Наследственность
| Часть серии о |
| Генетика |
|---|
 |
|
|
Наследственность , также называемая наследованием или биологическим наследованием , представляет собой передачу признаков от родителей к их потомству; либо посредством бесполого размножения , либо посредством полового размножения потомства , клетки или организмы приобретают генетическую информацию своих родителей. Благодаря наследственности различия между особями могут накапливаться и вызывать путем видов эволюцию естественного отбора . Наследственность в биологии изучает генетика .
Обзор
[ редактировать ]

У людей цвет глаз является примером наследственной характеристики: человек может унаследовать «черту карих глаз» от одного из родителей. [ 1 ] Наследственные признаки контролируются генами организма , а полный набор генов в геноме называется его генотипом . [ 2 ]
Полная совокупность наблюдаемых особенностей строения и поведения организма называется его фенотипом . Эти черты возникают в результате взаимодействия генотипа организма с окружающей средой . [ 3 ] В результате многие аспекты фенотипа организма не наследуются. Например, загорелая кожа возникает в результате взаимодействия генотипа человека и солнечного света; [ 4 ] таким образом, загар не передается детям. Однако некоторые люди загорают легче, чем другие, из-за различий в их генотипе: [ 5 ] Ярким примером являются люди с наследственной чертой альбинизма , которые вообще не загорают и очень чувствительны к солнечным ожогам . [ 6 ]
Известно, что наследственные черты передаются от одного поколения к другому через ДНК , — молекулу которая кодирует генетическую информацию. [ 2 ] ДНК представляет собой длинный полимер , в состав которого входят четыре типа оснований взаимозаменяемых . Последовательность нуклеиновой кислоты (последовательность оснований в определенной молекуле ДНК) определяет генетическую информацию: это можно сравнить с последовательностью букв, обозначающих отрывок текста. [ 7 ] Прежде чем клетка делится митозом , ДНК копируется, так что каждая из двух образовавшихся клеток унаследует последовательность ДНК. Часть молекулы ДНК, определяющая одну функциональную единицу, называется геном ; разные гены имеют разные последовательности оснований. Внутри клеток длинные нити ДНК образуют конденсированные структуры, называемые хромосомами . Организмы наследуют генетический материал от своих родителей в виде гомологичных хромосом , содержащих уникальную комбинацию последовательностей ДНК, кодирующих гены. Конкретное расположение последовательности ДНК внутри хромосомы известно как локус . Если последовательность ДНК в определенном локусе различается у разных людей, разные формы этой последовательности называются аллелями . Последовательности ДНК могут меняться в результате мутаций , создавая новые аллели. Если внутри гена происходит мутация, новая аллель может повлиять на признак, который контролирует ген, изменяя фенотип организма. [ 8 ]
Однако, хотя это простое соответствие между аллелем и признаком работает в некоторых случаях, большинство признаков более сложны и контролируются множеством взаимодействующих генов внутри и между организмами. [ 9 ] [ 10 ] Биологи развития предполагают, что сложные взаимодействия в генетических сетях и коммуникации между клетками могут привести к наследственным вариациям, которые могут лежать в основе некоторых механизмов пластичности и канализации развития . [ 11 ]
Недавние открытия подтвердили важные примеры наследственных изменений, которые нельзя объяснить прямым влиянием молекулы ДНК. Эти явления классифицируются как эпигенетические системы наследования, которые причинно или независимо развиваются в зависимости от генов. Исследования способов и механизмов эпигенетического наследования все еще находятся в зачаточном состоянии, но в последнее время эта область исследований привлекла много внимания, поскольку расширяет сферу наследственности и эволюционной биологии в целом. [ 12 ] Метилирование ДНК , маркирующее хроматин , самоподдерживающиеся метаболические петли , подавление генов интерференцией РНК и трехмерная конформация белков (таких как прионы ) — это области, в которых эпигенетические системы наследования были обнаружены на организменном уровне. [ 13 ] [ 14 ] Наследственность может также проявляться в еще более