БРЦА2
| БРЦА2 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | BRCA2 , BRCC2, BROVCA2, FACD, FAD, FAD1, FANCD, FANCD1, GLM3, PNCA2, XRCC11, рак молочной железы 2, связанный с репарацией ДНК, рак молочной железы 2, раннее начало, связанный с репарацией ДНК BRCA2, Гены | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | ОМИМ : 600185 ; МГИ : 109337 ; Гомологен : 41 ; Генные карты : BRCA2 ; OMA : BRCA2 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||





BRCA2 и BRCA2 ( / ˌ b r æ k ə ˈ t uː / [ 5 ] ) — гены человека и их белковые продукты соответственно. Официальный символ (BRCA2, курсив обозначает ген, некурсив — белок) и официальное название (первоначально рак молочной железы 2 ; в настоящее время BRCA2, связанный с репарацией ДНК ) поддерживаются Комитетом по номенклатуре генов HUGO . Один альтернативный символ, FANCD1 , указывает на его связь с белковым комплексом FANC . Ортологи , называемые Brca2 и Brca2, распространены у других позвоночных . видов [ 6 ] [ 7 ] BRCA2 человека . — ген-супрессор опухолей [ 8 ] [ 9 ] (в частности, ген-смотритель ), обнаруженный у всех людей; его белок , также называемый синонимом « белок предрасположенности к раку молочной железы 2-го типа» , отвечает за восстановление ДНК. [ 10 ]
BRCA2 и BRCA1 обычно экспрессируются в клетках молочной железы и других тканей, где они помогают восстановить поврежденную ДНК или уничтожить клетки, если ДНК не может быть восстановлена. Они участвуют в восстановлении хромосомных повреждений и играют важную роль в безошибочном ДНК . восстановлении двухцепочечных разрывов [ 11 ] [ 12 ] Если BRCA1 или сам BRCA2 поврежден мутацией , BRCA поврежденная ДНК не восстанавливается должным образом, и это увеличивает риск рака молочной железы . [ 13 ] [ 14 ] BRCA1 и BRCA2 были описаны как «гены предрасположенности к раку молочной железы» и «белки восприимчивости к раку молочной железы». Преобладающий аллель имеет нормальную функцию подавления опухоли, тогда как мутации с высокой пенетрантностью в этих генах вызывают потерю функции подавления опухоли, что коррелирует с повышенным риском рака молочной железы. [ 15 ]
Ген BRCA2 расположен на длинном (q) плече хромосомы 13 в положении 12,3 (13q12.3). [ 16 ] Эталонный ген BRCA2 человека содержит 27 экзонов, а кДНК имеет 10 254 пары оснований. [ 17 ] кодирующий белок, состоящий из 3418 аминокислот. [ 18 ] [ 19 ]
Функция
[ редактировать ]
Хотя структуры генов BRCA1 и BRCA2 сильно различаются, по крайней мере некоторые функции взаимосвязаны. Белки , вырабатываемые обоими генами , необходимы для восстановления поврежденной ДНК (см. Рисунок этапов рекомбинационного восстановления). BRCA2 связывает одноцепочечную ДНК и напрямую взаимодействует с рекомбиназой RAD51, стимулируя [ 27 ] и поддерживать [ 28 ] Инвазия цепи, жизненно важный этап гомологичной рекомбинации . Локализация RAD51 на двухцепочечном разрыве ДНК требует образования комплекса BRCA1-PALB2-BRCA2. PALB2 (партнер и локализатор BRCA2) [ 29 ] может функционировать синергически с химерой BRCA2 (называемой пикколо или piBRCA2), способствуя дальнейшей инвазии цепи. [ 30 ] Эти разрывы могут быть вызваны естественным и медицинским излучением или другими воздействиями окружающей среды, но также происходят, когда хромосомы обмениваются генетическим материалом во время особого типа деления клеток, в результате которого создаются сперматозоиды и яйцеклетки ( мейоз ). Двухцепочечные разрывы также возникают во время восстановления поперечных связей ДНК. Восстанавливая ДНК, эти белки играют роль в поддержании стабильности человеческого генома и предотвращении опасных перестроек генов, которые могут привести к гематологическим и другим видам рака.
Было показано, что BRCA2 играет решающую роль в защите от MRE11 -зависимой нуклеолитической деградации перевернутых вилок , которые образуются во время остановки вилки репликации ДНК (вызванной такими препятствиями, как мутации, интеркалирующие агенты и т. д.). [ 31 ]
Подобно BRCA1, BRCA2, вероятно, регулирует активность других генов и играет решающую роль в развитии эмбриона.
Клиническое значение
[ редактировать ]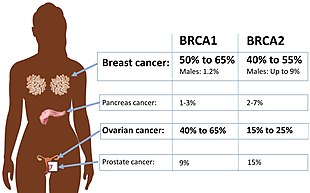
Определенные вариации гена BRCA2 повышают риск развития рака молочной железы как части наследственного синдрома рака молочной железы и яичников . Исследователи выявили сотни мутаций в гене BRCA2 , многие из которых вызывают повышенный риск развития рака. Мутации BRCA2 обычно представляют собой вставки или делеции небольшого количества пар оснований ДНК в гене. В результате этих мутаций белковый продукт гена BRCA2 является аномальным и не функционирует должным образом. Исследователи полагают, что дефектный белок BRCA2 не способен исправить повреждения ДНК, возникающие по всему геному. В результате происходит увеличение количества мутаций из-за склонного к ошибкам синтеза транслейкоза после невосстановленного повреждения ДНК, и некоторые из этих мутаций могут привести к неконтролируемому делению клеток и образованию опухоли.
Люди, у которых есть две мутированные копии гена BRCA2 , имеют один тип анемии Фанкони . Это состояние вызвано чрезвычайно низким уровнем белка BRCA2 в клетках, что приводит к накоплению поврежденной ДНК. Пациенты с анемией Фанкони склонны к нескольким типам лейкемии (разновидность рака клеток крови); солидные опухоли, особенно головы, шеи, кожи и репродуктивных органов; и подавление костного мозга (снижение производства клеток крови, что приводит к анемии ). Женщины, унаследовавшие дефектный ген BRCA1 или BRCA2, имеют настолько высокий риск развития рака молочной железы и яичников, который кажется настолько избирательным, что многие носители мутаций предпочитают профилактическое хирургическое вмешательство . Было много предположений, объясняющих столь поразительную тканевую специфичность. Основные факторы, определяющие возникновение BRCA1- и BRCA2 -ассоциированного наследственного рака, связаны с тканевой специфичностью возбудителя рака, агента, вызывающего хроническое воспаление, или канцерогена. Ткань-мишень может иметь рецепторы для возбудителя, избирательно подвергаться воздействию канцерогенов и инфекционному процессу. Врожденный геномный дефицит нарушает нормальные реакции и усугубляет восприимчивость к заболеваниям в органах-мишенях. Эта теория также соответствует данным по нескольким супрессорам опухолей, помимо BRCA1 или BRCA2 . Основным преимуществом этой модели является то, что она предполагает наличие некоторых вариантов помимо профилактической хирургии. [ 33 ]
Помимо рака молочной железы у мужчин и женщин, мутации в BRCA2 также приводят к повышенному риску рака яичников , маточной трубы , простаты и поджелудочной железы . В некоторых исследованиях мутации в центральной части гена были связаны с более высоким риском рака яичников и более низким риском рака простаты , чем мутации в других частях гена. Некоторые другие виды рака [ который? ] также наблюдались в некоторых семьях с BRCA2 . мутациями [ нужна ссылка ]
В целом, сильно наследственные мутации генов (включая мутации BRCA2 ) составляют лишь 5-10% случаев рака молочной железы; Конкретный риск заболевания раком молочной железы или другим раком для любого носителя мутации BRCA2 зависит от многих факторов. [ 34 ]
История
[ редактировать ]Ген BRCA2 был открыт в 1994 году. [ 35 ] [ 16 ] [ 36 ] В 1996 году Кеннет Оффит и его исследовательская группа в Онкологическом центре Мемориала Слоан-Кеттеринг успешно идентифицировали наиболее распространенную мутацию гена, связанного с раком груди и яичников среди лиц еврейского происхождения ашкенази . [ 37 ] [ 38 ] [ 39 ] [ 40 ]
Ген был впервые клонирован учеными из Myriad Genetics , Endo Recherche, Inc., HSC Research & Development Limited Partnership и Пенсильванского университета . [ 41 ]
Методы диагностики вероятности развития рака у пациента с мутациями в BRCA1 и BRCA2 защищены патентами , принадлежащими или контролируемыми Myriad Genetics . [ 42 ] [ 43 ] Бизнес-модель Myriad, предлагающая исключительно диагностические тесты, привела к тому, что Myriad зародилась как стартап в 1994 году и превратилась в публичную компанию с 1200 сотрудниками и годовым доходом около 500 миллионов долларов в 2012 году; [ 44 ] это также привело к разногласиям по поводу высоких цен на тесты и отсутствия второго мнения из других диагностических лабораторий, что, в свою очередь, привело к знаковому иску «Ассоциация молекулярной патологии против Myriad Genetics» . [ 45 ]
Зародышевые мутации и эффект основателя
[ редактировать ]Все выявленные на сегодняшний день мутации BRCA2 зародышевой линии были унаследованы, что предполагает возможность большого эффекта «основателя», при котором определенная мутация является общей для четко определенной группы населения и теоретически может быть прослежена до общего предка. Учитывая сложность скрининга мутаций BRCA2, эти распространенные мутации могут упростить методы, необходимые для скрининга мутаций в определенных популяциях. Анализ мутаций, встречающихся с высокой частотой, также позволяет изучить их клиническое выражение. [ 46 ] Яркий пример мутации-основателя обнаружен в Исландии, где одна мутация BRCA2 (999del5) ответственна практически за все семейства рака молочной железы/яичников. [ 47 ] [ 48 ] Эта мутация сдвига рамки приводит к образованию сильно укороченного белкового продукта. В большом исследовании, в котором изучались сотни больных раком и контрольной группы, мутация 999del5 была обнаружена у 0,6% населения в целом. Следует отметить, что в то время как 72% пациентов, которые были признаны носителями, имели умеренный или сильный семейный анамнез рака молочной железы, у 28% семейный анамнез заболевания был незначительным или вообще отсутствовал. Это убедительно свидетельствует о наличии модифицирующих генов, влияющих на фенотипическую экспрессию этой мутации, или, возможно, о взаимодействии мутации BRCA2 с факторами окружающей среды. Дополнительные примеры мутаций-основателей BRCA2 приведены в таблице ниже.
