Цитохром P450
| Цитохром P450 | |||
|---|---|---|---|
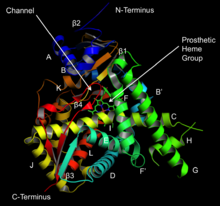 Структура ланостерол-14α-деметилазы ( CYP51 ) | |||
| Идентификаторы | |||
| Символ | стр.450 | ||
| Пфам | PF00067 | ||
| ИнтерПро | IPR001128 | ||
| PROSITE | PDOC00081 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 2cpp / СКОПе / СУПФАМ | ||
| Суперсемейство OPM | 39 | ||
| белок OPM | 2 спальни | ||
| CDD | cd00302 | ||
| Мембраном | 265 | ||
| |||
Цитохромы P450 ( P450 или CYP ) представляют собой суперсемейство ферментов , содержащих гем в качестве кофактора , которые в основном, но не исключительно, функционируют как монооксигеназы . [ 1 ] Однако они не вездесущи; например, они не обнаружены в Escherichia coli . [ 2 ] У млекопитающих эти ферменты окисляют стероиды , жирные кислоты , ксенобиотики , участвуют во многих биосинтезах. [ 1 ] Путем гидроксилирования ферменты CYP450 превращают ксенобиотики в гидрофильные производные, которые легче выводятся из организма.
P450, как правило, представляют собой концевые ферменты оксидазы в цепях переноса электронов , которые в широком смысле классифицируются как системы, содержащие P450 . Термин «P450» получен из спектрофотометрического пика на длине волны максимума поглощения фермента (450 нм ), когда он находится в восстановленном состоянии и образует комплекс с окисью углерода . Большинству P450 требуется белок-партнер для доставки одного или нескольких электронов для восстановления железа (и, в конечном итоге, молекулярного кислорода ).
Номенклатура
[ редактировать ]Гены, кодирующие ферменты P450, и сами ферменты обозначаются корневым символом CYP , обозначающим суперсемейство , за которым следует число, обозначающее семейство генов , заглавная буква, обозначающая подсемейство, и еще одна цифра, обозначающая отдельный ген. Принято выделять имя курсивом при упоминании гена. Например, CYP2E1 — это ген, который кодирует фермент CYP2E1 — один из ферментов, участвующих в метаболизме парацетамола (ацетаминофена). Номенклатура CYP является официальным соглашением об именах, хотя иногда CYP450 или CYP 450 используются как синонимы. Эти имена никогда не следует использовать согласно номенклатурному соглашению (поскольку они обозначают P450 в семействе номер 450). Однако некоторые названия генов или ферментов для P450 также называются историческими названиями (например, P450 BM3 для CYP102A1) или функциональными названиями, обозначающими каталитическую активность и название соединения, используемого в качестве субстрата. включают CYP5A1 , тромбоксан А 2 сокращенно TBXAS1 ( тромбоксан . А синтазу ) , 2 синтаза Примеры 1 ) и CYP51A1 , ланостерол-14-α-деметилаза, иногда неофициально сокращенно называемая LDM в зависимости от ее субстрата ( L -аностерин) и активности ( D e M -этилирование). [ 3 ]
Текущие рекомендации по номенклатуре предполагают, что члены новых семейств CYP имеют как минимум 40% аминокислотной идентичности, тогда как члены подсемейств должны иметь как минимум 55% аминокислотной идентичности. Номенклатурные комитеты присваивают и отслеживают как основные названия генов ( домашняя страница Cytochrome P450 , архивировано 27 июня 2010 г. в Wayback Machine ), так и аллелей названия ( Комитет по номенклатуре аллелей CYP ). [ 4 ] [ 5 ]
Классификация
[ редактировать ]В зависимости от природы белков-переносчиков электронов P450 можно разделить на несколько групп: [ 6 ]
- Микросомальные системы P450
- при котором электроны передаются от НАДФН посредством редуктазы цитохрома P450 (различно CPR, POR или CYPOR). Цитохром b 5 (cyb 5 ) также может способствовать снижению мощности этой системы после восстановления цитохром b 5 редуктазой (CYB 5 R).
- Митохондриальные системы P450
- которые используют адренодоксинредуктазу и адренодоксин для переноса электронов от НАДФН к P450.
- Бактериальные системы P450
- которые используют ферредоксинредуктазу и ферредоксин для переноса электронов на P450.
- CYB 5 R/cyb 5 /P450 Системы
- в котором оба электрона, необходимые для CYP, происходят от цитохрома b 5 .
- Системы FMN/Fd/P450
- первоначально обнаружен у видов Rhodococcus , у которых содержащая FMN -домен, редуктаза, слита с CYP.
- Только системы P450
- которые не требуют внешней восстанавливающей мощности. Известные из них включают тромбоксансинтазу (CYP5), простациклинсинтазу (CYP8) и CYP74A ( алленоксидсинтаза ).