крупных масштабах. Например, экологическое наследование в процессе построения ниш определяется регулярной и повторяющейся деятельностью организмов в окружающей среде. Это порождает наследие эффектов, которые модифицируются и возвращаются в режим отбора последующих поколений. Потомки наследуют гены плюс характеристики окружающей среды, созданные экологическими действиями предков. [ 15 ] Другие примеры наследственности в эволюции, не находящиеся под прямым контролем генов, включают наследование культурных признаков , групповую наследственность и симбиогенез . [ 16 ] [ 17 ] [ 18 ] Эти примеры наследственности, которые действуют выше гена, широко охватываются названием многоуровневого или иерархического отбора , который был предметом интенсивных дебатов в истории эволюционной науки. [ 17 ] [ 19 ]
Связь с теорией эволюции
[ редактировать ]Когда Чарльз Дарвин предложил свою теорию эволюции в 1859 году, одной из ее главных проблем было отсутствие основного механизма наследственности. [ 20 ] Дарвин верил в сочетание смешанного наследования и наследования приобретенных признаков ( пангенезис ). Смешение наследования привело бы к единообразию популяций всего за несколько поколений, а затем устранило бы вариации в популяции, на которые мог бы действовать естественный отбор. [ 21 ] Это привело к тому, что Дарвин принял некоторые идеи Ламарка в более поздних изданиях « Происхождения видов» и своих более поздних биологических работах. [ 22 ] Основной подход Дарвина к наследственности состоял в том, чтобы описать, как она работает (отмечая, что черты, которые не были явно выражены у родителя во время размножения, могут быть унаследованы, что определенные черты могут быть сцеплены с полом и т. д.), а не предлагать механизмы. . [ нужна ссылка ]
Первоначальная модель наследственности Дарвина была принята, а затем сильно модифицирована его двоюродным братом Фрэнсисом Гальтоном , который заложил основу биометрической школы наследственности. [ 23 ] Гальтон не нашел доказательств, подтверждающих те аспекты модели пангенезиса Дарвина, которые основывались на приобретенных признаках. [ 24 ]
В 1880-х годах было показано, что наследование приобретенных признаков не имеет большого значения, когда Август Вейсман отрезал хвосты многим поколениям мышей и обнаружил, что у их потомства хвосты продолжают развиваться. [ 25 ]
История
[ редактировать ]
Учёные древности имели множество представлений о наследственности: Теофраст предположил, что мужские цветки вызывают созревание женских цветков; [ 26 ] Гиппократ предположил, что «семена» производятся различными частями тела и передаются потомству во время зачатия; [ 27 ] и Аристотель считал, что мужская и женская жидкости смешиваются при зачатии. [ 28 ] Эсхил в 458 г. до н. э. предложил мужчину в качестве родителя, а женщину — в качестве «кормилицы молодой жизни, посеянной в ней». [ 29 ]
Древние представления о наследственности превратились в две обсуждаемые доктрины в 18 веке. Доктрина эпигенеза и доктрина преформации представляли собой два различных взгляда на понимание наследственности. Учение об эпигенезе, созданное Аристотелем , утверждало, что эмбрион постоянно развивается. Модификации родительских признаков передаются эмбриону в течение его жизни. В основу этого учения легла теория наследования приобретенных признаков . В противоположность этому, Доктрина преформации утверждала, что «подобное порождает подобное», когда зародыш будет развиваться, чтобы дать потомство, похожее на родителей. Преформаторская точка зрения считала, что деторождение — это акт раскрытия того, что было создано задолго до этого. Однако это было оспорено созданием в XIX веке клеточной теории , согласно которой фундаментальной единицей жизни является клетка, а не какие-то заранее сформированные части организма. Различные наследственные механизмы, включая смешанное наследование, также рассматривались без должного тестирования или количественной оценки и позже были оспорены. Тем не менее, люди смогли путем искусственного отбора вывести домашние породы животных, а также сельскохозяйственные культуры. Наследование приобретенных черт также составляло часть ранних идей Ламарка об эволюции. [ нужна ссылка ]