| Население или подгруппа | Мутация(и) BRCA2 [ 46 ] [ 49 ] | Ссылка(и) |
|---|---|---|
| Ашкеназский еврей | 6174delT | [ 50 ] |
| Голландский | 5579insA | [ 51 ] |
| Доступный | 8555Т>Г, 999дел5, ИВС23-2А>Г | [ 52 ] [ 53 ] |
| Французские канадцы | 8765делАГ, 3398делААААГ | [ 54 ] [ 55 ] [ 56 ] |
| венгры | 9326insA | [ 57 ] |
| Исландцы | 999del5 | [ 47 ] [ 48 ] |
| итальянцы | 8765delAG | [ 58 ] |
| Северо-Ирландский | 6503delTT | [ 59 ] |
| Пакистанцы | 3337C>Т | [ 60 ] |
| шотландский | 6503delTT | [ 59 ] |
| шотландский | c.517-2A>G | [ 61 ] |
| Словенцы | ИВС16-2А>Г | [ 62 ] |
| испанский | 3034delAAAC(кодон936), 9254del5 | [ 63 ] |
| Шведский | 4486delG | [ 64 ] |
Мейоз
[ редактировать ]У растения Arabidopsis thaliana потеря BRCA2 гомолога AtBRCA2 вызывает серьезные дефекты как мужского мейоза , так и развития женских гаметоцитов . [ 65 ] Белок AtBRCA2 необходим для правильной локализации белка синаптонемного комплекса AtZYP1 и рекомбиназ AtRAD51 и AtDMC1. Более того, AtBRCA2 необходим для правильного мейотического синапса. Таким образом, AtBRCA2, вероятно, важен для мейотической рекомбинации. По-видимому, AtBRCA2 действует во время мейоза, контролируя этапы одноцепочечной инвазии, опосредованные AtRAD51 и AtDMC1, происходящие во время мейотической гомологичной рекомбинационной репарации повреждений ДНК. [ 65 ]
Гомологи BRCA2 также необходимы для мейоза у гриба Ustilago maydis . [ 66 ] червь Caenorhabditis elegans , [ 67 ] [ 68 ] и плодовая мушка Drosophila melanogaster . [ 69 ]
Мыши, производящие усеченные версии BRCA2, жизнеспособны, но стерильны. [ 70 ] Мутантные крысы BRCA2 имеют фенотип задержки роста и бесплодия у обоих полов. [ 71 ] Асперматогенез у этих мутантных крыс обусловлен нарушением синапса гомологичных хромосом во время мейоза.
Последовательности повторов BRC
[ редактировать ]DMC1 (ДНК-мейотическая рекомбиназа 1) представляет собой для мейоза специфичный гомолог RAD51 , который опосредует обмен цепей во время гомологичной рекомбинационной репарации. DMC1 способствует образованию продуктов инвазии цепи ДНК (совместных молекул) между гомологичными молекулами ДНК. Человеческий DMC1 напрямую взаимодействует с каждой из серии повторяющихся последовательностей в белке BRCA2 (называемых повторами BRC), которые стимулируют образование совместных молекул с помощью DMC1. [ 72 ] Повторы BRC соответствуют мотиву, состоящему из последовательности примерно из 35 высококонсервативных аминокислот, которые присутствуют хотя бы один раз во всех BRCA2-подобных белках. Повторы BRC BRCA2 стимулируют образование совместных молекул, способствуя взаимодействию одноцепочечной ДНК (оцДНК) с DMC1. [ 72 ] ОцДНК в комплексе с DMC1 может соединяться с гомологичной оцДНК из другой хромосомы во время синопсисной стадии мейоза с образованием совместной молекулы, что является центральным этапом гомологичной рекомбинации . Таким образом, последовательности повторов BRC BRCA2, по-видимому, играют ключевую роль в рекомбинационной репарации повреждений ДНК во время мейотической рекомбинации.
В целом, похоже, что гомологичная рекомбинация во время мейоза восстанавливает повреждения ДНК. [ нужна ссылка ] и что BRCA2 играет ключевую роль в выполнении этой функции.
Нейрогенез
[ редактировать ]BRCA2 необходим мышам для нейрогенеза и подавления медуллобластомы . [ 73 ] Потеря «BRCA2» глубоко влияет на нейрогенез, особенно во время эмбрионального и постнатального развития нейронов. Эти неврологические дефекты возникают в результате повреждения ДНК. [ 73 ]
Эпигенетический контроль
[ редактировать ]Эпигенетические изменения в экспрессии BRCA2 (вызывающие избыточную или недостаточную экспрессию) очень часто встречаются при спорадических формах рака (см. таблицу ниже), тогда как мутации BRCA2 обнаруживаются редко. [ 74 ] [ 75 ] [ 76 ]
При немелкоклеточном раке легкого BRCA2 эпигенетически репрессируется за счет гиперметилирования промотора. [ 77 ] В этом случае гиперметилирование промотора в значительной степени связано с низкой экспрессией мРНК и низкой экспрессией белка, но не с потерей гетерозиготности гена.
При спорадическом раке яичников обнаруживается противоположный эффект. Промотор BRCA2 и области 5'-UTR содержат относительно мало или вообще не содержат метилированных CpG-динуклеотидов в опухолевой ДНК по сравнению с таковой в неопухолевой ДНК, и обнаруживается значительная корреляция между гипометилированием и >3-кратной сверхэкспрессией BRCA2. [ 78 ] Это указывает на то, что гипометилирование промотора BRCA2 и областей 5'-UTR приводит к сверхэкспрессии мРНК BRCA2.
В одном сообщении указывается на некоторый эпигенетический контроль экспрессии BRCA2 с помощью микроРНК miR-146a и miR-148a. [ 79 ]
Экспрессия BRCA2 при раке
[ редактировать ]У эукариот белок BRCA2 играет важную роль в гомологичной рекомбинационной репарации. У мышей и людей BRCA2 в первую очередь опосредует упорядоченную сборку RAD51 на одноцепочечной (ss) ДНК, форме, которая активна для гомологичного спаривания и инвазии цепей. [ 80 ] BRCA2 также перенаправляет RAD51 из двухцепочечной ДНК и предотвращает диссоциацию от оцДНК. [ 80 ] Кроме того, четыре паралога RAD51 RAD51L3 , состоящие из RAD51B ( RAD51L1 ), RAD51C (RAD51L2), RAD51D ( ) , XRCC2, образуют комплекс, называемый комплексом BCDX2 (см. Рисунок: Рекомбинационная репарация ДНК). Этот комплекс участвует в рекрутировании или стабилизации RAD51 в местах повреждения. [ 26 ] Комплекс BCDX2, по-видимому, действует, облегчая сборку или стабильность нуклеопротеиновой нити RAD51 . RAD51 катализирует перенос цепи между разорванной последовательностью и ее неповрежденным гомологом, обеспечивая повторный синтез поврежденной области (см. Модели гомологичной рекомбинации ).
Некоторые исследования рака сообщают о сверхэкспрессии BRCA2 , тогда как другие исследования сообщают о недостаточной экспрессии BRCA2 . По крайней мере, в двух сообщениях обнаружена повышенная экспрессия в некоторых спорадических опухолях молочной железы и недостаточная экспрессия в других спорадических опухолях молочной железы. [ 81 ] [ 82 ] (см. таблицу).
