама по себе не важна, но основную роль играет его молекулярный объем: чем больше места внутри мембраны занимает анестетик, тем сильнее анестезирующий эффект.Основываясь на этой теории, в 1954 году Маллинз предположил, что корреляцию Мейера-Овертона с эффективностью можно улучшить, если принять во внимание молекулярные объемы молекул анестетика. [22] Эта теория существовала более 60 лет и подтверждалась экспериментальным фактом, что повышение атмосферного давления приводит к обратному анестезирующему эффекту ( эффект реверсирования давления ). [21] [23] [24]
Затем появились другие, физико-химические теории анестезирующего действия, которые учитывали разнообразную химическую природу общих анестетиков и предполагали, что анестезирующий эффект осуществляется за счет некоторого возмущения липидного бислоя. [25] Было предложено несколько типов возмущений бислоя, вызывающих анестезирующий эффект, включая (1) изменения в разделении фаз, (2) изменения толщины бислоя, (3) изменения параметров порядка или (4) изменения эластичности кривизны. [26] [27] [28]
Согласно теории латерального фазового разделения [28] анестетики оказывают свое действие путем разжижения нервных мембран до такой степени, что фазовые разделения в критических липидных областях исчезают. Это псевдоожижение, вызванное анестезией, делает мембраны менее способными способствовать конформационным изменениям в белках, которые могут быть основой для таких мембранных событий, как ионное пропускание, высвобождение синаптического медиатора и связывание медиатора с рецепторами. Более поздние методы визуализации сверхвысокого разрешения показывают, что анестетики не преодолевают разделение фаз — разделение фаз сохраняется. Достаточно насыщенные липиды в процессе разделения фаз могут претерпевать переход от упорядоченного состояния к неупорядоченному, на что резко влияют анестетики. Тем не менее, теперь было показано, что концепция белков, перемещающихся между разделенными фазами липидами в ответ на анестетик, верна. [29]
Считалось, что все эти ранние теории липидов имеют четыре недостатка: [1] (полное описание с опровержениями смотрите в разделах ниже):
- Стереоизомеры анестезирующего препарата имеют очень разную анестезирующую эффективность, тогда как их коэффициенты распределения нефть/газ схожи.
- Некоторые препараты, которые хорошо растворяются в липидах и поэтому должны действовать как анестетики, вместо этого оказывают судорожный эффект (и поэтому были названы нониммобилизаторами ).
- Небольшое повышение температуры тела влияет на плотность и текучесть мембраны так же, как и общая анестезия, но не вызывает анестезии.
- Увеличение длины цепи в гомологическом ряду спиртов или алканов с прямой цепью увеличивает их растворимость в липидах, но их анестезирующая эффективность перестает увеличиваться после определенной предельной длины.
Считалось, что корреляция между растворимостью в липидах и эффективностью общих анестетиков является необходимым, но недостаточным условием для определения липидного участка-мишени. Общие анестетики с таким же успехом могут связываться с гидрофобными целевыми сайтами на белках головного мозга, но, учитывая химическое разнообразие анестетиков, вероятно, потребуется включать более одного сайта, и эти сайты по своей сути не будут исключать сайт в мембране. Что касается белков, то одна из причин, по которой более полярные общие анестетики могут быть менее эффективными, заключается в том, что им приходится преодолевать гематоэнцефалический барьер , чтобы оказать воздействие на нейроны головного мозга.
Современная липидная гипотеза
[ редактировать ]
Существуют две современные липидные гипотезы, которые не исключают прямого связывания с белками. Самая последняя гипотеза постулирует, что упорядоченные липиды в плазматической мембране содержат структурированный сайт связывания липид- пальмитата . Это сайт связывания липидов внутри липидной структуры, а не белковой структуры. Белки, которые содержат ковалентно присоединенный пальмитат ( пальмитоилирование ), нацеливаются на упорядоченные липиды посредством специфического липид-липидного взаимодействия. Связывание пальмитата с липидным доменом зависит от холестерина, и клетка регулирует белок посредством наноскопической локализации. Анестетики действуют путем неспецифического связывания с участком связывания пальмитата, что нарушает способность холестерина связываться с белком и переводить его в неактивное состояние. Этот мембранно-опосредованный механизм был экспериментально продемонстрирован Павлом и его коллегами в 2020 году. Они показали, что фермент фосфолипаза D2 (PLD2) чувствителен к анестетикам и активирует калиевый канал TREK-1 посредством Мембранно-опосредованный механизм . Анестетики вытеснили PLD2 из упорядоченных липидных доменов, что позволило ферменту активироваться путем презентации субстрата и активировать канал. [29] [30]
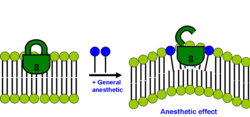
Вторая липидная гипотеза утверждает, что анестезирующий эффект возникает, если солюбилизация общего анестетика в бислое вызывает перераспределение латерального давления мембраны. [31] [32]
Каждая двухслойная мембрана имеет особый профиль распределения внутри нее латерального давления. Большинство мембранных белков (особенно ионные каналы) чувствительны к изменениям профиля распределения латерального давления. Эти латеральные напряжения довольно велики и изменяются в зависимости от глубины мембраны. Согласно современной липидной гипотезе, изменение профиля латерального давления мембраны смещает конформационное равновесие некоторых мембранных белков, на которые, как известно, влияют клинические концентрации анестетиков, такие как лиганд-управляемые ионные каналы. Этот механизм также неспецифичен, поскольку эффективность анестетика определяется не его фактической химической структурой, а позиционным и ориентационным распределением его сегментов и связей внутри бислоя.
В 1997 году Кантор предложил подробный механизм общей анестезии, основанный на решеточной статистической термодинамике. [32] Было высказано предположение, что включение амфифильных и других межфазно-активных растворов (например, средств для общей анестезии) в бислой избирательно увеличивает латеральное давление вблизи границ с водной поверхностью, что компенсируется снижением латерального давления по направлению к центру бислоя. Расчеты показали, что общая анестезия, вероятно, включает ингибирование открытия ионного канала в постсинаптическом лиганд-управляемом мембранном белке. [32] по следующему механизму:
- Канал пытается открыться в ответ на нервный импульс, тем самым увеличивая площадь поперечного сечения белка ближе к водной границе раздела, чем в середине бислоя;
- Затем вызванное анестезией увеличение латерального давления вблизи границы раздела сдвигает конформационное равновесие белка обратно в закрытое состояние, поскольку открытие канала потребует большей работы против более высокого давления на границе раздела. Это первая гипотеза, которая обеспечила не только корреляцию активности со структурными или термодинамическими свойствами, но и детальное механистическое и термодинамическое понимание анестезии.