В 18 веке голландский микроскопист Антони ван Левенгук (1632–1723) обнаружил «анимакулы» в сперме людей и других животных. [ 30 ] Некоторые учёные предположили, что видели «маленького человечка» ( гомункула ) внутри каждого сперматозоида . Эти ученые сформировали школу мысли, известную как «спермисты». Они утверждали, что единственным вкладом женщины в следующее поколение является матка, в которой вырос гомункул, и пренатальное влияние матки. [ 31 ] Противоположная школа мысли, овисты, считала, что будущий человек находится в яйцеклетке и что сперма просто стимулирует рост яйцеклетки. Овисты считали, что женщины носят яйца, содержащие детей мальчиков и девочек, и что пол потомства определяется задолго до зачатия. [ 32 ]
Ранняя исследовательская инициатива возникла в 1878 году, когда Алфеус Хаятт возглавил исследование по изучению законов наследственности путем сбора данных о семейных фенотипах (размер носа, форма ушей и т. д.), а также проявлении патологических состояний и аномальных характеристик, особенно в отношении возраста. внешности. Одной из целей проекта было сведение данных в таблицу, чтобы лучше понять, почему одни черты выражены постоянно, а другие крайне нерегулярно. [ 33 ]
Грегор Мендель: отец генетики
[ редактировать ]
Идею дисперсного наследования генов можно отнести к моравским [ 34 ] монах Грегор Мендель , опубликовавший свою работу о растениях гороха в 1865 году. Однако его работа не была широко известна и была заново открыта в 1901 году. Первоначально предполагалось, что менделевская наследственность объясняет только большие (качественные) различия, подобные тем, которые видел Мендель в его растения гороха – и идея аддитивного эффекта (количественных) генов не была реализована до тех пор, пока Р. А. Фишера не появилась статья (1918) « Корреляция между родственниками на основании предположения о менделевском наследовании ». были наследственными. Его демонстрация растений гороха стала основой изучения менделевских свойств. Эти черты можно проследить на одном локусе. [ 35 ]
Современное развитие генетики и наследственности
[ редактировать ]В 1930-х годах работы Фишера и других привели к объединению менделевской и биометрической школ в современный эволюционный синтез . Современный синтез преодолел разрыв между генетиками-экспериментаторами и натуралистами; и между обоими и палеонтологами, заявив, что: [ 36 ] [ 37 ]
- Все эволюционные явления можно объяснить в соответствии с известными генетическими механизмами и наблюдениями натуралистов.
- Эволюция постепенная: небольшие генетические изменения, рекомбинация, организованная естественным отбором . Различия между видами (или другими таксонами) объясняются тем, что они возникают постепенно в результате географического разделения и исчезновения (а не скачков).
- Отбор в подавляющем большинстве случаев является основным механизмом изменений; даже небольшие преимущества важны при продолжении. Объектом отбора является фенотип в окружающей его среде. Роль генетического дрейфа неоднозначна; хотя первоначально его активно поддерживал Добжанский , позже его рейтинг был понижен, когда были получены результаты экологической генетики.
- Примат популяционного мышления: генетическое разнообразие, заложенное в природных популяциях, является ключевым фактором эволюции. Сила естественного отбора в дикой природе оказалась больше, чем ожидалось; Влияние экологических факторов, таких как занятость ниши и значение барьеров для потока генов, имеют важное значение.
Идея о том, что видообразование происходит после репродуктивной изоляции популяций, широко обсуждалась. [ 38 ] У растений полиплоидия должна быть включена в любой вид видообразования. Формулировки типа «эволюция состоит прежде всего из изменения частот аллелей от одного поколения к другому» были предложены несколько позже. Традиционная точка зрения состоит в том, что биология развития (« эво-дево ») сыграла небольшую роль в синтезе, но отчет Гэвина де Бира о работе Стивена Джея Гулда предполагает, что он может быть исключением. [ 39 ]
Почти все аспекты синтеза время от времени подвергались сомнению с разной степенью успеха. Однако нет сомнений в том, что этот синтез стал великой вехой в эволюционной биологии. [ 40 ] Оно прояснило многие путаницы и непосредственно способствовало стимулированию большого количества исследований в эпоху после Второй мировой войны .