Наиболее распространенной реакцией, катализируемой цитохромами Р450, является монооксигеназная реакция, например, внедрение одного атома кислорода в алифатическое положение органического субстрата (RH), при этом другой атом кислорода восстанавливается до воды:
Родственные ферменты гидроксилирования
[ редактировать ]Во многих реакциях гидроксилирования (вставки гидроксильных групп) используются ферменты CYP, но существует множество других гидроксилаз. Альфа-кетоглутарат-зависимые гидроксилазы также основаны на промежуточном соединении Fe=O, но лишены гемов. Метанмонооксигеназа, превращающая метан в метанол, представляет собой негемовые ферменты на основе железа и меди. [ 7 ]
Механизм
[ редактировать ]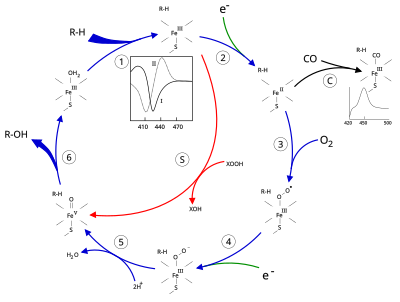
Структура
[ редактировать ]Активный центр цитохрома Р450 содержит гем-железный центр. Железо связано с белком через цистеина тиолатный лиганд . Этот цистеин и несколько фланкирующих остатков высококонсервативны в известных P450 и имеют формальный PROSITE консенсусный паттерн подписи [FW] - [SGNH] - x - [GD] - {F} - [RKHPT] - {P} - C - [ ЛИВМФАП] - [ГАД]. [ 8 ] В целом каталитический цикл P450 протекает следующим образом:
Каталитический цикл
[ редактировать ]- Субстрат связывается вблизи гемовой группы , на стороне, противоположной аксиальному тиолату. Связывание с субстратом вызывает изменение конформации активного центра, часто вытесняя молекулу воды из дистального аксиального координационного положения железа гема. [ 9 ] и изменение состояния гема железа с низкоспинового на высокоспиновое. [ 10 ]
- Связывание с субстратом индуцирует перенос электронов от НАД(Ф)Н через редуктазу цитохрома Р450 или другую ассоциированную редуктазу . [ 11 ] превращение Fe(III) в Fe(II).
- Молекулярный кислород связывается с образовавшимся центром гема железа в дистальной осевой координационной позиции, первоначально образуя дикислородный аддукт, аналогичный оксимиоглобину.
- Переносится второй электрон либо от редуктазы цитохрома P450 , либо от ферредоксина , либо от цитохрома b 5 , восстанавливая аддукт Fe-O 2 с образованием короткоживущего пероксо-состояния.
- Пероксогруппа, образовавшаяся на этапе 4, дважды быстро протонируется, высвобождая одну молекулу воды и образуя высокореактивную разновидность, называемую Соединением 1 P450 (или просто Соединением I). Это высокореактивное промежуточное соединение было выделено в 2010 году. [ 12 ] ) железо(IV) P450 Соединение 1 представляет собой оксо-(или феррил с дополнительным окислительным эквивалентом, делокализованным над порфириновым и тиолатным лигандами. Доказательства альтернативы перферрил- железо(V)-оксо. [ 9 ] отсутствует. [ 12 ]
- В зависимости от субстрата и фермента ферменты P450 могут катализировать любую из самых разнообразных реакций. Проиллюстрировано гипотетическое гидроксилирование. После того как гидроксилированный продукт высвобождается из активного центра, фермент возвращается в исходное состояние, при этом молекула воды возвращается и занимает дистальную координационную позицию ядра железа.

- Альтернативный путь монооксигенации — через «перекисный шунт» (путь «S» на рисунке). Этот путь влечет за собой окисление комплекса железо-субстрат донорами атомов кислорода, такими как пероксиды и гипохлориты. [ 13 ] Гипотетический пероксид «XOOH» показан на схеме.
Детали механизма, включая механизм отскока кислорода , были исследованы с использованием синтетических аналогов, состоящих из оксогемовых комплексов железа. [ 14 ]
Спектроскопия
[ редактировать ]Связывание субстрата отражается на спектральных свойствах фермента с увеличением оптической плотности при 390 нм и уменьшением при 420 нм. Его можно измерить с помощью разностной спектроскопии, и он называется разностным спектром «типа I» (см. врезной график на рисунке). Некоторые субстраты вызывают противоположное изменение спектральных свойств, спектр «обратного типа I», посредством процессов, которые пока неясны. Ингибиторы и некоторые субстраты, которые непосредственно связываются с железом гема, вызывают разностный спектр типа II с максимумом при 430 нм и минимумом при 390 нм (см. врезной график на рисунке). Если восстанавливающие эквиваленты недоступны, этот комплекс может оставаться стабильным, что позволяет определить степень связывания на основе измерений поглощения in vitro. [ 13 ] C: Если окись углерода (CO) связывается с восстановленным P450, каталитический цикл прерывается. Эта реакция дает классический разностный спектр CO с максимумом при 450 нм. Однако прерывающее и ингибирующее действие CO варьируется в зависимости от разных CYP, поэтому семейство CYP3A подвергается относительно меньшему воздействию. [ 15 ] [ 16 ]
См. также
[ редактировать ]Дальнейшее чтение
[ редактировать ]- Эстабрук Р.В. (декабрь 2003 г.). «Страсть к P450 (Воспоминания о ранней истории исследований цитохрома P450)». Метаболизм и распределение лекарств . 31 (12): 1461–1473. дои : 10.1124/dmd.31.12.1461 . ПМИД 14625342 . S2CID 43655270 .