Многие виды рака имеют эпигенетический дефицит различных генов репарации ДНК (см. Частота эпимутаций в генах репарации ДНК при раке ). Эти недостатки репарации, вероятно, вызывают увеличение невосстановленных повреждений ДНК. Сверхэкспрессия BRCA2, наблюдаемая при многих видах рака, может отражать компенсаторную BRCA2 сверхэкспрессию и усиление гомологичной рекомбинационной репарации, чтобы, по крайней мере, частично справиться с такими избыточными повреждениями ДНК. Эгава и др. [ 83 ] предполагают, что повышенная экспрессия BRCA2 может быть объяснена нестабильностью генома, часто наблюдаемой при раке, которая индуцирует экспрессию мРНК BRCA2 из-за повышенной потребности в BRCA2 для репарации ДНК.
Недостаточная экспрессия BRCA2 сама по себе приведет к увеличению невосстановленных повреждений ДНК. Ошибки репликации, выходящие за пределы этих повреждений (см. Синтез транслезий ), могут привести к увеличению числа мутаций и раку.
| Рак | Чрезмерное или недостаточное выражение | Частота измененного выражения | Метод оценки | Ссылка. |
|---|---|---|---|---|
| Спорадический рак яичников | Чрезмерное выражение | 80% | информационная РНК | [ 78 ] |
| Спорадический рак яичников | Недовыражение | 42% | иммуногистохимия | [ 84 ] |
| (рецидивирующий рак в исследовании выше) | Повышенное выражение | 71% | иммуногистохимия | [ 84 ] |
| Немелкоклеточный рак легкого | Недовыражение | 34% | иммуногистохимия | [ 77 ] |
| Рак молочной железы | Чрезмерное выражение | 66% | информационная РНК | [ 83 ] |
| Рак молочной железы | Чрезмерное выражение | 20% | информационная РНК | [ 81 ] |
| (то же исследование, что и выше) | Недовыражение | 11% | информационная РНК | [ 81 ] |
| Рак молочной железы | Чрезмерное выражение | 30% | иммуногистохимия | [ 82 ] |
| (то же исследование, что и выше) | Недовыражение | 30% | иммуногистохимия | [ 82 ] |
| Трижды негативный рак молочной железы | Недовыражение | 90% | иммуногистохимия | [ 85 ] |
Взаимодействия
[ редактировать ]Было показано, что BRCA2 взаимодействует с
- БРЭ , [ 86 ]
- БАРД1 , [ 86 ] [ 87 ]
- БЦИП , [ 88 ]
- БРЦА1 , [ 86 ] [ 89 ] [ 90 ] [ 91 ]
- БРЦЦ3 , [ 86 ]
- БУБ1Б , [ 92 ]
- КРЕББП , [ 93 ]
- C11orf30 , [ 94 ]
- ФАНКД2 , [ 95 ] [ 96 ] [ 97 ]
- ФАНЧГ , [ 98 ]
- ФЛНА , [ 99 ]
- ХМГ20Б , [ 100 ] [ 101 ]
- Р53 , [ 86 ] [ 102 ]
- ПАЛБ2 , [ 29 ] [ 103 ]
- ПКАФ , [ 104 ] [ 105 ]
- ПЛК1 , [ 104 ] [ 106 ]
- РАД51 , [ 86 ] [ 89 ] [ 104 ] [ 107 ] [ 108 ] [ 109 ] [ 110 ] [ 111 ] [ 112 ] [ 113 ] [ 88 ] [ 90 ] [ 102 ]
- РПА1 , [ 114 ]
- ШФМ1 [ 115 ] [ 116 ] и
- СМАД3 . [ 117 ]
Доменная архитектура
[ редактировать ]| BRCA2 повтор | |||
|---|---|---|---|
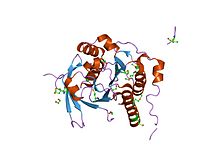 кристаллическая структура повторяющегося комплекса rad51-brca2 brc | |||
| Идентификаторы | |||
| Символ | БРЦА2 | ||
| Пфам | PF00634 | ||
| ИнтерПро | ИПР002093 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 1n0w / СКОПе / СУПФАМ | ||
| |||
BRCA2 содержит ряд из 39 аминокислотных повторов , которые имеют решающее значение для связывания с RAD51 (ключевого белка в рекомбинационной репарации ДНК) и устойчивости к лечению метилметансульфонатом. [ 102 ] [ 109 ] [ 110 ] [ 118 ]
Спиральный домен BRCA2 имеет спиральную структуру, состоящую из ядра кластера из четырех спиралей (альфа 1, альфа 8, альфа 9, альфа 10) и двух последовательных бета-шпильок (от бета 1 до бета 4). Сегмент, содержащий примерно 50 аминокислот и содержащий четыре короткие спирали (от альфа-2 до альфа-4), извивается вокруг поверхности основной структуры . В BRCA2 спирали альфа-9 и альфа-10 упаковываются с доменом OB1 BRCA2 посредством ван-дер-ваальсовых контактов, включающих гидрофобные и ароматические остатки, а также посредством боковой цепи и основной цепи водородных связей . Этот домен связывает из 70 аминокислот белок DSS1 (удаленный при синдроме разделенных рук/разделенных ног), который первоначально был идентифицирован как один из трех генов , которые картируются с локусом размером 1,5 Мб , удаленным при синдроме наследственного порока развития. [ 116 ]
Домен BRCA OB1 предполагает складку OB, которая состоит из сильно изогнутого пятинитевого бета-листа , который замыкается сам на себя, образуя бета-цилиндр . OB1 имеет неглубокую бороздку, образованную одной стороной изогнутого листа и разграниченную двумя петлями: одну между бета-1 и бета-2, а другую между бета-4 и бета-5, что обеспечивает слабое связывание одноцепочечной ДНК . Домен также связывает из 70 аминокислот белок DSS1 (удаленный при синдроме разделенных рук/разделенных ног). [ 116 ]
Домен BRCA OB3 предполагает складку OB, которая состоит из сильно изогнутого пятинитевого бета-листа , который замыкается сам на себя, образуя бета-цилиндр . OB3 имеет выраженную бороздку, образованную одной стороной изогнутого листа и разграниченную двумя петлями: одну между бета-1 и бета-2, а другую между бета-4 и бета-5, что обеспечивает прочное оцДНК связывание . [ 116 ]
Домен Башни имеет вторичную структуру, состоящую из пары длинных антипараллельных альфа-спиралей (стебель), которые поддерживают на своих концах пучок трех спиралей (3HB). 3HB содержит спираль-поворот-спираль и подобен ДНК- связывающим доменам бактериальных мотив сайт-специфических рекомбиназ , а также эукариотических Myb и гомеодоменовых факторов транскрипции . Домен Tower играет важную роль в функции подавления опухолей BRCA2 и необходим для соответствующего связывания BRCA2 с ДНК. [ 116 ] Исследования показали, что конформация этого домена башни аллостерически контролируется небольшим белком «DSS1», который взаимодействует со спиральными доменами OB1 и OB2 BRCA2. [ 119 ]
Патенты, правоприменение, судебные разбирательства и споры
[ редактировать ]Заявка на патент на изолированный ген BRCA1 и мутации, способствующие раку, а также на методы диагностики вероятности заболевания раком молочной железы была подана Университетом Юты, Национальным институтом наук об охране окружающей среды (NIEHS) и Myriad Genetics в 1994 году. ; [ 42 ] В течение следующего года Myriad в сотрудничестве с другими исследователями выделила и секвенировала ген BRCA2 и выявила соответствующие мутации, а первый патент на BRCA2 был подан в США Myriad и другими учреждениями в 1995 году. [ 41 ] Myriad является эксклюзивным лицензиатом этих патентов и применяет их в США в отношении клинических диагностических лабораторий. [ 45 ] Эта бизнес-модель привела Myriad от стартапа в 1994 году к публичной компании с 1200 сотрудниками и годовым доходом около 500 миллионов долларов в 2012 году; [ 44 ] это также привело к разногласиям по поводу высоких цен и невозможности получить второе мнение из других диагностических лабораторий, что, в свою очередь, привело к знаковому иску «Ассоциация молекулярной патологии против Myriad Genetics» . [ 45 ] [ 120 ] Срок действия патентов истекает в 2014 году.
Питер Мелдрам, генеральный директор Myriad Genetics, признал, что у Myriad есть «другие конкурентные преимущества, которые могут сделать такое [патентное] обеспечение ненужным» в Европе. [ 121 ]
Юридические решения, касающиеся патентов BRCA1 и BRCA2, повлияют на область генетического тестирования в целом. [ 122 ] В июне 2013 года в деле «Ассоциация молекулярной патологии против Myriad Genetics» (№ 12-398) Верховный суд США единогласно постановил, что «естественный сегмент ДНК является продуктом природы и не подлежит патентованию только потому, что он был изолирован». », аннулируя патенты Myriad на гены BRCA1 и BRCA2. Однако Суд также постановил, что манипуляция геном с целью создания чего-то, не встречающегося в природе, все еще может иметь право на патентную защиту. [ 123 ] Федеральный суд Австралии пришел к противоположному выводу, подтвердив в феврале 2013 года действительность австралийского патента Myriad Genetics на ген BRCA1. [ 124 ] но это решение обжалуется, и апелляция будет включать рассмотрение решения Верховного суда США. [ 125 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000139618 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000041147 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Хамель П.Дж. (29 мая 2007 г.). «BRCA1 и BRCA2: больше не единственные проблемные гены» . Центр здоровья . Проверено 2 июля 2010 г.
