Рост клеток
| Часть серии о |
| Биология |
|---|
клеток означает увеличение общей массы клетки и , включая цитоплазмы , ядра Рост органелл . объем [1] Рост клеток происходит, когда общая скорость клеточного биосинтеза (производство биомолекул или анаболизм) превышает общую скорость клеточной деградации (разрушение биомолекул посредством протеасом , лизосом или аутофагии или катаболизма). [2] [3] [4]
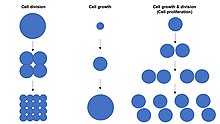
Рост клеток не следует путать с делением клеток или клеточным циклом , которые представляют собой отдельные процессы, которые могут происходить параллельно с ростом клеток в процессе пролиферации клеток , когда клетка, известная как материнская клетка, растет и делится с образованием двух дочерних клеток. . [1] Важно отметить, что рост и деление клеток также могут происходить независимо друг от друга. Во время раннего эмбрионального развития ( дробление зиготы происходят неоднократно без с образованием морулы и бластодермы ) клеточные деления роста клеток. И наоборот, некоторые клетки могут расти без деления клеток или без какого-либо прогресса клеточного цикла , например, рост нейронов во время поиска аксональных путей в развитии нервной системы .
В многоклеточных организмах рост тканей редко происходит исключительно за счет роста клеток без деления клеток , но чаще всего происходит за счет пролиферации клеток . [1] Это связано с тем, что одна клетка, имеющая только одну копию генома в клеточном ядре, может осуществлять биосинтез и, таким образом, подвергаться клеточному росту лишь вдвое медленнее, чем две клетки. Следовательно, две клетки растут (набирают массу) в два раза быстрее, чем одна клетка, а четыре клетки растут в 4 раза быстрее, чем одна клетка. Этот принцип приводит к экспоненциальному увеличению скорости роста тканей (накоплению массы) во время пролиферации клеток за счет экспоненциального увеличения количества клеток.
Размер клеток зависит как от роста клеток, так и от деления клеток : непропорциональное увеличение скорости роста клеток приводит к образованию более крупных клеток, а непропорциональное увеличение скорости деления клеток приводит к образованию множества более мелких клеток. Пролиферация клеток обычно включает сбалансированную скорость роста и деления клеток , которая поддерживает примерно постоянный размер клеток в экспоненциально пролиферирующей популяции клеток.
Некоторые специальные клетки могут вырасти до очень больших размеров посредством необычного клеточного цикла эндорепликации , в котором геном реплицируется во время S-фазы , но не происходит последующего митоза ( М-фаза ) или деления клеток ( цитокинез ). Эти крупные эндореплицирующиеся клетки имеют множество копий генома , поэтому являются высокополиплоидными .
Ооциты могут представлять собой необычно крупные клетки у видов, у которых эмбриональное развитие происходит вне тела матери в яйцеклетке, отложенной снаружи. Больших размеров некоторых яиц можно достичь либо закачиванием цитозольных компонентов из соседних клеток через цитоплазматические мостики, называемые кольцевыми каналами ( дрозофила ), либо за счет интернализации гранул хранения питательных веществ (гранул желтка) путем эндоцитоза ( лягушки ).
Механизмы контроля роста клеток
[ редактировать ]Клетки могут расти за счет увеличения общей скорости клеточного биосинтеза , так что производство биомолекул превышает общую скорость клеточной деградации биомолекул посредством протеасомы , лизосомы или аутофагии .
Биосинтез биомолекул , инициируется экспрессией генов кодирующих РНК и/или белки , в том числе ферментов , катализирующих синтез липидов и углеводов .
Отдельные гены обычно экспрессируются посредством транскрипции в информационную РНК (мРНК) и трансляции в белки , причем экспрессия каждого гена происходит на различных уровнях в зависимости от типа клеток (в ответ на регуляторные сети генов ).
Чтобы стимулировать рост клеток, можно увеличить глобальную скорость экспрессии генов за счет увеличения общей скорости транскрипции РНК -полимеразой II (для активных генов) или общей скорости мРНК трансляции в белок за счет увеличения количества рибосом и тРНК , биогенез которых зависит от РНК-полимеразы I и РНК-полимеразы III . Фактор Myc транскрипции является примером регуляторного белка, который может индуцировать общую активность РНК-полимеразы I , РНК-полимеразы II и РНК-полимеразы III, чтобы управлять глобальной транскрипцией и трансляцией и, следовательно, ростом клеток.