Трофим Лысенко, однако, вызвал негативную реакцию того, что сейчас называется лысенкоизмом в Советском Союзе , когда он подчеркнул Ламарка идеи о наследовании приобретенных черт . Это движение повлияло на сельскохозяйственные исследования и привело к нехватке продовольствия в 1960-е годы и серьезно повлияло на СССР. [ 41 ]
Появляется все больше доказательств того, что эпигенетические изменения у людей наследуются трансгенеративно. [ 42 ] и другие животные. [ 43 ]
Распространенные генетические нарушения
[ редактировать ]Типы
[ редактировать ]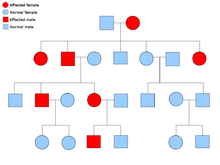
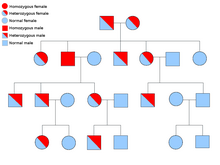
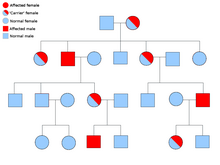
Описание типа биологического наследования состоит из трех основных категорий:
- 1. Количество задействованных локусов
- Моногенетический (также называемый «простым») - один локус .
- Олигогенный – мало локусов
- Полигенетический – много локусов
- 2. Задействованные хромосомы
- Аутосомный – локусы не расположены на половой хромосоме.
- Гоносомный – локусы расположены на половой хромосоме.
- Х-хромосомный – локусы расположены на Х-хромосоме (более распространенный случай)
- Y-хромосома – локусы расположены на Y-хромосоме.
- Митохондриальные – локусы расположены на митохондриальной ДНК.
- 3. Корреляция генотип – фенотип.
- Доминантный
- Промежуточный (также называемый « кодоминантным »)
- Рецессивный
- Чрезмерно доминантный
- Недоминантный
Эти три категории являются частью каждого точного описания способа наследования в указанном выше порядке. Кроме того, могут быть добавлены дополнительные характеристики:
- 4. Случайные совпадения и взаимодействия с окружающей средой
- Пенетрантность
- Полный
- Неполное (процентное число)
- Выразительность
- Неизменный
- Переменная
- Наследственность (при полигенетическом, а иногда и при олигогенетическом типе наследования)
- материнского или отцовского импринтинга Феномен (см. также эпигенетику )
- Пенетрантность
- 5. Взаимодействия, связанные с сексом
- Наследование, сцепленное с полом ( гоносомные локусы)
- Ограниченное по полу проявление фенотипа (например, крипторхизм )
- Наследование по материнской линии (в случае локусов митохондриальной ДНК )
- Наследование по отцовской линии (в случае Y-хромосомных локусов)
- 6. Локус-локусные взаимодействия
- Эпистаз с другими локусами (например, сверхдоминирование )
- Сцепление гена с другими локусами (см. также кроссинговер )
- Гомозиготные летальные факторы
- Полулетальные факторы
Определение и описание способа наследования достигается также прежде всего путем статистического анализа родословных данных. Если известны вовлеченные локусы, методы молекулярной генетики можно также использовать .
Доминантные и рецессивные аллели
[ редактировать ]Аллель . называют доминантной, если она всегда выражена во внешнем виде организма (фенотипе) при наличии хотя бы одной ее копии Например, у гороха аллель зеленых стручков G доминирует над аллелем желтых стручков g . Таким образом, растения гороха с парой аллелей или GG (гомозигота) Gg ( гетерозигота) будут иметь зеленые стручки. Аллель желтых стручков рецессивный. Эффекты этого аллеля заметны только тогда, когда он присутствует в обеих хромосомах gg (гомозигота). Это происходит от зиготности , степени, в которой обе копии хромосомы или гена имеют одинаковую генетическую последовательность, другими словами, степени сходства аллелей в организме.
-
 Наследственные дефекты ферментов обычно наследуются аутосомно, поскольку не-Х-хромосом больше, чем Х-хромосом, и рецессивно, поскольку ферментов непораженных генов обычно достаточно для предотвращения симптомов у носителей.
Наследственные дефекты ферментов обычно наследуются аутосомно, поскольку не-Х-хромосом больше, чем Х-хромосом, и рецессивно, поскольку ферментов непораженных генов обычно достаточно для предотвращения симптомов у носителей. -
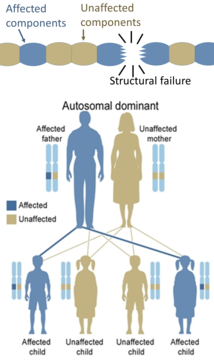 С другой стороны, наследственные дефекты структурных белков (такие как несовершенный остеогенез , синдром Марфана и многие синдромы Элерса-Данлоса ) обычно являются аутосомно-доминантными, поскольку дефектов некоторых компонентов достаточно, чтобы сделать всю структуру дисфункциональной. Это доминантно-негативный процесс, при котором мутированный генный продукт отрицательно влияет на немутантный генный продукт в той же клетке.