- ^ «Филогенетический маркер OrthoMaM: кодирующая последовательность BRCA2» . Архивировано из оригинала 3 марта 2016 г. Проверено 19 февраля 2010 г.
- ^ «Геновое дерево BRCA2» . ансамбль май 2021 г.
- ^ Дункан Дж.А., Ривз-младший, Кук Т.Г. (октябрь 1998 г.). «Белки BRCA1 и BRCA2: роль в здоровье и болезни» . Молекулярная патология . 51 (5): 237–47. дои : 10.1136/mp.51.5.237 . ПМЦ 395646 . ПМИД 10193517 .
- ^ Ёсида К., Мики Ю (ноябрь 2004 г.). «Роль BRCA1 и BRCA2 как регуляторов репарации ДНК, транскрипции и клеточного цикла в ответ на повреждение ДНК» . Раковая наука . 95 (11): 866–71. дои : 10.1111/j.1349-7006.2004.tb02195.x . ПМЦ 11159131 . ПМИД 15546503 . S2CID 24297965 .
- ^ Проверьте W (01 сентября 2006 г.). «BRCA: Что мы знаем сейчас» . Колледж американских патологов . Проверено 23 августа 2010 г.
- ^ Фриденсон Б. (август 2007 г.). «Путь BRCA1/2 предотвращает гематологический рак в дополнение к раку молочной железы и яичников» . БМК Рак . 7 (1): 152–162. дои : 10.1186/1471-2407-7-152 . ЧВК 1959234 . ПМИД 17683622 .
- ^ Фриденсон Б (8 июня 2008 г.). «Гены рака молочной железы защищают от некоторых лейкозов и лимфом» (видео) . SciVee.
- ^ «Генетический скрининг рака молочной железы и яичников» . Медицинский фонд Пало-Альто. Архивировано из оригинала 4 октября 2008 года . Проверено 11 октября 2008 г.
- ^ Фриденсон Б. (2007). «Путь BRCA1/2 предотвращает гематологический рак в дополнение к раку молочной железы и яичников» . БМК Рак . 7 (1): 152. дои : 10.1186/1471-2407-7-152 . ЧВК 1959234 . ПМИД 17683622 .
- ^ О'Донован П.Дж., Ливингстон DM (апрель 2010 г.). «BRCA1 и BRCA2: продукты генов предрасположенности к раку молочной железы/яичников и участники восстановления двухцепочечных разрывов ДНК» . Канцерогенез . 31 (6): 961–7. дои : 10.1093/carcin/bgq069 . ПМИД 20400477 .
- ^ Перейти обратно: а б Вустер Р., Нойхаузен С.Л., Мангион Дж., Квирк Ю., Форд Д., Коллинз Н. и др. (сентябрь 1994 г.). «Локализация гена предрасположенности к раку молочной железы, BRCA2, на хромосоме 13q12-13». Наука . 265 (5181): 2088–90. Бибкод : 1994Sci...265.2088W . дои : 10.1126/science.8091231 . ПМИД 8091231 .
- ^ «Рак молочной железы BRCA2 2, раннее начало [Homo sapiens]» . ЭнтрезГен . Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ «Белок восприимчивости к раку молочной железы 2 типа - Homo sapiens (Human)» . Р51587 . ЮниПрот.
- ^ Уильямс-Джонс Б. (2002). Генетическое тестирование на продажу: последствия коммерческого тестирования brca в Канаде (доктор философии). Университет Британской Колумбии.
- ^ Д'Андреа А.Д. (2010). «Пути восприимчивости при анемии Фанкони и раке молочной железы» . Н. англ. Дж. Мед . 362 (20): 1909–19. дои : 10.1056/NEJMra0809889 . ПМК 3069698 . ПМИД 20484397 .
- ^ Собек А., Стоун С., Ландэ И., де Грааф Б., Хоатлин М.Э. (2009). «Белок анемии Фанкони FANCM контролируется FANCD2 и путями ATR/ATM» . Ж. Биол. Хим . 284 (38): 25560–8. дои : 10.1074/jbc.M109.007690 . ПМЦ 2757957 . ПМИД 19633289 .
- ^ Кастильо П., Больоло М., Сюрраллес Дж. (2011). «Координированное действие путей анемии Фанкони и атаксии-телеангиэктазии в ответ на окислительное повреждение». Восстановление ДНК (Амст.) . 10 (5): 518–25. дои : 10.1016/j.dnarep.2011.02.007 . ПМИД 21466974 .
- ^ Штольц А., Эртыч Н., Бастианс Х (2011). «Опухолевый супрессор CHK2: регулятор реакции на повреждение ДНК и медиатор хромосомной стабильности» . Клин. Рак Рез . 17 (3): 401–5. дои : 10.1158/1078-0432.CCR-10-1215 . ПМИД 21088254 .
- ^ Танигучи Т., Гарсия-Игера И., Андреассен П.Р., Грегори Р.К., Громпе М., Д'Андреа А.Д. (2002). «S-фазоспецифическое взаимодействие белка анемии Фанкони, FANCD2, с BRCA1 и RAD51» . Кровь . 100 (7): 2414–20. дои : 10.1182/кровь-2002-01-0278 . ПМИД 12239151 .
- ^ Пак JY, Чжан Ф, Андреассен PR (2014). «PALB2: центр сети супрессоров опухолей, участвующих в реакциях на повреждение ДНК» . Биохим. Биофиз. Акта . 1846 (1): 263–75. дои : 10.1016/j.bbcan.2014.06.003 . ПМЦ 4183126 . ПМИД 24998779 .
- ^ Перейти обратно: а б Чун Дж., Бючелмайер Э.С., Пауэлл С.Н. (2013). «Паралоговые комплексы Rad51 BCDX2 и CX3 действуют на разных стадиях BRCA1-BRCA2-зависимого пути гомологичной рекомбинации» . Мол. Клетка. Биол . 33 (2): 387–95. дои : 10.1128/MCB.00465-12 . ПМЦ 3554112 . ПМИД 23149936 .
- ^ Йенсен РБ, Каррейра А, Ковальчиковски СК (октябрь 2010 г.). «Очищенный человеческий BRCA2 стимулирует рекомбинацию, опосредованную RAD51» . Природа . 467 (7316): 678–83. Бибкод : 2010Natur.467..678J . дои : 10.1038/nature09399 . ПМК 2952063 . ПМИД 20729832 .
- ^ Ван С.Х., Хименес-Сайнс Дж., Дженсен Р.Б., Мазин А.В. (март 2019 г.). «Постсинаптическая функция Brca2» . Научные отчеты . 9 (1): 4554. Бибкод : 2019NatSR...9.4554W . дои : 10.1038/s41598-019-41054-y . ПМК 6418147 . ПМИД 30872704 .
- ^ Перейти обратно: а б Ся Б., Шэн К., Наканиши К., Охаши А., Ву Дж., Крист Н. и др. (июнь 2006 г.). «Контроль клеточных и клинических функций BRCA2 с помощью ядерного партнера PALB2» . Молекулярная клетка . 22 (6): 719–29. doi : 10.1016/j.molcel.2006.05.022 . ПМИД 16793542 .
- ^ Бюиссон Р., Дион-Кот А.М., Куломб И., Лоне Х., Кай Х., Стасиак А.З. и др. (октябрь 2010 г.). «Сотрудничество белков рака молочной железы PALB2 и пикколо BRCA2 в стимуляции гомологичной рекомбинации» . Структурная и молекулярная биология природы . 17 (10): 1247–54. дои : 10.1038/nsmb.1915 . ПМК 4094107 . ПМИД 20871615 .
- ^ Михик С., Зеллвегер Р., Чаппиди Н., Берти М., Джейкобс К., Мутрея К. и др. (октябрь 2017 г.). «Реверс репликационной вилки вызывает деградацию вилки в BRCA2-дефектных клетках» . Природные коммуникации . 8 (1): 859. Бибкод : 2017NatCo...8..859M . дои : 10.1038/s41467-017-01164-5 . ПМЦ 5643541 . ПМИД 29038466 .
- ^ Петручелли Н., Дейли М.Б., Пал Т. (декабрь 2016 г.) [сентябрь 1998 г.]. «BRCA1- и BRCA2-ассоциированный наследственный рак молочной железы и яичников» . Адам М.П., Ардингер Х.Х., Пагон Р.А., Уоллес С.Е., Бин Л.Дж., Мирзаа Г., Амемия А. (ред.). Джин Обзоры . Вашингтонский университет, Сиэтл. ПМИД 20301425 .
- ^ Левин Б., Лех Д., Фриденсон Б. (2012). «Доказательства того, что рак, связанный с BRCA1 или BRCA2, не является неизбежным» . Молекулярная медицина . 18 (9): 1327–37. дои : 10.2119/molmed.2012.00280 . ПМЦ 3521784 . ПМИД 22972572 .
- ^ «Гены предрасположенности к раку молочной железы и/или яичников с высокой пенетрантностью» . Национальный институт рака . Проверено 7 декабря 2012 г.