Кроме того, активность отдельных рибосом можно увеличить, чтобы повысить общую эффективность мРНК трансляции посредством регуляции факторов инициации трансляции, включая комплекс «фактор инициации элонгации трансляции 4E» ( eIF4E ), который связывается и блокирует 5'-конец мРНК. мРНК . Белок TOR , входящий в состав комплекса TORC1 , является важным регулятором инициации трансляции , а также биогенеза рибосом . [5] TOR представляет собой серин/треониновую киназу , которая может напрямую фосфорилировать и инактивировать общий ингибитор eIF4E , называемый 4E-связывающим белком (4E-BP) , для повышения эффективности трансляции. TOR также непосредственно фосфорилирует и активирует рибосомальную протеин S6-киназу ( S6K ), которая способствует биогенезу рибосом .
Чтобы ингибировать рост клеток, можно снизить глобальную скорость экспрессии генов или глобальную скорость биомолекулярной увеличить деградации за счет увеличения скорости аутофагии . TOR обычно напрямую ингибирует функцию аутофагию индуцирующей киназы Atg1/ULK1, . Таким образом, снижение активности TOR снижает глобальную скорость трансляции и увеличивает степень аутофагии , снижая рост клеток.
Регуляция роста клеток у животных
[ редактировать ]Многие из сигнальных молекул, которые контролируют клеточный рост, называются факторами роста , многие из которых индуцируют передачу сигнала по пути PI3K/AKT/mTOR , который включает вышестоящую липидкиназу PI3K и нижележащую серин/треониновую протеинкиназу Akt , которая способна активировать другую протеинкиназу TOR , которая способствует трансляции и ингибирует аутофагию , стимулирующую рост клеток.
Доступность питательных веществ влияет на выработку факторов роста семейства инсулина / ИФР-1 , которые циркулируют в организме животных в виде гормонов, активируя путь PI3K/AKT/mTOR в клетках и стимулируя активность TOR , так что, когда животные сыты, они будут быстро расти, а когда они не смогут получать достаточное количество питательных веществ и замедлят темпы своего роста. Недавно также было продемонстрировано, что клеточный метаболизм бикарбоната, который отвечает за рост клеток, может регулироваться передачей сигналов mTORC1. [6]
Кроме того, доступность аминокислот для отдельных клеток также напрямую способствует активности TOR , хотя этот способ регуляции более важен для одноклеточных организмов, чем для многоклеточных , таких как животные, которые всегда поддерживают избыток аминокислот в кровообращении.
Одна из спорных теорий предполагает, что многие различные клетки млекопитающих претерпевают изменения, зависящие от размера, в течение клеточного цикла. Эти переходы контролируются циклинзависимой киназой Cdk1. [7] Хотя белки, контролирующие Cdk1, хорошо изучены, их связь с механизмами контроля размера клеток остается неясной.
Постулируемая модель контроля размера млекопитающих рассматривает массу как движущую силу клеточного цикла. Клетка не может вырасти до аномально большого размера, потому что при определенном размере клетки или клеточной массе инициируется S-фаза. Фаза S запускает последовательность событий, приводящих к митозу и цитокинезу. Клетка не может стать слишком маленькой, потому что более поздние события клеточного цикла, такие как S, G2 и M, задерживаются до тех пор, пока масса не увеличится достаточно, чтобы начать S-фазу. [8]
Популяции клеток
[ редактировать ]Популяции клеток проходят особый тип экспоненциального роста, называемый удвоением или пролиферацией клеток . Таким образом, каждое поколение клеток должно быть вдвое больше, чем предыдущее поколение. Однако количество поколений дает только максимальную цифру, поскольку не все клетки выживают в каждом поколении. Клетки могут размножаться на стадии митоза, когда они удваиваются и делятся на две генетически равные клетки.
Размер ячейки
[ редактировать ]Размер клеток у разных организмов сильно различается: некоторые водоросли, такие как Caulerpa Taxifolia, представляют собой одну клетку длиной несколько метров. [9] Растительные клетки намного крупнее клеток животных, а длина протистов, таких как Paramecium , может достигать 330 мкм, тогда как типичная человеческая клетка может иметь длину 10 мкм. Как эти клетки «решают», насколько большими они должны быть перед делением, остается открытым вопросом. Известно, что отчасти ответственными за это являются химические градиенты, и предполагается, что цитоскелета в этом участвует обнаружение механического стресса структурами . Для работы над этой темой обычно требуется организм, клеточный цикл которого хорошо охарактеризован.