Таким образом, согласно современной липидной гипотезе, анестетики не действуют непосредственно на мембранные белки-мишени, а скорее воздействуют на специализированные липидные матрицы на границе белок-липид, которые действуют как медиаторы. Это новый тип механизма трансдукции, отличающийся от обычного взаимодействия лиганда и рецептора по принципу «ключ-замок», при котором анестетик (лиганд) влияет на функцию мембранных белков путем связывания со специфическим участком белка. Таким образом, предполагается, что некоторые мембранные белки чувствительны к своему липидному окружению.В том же году был предложен немного другой подробный молекулярный механизм того, как возмущения бислоя могут влиять на ионный канал. Олеамид (амид жирной кислоты олеиновой кислоты) представляет собой эндогенный анестетик, обнаруженный in vivo (в мозге кошки), и известно, что он усиливает сон и снижает температуру тела, закрывая соединение каналов щелевого соединения. [33] Подробный механизм показан на рисунке: упорядоченное липидное (зеленое)/холестериновое (желтое) кольцо, существующее вокруг коннексона (пурпурный), нарушается при лечении анестетиком (красные треугольники), что способствует закрытию ионного канала коннексона. Это снижает активность мозга, вызывает летаргию и анестезирующий эффект.Недавно визуализация со сверхвысоким разрешением продемонстрировала прямые экспериментальные доказательства того, что летучие анестетики нарушают упорядоченные липидные домены, как и предполагалось. [34] В том же исследовании выявился схожий механизм, при котором анестетики высвобождали фермент фосфолипазу D (PLD) из липидных доменов, а фермент связывался с каналами TREK-1 и активировал их путем производства фосфатидной кислоты. Эти результаты экспериментально показали, что мембрана является физиологически значимой мишенью для общих анестетиков.
Мембранная белковая гипотеза действия общего анестетика
[ редактировать ]
В начале 1980-х годов Николас П. Фрэнкс и Уильям Р. Либ [35] продемонстрировали, что корреляцию Мейера-Овертона можно воспроизвести с использованием растворимого белка. Они обнаружили, что два класса белков инактивируются клиническими дозами анестетика при полном отсутствии липидов. Это люциферазы , которые используются биолюминесцентными животными и бактериями для производства света. [36] и цитохром Р450 , [37] Это группа гемовых белков, которые гидроксилируют разнообразную группу соединений, включая жирные кислоты , стероиды и ксенобиотики , такие как фенобарбитал . Примечательно, что ингибирование этих белков общими анестетиками напрямую коррелировало с их анестезирующей активностью. Ингибирование люциферазы также приводит к отсечению длинной цепи алкоголя, что связано с размером кармана, связывающего анестетик. [38]
Эти наблюдения были важны, поскольку они продемонстрировали, что общие анестетики оказывают свое действие неспецифически, в том числе при связывании с белками. Это также открыло возможность того, что анестетики могут действовать посредством прямого связывания с белками, а не влиять на мембранные белки косвенно через неспецифические взаимодействия с липидным бислоем в качестве медиатора. [14] [39] Показано, что анестетики изменяют функции многих цитоплазматических сигнальных белков, в том числе протеинкиназы С. [40] [41]
Однако белками, которые считаются наиболее вероятными молекулярными мишенями анестетиков, являются ионные каналы. Согласно этой теории, общие анестетики гораздо более селективны, чем в липидной гипотезе, и они напрямую связываются только с небольшим количеством мишеней в центральной нервной системе, в основном с лиганд-зависимыми ионными каналами в синапсах и рецепторах, связанных с G-белком , изменяя их поток ионов. В частности, рецепторы Cys-петли. [42] являются вероятными мишенями для общих анестетиков, которые связываются на границе между субъединицами. Суперсемейство рецепторов Cys-петли включает тормозные рецепторы ( ГАМК А рецепторы , ГАМК С рецепторы , глициновые рецепторы ) и возбуждающие рецепторы ( никотиновый ацетилхолиновый рецептор и 5-НТ3-серотониновый рецептор ). Общие анестетики могут ингибировать канальные функции возбуждающих рецепторов или потенцировать функции тормозных рецепторов соответственно.
Расположение сайтов неспецифического связывания в ионных каналах по-прежнему остается важным вопросом в этой области. В частности, как соединение, следующее за Овертоном-Мейером, напрямую вызывает конформационные изменения в белке? Обычно аллостерическая регуляция включает изменение формы белка, обеспечивающее связывание лиганда. Этот механизм отличается от механизма люциферазы. Второй важный вопрос: как неспецифические сайты связывания белков сохраняются у разных видов и почему они обычно ингибируют возбуждающие рецепторы и усиливают тормозные рецепторы?
Ряд экспериментальных и компьютерных исследований показали, что общие анестетики могут изменять динамику гибких петель, которые соединяют α-спирали в пучок и подвергаются воздействию мембранно-водной границы рецепторов Cys-петли. [43] [44] [45] [46] [47] [48] Однако основные карманы связывания общих анестетиков расположены внутри трансмембранных четырех-альфа-спиральных пучков рецепторов Cys-петли. [49] [50] [51]
Рецептор ГАМК А является основной мишенью общих анестетиков.