С другой стороны, наследственные дефекты структурных белков (такие как несовершенный остеогенез , синдром Марфана и многие синдромы Элерса-Данлоса ) обычно являются аутосомно-доминантными, поскольку дефектов некоторых компонентов достаточно, чтобы сделать всю структуру дисфункциональной. Это доминантно-негативный процесс, при котором мутированный генный продукт отрицательно влияет на немутантный генный продукт в той же клетке.


См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Штурм РА; Фрудакис Т.Н. (2004). «Цвет глаз: порталы в гены пигментации и происхождение». Тенденции Жене . 20 (8): 327–332. дои : 10.1016/j.tig.2004.06.010 . ПМИД 15262401 .
- ^ Перейти обратно: а б Пирсон Х (2006). «Генетика: что такое ген?» . Природа . 441 (7092): 398–401. Бибкод : 2006Natur.441..398P . дои : 10.1038/441398a . ПМИД 16724031 . S2CID 4420674 .
- ^ Вишер ПМ; Хилл РГ; Рэй Н.Р. (2008). «Наследственность в эпоху геномики – концепции и заблуждения». Нат. Преподобный Жене . 9 (4): 255–266. дои : 10.1038/nrg2322 . ПМИД 18319743 . S2CID 690431 .
- ^ Шоаг Дж; и др. (январь 2013 г.). «Коактиваторы PGC-1 регулируют MITF и реакцию на загар» . Мол Клетка . 49 (1): 145–157. дои : 10.1016/j.molcel.2012.10.027 . ПМЦ 3753666 . ПМИД 23201126 .
- ^ Фо Л.Н.; Личман С.А. (февраль 2010 г.). «Генетика пигментации и предрасположенности к меланоме» . G Итал Дерматол Венереол . 145 (1): 37–45. ПМИД 20197744 . Архивировано из оригинала 28 марта 2019 г. Проверено 26 марта 2013 г.
- ^ Эттинг В.С.; Блестящий МХ; Король РА (1996). «Клинический спектр альбинизма у человека и по действию». Молекулярная медицина сегодня . 2 (8): 330–335. дои : 10.1016/1357-4310(96)81798-9 . ПМИД 8796918 .
- ^ Гриффитс, Энтони, Дж. Ф.; Весслер, Сьюзен Р.; Кэрролл, Шон Б.; Добли Дж. (2012). Введение в генетический анализ (10-е изд.). Нью-Йорк: WH Freeman and Company. п. 3. ISBN 978-1-4292-2943-2 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Футуйма, Дуглас Дж. (2005). Эволюция . Сандерленд, Массачусетс: Sinauer Associates, Inc. ISBN 978-0-87893-187-3 .
- ^ Филлипс ПК (2008). «Эпистаз – существенная роль взаимодействий генов в структуре и эволюции генетических систем» . Нат. Преподобный Жене . 9 (11): 855–867. дои : 10.1038/nrg2452 . ПМК 2689140 . ПМИД 18852697 .
- ^ Ву Р; Лин М (2006). «Функциональное картирование - как составить карту и изучить генетическую архитектуру динамических сложных признаков». Нат. Преподобный Жене . 7 (3): 229–237. дои : 10.1038/nrg1804 . ПМИД 16485021 . S2CID 24301815 .
- ^ Яблонка, Э.; Лэмб, MJ (2002). «Изменяющаяся концепция эпигенетики» (PDF) . Анналы Нью-Йоркской академии наук . 981 (1): 82–96. Бибкод : 2002NYASA.981...82J . дои : 10.1111/j.1749-6632.2002.tb04913.x . ПМИД 12547675 . S2CID 12561900 . Архивировано из оригинала (PDF) 11 мая 2011 г.
- ^ Яблонка, Э.; Раз, Г. (2009). «Трансгенерационное эпигенетическое наследование: распространенность, механизмы и значение для изучения наследственности и эволюции» (PDF) . Ежеквартальный обзор биологии . 84 (2): 131–176. CiteSeerX 10.1.1.617.6333 . дои : 10.1086/598822 . ПМИД 19606595 . S2CID 7233550 . Архивировано (PDF) из оригинала 15 июля 2011 г. Проверено 18 февраля 2011 г.