Регулирование размера дрожжевых клеток
[ редактировать ]Взаимосвязь между размером клеток и их делением широко изучалась на дрожжах . Для некоторых клеток существует механизм, по которому деление клеток не начинается до тех пор, пока клетка не достигнет определенного размера. Если поступление питательных веществ ограничено (после времени t = 2 на диаграмме ниже) и скорость увеличения размера клеток замедляется, период времени между делениями клеток увеличивается. [10] Были выделены мутанты размера дрожжевых клеток, которые начинают деление клеток до достижения нормального/регулярного размера ( крошечные мутанты). [11]

Wee1 Белок представляет собой тирозинкиназу , которая в норме фосфорилирует белок, регулирующий клеточный цикл Cdc2 (гомолог CDK1 у человека), циклин-зависимую киназу, по остатку тирозина. Cdc2 обеспечивает вступление в митоз путем фосфорилирования широкого спектра мишеней. Эта ковалентная модификация молекулярной структуры Cdc2 ингибирует ферментативную активность Cdc2 и предотвращает деление клеток. Wee1 сохраняет Cdc2 неактивным во время раннего G2 , когда клетки еще малы. Когда клетки достигают достаточного размера во время G2, фосфатаза Cdc25 удаляет ингибирующее фосфорилирование и, таким образом, активирует Cdc2, обеспечивая вход в митоз. Баланс активности Wee1 и Cdc25 с изменениями размера клеток координируется системой контроля входа в митоз. На мутантах Wee1, клетках с ослабленной активностью Wee1, было показано, что Cdc2 становится активным, когда клетка становится меньше. Таким образом, митоз происходит до того, как дрожжи достигнут нормального размера. Это предполагает, что деление клеток может частично регулироваться за счет разбавления белка Wee1 в клетках по мере их роста.
Связывание Cdr2 с Wee1
[ редактировать ]Протеинкиназа Cdr2 (которая отрицательно регулирует Wee1) и родственная Cdr2 киназа Cdr1 (которая непосредственно фосфорилирует и ингибирует Wee1 in vitro ) [12] локализуются в полосе корковых узлов в середине интерфазных клеток. После вступления в митоз факторы цитокинеза, такие как миозин II , рекрутируются в аналогичные узлы; эти узлы в конечном итоге конденсируются, образуя цитокинетическое кольцо. [13] Было обнаружено , что ранее неохарактеризованный белок Blt1 колокализуется с Cdr2 в медиальных интерфазных узлах. Клетки, нокаутные по Blt1, имели увеличенную длину при делении, что согласуется с задержкой вступления в митоз. Это открытие связывает физическое местоположение, группу корковых узлов, с факторами, которые, как было показано, напрямую регулируют вход в митозы, а именно Cdr1, Cdr2 и Blt1.
Дальнейшие эксперименты с белками, меченными GFP , и мутантными белками показывают, что медиальные кортикальные узлы формируются посредством упорядоченной, Cdr2-зависимой сборки множества взаимодействующих белков во время интерфазы. Cdr2 находится на вершине этой иерархии и работает выше Cdr1 и Blt1. [14] Митозу способствует негативная регуляция Wee1 с помощью Cdr2. Также было показано, что Cdr2 рекрутирует Wee1 в медиальный кортикальный узел. Механизм этой вербовки еще предстоит выяснить. Мутант киназы Cdr2, который способен правильно локализоваться, несмотря на потерю функции фосфорилирования, нарушает рекрутирование Wee1 в медиальную кору и задерживает вступление в митоз. Т.о., Wee1 локализуется с помощью своей ингибирующей сети, что демонстрирует, что митоз контролируется посредством Cdr2-зависимой негативной регуляции Wee1 в медиальных кортикальных узлах. [14]
Факторы полярности клеток
[ редактировать ]Факторы полярности клеток, расположенные на кончиках клеток, обеспечивают пространственные сигналы, ограничивающие распространение Cdr2 в середину клетки. У делящихся дрожжей Schizosaccharomyces pombe ( S. Pombe ) клетки делятся на определенный, воспроизводимый размер во время митоза из-за регулируемой активности Cdk1. [15] Протеинкиназа клеточной полярности Pom1 , член семейства киназ, регулируемых тирозин-фосфорилированием двойной специфичности (DYRK), локализуется на концах клеток. В клетках с нокаутом Pom1 Cdr2 больше не ограничивался серединой клетки, а был виден диффузно через половину клетки. Из этих данных становится очевидным, что Pom1 обеспечивает ингибирующие сигналы, которые удерживают Cdr2 в середине клетки. Далее было показано, что Pom1-зависимые сигналы приводят к фосфорилированию Cdr2. Также было показано, что клетки, нокаутные Pom1, делятся с меньшим размером, чем клетки дикого типа, что указывает на преждевременное вступление в митоз. [14]