[ редактировать ]
Рецептор ГАМК А R) представляет собой ионотропный рецептор , (ГАМК А активируемый тормозным нейротрансмиттером γ-аминомасляной кислотой (ГАМК). Активация рецептора ГАМК А приводит к притоку ионов хлора , что вызывает гиперполяризацию мембран нейронов. [52] Рецептор ГАМК А был идентифицирован как основная мишень внутривенных анестетиков, таких как пропофол и этомидат . [4] [5] Сайт связывания пропофола с рецепторами ГАМК А млекопитающих был идентифицирован путем фотомечения с использованием производного диазирина . [53] Сильная активация тонической проводимости рецептора ГАМК А клиническими концентрациями пропофола была подтверждена электрофизиологическими записями нейронов СА1 гиппокампа на взрослых крыс срезах мозга . [54] Рецепторы ГАМК А , содержащие β3-субъединицы, являются основными молекулярными мишенями для анестезирующего действия этомидата , тогда как β2-содержащие рецепторы ГАМК А участвуют в седативном эффекте, вызываемом этим препаратом. [55] Электрофизиологические эксперименты с амнестическими концентрациями этомидата также показали усиление тонической проводимости ГАМК А СА1 пирамидных нейронов в срезах гиппокампа. [56] Мощная активация ГАМК - опосредованного ингибирования рецептора, приводящая к сильному снижению частоты возбуждения нейронов неокортекса, также была продемонстрирована для клинических концентраций летучих анестетиков, таких как изофлюран , энфлуран и галотан . [57]
Другие молекулярные мишени
[ редактировать ]Повышение активности рецептора ГАМК А вряд ли будет единственным механизмом, объясняющим широкий спектр поведенческих эффектов общих анестетиков. [1] что модуляция калиевых каналов двухпоровых доменов Накопление экспериментальных данных позволяет предположить , [58] [59] или потенциалзависимые натриевые каналы [60] может также объяснить некоторые действия летучих анестетиков. Альтернативно, ингибирование глутамат-управляемых N-метил-D-аспартатных рецепторов , кетамином ксеноном и закисью азота обеспечивает механизм действия, соответствующий преобладающему анальгетическому профилю. [1]
Исторические возражения против ранних липидных гипотез
[ редактировать ]1. Стереоизомеры анестетика.
[ редактировать ]Стереоизомеры, которые представляют собой зеркальные изображения друг друга, называются энантиомерами или оптическими изомерами (например, изомеры R-(+)- и S-(-)-этомидата). [1] Физико-химические эффекты энантиомеров всегда одинаковы в ахиральном окружении (например, в липидном бислое). Однако in vivo энантиомеры многих общих анестетиков (например, изофлуран , тиопентал , этомидат ) могут сильно различаться по своей анестезирующей активности, несмотря на схожие коэффициенты распределения масло/газ. [61] [62] Например, R-(+)-изомер этомидата в 10 раз более сильный анестетик, чем его S-(-)-изомер. [1] Это означает, что оптические изомеры одинаково распределяются в липиды, но оказывают различное влияние на ионные каналы и синаптическую передачу . Это возражение представляет собой убедительное доказательство того, что основной мишенью анестетиков является не сам ахиральный липидный бислой, а скорее стереоселективные сайты связывания на мембранных белках, которые обеспечивают хиральную среду для специфических взаимодействий стыковки анестетика и белка. [1]
Опровержение возражения: 1) Стереоселективный транспорт анестетика никогда не рассматривался. Анестетики гидрофобны и транспортируются в виде белков крови. Любое стереоселективное связывание с транспортным белком приведет к изменению концентрации в месте действия. Более того, сток белка в мембрану может немного лучше связывать один из изомеров и снижать эффективную концентрацию, которую испытывает мембрана. Все стереоизомеры являются эффективными анестетиками, они лишь меняют чувствительность, что позволяет предположить необходимость рассмотрения селективного транспорта и селективных поглотителей белка. 2) Липиды хиральны, как и белки. И, как и белки, липиды имеют упорядоченные и неупорядоченные области. [63] [64] В этой области не удалось исследовать хиральность упорядоченных липидов из-за отсутствия знаний об их существовании.
2. Неиммобилайзеры
[ редактировать ]Все общие анестетики вызывают иммобилизацию (отсутствие движения в ответ на вредные раздражители) посредством угнетения функций спинного мозга, тогда как их амнестическое действие проявляется в головном мозге. Согласно корреляции Мейера-Овертона, анестезирующая эффективность препарата прямо пропорциональна его растворимости в липидах, однако существует множество соединений, которые не удовлетворяют этому правилу. Эти препараты поразительно похожи на сильные общие анестетики и, по прогнозам, являются сильными анестетиками на основании их жирорастворимости, но они оказывают только одну составляющую анестезирующего действия (амнезию) и не подавляют движения (т. е. не угнетают функции спинного мозга), как все анестетики действуют. [65] [66] [67] [68] Эти препараты называются нониммобилайзерами. Существование ноиммобилайзеров позволяет предположить, что анестетики вызывают разные компоненты анестезирующего эффекта (амнезию и неподвижность), воздействуя на разные молекулярные мишени, а не только на одну мишень (нейронный бислой), как считалось ранее. [69] Хорошим примером неиммобилайзеров являются галогенированные алканы, которые очень гидрофобны, но не способны подавлять движение в ответ на вредное раздражение в соответствующих концентрациях. См. также: флуротил .
Опровержение возражения: Это логическая ошибка. Гипотеза не требует, чтобы каждая когда-либо протестированная молекула подчинялась гипотезе, чтобы гипотеза была верной. Существование менее 10-20 родственных соединений, которые, как известно, не подчиняются гипотезе Мейера-Овертона, никоим образом не отменяет существования сотен, если не тысяч химически разнообразных соединений, которые действительно подчиняются гипотезе Овертона-Мейера. Исключения могут существовать по причинам, не связанным с механизмом, лежащим в основе гипотезы Мейера-Овертона.
3. Повышение температуры не оказывает обезболивающего эффекта.
[ редактировать ]Экспериментальные исследования показали, что общие анестетики, включая этанол, являются мощными разжижителями естественных и искусственных мембран. Однако изменения плотности и текучести мембран в присутствии клинических концентраций общих анестетиков настолько малы, что относительно небольшое повышение температуры (~1 °C) может имитировать их, не вызывая анестезии. [70] Изменение температуры тела примерно на 1 °C находится в пределах физиологического диапазона и явно недостаточно для того, чтобы вызвать потерю сознания как таковую. Таким образом, мембраны разжижаются только большими количествами анестетиков, но текучесть мембран не меняется, когда концентрации анестетиков малы и ограничены фармакологически значимыми.
Опровержение возражения: ранние исследования учитывали только текучесть липидной мембраны. Недавняя работа показала, что изменения температуры могут происходить на несколько градусов в упорядоченных наноскопических липидных доменах. [71] Кроме того, текучесть активно регулируется десатуразами жирных кислот . И, наконец, конкуренция анестетиков с пальмитоилированными белками происходит независимо от температуры и несмотря на увеличение упорядоченных липидов. [29]
4. Эффект исчезает за определенной длиной цепочки.