Ссылки
[ редактировать ]- ^ Перейти обратно: а б «Цитохром Р450» . ИнтерПро .
- ^ Дэниелсон П.Б. (декабрь 2002 г.). «Суперсемейство цитохрома P450: биохимия, эволюция и метаболизм лекарств у человека». Современный метаболизм лекарств . 3 (6): 561–597. дои : 10.2174/1389200023337054 . ПМИД 12369887 .
- ^ «Просмотр последовательностей NCBI» . Проверено 19 ноября 2007 г.
- ^ Нельсон Д.Р. (октябрь 2009 г.). «Домашняя страница цитохрома p450» . Геномика человека . 4 (1): 59–65. дои : 10.1186/1479-7364-4-1-59 . ПМК 3500189 . ПМИД 19951895 .
- ^ Нельсон Д.Р. (январь 2011 г.). «Прогресс в отслеживании путей эволюции цитохрома P450». Biochimica et Biophysical Acta (BBA) - Белки и протеомика . 1814 (1): 14–18. дои : 10.1016/j.bbapap.2010.08.008 . ПМИД 20736090 .
- ^ Ханукоглу I (1996). «Белки-переносчики электронов систем цитохрома P450» (PDF) . Адв. Мол. Клеточная Биол . Достижения в области молекулярной и клеточной биологии. 14 :29–55. дои : 10.1016/S1569-2558(08)60339-2 . ISBN 978-0-7623-0113-3 .
- ^ Туччи FJ, Розенцвейг AC (февраль 2024 г.). «Прямое окисление метана медь- и железозависимыми метанмонооксигеназами» . Химические обзоры . 124 (3): 1288–1320. doi : 10.1021/acs.chemrev.3c00727 . ПМЦ 10923174 . ПМИД 38305159 .
- ^ [1] Архивировано 18 октября 2019 г. в Wayback Machine консенсусном шаблоне PROSITE для P450.
- ^ Перейти обратно: а б Менье Б., де Виссер С.П., Шайк С. (сентябрь 2004 г.). «Механизм реакций окисления, катализируемых ферментами цитохрома p450». Химические обзоры . 104 (9): 3947–3980. дои : 10.1021/cr020443g . ПМИД 15352783 . S2CID 33927145 .
- ^ Пулос Т.Л., Финзель Б.К., Ховард А.Дж. (июнь 1987 г.). «Кристаллическая структура цитохрома P450cam высокого разрешения». Журнал молекулярной биологии . 195 (3): 687–700. дои : 10.1016/0022-2836(87)90190-2 . ПМИД 3656428 .
- ^ Слайгар С.Г., Синти Д.Л., Гибсон Г.Г., Шенкман Дж.Б. (октябрь 1979 г.). «Контроль спинового состояния окислительно-восстановительного потенциала цитохрома P450 печени». Связь с биохимическими и биофизическими исследованиями . 90 (3): 925–932. дои : 10.1016/0006-291X(79)91916-8 . ПМИД 228675 .
- ^ Перейти обратно: а б Риттл Дж., Грин MT (ноябрь 2010 г.). «Соединение I цитохрома P450: захват, характеристика и кинетика активации связи CH». Наука . 330 (6006): 933–937. Бибкод : 2010Sci...330..933R . дои : 10.1126/science.1193478 . ПМИД 21071661 . S2CID 206528205 .
- ^ Перейти обратно: а б Ортис де Монтеллано PR (2005). Цитохром P450: структура, механизм и биохимия (3-е изд.). Нью-Йорк: Издательство Kluwer Academic/Plenum. ISBN 978-0-306-48324-0 .
- ^ Хуан Икс, Гроувс Дж.Т. (март 2018 г.). «Активация кислорода и радикальные превращения в гем-белках и металлопорфиринах» . Химические обзоры . 118 (5): 2491–2553. doi : 10.1021/acs.chemrev.7b00373 . ПМК 5855008 . ПМИД 29286645 .
- ^ Хоппер КП, Замбрана ПН, Гебель У, Уолборн Дж (июнь 2021 г.). «Краткая история угарного газа и его терапевтического происхождения». Оксид азота . 111–112: 45–63. дои : 10.1016/j.niox.2021.04.001 . ПМИД 33838343 . S2CID 233205099 .
- ^ Смит А.Т., Пазикни С., Марвин К.А., Стивенс Д.Д., Полсен К.М., Берстин Дж.Н. (апрель 2015 г.). «Функциональное расхождение геметиолатных белков: классификация, основанная на спектроскопических признаках». Химические обзоры . 115 (7): 2532–2558. дои : 10.1021/cr500056m . ПМИД 25763468 .
| Базы данных органов управления : Национальные |
|---|