Pom1 формирует полярные градиенты, достигающие максимума на концах клеток, что указывает на прямую связь между факторами контроля размера и конкретным физическим местоположением в клетке. [16] По мере увеличения размера ячейки градиент Pom1 увеличивается. Когда клетки маленькие, Pom1 диффузно распространяется по телу клетки. По мере увеличения размера клетки концентрация Pom1 уменьшается в середине и концентрируется на концах клетки. Маленькие клетки в раннем G2, которые содержат достаточные уровни Pom1 во всей клетке, имеют неактивный Cdr2 и не могут вступить в митоз. Только когда клетки перерастают в позднюю G2, когда Pom1 ограничивается концами клеток, Cdr2 в медиальных кортикальных узлах активируется и становится способным начать ингибирование Wee1. Это открытие показывает, как размер клеток играет прямую роль в регуляции начала митоза. В этой модели Pom1 действует как молекулярное звено между ростом клеток и входом в митоз через путь Cdr2-Cdr1-Wee1-Cdk1. [14] Полярный градиент Pom1 успешно передает информацию о размере и геометрии клеток в регуляторную систему Cdk1. Благодаря этому градиенту клетка гарантирует, что она достигла определенного размера, достаточного для вступления в митоз.
Другие экспериментальные системы для изучения регуляции размера клеток
[ редактировать ]Одним из распространенных способов получения очень крупных клеток является слияние клеток с образованием синцития . Например, очень длинные (несколько дюймов) клетки скелетных мышц образуются путем слияния тысяч миоцитов . Генетические исследования плодовой мушки Drosophila выявили несколько генов, необходимых для образования многоядерных мышечных клеток путем слияния миобластов . [17] Некоторые из ключевых белков важны для клеточной адгезии между миоцитами, а некоторые участвуют в зависимой от адгезии передаче сигнала от клетки к клетке , что обеспечивает каскад событий слияния клеток.
Увеличение размеров растительных клеток осложняется тем фактом, что почти все растительные клетки находятся внутри твердой клеточной стенки . Под влиянием определенных растительных гормонов клеточная стенка может реконструироваться, что позволяет увеличить размер клеток, что важно для роста некоторых тканей растения.
Большинство одноклеточных организмов имеют микроскопические размеры, но есть некоторые гигантские бактерии и простейшие , видимые невооруженным глазом. (См. Таблицу размеров клеток — Плотные популяции гигантской серной бактерии в отложениях шельфа Намибии. [18] — Крупные протисты рода Chaos , тесно связанные с родом Amoeba .)
У палочковидных бактерий E. coli , Caulobacter crescentus и B. subtilis размер клеток контролируется простыми механизмами, при которых деление клеток происходит после добавления постоянного объема с момента предыдущего деления. [19] [20] Постоянно увеличиваясь на одну и ту же величину, клетки, рождающиеся меньше или крупнее среднего, естественным образом сходятся к среднему размеру, эквивалентному количеству, добавляемому в каждом поколении.
Деление клеток
[ редактировать ]Размножение клеток бесполое . Для большинства компонентов клетки рост представляет собой устойчивый, непрерывный процесс, прерывающийся лишь ненадолго в фазе М, когда ядро, а затем клетка делятся на две части.
Процесс деления клеток, называемый клеточным циклом , состоит из четырех основных частей, называемых фазами. Первая часть, называемая фазой G1, отмечена синтезом различных ферментов , необходимых для репликации ДНК.Вторая часть клеточного цикла — это фаза S, когда репликация ДНК производит два идентичных набора хромосом . Третья часть — это фаза G2, в которой происходит значительный синтез белка , в основном связанный с производством микротрубочек , которые необходимы в процессе деления, называемом митозом .Четвертая фаза, М-фаза, состоит из деления ядра ( кариокинез ) и деления цитоплазмы ( цитокинез ), сопровождающихся образованием новой клеточной мембраны . Это физическое деление материнских и дочерних клеток. Фаза М разделена на несколько отдельных фаз, последовательно известных как профаза , прометафаза , метафаза , анафаза и телофаза , приводящие к цитокинезу.