[ редактировать ]Согласно корреляции Мейера-Овертона, в гомологическом ряду любого общего анестетика (например, н - спиртов или алканов) увеличение длины цепи увеличивает растворимость в липидах и, таким образом, должно вызывать соответствующее увеличение эффективности анестетика. Однако за пределами определенной длины цепи анестезирующий эффект исчезает. Для н -спиртов это обрыв происходит при длине углеродной цепи около 13 [72] и для н -алканов с длиной цепи от 6 до 10, в зависимости от вида. [73]

Если общие анестетики разрушают ионные каналы, разделяя и нарушая липидный бислой, то можно было бы ожидать, что их растворимость в липидных бислоях также будет проявлять эффект отсечки. Однако разделение спиртов на липидные бислои не обнаруживает границы для спиртов с длинной цепью от н - деканола до н - пентадеканола . График зависимости длины цепи от логарифма коэффициента распределения липидного бислоя/буфера K является линейным, при этом добавление каждой метиленовой группы вызывает изменение свободной энергии Гиббса на -3,63 кДж/моль.
Эффект отсечки был впервые интерпретирован как доказательство того, что анестетики оказывают свое действие не за счет глобального воздействия на липиды мембран, а скорее за счет непосредственного связывания с гидрофобными карманами четко определенных объемов в белках. По мере роста алкильной цепи анестетик заполняет большую часть гидрофобного кармана и связывается с большей аффинностью. Когда молекула слишком велика, чтобы полностью поместиться в гидрофобный карман, аффинность связывания больше не увеличивается с увеличением длины цепи. Таким образом, объем цепи н-алканола на предельной длине дает оценку объема сайта связывания. Это возражение легло в основу белковой гипотезы обезболивающего эффекта (см. ниже).

Однако эффект отсечения все же можно объяснить в рамках липидной гипотезы. [31] [74] В короткоцепочечных алканолах (А) участки цепи достаточно жесткие (с точки зрения конформационной энтропии) и очень близки к гидроксильной группе, привязанной к водной межфазной области («буй»). Следовательно, эти сегменты эффективно перераспределяют латеральные напряжения из внутренней части бислоя в сторону границы раздела. В длинноцепочечных алканолах (В) участки углеводородной цепи расположены дальше от гидроксильной группы и более гибки, чем в короткоцепочечных алканолах. Эффективность перераспределения давления снижается по мере увеличения длины углеводородной цепи до тех пор, пока в какой-то момент эффективность анестетика не потеряется. Было высказано предположение, что полиалканолы (С) будут оказывать анестезирующее действие, аналогичное короткоцепочечным 1-алканолам, если длина цепи между двумя соседними гидроксильными группами меньше предельной. [75] Эта идея была подтверждена экспериментальными данными, поскольку полигидроксиалканы 1,6,11,16-гексадекантетраол и 2,7,12,17-октадекантетраол проявляли значительную анестезирующую эффективность, как предполагалось первоначально. [74]
Опровержение возражения: Аргумент предполагает, что все классы анестетиков должны одинаково действовать на мембрану. Вполне возможно, что один или два класса молекул могут действовать по немембранному механизму. Например, было показано, что спирты внедряются в липидную мембрану посредством ферментативной реакции трансфосфатидилирования. [76] Метаболит этанола связывался с анестетическим каналом и ингибировал его. И хотя этот механизм может противоречить единому механизму анестезии, он не исключает мембранопосредованного механизма.
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я Вейр, Кэмерон Дж. (2006). «Молекулярные механизмы общей анестезии: анализ рецептора ГАМК А » . Непрерывное образование в области анестезиологии, интенсивной терапии и боли . 6 (2): 49–53. doi : 10.1093/bjaceaccp/mki068 .
- ^ Миллер, Рональд Д.; Коэн, Нил Х.; Эрикссон, Ларс И.; Флейшер, Ли А.; Винер-Крониш, Жанин П.; Янг, Уильям Л. (2014). Анестезия Миллера (8-е изд.). Филадельфия: Сондерс. ISBN 978-0-7020-5283-5 . OCLC 892338436 .
- ^ Иган, Талмейдж Д. (2019). «Незаменимы ли опиоиды для общей анестезии?» . Британский журнал анестезии . 122 (6): е127–е135. дои : 10.1016/j.bja.2019.02.018 . ПМИД 31104756 . S2CID 133023216 .
- ^ Jump up to: а б Урбан, BW (2002). «Современная оценка целей и теорий анестезии» . Британский журнал анестезии . 89 (1): 167–183. дои : 10.1093/bja/aef165 . ПМИД 12173228 .
- ^ Jump up to: а б с Фрэнкс, Николас П. (2006). «Молекулярные мишени, лежащие в основе общей анестезии» . Британский журнал фармакологии . 147 (С1): С72–С81. дои : 10.1038/sj.bjp.0706441 . ПМК 1760740 . ПМИД 16402123 .
- ^ Jump up to: а б Вейр, CJ; Митчелл, С.Дж.; Ламберт, Джей-Джей (2017). «Роль подтипов рецепторов ГАМК А в поведенческих эффектах внутривенных общих анестетиков» . Британский журнал анестезии . 119 (дополнение_1): i167–i175. дои : 10.1093/bja/aex369 . ПМИД 29161398 .
- ^ Jump up to: а б Дрекслер, Бертольд; Антковяк, Бернд; Энгин, Элиф; Рудольф, Уве (2011). «Идентификация и характеристика мишеней для анестетика с помощью подходов молекулярной генетики мышей» . Канадский журнал анестезии . 58 (2): 178–190. дои : 10.1007/s12630-010-9414-1 . ПМК 3330822 . ПМИД 21174184 .
- ^ Мэле, Андреас-Хольгер (2009). «Обязательный вопрос: эволюция концепции рецептора» . Стараться . 33 (4): 135–140. doi : 10.1016/j.endeavour.2009.09.001 . ПМК 2812702 . ПМИД 19837460 .
- ^ Харлесс, Эмиль; Бибра, Эрнст (1847). Результаты опытов по воздействию серного эфира . Эрланген.
- ^ Jump up to: а б Мейер, Ганс Хорст (1899). «К теории алкогольного наркоза. Первое сообщение. Какое свойство анестетиков определяет их наркотический эффект?» . Архив экспериментальной патологии и фармакологии . 42 (2–4): 109–118. дои : 10.1007/BF01834479 . S2CID 7040253 .