- ^ Вустер Р., Бигнелл Дж., Ланкастер Дж., Свифт С., Сил С., Мангион Дж. и др. (1995). «Идентификация гена предрасположенности к раку молочной железы BRCA2». Природа . 378 (6559): 789–792. Бибкод : 1995Natur.378..789W . дои : 10.1038/378789a0 . ПМИД 8524414 . S2CID 4346791 .
- ^ Высокоэффективная наука: отслеживание генов BRCA (Часть 2). Архивировано 3 марта 2012 г. в Wayback Machine - научный блог Cancer Research UK, 2012 г.
- ^ «Кеннет Оффит | Фонд исследования рака молочной железы | BCRF» . Bcrfcure.org. 23 июня 2014 года . Проверено 16 июля 2015 г.
- ^ «Революция в 50 лет; Кеннет Оффит» . Нью-Йорк Таймс . 25 февраля 2003 г. ISSN 0362-4331 . Проверено 2 июля 2015 г.
- ^ «20 лет прогресса в понимании рака молочной железы» (JPG) . Mskcc.org . Проверено 17 июля 2015 г.
- ^ Колата Г (02 октября 1996 г.). «2-й ген рака молочной железы обнаружен у еврейских женщин» . Нью-Йорк Таймс . ISSN 0362-4331 . Проверено 7 июля 2015 г.
- ^ Перейти обратно: а б Патент США 5837492 , Тавтигян С.В., Камб А., Симард Дж., Коуч Ф., Ромменс Дж.М., Вебер Б.Л., «Ген предрасположенности к раку молочной железы, связанный с хромосомой 13», выдан 17 ноября 1998 г., передан Myriad Genetics, Inc., Endo Recherche. , Inc., Товарищество с ограниченной ответственностью HSC Research & Development, Попечители Пенсильванского университета
- ^ Перейти обратно: а б Патент США 5747282 , Сколник Х.С., Голдгар Д.Е., Мики Й., Свенсон Дж., Камб А., Харшман К.Д., Шаттук-Эйденс Д.М., Тавтигян С.В., Уайзман Р.В., Фуреал П.А., «7Q-связанный ген предрасположенности к раку молочной железы и яичников», выдан в 1998 г. -05-05, закреплено за Myriad Genetics, Inc., Соединенные Штаты Америки в лице министра здравоохранения и социальных служб, и Исследовательского фонда Университета Юты.
- ^ Патент США 5837492 , Тавтигян С.В., Камб А., Симард Дж., Коуч Ф., Ромменс Дж.М., Вебер Б.Л., «Ген предрасположенности к раку молочной железы, связанный с хромосомой 13», выдан 17 ноября 1998 г., передан Myriad Genetics, Inc., Endo Recherche. , Inc., Товарищество с ограниченной ответственностью HSC Research & Development, Попечители Пенсильванского университета
- ^ Перейти обратно: а б Страница Myriad Investor - см. «Мириады с первого взгляда». Архивировано 18 октября 2012 г. на Wayback Machine , доступ октябрь 2012 г.
- ^ Перейти обратно: а б с Шварц Дж. (12 мая 2009 г.). «Больные раком бросают вызов патентованию гена» . Здоровье . Нью-Йорк Таймс.
- ^ Перейти обратно: а б Лакруа М., Леклерк Г. (2005). «Портрет» наследственного рака молочной железы». Исследование и лечение рака молочной железы . 89 (3): 297–304. дои : 10.1007/s10549-004-2172-4 . ПМИД 15754129 . S2CID 23327569 .
- ^ Перейти обратно: а б Торлациус С., Олафсдоттир Г., Трюггвадоттир Л., Нойхаузен С., Йонассон Дж.Г., Тавтигян С.В. и др. (1996). «Одна мутация BRCA2 в семьях мужчин и женщин с раком молочной железы из Исландии с различными фенотипами рака». Природная генетика . 13 (1): 117–119. дои : 10.1038/ng0596-117 . ПМИД 8673089 . S2CID 8443452 .
- ^ Перейти обратно: а б Торлациус С., Сигурдссон С., Бьярнадоттир Х., Олафсдоттир Г., Йонассон Дж.Г., Триггвадоттир Л. и др. (1997). «Исследование одиночной мутации BRCA2 с высокой частотой носительства в небольшой популяции» . Американский журнал генетики человека . 60 (5): 1079–1085. ПМЦ 1712443 . ПМИД 9150155 .
- ^ ден Даннен Дж.Т., Антонаракис С.Е. (2000). «Расширение номенклатуры мутаций и предложения для описания сложных мутаций: обсуждение» . Человеческая мутация . 15 (1): 7–12. doi : 10.1002/(SICI)1098-1004(200001)15:1<7::AID-HUMU4>3.0.CO;2-N . ПМИД 10612815 .
- ^ Нойхаузен С., Гилевски Т., Нортон Л., Тран Т., Макгуайр П., Свенсен Дж. и др. (1996). «Рецидивирующие мутации BRCA2 6174delT у еврейских женщин-ашкенази, больных раком молочной железы». Природная генетика . 13 (1): 126–128. дои : 10.1038/ng0596-126 . ПМИД 8673092 . S2CID 11909356 .
- ^ Верхоог Л.С., ван ден Оувеланд А.М., Бернс Э., ван Вегел-Пландсоен М.М., ван Ставерен И.Л., Вагнер А. и др. (2001). «Большие региональные различия в частоте различных мутаций BRCA1/BRCA2 в 517 голландских семьях рака молочной железы и / или яичников». Европейский журнал рака . 37 (16): 2082–2090. дои : 10.1016/S0959-8049(01)00244-1 . ПМИД 11597388 .
- ^ Хууско П., Пяакконен К., Лаунонен В., Пёйхёнен М., Бланко Г., Кауппила А. и др. (1998). «Доказательства мутаций-основателей финских семейств BRCA1 и BRCA2» . Американский журнал генетики человека . 62 (6): 1544–1548. дои : 10.1086/301880 . ПМЦ 1377159 . ПМИД 9585608 .
- ^ Пяакконен К., Саурамо С., Сарантаус Л., Вахтеристо П., Хартикайнен А., Вехманен П. и др. (2001). «Вовлечение BRCA1 и BRCA2 в развитие рака молочной железы в западной части населения Финляндии». Генетическая эпидемиология . 20 (2): 239–246. doi : 10.1002/1098-2272(200102)20:2<239::AID-GEPI6>3.0.CO;2-Y . ПМИД 11180449 . S2CID 41804152 .
- ^ Тонин П.Н., Мес-Массон А.М., Народ С.А., Гадирян П., Прованшер Д. (1999). «Мутации-основатели BRCA1 и BRCA2 во франко-канадских случаях рака яичников, не выбранные для семейного анамнеза». Клиническая генетика . 55 (5): 318–324. дои : 10.1034/j.1399-0004.1999.550504.x . ПМИД 10422801 . S2CID 23931343 .
- ^ Орос К.К., Леблан Г., Арканд С.Л., Шен З., Перре С., Мес-Массон А.М. и др. (2006). «Анализ гаплотипов предполагает наличие общих основателей у носителей рецидивирующей мутации BRCA2, 3398delAAAAG, во франко-канадских семьях с наследственным раком молочной железы и/или яичников» . BMC Медицинская генетика . 7 (23): 23. дои : 10.1186/1471-2350-7-23 . ПМК 1464093 . ПМИД 16539696 .
- ^ Тонин П.Н. (2006). «Ограниченный спектр патогенных мутаций BRCA1 и BRCA2 во франко-канадских семьях рака молочной железы и молочной железы яичников, основной популяции в Квебеке, Канада». Бык-Рак . 93 (9): 841–846. ПМИД 16980226 .
- ^ Ван дер Лоой М., Сабо С., Бесняк И., Лишка Г., Чокай Б., Пулай Т. и др. (2000). «Распространенность мутаций-основателей BRCA1 и BRCA2 среди больных раком молочной железы и яичников в Венгрии» . Международный журнал рака . 86 (5): 737–740. doi : 10.1002/(SICI)1097-0215(20000601)86:5<737::AID-IJC21>3.0.CO;2-1 . ПМИД 10797299 . S2CID 25394976 .
- ^ Пизано М., Коссу А., Персико I, Палмьери Г., Ангиус А., Касу Г. и др. (2000). «Идентификация мутации-основателя BRCA2 на Сардинии» . Британский журнал рака . 82 (3): 553–559. дои : 10.1054/bjoc.1999.0963 . ПМК 2363305 . ПМИД 10682665 .
- ^ Перейти обратно: а б Шотландско-Северо-Ирландский консорциум BRCAI/BRCA2 (2003 г.). «Мутации BRCA1 и BRCA2 в Шотландии и Северной Ирландии» . Британский журнал рака . 88 (8): 1256–1262. дои : 10.1038/sj.bjc.6600840 . ПМЦ 2747571 . ПМИД 12698193 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Лиеде А., Малик И.А., Азиз З., Риос Пд Пде Л., Кван Э., Народ С.А. (2002). «Вклад мутаций BRCA1 и BRCA2 в рак молочной железы и яичников в Пакистане» . Американский журнал генетики человека . 71 (3): 595–606. дои : 10.1086/342506 . ПМК 379195 . ПМИД 12181777 .