более сложное, Деление клеток у эукариот чем у других организмов. Прокариотические клетки, такие как бактериальные клетки, размножаются путем бинарного деления — процесса, который включает репликацию ДНК, сегрегацию хромосом и цитокинез. Деление эукариотических клеток включает либо митоз , либо более сложный процесс, называемый мейозом . Митоз и мейоз иногда называют двумя процессами деления ядра . Бинарное деление похоже на размножение клеток эукариот, которое включает митоз. Оба приводят к образованию двух дочерних клеток с тем же количеством хромосом, что и родительская клетка. Мейоз используется для особого процесса размножения клеток диплоидных организмов. Он производит четыре специальные дочерние клетки ( гаметы ), которые содержат половину нормального клеточного количества ДНК. Затем мужская женская и зиготу гамета могут объединиться, чтобы произвести — клетку, которая снова имеет нормальное количество хромосом.
Оставшаяся часть статьи представляет собой сравнение основных особенностей трех типов размножения клеток, которые включают бинарное деление, митоз или мейоз. На диаграмме ниже показаны сходства и различия этих трех типов размножения клеток.

Сравнение трех типов деления клеток
[ редактировать ]Содержимое ДНК клетки дублируется в начале процесса воспроизводства клеток. До репликации ДНК содержание ДНК в клетке можно представить как количество Z (клетка имеет Z-хромосомы). После процесса репликации ДНК количество ДНК в клетке составляет 2Z (умножение: 2 x Z = 2Z). Во время бинарного деления и митоза дублированное содержимое ДНК воспроизводящей родительской клетки разделяется на две равные половины, которым суждено оказаться в двух дочерних клетках. Заключительной частью процесса воспроизводства клеток является деление клеток , когда дочерние клетки физически отделяются от родительской клетки. Во время мейоза происходят два этапа деления клеток, которые вместе производят четыре дочерние клетки.
После завершения бинарного деления или размножения клеток с участием митоза каждая дочерняя клетка имеет такое же количество ДНК (Z), как и родительская клетка до того, как она реплицировала свою ДНК. Эти два типа размножения клеток дали две дочерние клетки, имеющие такое же количество хромосом, что и родительская клетка. Хромосомы дублируются перед делением клеток при формировании новых клеток кожи для размножения. После мейотического размножения клеток четыре дочерние клетки имеют вдвое меньше хромосом, чем исходная родительская клетка. Это гаплоидное количество ДНК, часто обозначаемое как N. Мейоз используется диплоидными организмами для производства гаплоидных гамет. В диплоидном организме, таком как человеческий организм, большинство клеток тела имеют диплоидное количество ДНК, 2N. Используя эти обозначения для подсчета хромосом, мы говорим, что соматические клетки человека имеют 46 хромосом (2N = 46), а сперматозоиды и яйцеклетки человека имеют 23 хромосомы (N = 23). У человека есть 23 различных типа хромосом, 22 аутосомы и особая категория хромосом. половые хромосомы . Есть две разные половые хромосомы: Х-хромосома и Y-хромосома. Диплоидная человеческая клетка имеет 23 хромосомы от отца этого человека и 23 от матери. То есть в вашем организме есть две копии человеческой хромосомы номер 2, по одной от каждого из ваших родителей.

Сразу после репликации ДНК человеческая клетка будет иметь 46 «двойных хромосом». В каждой двойной хромосоме есть две копии молекулы ДНК этой хромосомы. Во время митоза двойные хромосомы расщепляются, образуя 92 «одиночные хромосомы», половина из которых переходит в каждую дочернюю клетку. Во время мейоза происходят два этапа разделения хромосом, которые гарантируют, что каждая из четырех дочерних клеток получает по одной копии каждого из 23 типов хромосом.
Половое размножение
[ редактировать ]Хотя размножение клеток с использованием митоза может воспроизводить эукариотические клетки, эукариоты беспокоятся о более сложном процессе мейоза, поскольку половое размножение , такое как мейоз, дает селективное преимущество . Обратите внимание, что когда начинается мейоз, две копии сестринских хроматид номер 2 примыкают друг к другу. В это время могут произойти события генетической рекомбинации . Информация из ДНК хромосомы 2, полученная от одного родителя (красный), будет передана молекуле ДНК хромосомы 2, полученной от другого родителя (зеленый). Обратите внимание, что при митозе две копии хромосомы номер 2 не взаимодействуют. Рекомбинация генетической информации между гомологичными хромосомами во время мейоза — это процесс восстановления повреждений ДНК . Этот процесс также может создавать новые комбинации генов, некоторые из которых могут быть адаптивно полезными и влиять на ход эволюции. Однако у организмов с более чем одним набором хромосом на основной стадии жизненного цикла пол также может давать преимущество, поскольку при случайном спаривании он производит гомозиготы и гетерозиготы по соотношению Харди-Вайнберга .