- ^ Jump up to: а б Мейер, Ганс Хорст (1901). «К теории алкогольного наркоза. Третье сообщение. Влияние изменения температуры на эффективность и коэффициент распределения наркотических средств» . Архив экспериментальной патологии и фармакологии . 46 (5–6): 338–346. дои : 10.1007/BF01978064 . S2CID 30441885 .
- ^ Овертон, Чарльз Эрнест (1901). Исследования по анестезии: одновременно вклад в общую фармакологию . Йена: Густав Фишер. OCLC 876369243 .
- ^ Jump up to: а б Мейер, Курт Х. (1937). «Вклад в теорию наркоза». Труды Фарадеевского общества . 33 : 1062–1068. дои : 10.1039/tf9373301062 .
- ^ Jump up to: а б Фрэнкс, Николас П .; Либ, Уильям Р. (1978). «Где действуют общие анестетики?». Природа . 274 (5669): 339–342. Бибкод : 1978Natur.274..339F . дои : 10.1038/274339a0 . ПМИД 672957 . S2CID 4200246 .
- ^ Янофф А.С., Прингл М.Дж., Миллер К.В. (1981). «Корреляция эффективности общего анестетика с растворимостью в мембранах». Biochimica et Biophysical Acta (BBA) – Биомембраны . 649 (1): 125–128. дои : 10.1016/0005-2736(81)90017-1 . ПМИД 7306543 .
- ^ Тахери С., Хэлси М.Дж., Лю Дж., Эгер Э.И., Коблин Д.Д., Ластер М.Дж. (1991). «Какой растворитель лучше всего представляет место действия ингаляционных анестетиков у людей, крыс и собак?» . Анестезия и анальгезия . 72 (5): 627–634. дои : 10.1213/00000539-199105000-00010 . ПМИД 2018220 . S2CID 39187918 .
- ^ Ваес В.Х., Рамос ЕС, Хамвейк С., ван Холстейн И., Блааубоер Б.Дж., Сейнен В., Верхаар Х.Дж., Херменс Дж.Л. (1997). «Твердофазная микроэкстракция как инструмент для определения коэффициентов распределения мембрана/вода и биодоступных концентраций в системах in vitro». Химические исследования в токсикологии . 10 (10): 1067–1072. дои : 10.1021/tx970109t . ПМИД 9348427 .
- ^ Мейер Л.А., Леермейкерс Ф.А., Ликлема Дж. (1999). «Моделирование самосогласованных полей сложных молекул с деталями объединенных атомов в неоднородных системах. Циклические и разветвленные чужеродные молекулы в мембранах димиристоилфосфатидилхолина». Журнал химической физики . 110 (13): 6560–6579. Бибкод : 1999ЖЧФ.110.6560М . дои : 10.1063/1.478562 .
- ^ Павел, Махмуд Ариф; Петерсен, Э. Николас; Ван, Хао; Лернер, Ричард А.; Хансен, Скотт Б. (16 июня 2020 г.). «Исследования о механизме общей анестезии» . Труды Национальной академии наук . 117 (24): 13757–13766. Бибкод : 2020PNAS..11713757P . дои : 10.1073/pnas.2004259117 . ПМК 7306821 . ПМИД 32467161 .
- ^ Карри, С; Либ, ВР; Фрэнкс, НП (15 мая 1990 г.). «Влияние общих анестетиков на бактериальный фермент люцифераза Vibrio harveyi: целевой участок анестетика с дифференциальной чувствительностью». Биохимия . 29 (19): 4641–52. дои : 10.1021/bi00471a020 . ПМИД 2372547 .
- ^ Jump up to: а б Миллер К.В., Патон В.Д., Смит Р.А., Смит Э.Б. (1973). «Реверсирование давления при общей анестезии и гипотеза критического объема». Молекулярная фармакология . 9 (2): 131–143. ПМИД 4711696 .
- ^ Маллинз Л.И. (1954). «Некоторые физические механизмы наркоза». Химические обзоры . 54 (2): 289–323. дои : 10.1021/cr60168a003 .
- ^ Труделл-младший; Паян, Д.Г.; Чин, Дж. Х.; Коэн, EN (1975). «Антагонистическое влияние ингаляционного анестетика и высокого давления на фазовую диаграмму смешанных бислоев дипальмитоил-димиристоилфосфатидилхолина» . Труды Национальной академии наук . 72 (1): 210–213. Бибкод : 1975ПНАС...72..210Т . дои : 10.1073/pnas.72.1.210 . ПМЦ 432272 . ПМИД 164016 .
- ^ Кендиг, Джей-Джей; Гроссман, Ю.; Макивер, М. Брюс (1988). «Реверсирование давления анестезии: синаптический механизм» . Британский журнал анестезии . 60 (7): 806–816. дои : 10.1093/бья/60.7.806 . ПМИД 2840107 .
- ^ Миллер К.В. (1985). «Характер места общего наркоза». Международное обозрение нейробиологии . 27 (1): 1–61. дои : 10.1016/S0074-7742(08)60555-3 . ISBN 9780123668271 . ПМИД 3910602 .
- ^ Симан, П. (1974). «Теория расширения мембран в анестезии: прямые доказательства с использованием этанола и высокоточного плотномера». Эксперименты . 30 (7): 759–760. дои : 10.1007/BF01924170 . ПМИД 4847658 . S2CID 25056954 .
- ^ Джайн, Махендра К.; Йен-Мин Ву, Нора; Рэй, Льюис В. (1975). «Вызванное лекарством фазовое изменение в бислое как возможный механизм действия препаратов, расширяющих мембрану». Природа . 255 (5508): 494–496. Бибкод : 1975Natur.255..494J . дои : 10.1038/255494a0 . ПМИД 1138201 . S2CID 2033461 .
- ^ Jump up to: а б Труделл-младший (1977). «Единая теория анестезии, основанная на латеральном разделении фаз в нервных оболочках» . Анестезиология . 46 (1): 5–10. дои : 10.1097/00000542-197701000-00003 . ПМИД 12686 . S2CID 24107213 .