- ^ Керр С.М., Кларик Л., Макиан М.Д., Коуэн Э., Снадден Л., Цонева Г. и др. (2024). «Два варианта-основателя составляют более 90% патогенных аллелей BRCA на Оркнейских и Шетландских островах». medRxiv 10.1101/2024.04.03.24305239 .
- ^ Крайц М., Де Грев Дж., Гоелен Г., Теугельс Э. (2002). «Мутация-основатель BRCA2 в словенских семьях, больных раком молочной железы» . Европейский журнал генетики человека . 10 (12): 879–882. дои : 10.1038/sj.ejhg.5200886 . ПМИД 12461697 .
- ^ Осорио А., Робледо М., Мартинес Б., Себриан А., Сан-Роман Х.М., Альбертос Дж. и др. (1998). «Молекулярный анализ гена BRCA2 в 16 испанских семьях с раком молочной железы и яичников». Клин. Жене . 54 (2): 142–7. дои : 10.1111/j.1399-0004.1998.tb03717.x . ПМИД 9761393 . S2CID 30388365 .
- ^ Нойхаузен С.Л. (2000). «Популяции-основатели и их использование в генетике рака молочной железы» . Исследования рака . 2 (2): 77–81. дои : 10.1186/bcr36 . ПМК 139426 . ПМИД 11250694 .
- ^ Перейти обратно: а б Зилигер К., Дукович-Шульце С., Вурц-Вильдерсинн Р., Пахер М., Пухта Х (2012). «BRCA2 является медиатором гомологичной рекомбинации, облегчаемой RAD51 и DMC1, у Arabidopsis thaliana» . Новый Фитол . 193 (2): 364–75. дои : 10.1111/j.1469-8137.2011.03947.x . ПМИД 22077663 .
- ^ Койич М., Коструб К.Ф., Бухман А.Р., Холломан В.К. (2002). «Гомолог BRCA2 необходим для навыков репарации ДНК, рекомбинации и стабильности генома Ustilago maydis» . Мол. Клетка . 10 (3): 683–91. дои : 10.1016/s1097-2765(02)00632-9 . ПМИД 12408834 .
- ^ Ко Э, Ли Дж, Ли Х (2008). «Основная роль brc-2 в целостности хромосом зародышевых клеток C. elegans» . Мол. Клетки . 26 (6): 590–4. дои : 10.1016/S1016-8478(23)14041-6 . ПМИД 18779660 .
- ^ Мартин Дж.С., Винкельманн Н., Петалкорин М.И., Макилрайт М.Дж., Бултон С.Дж. (2005). «RAD-51-зависимая и независимая роль белка, родственного BRCA2 Caenorhabditis elegans, во время восстановления двухцепочечного разрыва ДНК» . Мол. Клетка. Биол . 25 (8): 3127–39. дои : 10.1128/MCB.25.8.3127-3139.2005 . ПМЦ 1069622 . ПМИД 15798199 .
- ^ Кловстад М., Абду У., Шюпбах Т. (2008). «Brca2 дрозофилы необходим для восстановления митотической и мейотической ДНК и эффективной активации контрольной точки мейотической рекомбинации» . ПЛОС Генет . 4 (2): е31. дои : 10.1371/journal.pgen.0040031 . ПМЦ 2233675 . ПМИД 18266476 .
- ^ Коннор Ф., Бертвистл Д., Ми П.Дж., Росс Г.М., Свифт С., Григорьева Е. и др. (1997). «Онкогенез и дефект репарации ДНК у мышей с укороченной мутацией Brca2». Нат. Жене . 17 (4): 423–30. дои : 10.1038/ng1297-423 . ПМИД 9398843 . S2CID 42462448 .
- ^ Котронео М.С., Хааг Дж.Д., Зан Й., Лопес К.С., Туваджит П., Петухова Г.В. и др. (2007). «Характеристика крысиной модели нокаута Brca2» . Онкоген . 26 (11): 1626–35. дои : 10.1038/sj.onc.1209960 . ПМИД 16964288 .
- ^ Перейти обратно: а б Мартинес Дж.С., фон Николай С., Ким Т., Элен О., Мазин А.В., Ковальчиковский С.С. и др. (2016). «BRCA2 регулирует рекомбинацию, опосредованную DMC1, посредством повторов BRC» . Учеб. Натл. акад. наук. США . 113 (13): 3515–20. Бибкод : 2016PNAS..113.3515M . дои : 10.1073/pnas.1601691113 . ПМЦ 4822569 . ПМИД 26976601 .
- ^ Перейти обратно: а б Фраппарт П.О., Ли Ю., Ламонт Дж., Маккиннон П.Дж. (2007). «BRCA2 необходим для нейрогенеза и подавления медуллобластомы» . ЭМБО Дж . 26 (11): 2732–42. дои : 10.1038/sj.emboj.7601703 . ПМК 1888666 . ПМИД 17476307 .
- ^ Тенг Д.Х., Богден Р., Митчелл Дж., Баумгард М., Белл Р., Берри С. и др. (1996). «Низкая частота мутаций BRCA2 при раке молочной железы и других видах рака». Нат. Жене . 13 (2): 241–4. дои : 10.1038/ng0696-241 . ПМИД 8640236 . S2CID 9831745 .
- ^ Мики Ю, Катагири Т, Касуми Ф, Ёсимото Т, Накамура Ю (1996). «Анализ мутаций гена BRCA2 при первичном раке молочной железы». Нат. Жене 13 (2): 245–7. дои : 10.1038/ng0696-245 . ПМИД 8640237 . S2CID 3203046 .
- ^ Ланкастер Дж. М., Вустер Р., Мангион Дж., Фелан С. М., Кокран С., Гамбс С. и др. (1996). «Мутации BRCA2 при первичном раке молочной железы и яичников». Нат. Жене . 13 (2): 238–40. дои : 10.1038/ng0696-238 . ПМИД 8640235 . S2CID 26808443 .
- ^ Перейти обратно: а б Ли М.Н., Ценг Р.К., Сюй Х.С., Чен Дж.Ю., Цао С., Хо В.Л. и др. (2007). «Эпигенетическая инактивация генов контроля хромосомной стабильности BRCA1, BRCA2 и XRCC5 при немелкоклеточном раке легкого» . Клин. Рак Рез . 13 (3): 832–8. дои : 10.1158/1078-0432.CCR-05-2694 . ПМИД 17289874 .
- ^ Перейти обратно: а б Чан К.Ю., Озчелик Х., Чунг А.Н., Нган Х.И., Ху США (2002). «Эпигенетические факторы, контролирующие гены BRCA1 и BRCA2 при спорадическом раке яичников». Рак Рез . 62 (14): 4151–6. ПМИД 12124354 .
- ^ Гу Ю, Чжан М, Пэн Ф, Фанг Л, Чжан Ю, Лян Х и др. (2015). «Сигнатура микроРНК, направленная на BRCA1/2, предсказывает хороший прогноз у больных раком яичников с BRCA1/2 дикого типа» . Онкотаргет . 6 (4): 2397–406. дои : 10.18632/oncotarget.2963 . ПМЦ 4385859 . ПМИД 25537514 .
- ^ Перейти обратно: а б Холломан В.К. (2011). «Разгадка механизма гомологичной рекомбинации BRCA2» . Нат. Структура. Мол. Биол . 18 (7): 748–54. дои : 10.1038/nsmb.2096 . ПМЦ 3647347 . ПМИД 21731065 .
- ^ Перейти обратно: а б с Бьеш I, Ноге С, Лидеро Р (1999). «Сверхэкспрессия гена BRCA2 в спорадических опухолях молочной железы» . Онкоген . 18 (37): 5232–8. дои : 10.1038/sj.onc.1202903 . ПМИД 10498873 .
- ^ Перейти обратно: а б с Хедау С., Батра М., Сингх УР, Бхарти AC, Рэй А., Дас BC (2015). «Экспрессия белков BRCA1 и BRCA2 и их корреляция с клинической стадией рака молочной железы» . J Рак Res Ther . 11 (1): 158–63. дои : 10.4103/0973-1482.140985 . ПМИД 25879355 .
- ^ Перейти обратно: а б Эгава С., Миёси Ю., Тагучи Т., Тамаки Ю., Ногучи С. (2002). «Высокая экспрессия мРНК BRCA2 предсказывает плохой прогноз у пациентов с раком молочной железы» . Межд. Дж. Рак . 98 (6): 879–82. дои : 10.1002/ijc.10231 . ПМИД 11948466 . S2CID 9083282 .
- ^ Перейти обратно: а б Свишер Э.М., Гонсалес Р.М., Танигучи Т., Гарсия Р.Л., Уолш Т., Гофф Б.А. и др. (2009). «Метилирование и экспрессия белков генов репарации ДНК: связь с воздействием химиотерапии и выживаемостью при спорадических карциномах яичников и брюшины» . Мол. Рак . 8 (1): 48. дои : 10.1186/1476-4598-8-48 . ПМК 2719582 . ПМИД 19602291 .