расстройства
[ редактировать ]На клеточном уровне может возникнуть ряд нарушений роста, которые, следовательно, лежат в основе большей части последующего течения рака , при котором группа клеток демонстрирует неконтролируемый рост и деление, выходящее за нормальные пределы, инвазию (вторжение и разрушение соседних тканей), а иногда и метастазы (распространяются в другие части тела через лимфу или кровь). Некоторые ключевые детерминанты роста клеток, такие как плоидность и регуляция клеточного метаболизма , обычно нарушаются в опухолях . [21] Таким образом, гетерогенный рост клеток и плеоморфизм являются одними из самых ранних признаков прогрессирования рака . [22] [23] Несмотря на распространенность плеоморфизма в патологии человека, его роль в прогрессировании заболевания неясна. В эпителиальных тканях неправильная регуляция размера клеток может вызывать дефекты упаковки и рассеивать аберрантные клетки. [24] Но последствия атипичного роста клеток в других тканях животных неизвестны.
Методы измерения
[ редактировать ]Рост клеток можно обнаружить различными методами.Рост размера клеток можно визуализировать с помощью микроскопии с использованием подходящих красителей. Но увеличение количества клеток обычно более существенное. Его можно измерить путем ручного подсчета клеток под микроскопом, используя метод исключения красителя (т.е. трипановый синий ) для подсчета только жизнеспособных клеток. Менее привередливые и масштабируемые методы включают использование цитометров , в то время как проточная цитометрия позволяет комбинировать подсчет клеток («события») с другими конкретными параметрами: флуоресцентные зонды для мембран, цитоплазмы или ядер позволяют различать мертвые/жизнеспособные клетки, типы клеток, дифференцировку клеток, экспрессия биомаркера , такого как Ki67 . Общая масса клетки, которая включает массу всех ее компонентов, включая содержание воды, представляет собой динамическую величину, и ее можно измерять в режиме реального времени и отслеживать в течение часов или даже дней с помощью инерционных пикобалансов. [25] [26] Плавучую массу клетки, которая соответствует общей массе клетки за вычетом массы жидкости, которую она вытесняет, можно измерить с помощью подвешенных микроканальных резонаторов. [27]
Помимо увеличения количества клеток, можно оценить рост метаболической активности, то есть CFDA и кальцеин -АМ измеряют (флуориметрически) не только функциональность мембраны (удержание красителя), но и функциональность цитоплазматических ферментов (эстераз). . Анализы МТТ (колориметрические) и анализы резазурина (флуориметрические) измеряют окислительно-восстановительный потенциал митохондрий.
Все эти анализы могут хорошо или плохо коррелировать, в зависимости от условий роста клеток и желаемых аспектов (активность, пролиферация). Задача еще более усложняется с популяциями разных клеток, а также при сочетании помех клеточному росту или токсичности .
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с Конлон, Ян; Рафф, Мартин (1999). «Контроль размера в развитии животных» . Клетка . 96 (2): 235–244. дои : 10.1016/S0092-8674(00)80563-2 . ISSN 0092-8674 . ПМИД 9988218 . S2CID 15738174 .
- ^ Гревал, Савраж С; Эдгар, Брюс А. (2003). «Контроль деления клеток у дрожжей и животных: имеет ли значение размер?» . Журнал биологии . 2 (1): 5. дои : 10.1186/1475-4924-2-5 . ISSN 1475-4924 . ПМК 156596 . ПМИД 12733996 .
- ^ Нойфельд, Томас П; де ла Крус, Аида Флор А; Джонстон, Лаура А; Эдгар, Брюс А. (1998). «Координация роста и деления клеток крыла дрозофилы» . Клетка . 93 (7): 1183–1193. дои : 10.1016/S0092-8674(00)81462-2 . ISSN 0092-8674 . ПМИД 9657151 . S2CID 14608744 .
- ^ Томпсон, Барри Дж. (2010). «Контроль развития роста и деления клеток у дрозофилы». Современное мнение в области клеточной биологии . 22 (6): 788–794. дои : 10.1016/j.ceb.2010.08.018 . ПМИД 20833011 .
- ^ Хафен, Э. (2004). «Взаимодействие между фактором роста и передачей сигналов питательных веществ: уроки TOR дрозофилы». ТОР . Актуальные темы микробиологии и иммунологии. Том. 279. стр. 153–167. дои : 10.1007/978-3-642-18930-2_10 . ISBN 978-3-642-62360-8 . ISSN 0070-217X . ПМИД 14560957 .