- ^ Jump up to: а б с Павел, М.А.; Петерсен, Э.Н.; Ван, Х; Лернер, РА; Хансен, С.Б. (16 июня 2020 г.). «Исследования о механизме общей анестезии» . Труды Национальной академии наук Соединенных Штатов Америки . 117 (24): 13757–13766. Бибкод : 2020PNAS..11713757P . дои : 10.1073/pnas.2004259117 . ПМК 7306821 . ПМИД 32467161 .
- ^ Петерсен, Э.Н.; Павел, М.А.; Ван, Х; Хансен, С.Б. (1 января 2020 г.). «Нарушение пальмитат-опосредованной локализации; общий путь силовой и анестезирующей активации каналов TREK-1» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1862 (1): 183091. doi : 10.1016/j.bbamem.2019.183091 . ПМК 6907892 . ПМИД 31672538 .
- ^ Jump up to: а б Экенхофф Р.Г., Таннер Дж.В., Йоханссон Дж.С. (1999). «Стерические препятствия не требуются для отсечения н-алканола в растворимых белках». Молекулярная фармакология . 56 (2): 414–418. дои : 10.1124/моль.56.2.414 . ПМИД 10419562 .
- ^ Jump up to: а б с Кантор РС (1997). «Профиль латерального давления в мембранах: физический механизм общей анестезии». Биохимия . 36 (9): 2339–2344. дои : 10.1021/bi9627323 . ПМИД 9054538 .
- ^ Лернер, Ричард А. (1997). «Гипотеза об эндогенном аналоге общей анестезии» . Труды Национальной академии наук . 94 (25): 13375–13377. Бибкод : 1997PNAS...9413375L . дои : 10.1073/pnas.94.25.13375 . ПМЦ 33784 . ПМИД 9391028 .
- ^ Павел, Махмуд Ариф; Петерсен, Э. Николас; Ван, Хао; Лернер, Ричард А.; Хансен, Скотт Б. (2020). «Исследования о механизме общей анестезии» . Труды Национальной академии наук . 117 (24): 13757–13766. Бибкод : 2020PNAS..11713757P . дои : 10.1073/pnas.2004259117 . ПМК 7306821 . ПМИД 32467161 .
- ^ Фрэнкс, Николас П .; Либ, Уильям Р. (1984). «Действуют ли общие анестетики путем конкурентного связывания со специфическими рецепторами?». Природа . 310 (16): 599–601. Бибкод : 1984Natur.310..599F . дои : 10.1038/310599a0 . ПМИД 6462249 . S2CID 4350646 .
- ^ Фрэнкс Н.П., Дженкинс А., Конти Э., Либ В.Р., Брик П. (1998). «Структурная основа ингибирования люциферазы светлячков общим анестетиком» . Биофизический журнал . 75 (5): 2205–2211. Бибкод : 1998BpJ....75.2205F . дои : 10.1016/S0006-3495(98)77664-7 . ПМЦ 1299894 . ПМИД 9788915 .
- ^ ЛаБелла Ф.С., Штейн Д., Королева Дж. (1998). «Занятие субстратного кармана цитохрома P450 различными соединениями в концентрациях для общей анестезии». Европейский журнал фармакологии . 358 (2): 177–185. дои : 10.1016/S0014-2999(98)00596-2 . ПМИД 9808268 .
- ^ Фрэнкс, Николас П .; Либ, Уильям Р. (1985). «Картирование целевых участков общего анестетика обеспечивает молекулярную основу для эффектов отсечения». Природа . 316 (6026): 349–351. Бибкод : 1985Natur.316..349F . дои : 10.1038/316349a0 . ПМИД 4022125 . S2CID 4239192 .
- ^ Миллер, Кейт В. (1985). «Характер места общего наркоза». Международное обозрение нейробиологии . 27 : 1–61. дои : 10.1016/S0074-7742(08)60555-3 . ISBN 9780123668271 . ПМИД 3910602 .
- ^ Слейтер С.Дж., Кокс К.Дж., Ломбарди Дж.В., Хо С., Келли М.Б., Рубин Э., Стаббс, компакт-диск (1993). «Ингибирование протеинкиназы С спиртами и анестетиками». Природа . 364 (6432): 82–84. Бибкод : 1993Natur.364...82S . дои : 10.1038/364082a0 . ПМИД 8316305 . S2CID 4343565 .
- ^ Хеммингс-младший, ХК; Адамо, А.И. (1994). «Влияние галотана и пропофола на активацию очищенной протеинкиназы C мозга». Анестезиология . 81 (1): 147–155. дои : 10.1097/00000542-199409001-00886 . ПМИД 8042784 .
- ^ Фрэнкс, Николас П .; Либ, Уильям Р. (1994). «Молекулярные и клеточные механизмы общей анестезии». Природа . 367 (6464): 607–614. Бибкод : 1994Natur.367..607F . дои : 10.1038/367607a0 . ПМИД 7509043 . S2CID 4357493 .
- ^ Йоханссон Дж.С., Гибни Б.Р., Рабанал Ф., Редди К.С., Даттон П.Л. (1998). «Спроектированная полость в гидрофобном ядре пучка из четырех α-спиралей улучшает сродство связывания летучих анестетиков». Биохимия . 37 (5): 1421–1429. дои : 10.1021/bi9721290 . ПМИД 9477971 .
- ^ Цуй Т, Бондаренко В, Ма Д, Канлас С, Брэндон Н.Р., Йоханссон Дж.С., Сюй Ю, Тан П (2008). «Пучок четырех альфа-спиралей с разработанными карманами для связывания анестетика. Часть II: Влияние галотана на структуру и динамику» . Биофизический журнал . 94 (11): 4464–4472. Бибкод : 2008BpJ....94.4464C . дои : 10.1529/biophysj.107.117853 . ПМК 2480694 . ПМИД 18310239 .
- ^ Ма Д., Брэндон Н.Р., Цуй Т., Бондаренко В., Канлас С., Йоханссон Дж.С., Тан П., Сюй Ю (2008). «Пучок четырех альфа-спиралей с разработанными карманами для связывания анестетика. Часть I: Структурный и динамический анализ» . Биофизический журнал . 94 (11): 4454–4463. Бибкод : 2008BpJ....94.4454M . дои : 10.1529/biophysj.107.117838 . ПМК 2480675 . ПМИД 18310240 .
- ^ Лю Р., Лолл П.Дж., Экенхофф Р.Г. (2005). «Структурная основа высокоаффинного связывания летучих анестетиков в природном белке из четырехспирального пучка» . Журнал ФАСЭБ . 19 (6): 567–576. дои : 10.1096/fj.04-3171com . ПМИД 15791007 . S2CID 27832370 .