- ^ Боссдорф, О.; Аркури, Д.; Ричардс, КЛ; Пильуччи, М. (2010). «Экспериментальное изменение метилирования ДНК влияет на фенотипическую пластичность экологически значимых признаков Arabidopsis thaliana » (PDF) . Эволюционная экология . 24 (3): 541–553. дои : 10.1007/s10682-010-9372-7 . S2CID 15763479 . Архивировано (PDF) из оригинала 01 марта 2020 г. Проверено 15 августа 2019 г.
- ^ Яблонка, Э.; Лэмб, М. (2005). Эволюция в четырех измерениях: генетическом, эпигенетическом, поведенческом и символическом . МТИ Пресс. ISBN 978-0-262-10107-3 . Архивировано из оригинала 27 декабря 2021 г. Проверено 27 июня 2015 г.
- ^ Лаланд, КНЦ; Стерельный, К. (2006). «Перспектива: семь причин (не) пренебрегать строительством ниш» . Эволюция . 60 (8): 1751–1762. дои : 10.1111/j.0014-3820.2006.tb00520.x . ПМИД 17089961 .
- ^ Чепмен, MJ; Маргулис, Л. (1998). «Морфогенез посредством симбиогенеза» (PDF) . Международная микробиология . 1 (4): 319–326. ПМИД 10943381 . Архивировано из оригинала (PDF) 23 августа 2014 г.
- ^ Перейти обратно: а б Уилсон, Д.С.; Уилсон, Э.О. (2007). «Переосмысление теоретических основ социобиологии» (PDF) . Ежеквартальный обзор биологии . 82 (4): 327–348. дои : 10.1086/522809 . ПМИД 18217526 . S2CID 37774648 . Архивировано из оригинала (PDF) 11 мая 2011 г.
- ^ Биджма, П.; Уэйд, MJ (2008). «Совместное влияние родства, многоуровневого отбора и косвенного генетического воздействия на реакцию на генетический отбор» . Журнал эволюционной биологии . 21 (5): 1175–1188. дои : 10.1111/j.1420-9101.2008.01550.x . ПМИД 18547354 . S2CID 7204089 .
- ^ Врба, ЕС; Гулд, С.Дж. (1986). «Иерархическое расширение сортировки и выбора: сортировку и выбор нельзя отождествлять» (PDF) . Палеобиология . 12 (2): 217–228. Бибкод : 1986Pbio...12..217V . дои : 10.1017/S0094837300013671 . S2CID 86593897 . Архивировано из оригинала (PDF) 4 августа 2016 г. Проверено 18 февраля 2011 г.
- ^ Гриффитс, Энтони, Дж. Ф.; Весслер, Сьюзен Р.; Кэрролл, Шон Б.; Добли, Джон (2012). Введение в генетический анализ (10-е изд.). Нью-Йорк: WH Freeman and Company. п. 14. ISBN 978-1-4292-2943-2 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Чарльзуорт, Брайан и Чарльзуорт, Дебора (ноябрь 2009 г.). «Дарвин и генетика» . Генетика . 183 (3): 757–766. дои : 10.1534/genetics.109.109991 . ПМЦ 2778973 . ПМИД 19933231 . Архивировано из оригинала 29 апреля 2019 г. Проверено 26 марта 2013 г.
- ^ Бард, Джонатан Б.Л. (2011). «Следующий эволюционный синтез: от Ламарка и Дарвина к геномной изменчивости и системной биологии» . Сотовая связь и сигнализация . 9 (30): 30. дои : 10.1186/1478-811X-9-30 . ПМЦ 3215633 . ПМИД 22053760 .
- ^ «Фрэнсис Гальтон (1822-1911)» . Музей науки . Архивировано из оригинала 30 января 2016 года . Проверено 26 марта 2013 г.
- ^ Лю Ю. (май 2008 г.). «Новый взгляд на пангенезис Дарвина». Biol Rev Camb Philos Soc . 83 (2): 141–149. дои : 10.1111/j.1469-185X.2008.00036.x . ПМИД 18429766 . S2CID 39953275 .