- ^ Али Э, Липонска А, О'Хара Б, Амичи Д, Торно М, Гао П, Асара Дж, Яп МН Ф, Мендилло М, Бен-Сахра I (июнь 2022 г.). «Ось mTORC1-SLC4A7 стимулирует импорт бикарбоната для усиления синтеза нуклеотидов de novo» . Молекулярная клетка . 82 (1): 3284–3298.e7. doi : 10.1016/j.molcel.2022.06.008 . ПМЦ 9444906 . ПМИД 35772404 .
- ^ Митчисон Дж. М. (2003). «Рост в течение клеточного цикла». Межд. Преподобный Цитол . Международный обзор цитологии. 226 : 165–258. дои : 10.1016/S0074-7696(03)01004-0 . ISBN 978-0-12-364630-9 . ПМИД 12921238 .
- ^ Купер, Стивен (2004). «Контроль и поддержание размера клеток млекопитающих» . Клеточная биология BMC . 5 (1): 35. дои : 10.1186/1471-2121-5-35 . ПМК 524481 . ПМИД 15456512 .
- ^ Пеплоу, Марк (23 марта 2005 г.). «Водоросли создают клей для восстановления повреждений клеток» . Nature.com . Проверено 4 июля 2016 г.
- ^ Славов Н.; Ботштейн Д. (июнь 2011 г.). «Связь между реакцией скорости роста, метаболическим циклом и циклом деления клеток у дрожжей» . Молекулярная биология клетки . 22 (12): 1997–2009. дои : 10.1091/mbc.E11-02-0132 . ПМК 3113766 . ПМИД 21525243 .
- ^ Wee1 Мутанты S. pombe имеют небольшой размер клеток , а гомологичные белки у человека также регулируют вступление клеток в митоз ; в Лодиш Х.Ф., Берк А., Зипурский Л.С., Мацудайра П. и др., ред. (2000). Молекулярно-клеточная биология (4-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3136-8 .
- ^ Ву Л, Рассел П. (июнь 1993 г.). «Киназа Nim1 способствует митозу, инактивируя тирозинкиназу Wee1». Природа . 363 (6431): 738–41. Бибкод : 1993Natur.363..738W . дои : 10.1038/363738a0 . ПМИД 8515818 . S2CID 4320080 .
- ^ Ву Дж.К., Кун Дж.Р., Ковар Д.Р., Поллард Т.Д. (ноябрь 2003 г.). «Пространственный и временной путь сборки и сужения сократительного кольца при цитокинезе делящихся дрожжей» . Дев. Клетка . 5 (5): 723–34. дои : 10.1016/S1534-5807(03)00324-1 . ПМИД 14602073 .
- ^ Jump up to: а б с д Мозли Дж. Б., Майе А., Паолетти А., медсестра П. (июнь 2009 г.). «Пространственный градиент координирует размер клеток и вступление в митоз у делящихся дрожжей». Природа . 459 (7248): 857–60. Бибкод : 2009Natur.459..857M . дои : 10.1038/nature08074 . ПМИД 19474789 . S2CID 4330336 .
- ^ Рупес I (сентябрь 2002 г.). «Проверка размера клеток дрожжей». Тенденции Жене . 18 (9): 479–85. дои : 10.1016/S0168-9525(02)02745-2 . ПМИД 12175809 .
- ^ Падте Н.Н., Мартин С.Г., Ховард М., Чанг Ф. (декабрь 2006 г.). «Фактор конца клетки pom1p ингибирует Mid1p в спецификации плоскости деления клеток у делящихся дрожжей» . Курс. Биол . 16 (24): 2480–7. дои : 10.1016/j.cub.2006.11.024 . ПМИД 17140794 .
- ^ Менон С.Д., Осман З., Ченчилль К., Чиа В. (июнь 2005 г.). «Петля положительной обратной связи между камешками Dumbfounded и Rolling приводит к увеличению мышечных трубок у дрозофилы» . Дж. Клеточная Биол . 169 (6): 909–20. дои : 10.1083/jcb.200501126 . ПМК 2171639 . ПМИД 15955848 .
- ^ Шульц Х.Н., Бринкхофф Т., Фердельман Т.Г., Марине М.Х., Теске А., Йоргенсен Б.Б. (апрель 1999 г.). «Плотные популяции гигантской серной бактерии в отложениях шельфа Намибии». Наука . 284 (5413): 493–5. Бибкод : 1999Sci...284..493S . дои : 10.1126/science.284.5413.493 . ПМИД 10205058 . S2CID 32571118 .