- ^ Тан П, Сюй Ю (2002). «Крупномасштабное молекулярно-динамическое моделирование воздействия общего анестетика на ионный канал в полностью гидратированной мембране: значение молекулярных механизмов общей анестезии» . Труды Национальной академии наук . 99 (25): 16035–16040. Бибкод : 2002PNAS...9916035T . дои : 10.1073/pnas.252522299 . ПМК 138560 . ПМИД 12438684 .
- ^ Канлас К.Г., Цуй Т, Ли Л, Сюй Ю, Тан П (2008). «Анестетическая модуляция динамики белка: результаты исследования ЯМР» . Журнал физической химии Б. 112 (45): 14312–14318. дои : 10.1021/jp805952w . ПМК 2669902 . ПМИД 18821786 .
- ^ Михик С.Дж., Йе Кью, Вик М.Дж., Колчин В.В., Красовский М.Д., Финн С.Е., Массия М.П., Валенсуэла К.Ф., Хэнсон К.К., Гринблатт Э.П., Харрис Р.А., Харрисон Н.Л. (1997). «Места действия алкоголя и летучих анестетиков на рецепторы ГАМК А и глицина». Природа . 389 (6649): 385–389. Бибкод : 1997Natur.389..385M . дои : 10.1038/38738 . ПМИД 9311780 . S2CID 4393717 .
- ^ Ким, Чон Джу; Гарпур, Анант; Тэн, Цзиньфэн; Чжуан, Юйсюань; Ховард, Ребекка Дж.; Чжу, Шаотун; Новиелло, Коллин М.; Уолш, Ричард М.; Линдал, Эрик; Хиббс, Райан Э. (2020). «Человеческий рецептор ГАМК А подтипа α1-β2-γ2 в комплексе с ГАМК плюс пропофол». РЦСБ ПДБ . дои : 10.2210/pdb6X3T/pdb . S2CID 225185057 .
- ^ Ким, Чон Джу; Гарпур, Анант; Тэн, Цзиньфэн; Чжуан, Юйсюань; Ховард, Ребекка Дж.; Чжу, Шаотун; Новиелло, Коллин М.; Уолш, Ричард М.; Линдал, Эрик; Хиббс, Райан Э. (2020). «Общие структурные механизмы общих анестетиков и бензодиазепинов» . Природа . 585 (7824): 303–308. дои : 10.1038/s41586-020-2654-5 . ПМЦ 7486282 . ПМИД 32879488 .
- ^ Саллард, Эрван; Летурнер, Дайан; Лежандр, Паскаль (2021). «Электрофизиология ионотропных ГАМК-рецепторов» . Клеточные и молекулярные науки о жизни . 78 (13): 5341–5370. дои : 10.1007/s00018-021-03846-2 . ПМЦ 8257536 . ПМИД 34061215 .
- ^ Да, Грейс М.С.; Чен, Цзы-Вэй; Эдж, Кристофер Дж.; Смит, Эдвард Х.; Дикинсон, Роберт; Хоэнестер, Эрхард; Таунсенд, Р. Рид; Фукс, Каролина; Зигхарт, Вернер; Эверс, Алекс С.; Фрэнкс, Николас П. (2013). «Сайт связывания пропофола на рецепторах ГАМК А млекопитающих , выявленный с помощью фотомечения» . Химическая биология природы . 9 (11): 715–720. дои : 10.1038/nchembio.1340 . ПМЦ 3951778 . ПМИД 24056400 .
- ^ Биеда, Марк К.; Макивер, М. Брюс (2004). «Основная роль тонической проводимости ГАМК А в анестезирующем подавлении внутренней возбудимости нейронов». Журнал нейрофизиологии . 92 (3): 1658–1667. дои : 10.1152/jn.00223.2004 . ПМИД 15140905 .
- ^ Кьяра Д.С., Досталова З., Джаякар С.С., Чжоу Х, Миллер К.В., Коэн Дж.Б. (2012). «Картирование сайтов связывания общего анестетика в человеческих рецепторах α1β3 γ-аминомасляной кислоты типа А с [3H]TDBzl-этомидатом, фотореактивным аналогом этомидата» . Биохимия . 51 (4): 836–47. дои : 10.1021/bi201772m . ПМЦ 3274767 . ПМИД 22243422 .
- ^ Дай, Шуйпин; Перуанский, Миша; Пирс, Роберт А. (2009). «Амнестические концентрации этомидата модулируют ГАМК А, медленное синаптическое торможение в гиппокампе» . Анестезиология . 111 (4): 766–773. дои : 10.1097/ALN.0b013e3181b4392d . ПМЦ 2797577 . ПМИД 19741493 .
- ^ Хентшке, Харальд; Шварц, Корнелиус; Антковяк, Бернд (2005). «Неокортекс является основной мишенью седативных концентраций летучих анестетиков: сильное снижение скорости срабатывания и усиление ингибирования, ГАМК А опосредованного рецептором ». Европейский журнал неврологии . 21 (1): 93–102. дои : 10.1111/j.1460-9568.2004.03843.x . ПМИД 15654846 . S2CID 12707025 .
- ^ Патель, Аманда Дж.; Оноре, Эрик; Лесаж, Флориан; Финк, Мишель; Роми, Жорж; Лаздунски, Мишель (1999). «Ингаляционные анестетики активируют двухпоровые фоновые K+-каналы». Природная неврология . 2 (5): 422–426. дои : 10.1038/8084 . ПМИД 10321245 . S2CID 23092576 .
- ^ Стейнберг, Э.А.; Уоффорд, Калифорния; Брикли, СГ; Фрэнкс, Николас П .; Висден, В. (2015). «Роль каналов K2P в анестезии и сне» . Архив Pflügers: Европейский журнал физиологии . 467 (5): 907–916. дои : 10.1007/s00424-014-1654-4 . ПМЦ 4428837 . ПМИД 25482669 .
- ^ Деномм, Николас; Халл, Джейкоб М.; Машур, Джордж А. (2019). «Роль потенциалзависимых натриевых каналов в механизме бессознательного состояния, вызванного эфиром» . Фармакологические обзоры . 71 (4): 450–466. дои : 10.1124/пр.118.016592 . ПМИД 31471460 . S2CID 201757964 .