- ^ Тике А.А., Тан П.Х., Икеда М., Икбал Дж. (2016). «Повышение экспрессии ID4, сопровождающееся накоплением мутантного р53 и потерей белков BRCA1/2 при трижды негативном раке молочной железы, отрицательно влияет на выживаемость». Гистопатология . 68 (5): 702–12. дои : 10.1111/его.12801 . ПМИД 26259780 . S2CID 3566545 .
- ^ Перейти обратно: а б с д и ж Донг Ю, Хакими М.А., Чен Х, Кумарасвами Э, Куч Н.С., Годвин А.К. и др. (ноябрь 2003 г.). «Регуляция BRCC, голоферментного комплекса, содержащего BRCA1 и BRCA2, с помощью сигналосомоподобной субъединицы и ее роль в репарации ДНК» . Мол. Клетка . 12 (5): 1087–99. дои : 10.1016/S1097-2765(03)00424-6 . ПМИД 14636569 .
- ^ Райсер С., Дизин Е., Джеффорд С.Э., Делаваль Б., Гагос С., Христодулиду А. и др. (февраль 2009 г.). «Различные роли изоформ BARD1 в митозе: полноразмерный BARD1 опосредует деградацию Aurora B, связанные с раком каркасы BARD1beta Aurora B и BRCA2» . Рак Рез . 69 (3): 1125–34. дои : 10.1158/0008-5472.CAN-08-2134 . ПМИД 19176389 .
- ^ Перейти обратно: а б Лю Дж, Юань Ю, Хуань Дж, Шэнь З (январь 2001 г.). «Ингибирование роста клеток рака молочной железы и мозга с помощью BCCIPalpha, эволюционно консервативного ядерного белка, который взаимодействует с BRCA2» . Онкоген . 20 (3): 336–45. дои : 10.1038/sj.onc.1204098 . ПМИД 11313963 .
- ^ Перейти обратно: а б Саркисян CJ, Мастер SR, Хубер LJ, Ха С.И., Чодош Л.А. (октябрь 2001 г.). «Анализ мышиного Brca2 показывает сохранение межбелковых взаимодействий, но различия в сигналах ядерной локализации» . Ж. Биол. Хим . 276 (40): 37640–8. дои : 10.1074/jbc.M106281200 . ПМИД 11477095 .
- ^ Перейти обратно: а б Чен Дж., Сильвер Д.П., Уолпита Д., Кантор С.Б., Газдар А.Ф., Томлинсон Дж. и др. (сентябрь 1998 г.). «Стабильное взаимодействие продуктов генов-супрессоров опухолей BRCA1 и BRCA2 в митотических и мейотических клетках» . Мол. Клетка . 2 (3): 317–28. дои : 10.1016/S1097-2765(00)80276-2 . ПМИД 9774970 .
- ^ Рейтер Т., Медхерст А.Л., Вайсфиш К., Чжи Ю., Хертерих С., Хоэн Х. и др. (октябрь 2003 г.). «Двугибридные дрожжевые скрининги подразумевают участие белков анемии Фанкони в регуляции транскрипции, передаче сигналов в клетках, окислительном метаболизме и клеточном транспорте». Эксп. Сотовый Res . 289 (2): 211–21. дои : 10.1016/S0014-4827(03)00261-1 . ПМИД 14499622 .
- ^ Футамура М., Аракава Х., Мацуда К., Катагири Т., Саджи С., Мики Ю. и др. (март 2000 г.). «Потенциальная роль BRCA2 в митотической контрольной точке после фосфорилирования hBUBR1». Рак Рез . 60 (6): 1531–5. ПМИД 10749118 .
- ^ Сиддик Х., Рао В.Н., Редди Э.С. (август 2009 г.). «CBP-опосредованное посттрансляционное N-гликозилирование BRCA2» . Инт Джей Онкол . 35 (2): 16387–91. дои : 10.3892/ijo_00000351 . ПМИД 19578754 .
- ^ Хьюз-Дэвис Л., Хантсман Д., Руас М., Фукс Ф., Бай Дж., Чин С.Ф. и др. (ноябрь 2003 г.). «EMSY связывает путь BRCA2 со спорадическим раком молочной железы и яичников» . Клетка . 115 (5): 523–35. дои : 10.1016/S0092-8674(03)00930-9 . ПМИД 14651845 . S2CID 18911371 .
- ^ Ван X, Андреассен П.Р., Д'Андреа А.Д. (июль 2004 г.). «Функциональное взаимодействие моноубиквитинированных FANCD2 и BRCA2/FANCD1 в хроматине» . Мол. Клетка. Биол . 24 (13): 5850–62. дои : 10.1128/MCB.24.13.5850-5862.2004 . ПМК 480901 . ПМИД 15199141 .
- ^ Хусейн С., Уилсон Дж.Б., Медхерст А.Л., Хейна Дж., Витт Э., Анант С. и др. (июнь 2004 г.). «Прямое взаимодействие FANCD2 с BRCA2 в путях реакции на повреждение ДНК» . Хм. Мол. Жене . 13 (12): 1241–8. дои : 10.1093/hmg/ddh135 . ПМИД 15115758 .
- ^ Хейна Дж., Холторф М., Хайнс Дж., Мэтьюсон Л., Хемфилл А., Аль-Дхалими М. и др. (апрель 2008 г.). «Tip60 необходим для восстановления межцепочечных поперечных связей ДНК в пути анемии Фанкони» . Ж. Биол. Хим . 283 (15): 9844–51. дои : 10.1074/jbc.M709076200 . ПМК 2398728 . ПМИД 18263878 .
- ^ Хусейн С., Витт Э., Хубер П.А., Медхерст А.Л., Эшворт А., Мэтью К.Г. (октябрь 2003 г.). «Прямое взаимодействие белка анемии Фанкони FANCG с BRCA2/FANCD1» . Хм. Мол. Жене . 12 (19): 2503–10. дои : 10.1093/hmg/ddg266 . ПМИД 12915460 .
- ^ Юань Ю, Шэнь Цзи (декабрь 2001 г.). «Взаимодействие с BRCA2 предполагает роль филамина-1 (hsFLNa) в реакции на повреждение ДНК» . Ж. Биол. Хим . 276 (51): 48318–24. дои : 10.1074/jbc.M102557200 . ПМИД 11602572 .
- ^ Марморштейн Л.Ю., Кинев А.В., Чан Г.К., Бочар Д.А., Бения Х., Эпштейн Дж.А. и др. (январь 2001 г.). «Комплекс BRCA2 человека, содержащий структурный компонент, связывающийся с ДНК, влияет на развитие клеточного цикла» . Клетка . 104 (2): 247–57. дои : 10.1016/S0092-8674(01)00209-4 . ПМИД 11207365 . S2CID 5822368 .
- ^ Хакими М.А., Бочар Д.А., Ченовет Дж., Лейн В.С., Мандель Г., Шихаттар Р. (май 2002 г.). «Комплекс ядро-BRAF35, содержащий деацетилазу гистонов, опосредует репрессию нейронально-специфичных генов» . Учеб. Натл. акад. наук. США 99 (11): 7420–5. Бибкод : 2002PNAS...99.7420H . дои : 10.1073/pnas.112008599 . ПМК 124246 . ПМИД 12032298 .
- ^ Перейти обратно: а б с Марморштейн Л.Ю., Оучи Т., Ааронсон С.А. (ноябрь 1998 г.). «Продукт гена BRCA2 функционально взаимодействует с p53 и RAD51» . Учеб. Натл. акад. наук. США 95 (23): 13869–74. Бибкод : 1998PNAS...9513869M . дои : 10.1073/pnas.95.23.13869 . ПМК 24938 . ПМИД 9811893 .
- ^ «Энтрез Джин: партнер PALB2 и локализатор BRCA2» .
- ^ Перейти обратно: а б с Лин Х.Р., Тинг Н.С., Цинь Дж., Ли WH (сентябрь 2003 г.). «М-фазное специфическое фосфорилирование BRCA2 с помощью Polo-подобной киназы 1 коррелирует с диссоциацией комплекса BRCA2-P/CAF» . Ж. Биол. Хим . 278 (38): 35979–87. дои : 10.1074/jbc.M210659200 . ПМИД 12815053 .
- ^ Фукс Ф., Милнер Дж., Кузаридес Т. (ноябрь 1998 г.). «BRCA2 связывается с активностью ацетилтрансферазы при связывании с P/CAF» . Онкоген . 17 (19): 2531–4. дои : 10.1038/sj.onc.1202475 . ПМИД 9824164 .
- ^ Ли М., Дэниелс М.Дж., Венкитараман А.Р. (январь 2004 г.). «Фосфорилирование BRCA2 с помощью Polo-подобной киназы Plk1 регулируется повреждением ДНК и митотической прогрессией» . Онкоген . 23 (4): 865–72. дои : 10.1038/sj.onc.1207223 . ПМИД 14647413 .
- ^ Шаран С.К., Моримацу М., Альбрехт У., Лим Д.С., Регель Э., Динь С. и др. (апрель 1997 г.). «Эмбриональная летальность и радиационная гиперчувствительность, опосредованная Rad51, у мышей, лишенных Brca2». Природа . 386 (6627): 804–10. Бибкод : 1997Natur.386..804S . дои : 10.1038/386804a0 . hdl : 11858/00-001M-0000-0010-5059-F . ПМИД 9126738 . S2CID 4238943 .