- ^ Тахери-Араги, С; Брэдд, С; Саулс, Джей Ти; Хилл, Северная Каролина; Левин, Пенсильвания; Паулссон, Дж; Вергассола, М; Июнь, С (февраль 2015 г.). «Контроль размера клеток и гомеостаз у бактерий» . Современная биология . 25 (3): 385–391. дои : 10.1016/j.cub.2014.12.009 . ПМЦ 4323405 . ПМИД 25544609 .
- ^ Кампос, М; Суровцев И.В.; Като, С; Пайнтдахи, А; Бельтран, Б; Эбмайер, SE; Джейкобс-Вагнер, C (декабрь 2014 г.). «Постоянное увеличение размера обеспечивает гомеостаз размера бактериальных клеток» . Клетка . 159 (6): 1433–1446. дои : 10.1016/j.cell.2014.11.022 . ПМЦ 4258233 . ПМИД 25480302 .
- ^ Шмоллер, Курт М.; Скотхайм, Ян М. (декабрь 2015 г.). «Биосинтетическая основа контроля размера клеток» . Тенденции клеточной биологии . 25 (12): 793–802. дои : 10.1016/j.tcb.2015.10.006 . ПМК 6773270 . ПМИД 26573465 .
- ^ Трэвис, штат Вашингтон; Брамбилла, Б.; Берк, AP; Маркс, А.; Николсон, АГ (2015). Классификация ВОЗ опухолей легких, плевры, тимуса и сердца . Лион: Международное агентство по исследованию рака. ISBN 978-92-832-2436-5 .
- ^ Эль-Наггар, АК; Чан, JCK; Грандис, младший; Таката, Т.; Слотвег, Пи Джей (23 января 2017 г.). Классификация ВОЗ опухолей головы и шеи . Лион: Международное агентство по исследованию рака (также известное как Adman). ISBN 978-92-832-2438-9 . Архивировано из оригинала 31 октября 2019 г. Проверено 31 октября 2019 г.
- ^ Раманатан, Субраманиан П.; Крайнц, Матей; Гибсон, Мэтью К. (октябрь 2019 г.). «Плеоморфизм клеточного размера приводит к аберрантному распространению клонов в пролиферирующем эпителии» . Развивающая клетка . 51 (1): 49–61.e4. дои : 10.1016/j.devcel.2019.08.005 . ПМК 6903429 . ПМИД 31495693 .
- ^ Мартинес-Мартин, Давид; Флешнер, Готхольд; Гауб, Бенджамин; Мартин, Саша; Ньютон, Ричард; Берли, Корина; Мерсер, Джейсон; Гербер, Кристофер; Мюллер, Дэниел Дж. (октябрь 2017 г.). «Инерционный пикобаланс выявляет быстрые колебания массы в клетках млекопитающих» . Природа . 550 (7677): 500-505. дои : 10.1038/nature24288 . ISSN 1476-4687 . ПМИД 29072271 .
- ^ Куни, Андреас П.; Танудж Сапра, К.; Мартинес-Мартин, Дэвид; Флешнер, Готхольд; Адамс, Джонатан Д.; Мартин, Саша; Гербер, Кристоф; Рудольф, Фабиан; Мюллер, Дэниел Дж. (22 июня 2022 г.). «Измерения массы одиночных почкующихся дрожжей с высоким разрешением позволяют выявить сегменты линейного роста» . Природные коммуникации . 13 (1): 3483. doi : 10.1038/s41467-022-30781-y . ISSN 2041-1723 . ПМЦ 9217925 . ПМИД 35732645 .
- ^ Бург, Томас П.; Годен, Мишель; Кнудсен, Скотт М.; Шен, Вэньцзян; Карлсон, Грег; Фостер, Джон С.; Бэбкок, Кен; Маналис, Скотт Р. (апрель 2007 г.). «Взвешивание биомолекул, одиночных клеток и одиночных наночастиц в жидкости» . Природа . 446 (7139): 1066–1069. дои : 10.1038/nature05741 . hdl : 11858/00-001M-0000-0014-9C58-F . ISSN 1476-4687 . ПМИД 17460669 .
Книги
[ редактировать ]- Морган, Дэвид О. (2007). Клеточный цикл: принципы управления . Лондон: Сандерленд, Массачусетс, ISBN . 978-0-9539181-2-6 .
Внешние ссылки
[ редактировать ]- Сравнение поколенческой и экспоненциальной моделей роста популяции клеток
- Локальный рост в массиве дисков Демонстрационный проект Wolfram