- ^ Липтон, Брюс Х. (2008). Биология веры: раскрытие силы сознания, материи и чудес . Hay House, Inc., стр. 12 . ISBN 978-1-4019-2344-0 .
- ^ Негби, Моше (лето 1995 г.). «Мужчина и женщина в ботанических трудах Теофраста». Журнал истории биологии . 28 (2): 317–332. дои : 10.1007/BF01059192 . S2CID 84754865 .
- ^ Гипократ (1981). Трактаты Гиппократа: О поколении – Природа ребенка – Болезни Ic . Вальтер де Грютер. п. 6. ISBN 978-3-11-007903-6 .
- ^ «Биология Аристотеля – 5.2. От исследования к пониманию; от хоти к диоти» . Стэнфордский университет. 15 февраля 2006 г. Архивировано из оригинала 7 мая 2019 г. . Проверено 26 марта 2013 г.
- ^ Евменид 658–661
- ^ Сноу, Курт. «Удивительные маленькие «животные» Антони ван Левенгука » . Лебен. Архивировано из оригинала 24 апреля 2013 года . Проверено 26 марта 2013 г.
- ^ Лоуренс, Сера Р. (2008). Эскиз гомункула Хартсокера из «Эссай де Диоптрик» . Энциклопедия проекта «Эмбрион». ISSN 1940-5030 . Архивировано из оригинала 9 апреля 2013 г. Проверено 26 марта 2013 г.
- ^ Готлиб, Гилберт (2001). Индивидуальное развитие и эволюция: генезис нового поведения . Психология Пресс. п. 4. ISBN 978-1-4106-0442-2 .
- ^ Журнал Scientific American, «Наследственность» . Манн и компания. 1878-11-30. п. 343. Архивировано из оригинала 18 мая 2022 г. Проверено 6 августа 2021 г.
- ^ Хениг, Робин Маранц (2001). Монах в саду: потерянный и найденный гений Грегора Менделя, отца генетики . Хоутон Миффлин. ISBN 978-0-395-97765-1 .
Статья, написанная малоизвестным моравским монахом по имени Грегор Мендель.
- ^ Перейти обратно: а б Карлсон, Нил Р. (2010). Психология: наука о поведении , с. 206. Торонто: Пирсон Канада. ISBN 978-0-205-64524-4 . ОСЛК 1019975419
- ^ Майр и Провайн, 1998 г.
- ^ Майр Э. 1982. Рост биологической мысли: разнообразие, эволюция и наследственность . Гарвард, Кэмбс. стр. 567 и след.
- ^ Палумби, Стивен Р. (1994). «Генетическая дивергенция, репродуктивная изоляция и морское видообразование». Ежегодный обзор экологии и систематики . 25 : 547–572. doi : 10.1146/annurev.es.25.110194.002555 .
- ^ Гулд С.Дж. Онтогенез и филогения . Гарвард, 1977. стр. 221–222.
- ^ Хандшу, Стефан; Миттерёкер, Филипп (июнь 2012 г.). «Эволюция - расширенный синтез. Исследовательское предложение, достаточно убедительное для большинства биологов-эволюционистов?». Бюллетень этологии человека . 27 (1–2): 18–21. ISSN 2224-4476 .
- ^ Харпер, Питер С. (3 августа 2017 г.). «Генетика человека в смутные времена и места» . Эредитас . 155 :7. дои : 10.1186/s41065-017-0042-4 . ISSN 1601-5223 . ПМК 5541658 . ПМИД 28794693 .
- ^ Шиф, М (2015). «Негенетическое наследование и трансгенерационная эпигенетика». Тенденции молекулярной медицины . 21 (2): 134–144. doi : 10.1016/j.molmed.2014.12.004 . ПМИД 25601643 .
- ^ Кисимото, С; и др. (2017). «Стрессы окружающей среды вызывают трансгенерационно наследуемые преимущества выживания посредством связи между зародышевой линией и сомой у Caenorhabditis elegans» . Природные коммуникации . 8 : 14031. Бибкод : 2017NatCo...814031K . дои : 10.1038/ncomms14031 . HDL : 2433/217772 . ПМК 5227915 . ПМИД 28067237 .