- ^ Нау С., Стрихарц Г.Р. (2002). «Лекарственная хиральность в анестезии» . Анестезиология . 97 (2): 497–502. дои : 10.1097/00000542-200208000-00029 . ПМИД 12151942 . S2CID 2388540 .
- ^ Фрэнкс, Николас П .; Либ, Уильям Р. (1991). «Стереоспецифическое воздействие оптических изомеров ингаляционных анестетиков общего действия на нервные ионные каналы». Наука . 254 (5030): 427–430. Бибкод : 1991Sci...254..427F . дои : 10.1126/science.1925602 . ПМИД 1925602 .
- ^ Себекауэр, М; Амаро, М; Юркевич, П; Сарменто, MJ; Шахль, Р; Цвиклик, Л; Хоф, М. (12 декабря 2018 г.). «Мембранные липидные нанодомены». Химические обзоры . 118 (23): 11259–11297. doi : 10.1021/acs.chemrev.8b00322 . ПМИД 30362705 . S2CID 53096675 .
- ^ Сезгин Е; Левенталь, Я; Мэр, С; Эггелинг, К. (июнь 2017 г.). «Тайна мембранной организации: состав, регуляция и роль липидных рафтов» . Обзоры природы. Молекулярно-клеточная биология . 18 (6): 361–374. дои : 10.1038/номер.2017.16 . ПМК 5500228 . ПМИД 28356571 .
- ^ Кандель Л., Чортков Б.С., Зоннер Дж., Ластер М.Дж., Эгер Э.И. (1996). «Неанестетики могут подавлять обучение» . Анестезия и анальгезия . 82 (2): 321–326. дои : 10.1097/00000539-199602000-00019 . ПМИД 8561335 . S2CID 32518667 .
- ^ Коблин Д.Д., Чортков Б.С., Ластер М.Дж., Эгер Э.И. II, Хэлси М.Дж., Ионеску П. (1994). «Полигалогенированные и перфторированные соединения, не подчиняющиеся гипотезе Мейера-Овертона» . Анестезия и анальгезия . 79 (6): 1043–1048. дои : 10.1213/00000539-199412000-00004 . ПМИД 7978424 .
- ^ Фанг З., Соннер Дж., Ластер М.Дж., Ионеску П., Кандел Л., Коблин Д.Д., Эгер Э.И. II, Хэлси М.Дж. (1996). «Анестетические и противосудорожные свойства ароматических соединений и циклоалканов: значение для механизмов наркоза» . Анестезия и анальгезия . 83 (5): 1097–1104. дои : 10.1097/00000539-199611000-00035 . ПМИД 8895293 . S2CID 25929855 .
- ^ Тахери С., Ластер М.Дж., Лю Дж., Эгер Э.И. II, Хэлси М.Дж., Коблин Д.Д. (1993). «Анестезия н-алканами, не согласующаяся с гипотезой Мейера-Овертона: определение растворимости алканов в физиологическом растворе и различных липидах» . Анестезия и анальгезия . 77 (1): 7–11. дои : 10.1213/00000539-199307000-00003 . ПМИД 8317750 .
- ^ Эгер Э.И., Коблин Д.Д., Харрис Р.А., Кендиг Дж.Дж., Похорилле А., Хэлси М.Дж., Труделл Дж.Р. (1997). «Гипотеза: ингаляционные анестетики вызывают обездвиженность и амнезию с помощью разных механизмов в разных местах» . Анестезия и анальгезия . 84 (4): 915–918. дои : 10.1097/00000539-199704000-00039 . ПМИД 9085981 . S2CID 890662 .
- ^ Фрэнкс, Николас П .; Либ, Уильям Р. (1982). «Молекулярные механизмы общей анестезии». Природа . 300 (5892): 487–493. Бибкод : 1982Natur.300..487F . дои : 10.1038/300487a0 . ПМИД 6755267 . S2CID 4277388 .
- ^ Грей, Е; Карслейк, Дж; Мачта, Б.Б.; Витч, SL (17 декабря 2013 г.). «Жидкие общие анестетики снижают критические температуры в пузырьках плазматической мембраны» . Биофизический журнал . 105 (12): 2751–9. arXiv : 1309.2684 . Бибкод : 2013BpJ...105.2751G . дои : 10.1016/j.bpj.2013.11.005 . ПМЦ 3882514 . ПМИД 24359747 .
- ^ Прингл М.Дж., Браун К.Б., Миллер К.В. (1981). «Могут ли липидные теории анестезии объяснить снижение анестезирующей активности гомологического ряда спиртов?» . Молекулярная фармакология . 19 (1): 49–55. ПМИД 7207463 .
- ^ Лю Дж., Ластер М.Дж., Тахери С., Эгер Э.И., Коблин Д.Д., Хэлси М.Дж. (1993). «Есть ли ограничение по анестезирующей активности нормальных алканов?» . Анестезия и анальгезия . 77 (1): 12–18. дои : 10.1213/00000539-199307000-00004 . ПМИД 8317717 . S2CID 24811390 .
- ^ Jump up to: а б Мор Дж.Т., Гриббл Г.В., Лин С.С., Экенхофф Р.Г., Кантор Р.С. (2005). «Анестетическая эффективность двух новых синтетических многоатомных алканолов, длиннее порогового уровня н-алканола: доказательства двухслойного механизма анестезии?». Журнал медицинской химии . 48 (12): 4172–76. дои : 10.1021/jm049459k . ПМИД 15943489 .
- ^ Кантор РС (2001). «Нарушение правила Мейера-Овертона: прогнозируемые эффекты различной жесткости и межфазной активности на внутреннюю эффективность анестетиков» . Биофизический журнал . 80 (5): 2284–2297. Бибкод : 2001BpJ....80.2284C . дои : 10.1016/S0006-3495(01)76200-5 . ПМЦ 1301419 . ПМИД 11325730 .
- ^ Чунг, Х.В.; Петерсен, Э.Н.; Кабанос, К; Мерфи, КР; Павел, М.А.; Хансен, А.С.; Да, WW; Хансен, С.Б. (18 января 2019 г.). «Молекулярная мишень для ограничения длины алкогольной цепи» . Журнал молекулярной биологии . 431 (2): 196–209. дои : 10.1016/j.jmb.2018.11.028 . ПМК 6360937 . ПМИД 30529033 .