- ^ Ю Д.С., Сонода Э., Такеда С., Хуанг К.Л., Пеллегрини Л., Бланделл Т.Л. и др. (октябрь 2003 г.). «Динамический контроль рекомбиназы Rad51 путем самоассоциации и взаимодействия с BRCA2» . Мол. Клетка . 12 (4): 1029–41. дои : 10.1016/S1097-2765(03)00394-0 . ПМИД 14580352 .
- ^ Перейти обратно: а б Чен П.Л., Чен К.Ф., Чен Ю, Сяо Дж., Шарп З.Д., Ли В.Х. (апрель 1998 г.). «Повторы BRC в BRCA2 имеют решающее значение для связывания RAD51 и устойчивости к обработке метилметансульфонатом» . Учеб. Натл. акад. наук. США 95 (9): 5287–92. Бибкод : 1998PNAS...95.5287C . дои : 10.1073/pnas.95.9.5287 . ЧВК 20253 . ПМИД 9560268 .
- ^ Перейти обратно: а б Вонг А.К., Перо Р., Ормонд П.А., Тавтигян С.В., Бартель П.Л. (декабрь 1997 г.). «RAD51 взаимодействует с эволюционно консервативными мотивами BRC в гене предрасположенности к раку молочной железы человека brca2» . Ж. Биол. Хим . 272 (51): 31941–4. дои : 10.1074/jbc.272.51.31941 . ПМИД 9405383 .
- ^ Катагири Т., Сайто Х., Шинохара А., Огава Х., Камада Н., Накамура Ю. и др. (март 1998 г.). «Множество возможных сайтов BRCA2, взаимодействующих с белком репарации ДНК RAD51». Гены Хромосомы Рак . 21 (3): 217–22. doi : 10.1002/(SICI)1098-2264(199803)21:3<217::AID-GCC5>3.0.CO;2-2 . ПМИД 9523196 . S2CID 45954246 .
- ^ Пеллегрини Л., Ю Д.С., Ло Т., Ананд С., Ли М., Бланделл Т.Л. и др. (ноябрь 2002 г.). «Понимание рекомбинации ДНК на основе структуры комплекса RAD51-BRCA2». Природа . 420 (6913): 287–93. Бибкод : 2002Natur.420..287P . дои : 10.1038/nature01230 . ПМИД 12442171 . S2CID 4359383 .
- ^ Тарсунас М., Дэвис А.А., West SC (январь 2004 г.). «Локализация и активация RAD51 после повреждения ДНК» . Филос. Пер. Р. Сок. Лонд. Б Биол. Наука . 359 (1441): 87–93. дои : 10.1098/rstb.2003.1368 . ПМК 1693300 . ПМИД 15065660 .
- ^ Вонг Дж. М., Ионеску Д., Инглес CJ (январь 2003 г.). «Взаимодействие между BRCA2 и репликационным белком А нарушается предрасполагающей к раку мутацией в BRCA2» . Онкоген . 22 (1): 28–33. дои : 10.1038/sj.onc.1206071 . ПМИД 12527904 .
- ^ Марстон, Нью-Джерси, Ричардс В.Дж., Хьюз Д., Бертвистл Д., Маршалл С.Дж., Эшворт А. (июль 1999 г.). «Взаимодействие между продуктом гена предрасположенности к раку молочной железы BRCA2 и DSS1, белком, функционально консервативным от дрожжей до млекопитающих» . Мол. Клетка. Биол . 19 (7): 4633–42. дои : 10.1128/MCB.19.7.4633 . ПМК 84261 . ПМИД 10373512 .
- ^ Перейти обратно: а б с д и Ян Х., Джеффри П.Д., Миллер Дж., Киннукан Э., Сан Ю., Тома Н.Х. и др. (сентябрь 2002 г.). «Функция BRCA2 в связывании ДНК и рекомбинации из структуры BRCA2-DSS1-ssDNA». Наука . 297 (5588): 1837–48. Бибкод : 2002Sci...297.1837Y . дои : 10.1126/science.297.5588.1837 . ПМИД 12228710 .
- ^ Преображенская О, Якимовых М, Канамото Т, Якимовых I, Стоика R, Heldin CH, et al. (August 2002). "BRCA2 and Smad3 synergize in regulation of gene transcription" . Oncogene . 21 (36): 5660-4. doi : 10.1038/sj.onc.1205732 . PMID 12165866 .
- ^ Борк П., Бломберг Н., Нильгес М. (май 1996 г.). «Внутренние повторы в последовательности белка BRCA2». Нат. Жене . 13 (1): 22–3. дои : 10.1038/ng0596-22 . ПМИД 8673099 . S2CID 2312211 .
- ^ Алагар С., Бахадур Р.П. (2020). «DSS1 аллостерически регулирует конформацию башенного домена BRCA2, который обладает специфичностью связывания дцДНК для гомологичной рекомбинации». Международный журнал биологических макромолекул . 165 (Часть А): 918–929. doi : 10.1016/j.ijbiomac.2020.09.230 . ПМИД 33011260 . S2CID 222165754 .
- ^ «ACLU подает в суд на патенты на гены рака молочной железы» . Си-Эн-Эн. Архивировано из оригинала 15 мая 2009 года . Проверено 14 мая 2009 г.
- ^ Конли Дж., Ворхаус Д., Кук-Диган Дж. (01.03.2011). «Как Myriad отреагирует на следующее поколение тестов BRCA?» . Робинсон, Брэдшоу и Хинсон . Проверено 9 декабря 2012 г.
- ^ «Генетика и патентование» . Информация о проекте «Геном человека» . Программы генома Министерства энергетики США. 07.07.2010.
- ^ Липтак А (13 июня 2013 г.). «Верховный суд постановил, что человеческие гены не могут быть запатентованы» . Нью-Йорк Таймс . Проверено 13 июня 2013 г.
- ^ Кордерой А. (15 февраля 2013 г.). «Веховое патентное решение в отношении гена рака молочной железы BRCA1» . Сидней Морнинг Геральд . Проверено 14 июня 2013 г.
- ^ Кордерой А (14 июня 2013 г.). «Компании не могут патентовать гены, постановил суд США» . Сидней Морнинг Геральд . Проверено 14 июня 2013 г.
Дальнейшее чтение
[ редактировать ]- Цзоу Дж.П., Хиросе Ю., Сиддик Х., Рао В.Н., Редди Э.С. (1999). «Структура и экспрессия варианта BRCA2a, лишенного домена трансактивации». Отчеты онкологии . 6 (2): 437–40. дои : 10.3892/или.6.2.437 . ПМИД 10023017 .
- Венкитараман А.Р. (2001). «Хромосомная стабильность, рекомбинация ДНК и супрессор опухоли BRCA2». Современное мнение в области клеточной биологии . 13 (3): 338–43. дои : 10.1016/S0955-0674(00)00217-9 . ПМИД 11343905 .
- Орелли Б.Дж., епископ Д.К. (2001). «BRCA2 и гомологичная рекомбинация» . Исследование рака молочной железы . 3 (5): 294–8. дои : 10.1186/bcr310 . ПМК 138691 . ПМИД 11597317 .
- Дэниел, округ Колумбия (2002). «Основной момент: белки BRCA1 и BRCA2 при раке молочной железы» . Микроскопические исследования и техника . 59 (1): 68–83. дои : 10.1002/jemt.10178 . ПМИД 12242698 . S2CID 30091586 .
- Тутт А., Эшворт А. (2003). «Взаимосвязь между ролью генов BRCA в репарации ДНК и предрасположенностью к раку». Тенденции молекулярной медицины . 8 (12): 571–6. дои : 10.1016/S1471-4914(02)02434-6 . ПМИД 12470990 .
- Гонсалвеш А., Вьенс П., Соболь Х., Маранинчи Д., Бертуччи Ф. (2005). «[Молекулярные изменения при раке молочной железы: клинические последствия и новые аналитические инструменты]». Revue de Médecine Interne . 26 (6): 470–8. doi : 10.1016/j.revmed.2004.11.012 . ПМИД 15936476 .
- Хэй Т., Кларк А.Р. (2005). «Гиперчувствительность к повреждению ДНК в клетках, лишенных BRCA2: обзор данных in vitro и in vivo». Труды Биохимического общества . 33 (Часть 4): 715–7. дои : 10.1042/BST0330715 . ПМИД 16042582 .
- Домчек С.М., Вебер Б.Л. (2006). «Клиническое ведение носителей мутаций BRCA1 и BRCA2» . Онкоген . 25 (43): 5825–31. дои : 10.1038/sj.onc.1209881 . ПМИД 16998496 .
- Хонрадо Э., Осорио А., Паласиос Дж., Бенитес Дж. (2006). «Патология и экспрессия генов наследственных опухолей молочной железы, связанных с мутациями генов BRCA1, BRCA2 и CHEK2» . Онкоген . 25 (43): 5837–45. дои : 10.1038/sj.onc.1209875 . ПМИД 16998498 . S2CID 20960561 .
Внешние ссылки
[ редактировать ]- Белок BRCA2 Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)
галерея PDB |
|---|

|}